
Targeting CA-125 Transcription by Development of a Conditionally Replicative Adenovirus for Ovarian Cancer Treatment


 and
and 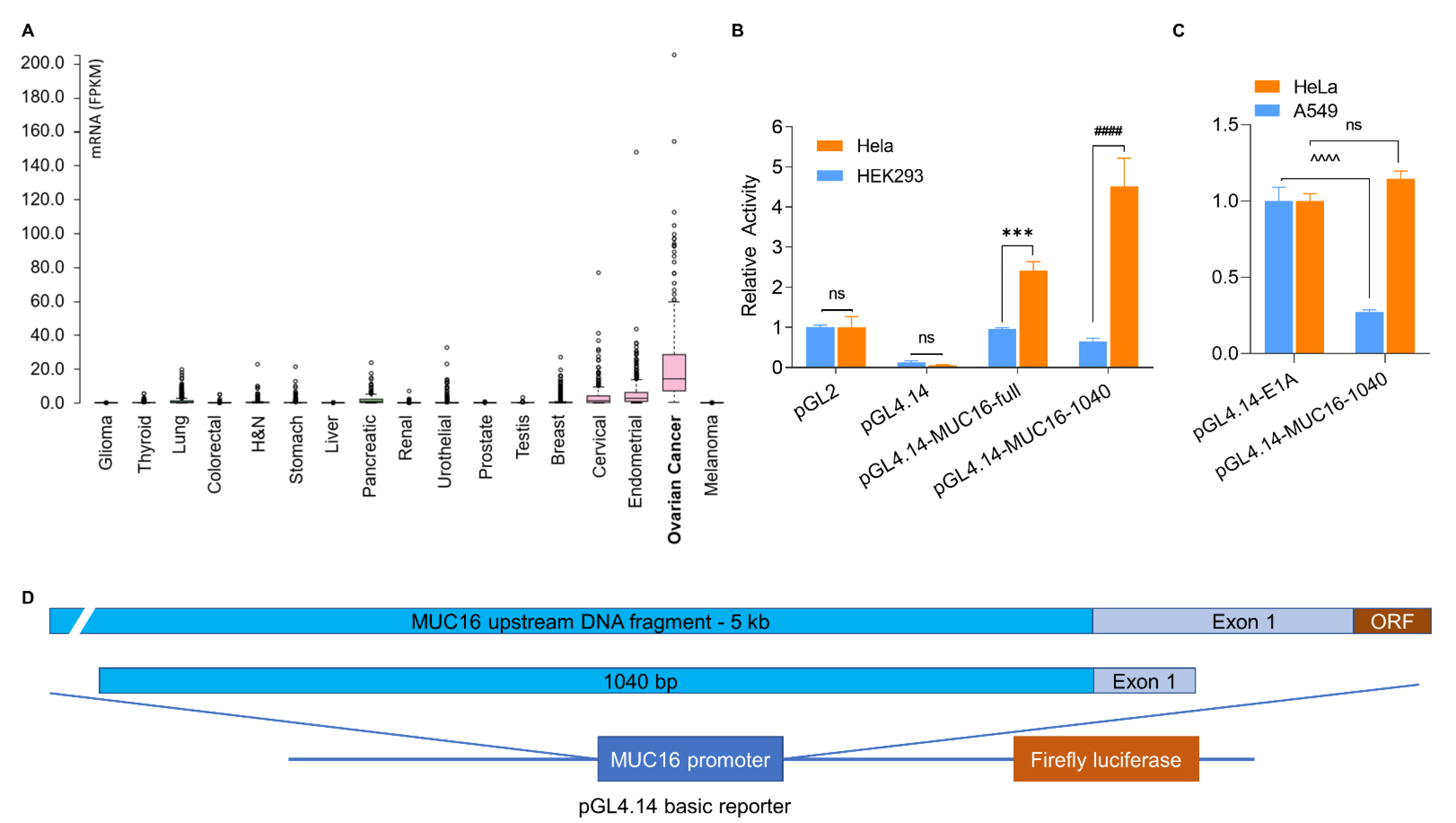{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture
2.2. Cell Viability Assay
2.3. Primary Ovarian Cancer Cell Collection and Culture
2.4. Adenovirus Construction, Amplification and Purification
2.5. Reporter Vector Construction and Dual-Luciferase Assay
2.6. mRNA Isolation and Quantitative Real Time PCR (qRT-PCR)
2.7. Virus Oncolytic Activity in NSG Tumor Xenograft Models
2.8. Bioluminescent Imaging of Tumor-Bearing Mice
2.9. Statistical Analysis
3. Results
3.1. Identification of an Upstream Region of the MUC16 Gene with Specific Transcriptional Activity
3.2. Generation of a CRAd with MUC16 Fragment with Promoter Activity to Control E1A Expression
3.3. Replication and Oncolysis of Ad5/MUC16-1040 in Ovarian Cancer Cell Lines Is Dependent on CA-125 Expression
3.4. Selective Replication and Oncolysis in Primary Ovarian Cancer Cells
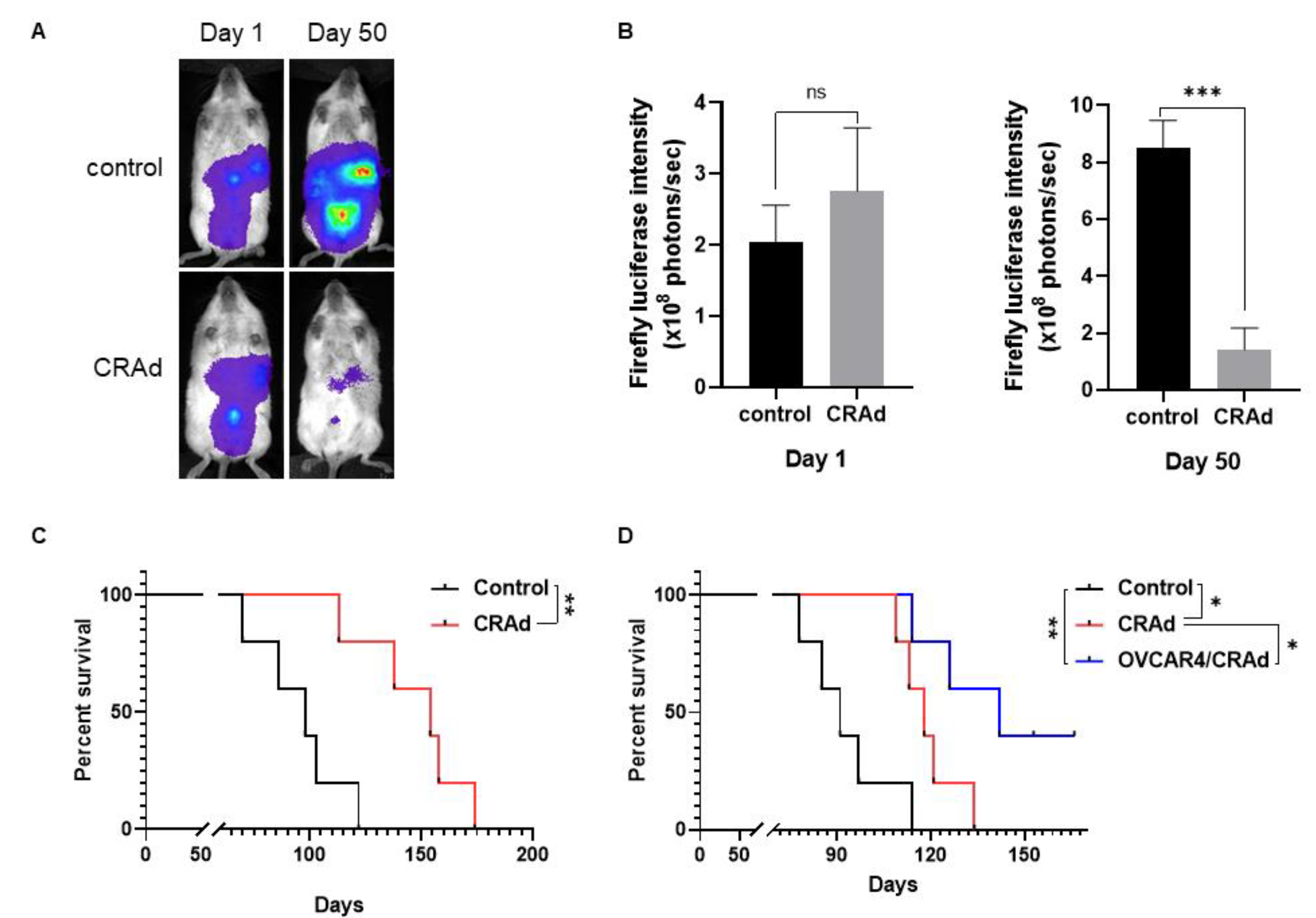
3.5. Oncolytic Activity in Ovarian Cancer Xenograft Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- PDQ Adult Treatment Editorial Board. Ovarian Epithelial, Fallopian Tube, and Primary Peritoneal Cancer Treatment (PDQ(R)): Health Professional Version. In PDQ Cancer Information Summaries; National Cancer Institute: Bethesda, MD, USA, 2002. [Google Scholar]
- Nebgen, D.R.; Lu, K.H.; Bast, R.C., Jr. Novel Approaches to Ovarian Cancer Screening. Curr. Oncol. Rep. 2019, 21, 75. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Dochez, V.; Caillon, H.; Vaucel, E.; Dimet, J.; Winer, N.; Ducarme, G. Biomarkers and algorithms for diagnosis of ovarian cancer: CA125, HE4, RMI and ROMA, a review. J. Ovarian. Res. 2019, 12, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, T.; Bhatt, H. Cancer Antigen 125. Available online: https://www.ncbi.nlm.nih.gov/books/NBK562245/ (accessed on 13 August 2021).
- Bast, R.C., Jr.; Feeney, M.; Lazarus, H.; Nadler, L.M.; Colvin, R.B.; Knapp, R.C. Reactivity of a monoclonal antibody with human ovarian carcinoma. J. Clin. Investig. 1981, 68, 1331–1337. [Google Scholar] [CrossRef] [Green Version]
- Aithal, A.; Rauth, S.; Kshirsagar, P.; Shah, A.; Lakshmanan, I.; Junker, W.M.; Jain, M.; Ponnusamy, M.P.; Batra, S.K. MUC16 as a novel target for cancer therapy. Expert Opin. Ther. Targets 2018, 22, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Moore, K.N.; Birrer, M.J.; Berlin, S.; Matulonis, U.A.; Infante, J.R.; Wolpin, B.; Poon, K.A.; Firestein, R.; Xu, J.; et al. Phase I study of safety and pharmacokinetics of the anti-MUC16 antibody-drug conjugate DMUC5754A in patients with platinum-resistant ovarian cancer or unresectable pancreatic cancer. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2016, 27, 2124–2130. [Google Scholar] [CrossRef]
- Paijens, S.T.; Leffers, N.; Daemen, T.; Helfrich, W.; Boezen, H.M.; Cohlen, B.J.; Melief, C.J.; de Bruyn, M.; Nijman, H.W. Antigen-specific active immunotherapy for ovarian cancer. Cochrane Database Syst. Rev. 2018, 9, Cd007287. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini, P.; Harter, P.; Scambia, G.; Sehouli, J.; Meier, W.; Wimberger, P.; Baumann, K.H.; Kurzeder, C.; Schmalfeldt, B.; Cibula, D.; et al. Abagovomab as maintenance therapy in patients with epithelial ovarian cancer: A phase III trial of the AGO OVAR, COGI, GINECO, and GEICO--the MIMOSA study. J. Clin. Oncol. 2013, 31, 1554–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berek, J.S.; Taylor, P.T.; Gordon, A.; Cunningham, M.J.; Finkler, N.; Orr, J., Jr.; Rivkin, S.; Schultes, B.C.; Whiteside, T.L.; Nicodemus, C.F. Randomized, placebo-controlled study of oregovomab for consolidation of clinical remission in patients with advanced ovarian cancer. J. Clin. Oncol. 2004, 22, 3507–3516. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, A.; Fossati, M.; Buzzonetti, A.; Scambia, G.; Fattorossi, A. A robust immune system conditions the response to abagovomab (anti-idiotypic monoclonal antibody mimicking the CA125 protein) vaccination in ovarian cancer patients. Immunol. Lett. 2017, 191, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Chekmasova, A.A.; Rao, T.D.; Nikhamin, Y.; Park, K.J.; Levine, D.A.; Spriggs, D.R.; Brentjens, R.J. Successful eradication of established peritoneal ovarian tumors in SCID-Beige mice following adoptive transfer of T cells genetically targeted to the MUC16 antigen. Clin. CancerRes. Off. J. Am. Assoc. Cancer Res. 2010, 16, 3594–3606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koneru, M.; O’Cearbhaill, R.; Pendharkar, S.; Spriggs, D.R.; Brentjens, R.J. A phase I clinical trial of adoptive T cell therapy using IL-12 secreting MUC-16(ecto) directed chimeric antigen receptors for recurrent ovarian cancer. J. Transl. Med. 2015, 13, 102. [Google Scholar] [CrossRef] [Green Version]
- Yin, B.W.; Lloyd, K.O. Molecular cloning of the CA125 ovarian cancer antigen: Identification as a new mucin, MUC16. J. Biol. Chem. 2001, 276, 27371–27375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapi, D.; Binder, D.; Rosales, N.; Yan, X.J.; Lash, A.E.; Spriggs, D.R. Abstract 2033: Transcription factors that regulate MUC16 expression in ovarian cancer. Cancer Res. 2011, 71, 2033. [Google Scholar] [CrossRef]
- Zhang, M.X.; Hong, S.S.; Cai, Q.Q.; Zhang, M.; Chen, J.; Zhang, X.Y.; Xu, C.J. Transcriptional control of the MUC16 promoter facilitates follicle-stimulating hormone peptide-conjugated shRNA nanoparticle-mediated inhibition of ovarian carcinoma in vivo. Drug Deliv. 2018, 25, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Flint, J.; Shenk, T. Adenovirus E1A protein paradigm viral transactivator. Annu. Rev. Genet. 1989, 23, 141–161. [Google Scholar] [CrossRef]
- Moran, E.; Mathews, M.B. Multiple functional domains in the adenovirus E1A gene. Cell 1987, 48, 177–178. [Google Scholar] [CrossRef]
- Frisch, S.M.; Mymryk, J.S. Adenovirus-5 E1A: Paradox and paradigm. Nat. Rev. Mol. Cell Biol. 2002, 3, 441–452. [Google Scholar] [CrossRef]
- The Human Protein Atlas: RNA Expression Overview. Available online: https://www.proteinatlas.org/ENSG00000181143-MUC16/cell (accessed on 13 August 2021).
- Loimas, S.; Wahlfors, J.; Jänne, J. Herpes simplex virus thymidine kinase-green fluorescent protein fusion gene: New tool for gene transfer studies and gene therapy. Biotechniques 1998, 24, 614–618. [Google Scholar] [CrossRef]
- Steffens, S.; Frank, S.; Fischer, U.; Heuser, C.; Meyer, K.L.; Dobberstein, K.U.; Rainov, N.G.; Kramm, C.M. Enhanced green fluorescent protein fusion proteins of herpes simplex virus type 1 thymidine kinase and cytochrome P450 4B1: Applications for prodrug-activating gene therapy. Cancer Gene Ther. 2000, 7, 806–812. [Google Scholar] [CrossRef] [Green Version]
- Louis, N.; Evelegh, C.; Graham, F.L. Cloning and sequencing of the cellular-viral junctions from the human adenovirus type 5 transformed 293 cell line. Virology 1997, 233, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Ganly, I.; Kim, Y.T.; Hann, B.; Balmain, A.; Brown, R. Replication and cytolysis of an E1B-attenuated adenovirus in drug-resistant ovarian tumour cells is associated with reduced apoptosis. Gene Ther. 2001, 8, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Flak, M.B.; Connell, C.M.; Chelala, C.; Archibald, K.; Salako, M.A.; Pirlo, K.J.; Lockley, M.; Wheatley, S.P.; Balkwill, F.R.; McNeish, I.A. p21 Promotes oncolytic adenoviral activity in ovarian cancer and is a potential biomarker. Mol. Cancer 2010, 9, 175. [Google Scholar] [CrossRef] [Green Version]
- Tookman, L.A.; Browne, A.K.; Connell, C.M.; Bridge, G.; Ingemarsdotter, C.K.; Dowson, S.; Shibata, A.; Lockley, M.; Martin, S.A.; McNeish, I.A. RAD51 and BRCA2 Enhance Oncolytic Adenovirus Type 5 Activity in Ovarian Cancer. Mol. Cancer Res. 2016, 14, 44–55. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Lee, S.H.; Cho, Y.S.; Choi, J.J.; Kim, Y.H.; Lee, J.H. Enhancement of the adenoviral sensitivity of human ovarian cancer cells by transient expression of coxsackievirus and adenovirus receptor (CAR). Gynecol. Oncol. 2002, 85, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K. Striking out at disseminated metastases: The systemic delivery of oncolytic viruses. Curr. Opin. Mol. Ther. 2006, 8, 301–313. [Google Scholar]
- Neukirch, L.; Fougeroux, C.; Andersson, A.C.; Holst, P.J. The potential of adenoviral vaccine vectors with altered antigen presentation capabilities. Expert Rev. Vaccines 2020, 19, 25–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanpourghadi, M.; Novikov, M.; Ertl, H.C.J. COVID-19 Vaccines Based on Adenovirus Vectors. Trends Biochem. Sci. 2021, 46, 429–430. [Google Scholar] [CrossRef] [PubMed]
- González-Pastor, R.; Ashshi, A.M.; El-Shemi, A.G.; Dmitriev, I.P.; Kashentseva, E.A.; Lu, Z.H.; Goedegebuure, S.P.; Podhajcer, O.L.; Curiel, D.T. Defining a murine ovarian cancer model for the evaluation of conditionally-replicative adenovirus (CRAd) virotherapy agents. J. Ovarian. Res. 2019, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haitz, K.; Khosravi, H.; Lin, J.Y.; Menge, T.; Nambudiri, V.E. Review of talimogene laherparepvec: A first-in-class oncolytic viral treatment of advanced melanoma. J. Am. Acad. Dermatol. 2020, 83, 189–196. [Google Scholar] [CrossRef]
- Vasey, P.A.; Shulman, L.N.; Campos, S.; Davis, J.; Gore, M.; Johnston, S.; Kirn, D.H.; O’Neill, V.; Siddiqui, N.; Seiden, M.V.; et al. Phase I trial of intraperitoneal injection of the E1B-55-kd-gene-deleted adenovirus ONYX-015 (dl1520) given on days 1 through 5 every 3 weeks in patients with recurrent/refractory epithelial ovarian cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 1562–1569. [Google Scholar] [CrossRef]
- Bischoff, J.R.; Kirn, D.H.; Williams, A.; Heise, C.; Horn, S.; Muna, M.; Ng, L.; Nye, J.A.; Sampson-Johannes, A.; Fattaey, A.; et al. An adenovirus mutant that replicates selectively in p53-deficient human tumor cells. Science 1996, 274, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Sauthoff, H.; Hu, J.; Maca, C.; Goldman, M.; Heitner, S.; Yee, H.; Pipiya, T.; Rom, W.N.; Hay, J.G. Intratumoral spread of wild-type adenovirus is limited after local injection of human xenograft tumors: Virus persists and spreads systemically at late time points. Hum. Gene Ther. 2003, 14, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.; Sauthoff, H.; Heitner, S.; Jagirdar, J.; Rom, W.N.; Hay, J.G. Wild-type adenovirus decreases tumor xenograft growth, but despite viral persistence complete tumor responses are rarely achieved--deletion of the viral E1b-19-kD gene increases the viral oncolytic effect. Hum. Gene Ther. 2001, 12, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.M.; Havunen, R.; Hemminki, A. Modulation of the tumor microenvironment with an oncolytic adenovirus for effective T-cell therapy and checkpoint inhibition. Methods Enzymol. 2020, 635, 205–230. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.M.; Heiniö, C.; Cervera-Carrascon, V.; Quixabeira, D.C.A.; Siurala, M.; Havunen, R.; Butzow, R.; Zafar, S.; de Gruijl, T.; Lassus, H.; et al. Oncolytic adenovirus shapes the ovarian tumor microenvironment for potent tumor-infiltrating lymphocyte tumor reactivity. J. Immunother. Cancer 2020, 8, e000188. [Google Scholar] [CrossRef] [Green Version]
- Loskog, A. Immunostimulatory Gene Therapy Using Oncolytic Viruses as Vehicles. Viruses 2015, 7, 5780–5791. [Google Scholar] [CrossRef]
- Hawkins, L.K.; Johnson, L.; Bauzon, M.; Nye, J.A.; Castro, D.; Kitzes, G.A.; Young, M.D.; Holt, J.K.; Trown, P.; Hermiston, T.W. Gene delivery from the E3 region of replicating human adenovirus: Evaluation of the 6.7 K/gp19 K region. Gene Ther. 2001, 8, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, M.S. Function of adenovirus E3 proteins and their interactions with immunoregulatory cell proteins. J. Gene Med. 2004, 6 (Suppl. S1), S172–S183. [Google Scholar] [CrossRef]
- Kanerva, A.; Wang, M.; Bauerschmitz, G.J.; Lam, J.T.; Desmond, R.A.; Bhoola, S.M.; Barnes, M.N.; Alvarez, R.D.; Siegal, G.P.; Curiel, D.T.; et al. Gene transfer to ovarian cancer versus normal tissues with fiber-modified adenoviruses. Mol. Ther. 2002, 5, 695–704. [Google Scholar] [CrossRef]
- Atasheva, S.; Yao, J.; Shayakhmetov, D.M. Innate immunity to adenovirus: Lessons from mice. FEBS Lett. 2019, 593, 3461–3483. [Google Scholar] [CrossRef] [PubMed]
- Shayakhmetov, D.M.; Gaggar, A.; Ni, S.; Li, Z.Y.; Lieber, A. Adenovirus binding to blood factors results in liver cell infection and hepatotoxicity. J. Virol. 2005, 79, 7478–7491. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, E.; Yang, G.; Yao, Y.; Wang, G.; Mohanty, A.; Fan, F.; Zhao, L.; Zhang, Y.; Mirzapoiazova, T.; Walser, T.C.; et al. Targeting CA-125 Transcription by Development of a Conditionally Replicative Adenovirus for Ovarian Cancer Treatment. Cancers 2021, 13, 4265. https://doi.org/10.3390/cancers13174265
Yue E, Yang G, Yao Y, Wang G, Mohanty A, Fan F, Zhao L, Zhang Y, Mirzapoiazova T, Walser TC, et al. Targeting CA-125 Transcription by Development of a Conditionally Replicative Adenovirus for Ovarian Cancer Treatment. Cancers. 2021; 13(17):4265. https://doi.org/10.3390/cancers13174265
Chicago/Turabian StyleYue, Er, Guangchao Yang, Yuanfei Yao, Guangyu Wang, Atish Mohanty, Fang Fan, Ling Zhao, Yanqiao Zhang, Tamara Mirzapoiazova, Tonya C. Walser, and et al. 2021. "Targeting CA-125 Transcription by Development of a Conditionally Replicative Adenovirus for Ovarian Cancer Treatment" Cancers 13, no. 17: 4265. https://doi.org/10.3390/cancers13174265