
Ovarian Cancer-Associated Mesothelial Cells: Transdifferentiation to Minions of Cancer and Orchestrate Developing Peritoneal Dissemination

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Characteristics and Role of MCs and the Association with OvCa
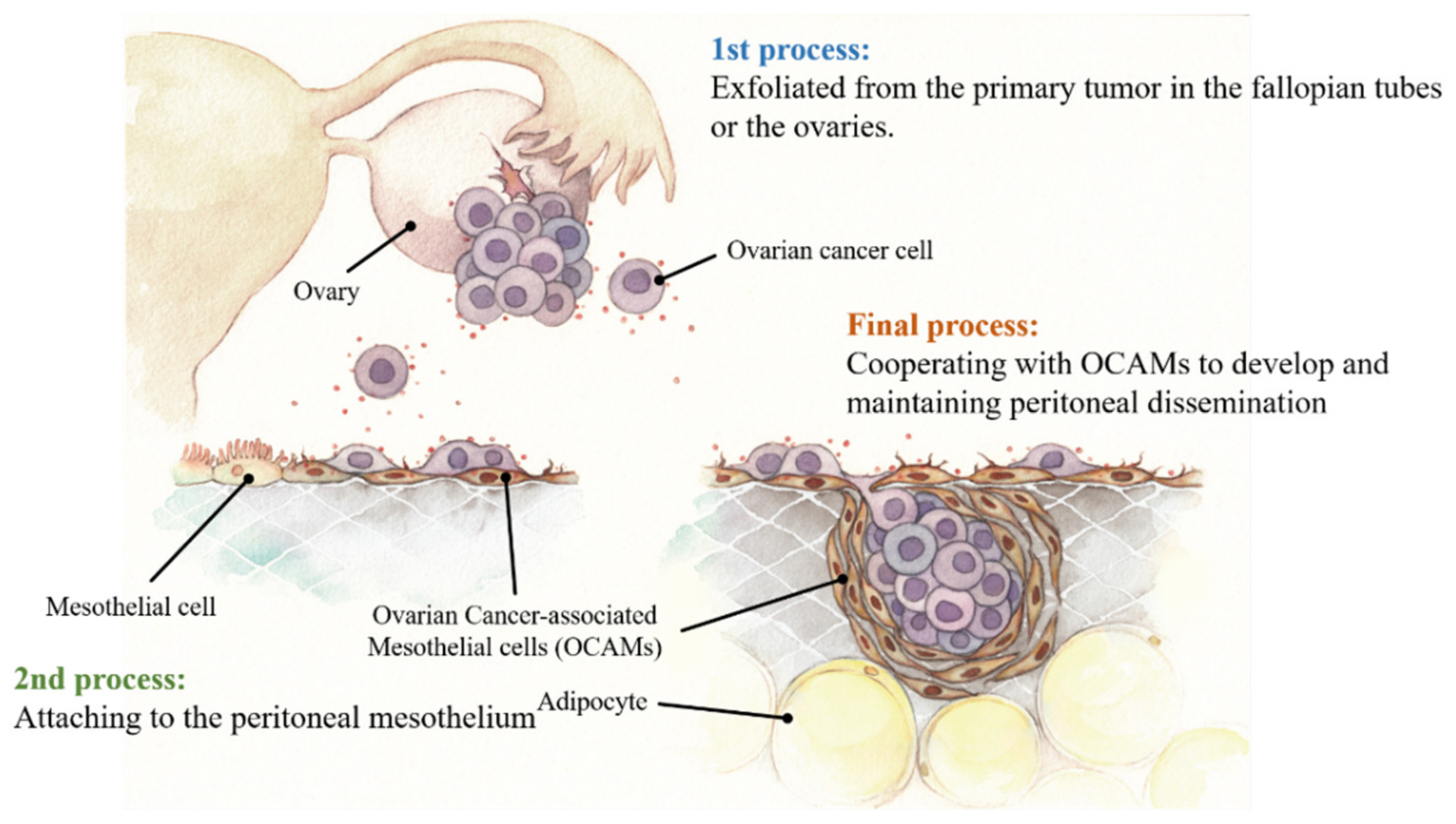
3. Biological Process of Peritoneal Dissemination of OvCa
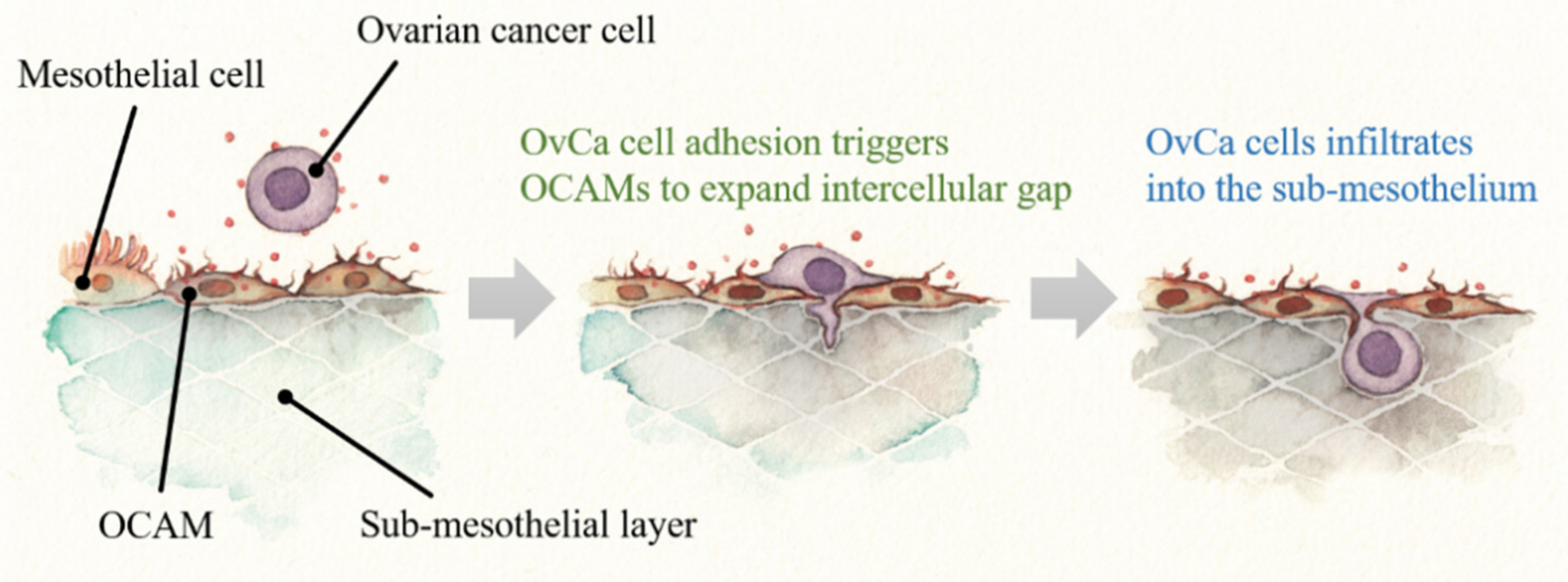
4. OCAMs Affect OvCa Adhesion on the Peritoneum
4.1. Adhesion Molecules
4.2. OCAMs and OvCa Adhesion
5. OCAMs Represent One of the Stromal Components Promoting Tumor Microenvironment of Peritoneal Dissemination
6. Future Prospects
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OvCa | Ovarian cancer |
| MCs | Mesothelial cells |
| OCAMs | Ovarian cancer-associated mesothelial cells |
References
- Torre, L.A.; Trabert, B.; De Santis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian Cancer Statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Machida, H.; Matsuo, K.; Yamagami, W.; Ebina, Y.; Kobayashi, Y.; Tabata, T.; Kanauchi, M.; Nagase, S.; Enomoto, T.; Mikami, M. Trends and characteristics of epithelial ovarian cancer in Japan between 2002 and 2015: A JSGO-JSOG joint study. Gynecol. Oncol. 2019, 153, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Prat, J.; FIGO Committee on Gynecologic Oncology. Abridged Republication of FIGO’s Staging Classification for Cancer of the Ovary, Fallopian Tube, and Peritoneum. Cancer 2015, 121, 3452–3454. [Google Scholar] [CrossRef] [PubMed]
- Thibault, B.; Castells, M.; Delord, J.P.; Couderc, B. Ovarian Cancer Microenvironment: Implications for Cancer Dissemination and Chemoresistance Acquisition. Cancer Metastasis Rev. 2014, 33, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Perren, T.J.; Swart, A.M.; Pfisterer, J.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; Kurzeder, C.; et al. A phase 3 Trial of Bevacizumab in Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2484–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsumata, N.; Yasuda, M.; Takahashi, F.; Isonishi, S.; Jobo, T.; Aoki, D.; Tsuda, H.; Sugiyama, T.; Kodama, S.; Kimura, E.; et al. Dose-Dense Paclitaxel Once a Week in Combination with Carboplatin Every 3 Weeks for Advanced Ovarian Cancer: A phase 3, Open-Label, Randomised Controlled Trial. Lancet 2009, 374, 1331–1338. [Google Scholar] [CrossRef]
- Paget, S. The Distribution of Secondary Growths in Cancer of the Breast. Lancet 1889, 133, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Langley, R.R.; Fidler, I.J. The Seed and Soil Hypothesis Revisited—The Role of Tumor-Stroma Interactions in Metastasis to Different Organs. Int. J. Cancer 2011, 128, 2527–2535. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, M.; Kajiyama, H.; Yokoi, A.; Sugiyama, M.; Koya, Y.; Yamakita, Y.; Liu, W.; Nakamura, K.; Moriyama, Y.; Yasui, H.; et al. Ovarian Cancer-Associated Mesothelial Cells Induce Acquired Platinum-Resistance in Peritoneal Metastasis via the FN1/Akt Signaling Pathway. Int. J. Cancer 2020, 146, 2268–2280. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.M.H.; Gardner, M.J.; Catterall, J.B.; Turner, G.A. Hyaluronic Acid Secreted by Mesothelial Cells: A Natural Barrier to Ovarian Cancer Cell Adhesion. Clin. Exp. Metastasis 1995, 13, 373–380. [Google Scholar] [CrossRef]
- Yoshida, M.; Taguchi, A.; Kawana, K.; Adachi, K.; Kawata, A.; Ogishima, J.; Nakamura, H.; Fujimoto, A.; Sato, M.; Inoue, T.; et al. Modification of the Tumor Microenvironment in KRAS or c-MYC-Induced Ovarian Cancer-Associated Peritonitis. PLoS ONE 2016, 11, e0160330. [Google Scholar]
- Mutsaers, S.E.; Prêle, C.M.A.; Pengelly, S.; Herrick, S.E. Mesothelial Cells and Peritoneal Homeostasis. Fertil. Steril. 2016, 106, 1018–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, P.; Jiménez-Heffernan, J.A.; Rynne-Vidal, Á.; Pérez-Lozano, M.L.; Gilsanz, Á.; Ruiz-Carpio, V.; Reyes, R.; García-Bordas, J.; Stamatakis, K.; Dotor, J.; et al. Carcinoma-Associated Fibroblasts Derive From Mesothelial Cells via Mesothelial-to-Mesenchymal Transition in Peritoneal Metastasis. J. Pathol. 2013, 231, 517–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rynne-Vidal, A.; Au-Yeung, C.L.; Jiménez-Heffernan, J.A.; Pérez-Lozano, M.L.; Cremades-Jimeno, L.; Bárcena, C.; Cristóbal-García, I.; Fernández-Chacón, C.; Yeung, T.L.; Mok, S.C.; et al. Mesothelial-to-Mesenchymal Transition as a Possible Therapeutic Target in Peritoneal Metastasis of Ovarian Cancer. J. Pathol. 2017, 242, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Kajiyama, H.; Shibata, K.; Terauchi, M.; Ino, K.; Nawa, A.; Kikkawa, F. Involvement of SDF-1α/CXCR4 Axis in the Enhanced Peritoneal Metastasis of Epithelial Ovarian Carcinoma. Int. J. Cancer 2008, 122, 91–99. [Google Scholar] [CrossRef]
- Sugiyama, K.; Kajiyama, H.; Shibata, K.; Yuan, H.; Kikkawa, F.; Senga, T. Expression of the miR200 Family of MicroRNAs in Mesothelial Cells Suppresses the Dissemination of Ovarian Cancer Cells. Mol. Cancer Ther. 2014, 13, 2081–2091. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Kajiyama, H.; Yuan, H.; Nakamura, K.; Yoshihara, M.; Yokoi, A.; Fujikake, K.; Yasui, H.; Yoshikawa, N.; Suzuki, S.; et al. PAI-1 Secreted From Metastatic Ovarian Cancer Cells Triggers the Tumor-Promoting Role of the Mesothelium in a Feedback Loop to Accelerate Peritoneal Dissemination. Cancer Lett. 2019, 442, 181–192. [Google Scholar] [CrossRef]
- Yokoi, A.; Yoshioka, Y.; Yamamoto, Y.; Ishikawa, M.; Ikeda, S.I.; Kato, T.; Kiyono, T.; Takeshita, F.; Kajiyama, H.; Kikkawa, F.; et al. Malignant Extracellular Vesicles Carrying MMP1 mRNA Facilitate Peritoneal Dissemination in Ovarian Cancer. Nat. Commun. 2017, 8, 14470. [Google Scholar] [CrossRef] [Green Version]
- Fujikake, K.; Kajiyama, H.; Yoshihara, M.; Nishino, K.; Yoshikawa, N.; Utsumi, F.; Suzuki, S.; Niimi, K.; Sakata, J.; Mitsui, H.; et al. A Novel Mechanism of Neovascularization in Peritoneal Dissemination via Cancer-Associated Mesothelial Cells Affected by TGF-β Derived from Ovarian Cancer. Oncol. Rep. 2018, 39, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Mutsaers, S.E. Mesothelial Cells: Their Structure, Function and Role in Serosal Repair. Respirology 2002, 7, 171–191. [Google Scholar] [CrossRef]
- Blackburn, S.C.; Stanton, M.P. Anatomy and Physiology of the Peritoneum. Semin. Pediatr. Surg. 2014, 23, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Albanese, A.M.; Albanese, E.F.; Miño, J.H.; Gómez, E.; Gómez, M.; Zandomeni, M.; Merlo, A.B. Peritoneal Surface Area: Measurements of 40 Structures Covered by Peritoneum: Correlation between Total Peritoneal Surface Area and the Surface Calculated by Formulas. Surg. Radiol. Anat. 2009, 31, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Yoshida, S.; Hara, A.; Ogura, S.; Tomita, H. Vascular Endothelial Injury Exacerbates Coronavirus disease 2019: The Role of Endothelial Glycocalyx Protection. Microcirculation 2020, e12654. [Google Scholar] [CrossRef] [PubMed]
- Bodega, F.; Sironi, C.; Porta, C.; Pecchiari, M.; Zocchi, L.; Agostoni, E. Mixed Lubrication After Rewetting of Blotted Pleural Mesothelium. Respir. Physiol. Neurobiol. 2013, 185, 369–373. [Google Scholar] [CrossRef]
- Van Teeffelen, J.W.; Brands, J.; Stroes, E.S.; Vink, H. Endothelial Glycocalyx: Sweet Shield of Blood Vessels. Trends Cardiovasc. Med. 2007, 17, 101–105. [Google Scholar] [CrossRef]
- Bodega, F.; Sironi, C.; Porta, C.; Zocchi, L.; Agostoni, E. Pleural Mesothelium Lubrication after Phospholipase Treatment. Respir. Physiol. Neurobiol. 2014, 194, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Chew, S.H.; Okazaki, Y.; Funahashi, S.; Namba, T.; Kato, T.; Enomoto, A.; Jiang, L.; Akatsuka, S.; Toyokuni, S. Metamorphosis of Mesothelial Cells with Active Horizontal Motility in Tissue Culture. Sci. Rep. 2013, 3, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley-Comer, A.J.; Herrick, S.E.; Al-Mishlab, T.; Prêle, C.M.; Laurent, G.J.; Mutsaers, S.E. Evidence for Incorporation of Free-Floating Mesothelial Cells as a Mechanism of Serosal Healing. J. Cell Sci. 2002, 115, 1383–1389. [Google Scholar]
- Mutsaers, S.E.; Whitaker, D.; Papadimitriou, J.M. Mesothelial Regeneration Is Not Dependent on Subserosal Cells. J. Pathol. 2000, 190, 86–92. [Google Scholar] [CrossRef]
- Hausmann, M.J.; Rogachev, B.; Weiler, M.; Chaimovitz, C.; Douvdevani, A. Accessory Role of Human Peritoneal Mesothelial Cells in Antigen Presentation and T-Cell Growth. Kidney Int. 2000, 57, 476–486. [Google Scholar] [CrossRef]
- Van Der Wal, B.C.H.; Hofland, L.J.; Marquet, R.L.; van Koetsveld, P.M.; van Rossen, M.E.; van Eijck, C.H. Paracrine Interactions between Mesothelial and Colon-Carcinoma Cells in a Rat Model. Int. J. Cancer 1997, 73, 885–890. [Google Scholar] [CrossRef]
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 14, 9–32. [Google Scholar] [PubMed] [Green Version]
- Lengyel, E. Ovarian Cancer Development and Metastasis. Am. J. Pathol. 2010, 177, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Dubeau, L. The Cell of Origin of Ovarian Epithelial Tumours. Lancet Oncol. 2008, 9, 1191–1197. [Google Scholar] [CrossRef] [Green Version]
- Labidi-Galy, S.I.; Papp, E.; Hallberg, D.; Niknafs, N.; Adleff, V.; Noe, M.; Bhattacharya, R.; Novak, M.; Jones, S.; Phallen, J.; et al. High grade serous ovarian carcinomas originate in the fallopian tube. Nat. Commun. 2017, 8, 1093. [Google Scholar] [CrossRef] [PubMed]
- Doglioni, C.; Dei Tos, A.P.; Laurino, L.; Iuzzolino, P.; Chiarelli, C.; Celio, M.R.; Viale, G. Calretinin: A Novel Immunocytochemical Marker for Mesothelioma. Am. J. Surg. Pathol. 1996, 20, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Gulyás, M.; Hjerpe, A. Proteoglycans and WT1 as Markers for Distinguishing Adenocarcinoma, Epithelioid Mesothelioma, and Benign Mesothelium. J. Pathol. 2003, 199, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Schalling, M.; Bernard, A.; Maheswaran, S.; Shipley, G.C.; Roberts, D.; Fletcher, J.; Shipman, R.; Rheinwald, J.; Demetri, G.; et al. The Wilms Tumour Gene WT1 is Expressed in Murine Mesoderm-Derived Tissues and Mutated in a Human Mesothelioma. Nat. Genet. 1993, 4, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, N.G. Podoplanin: A Novel Diagnostic Immunohistochemical Marker. Adv. Anat. Pathol. 2006, 13, 83–88. [Google Scholar] [CrossRef]
- Hanna, A.; Pang, Y.; Bedrossian, C.W.; Dejmek, A.; Michael, C.W. Podoplanin Is a Useful Marker for Identifying Mesothelioma in Malignant Effusions. Diagn. Cytopathol. 2008, 36, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Hassan, R.; Kreitman, R.J.; Pastan, I.; Willingham, M.C. Localization of Mesothelin in Epithelial Ovarian Cancer. Appl. Immunohistochem. Mol. Morphol. 2005, 13, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.; Pastan, I. Molecular Cloning of Mesothelin, a Differentiation Antigen Present on Mesothelium, Mesotheliomas, and Ovarian Cancers. Proc. Natl. Acad. Sci. USA 1996, 93, 136–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rump, A.; Morikawa, Y.; Tanaka, M.; Minami, S.; Umesaki, N.; Takeuchi, M.; Miyajima, A. Binding of Ovarian Cancer Antigen CA125/MUC16 to Mesothelin Mediates Cell Adhesion. J. Biol. Chem. 2004, 279, 9190–9198. [Google Scholar] [CrossRef] [Green Version]
- Gubbels, J.A.A.; Belisle, J.; Onda, M.; Rancourt, C.; Migneault, M.; Ho, M.; Bera, T.K.; Connor, J.; Sathyanarayana, B.K.; Lee, B.; et al. Mesothelin-MUC16 Binding Is a High Affinity, N-Glycan Dependent Interaction That Facilitates Peritoneal Metastasis of Ovarian Tumors. Mol. Cancer 2006, 5, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamori-Katayama, M.; Kaiho, A.; Ishizu, Y.; Okamura-Oho, Y.; Hino, O.; Abe, M.; Kishimoto, T.; Sekihara, H.; Nakamura, Y.; Suzuki, H.; et al. LRRN4 and UPK3B Are Markers of Primary Mesothelial Cells. PLoS ONE 2011, 6, e25391. [Google Scholar] [CrossRef] [Green Version]
- Davidson, B.; Nielsen, S.; Christensen, J.; Asschenfeldt, P.; Berner, A.; Risberg, B.; Johansen, P. The Role of Desmin and N-Cadherin in Effusion Cytology: A Comparative Study Using Established Markers of Mesothelial and Epithelial Cells. Am. J. Surg. Pathol. 2001, 25, 1405–1412. [Google Scholar] [CrossRef]
- Yamada, T.; Jiping, J.; Endo, R.; Gotoh, M.; Shimosato, Y.; Hirohashi, S. Molecular Cloning of a Cell-Surface Glycoprotein That Can Potentially Discriminate Mesothelium From Epithelium: Its Identification as Vascular Cell Adhesion Molecule 1. Br. J. Cancer 1995, 71, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.T.; Chang, Y.T.; Pan, S.Y.; Chou, Y.H.; Chang, F.C.; Yeh, P.Y.; Liu, Y.H.; Chiang, W.C.; Chen, Y.M.; Wu, K.D.; et al. Lineage Tracing Reveals Distinctive Fates for Mesothelial Cells and Submesothelial Fibroblasts During Peritoneal Injury. J. Am. Soc. Nephrol. 2014, 25, 2847–2858. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, C.X.; Müller, S.; Keerthivasan, S.; Koeppen, H.; Hung, J.; Gierke, S.; Breart, B.; Foreman, O.; Bainbridge, T.W.; Castiglioni, A.; et al. Single-Cell RNA Sequencing Reveals Stromal Evolution Into LRRC15+ Myofibroblasts as a Determinant of Patient Response to Cancer Immunotherapy. Cancer Discov. 2020, 10, 232–253. [Google Scholar] [CrossRef] [Green Version]
- Dasari, S.; Fang, Y.; Mitra, A.K. Cancer Associated Fibroblasts: Naughty Neighbors That Drive Ovarian Cancer Progression. Cancers 2018, 10, 406. [Google Scholar] [CrossRef] [Green Version]
- Kenny, H.A.; Chiang, C.Y.; White, E.A.; Schryver, E.M.; Habis, M.; Romero, I.L.; Ladanyi, A.; Penicka, C.V.; George, J.; Matlin, K.; et al. Mesothelial Cells Promote Early Ovarian Cancer Metastasis Through Fibronectin Secretion. J. Clin. Investig. 2014, 124, 4614–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Yang, Z.; Xu, S.; Li, X.; Yang, X.; Jin, P.; Liu, Y.; Zhou, X.; Zhang, T.; Gong, C.; et al. Heterotypic CAF-Tumor Spheroids Promote Early Peritoneal Metastatis of Ovarian Cancer. J. Exp. Med. 2019, 216, 688–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matte, I.; Lane, D.; Laplante, C.; Rancourt, C.; Piché, A. Profiling of Cytokines in Human Epithelial Ovarian Cancer Ascites. Am. J. Cancer Res. 2012, 2, 566–580. [Google Scholar] [PubMed]
- Nakamura, M.; Ono, Y.J.; Kanemura, M.; Tanaka, T.; Hayashi, M.; Terai, Y.; Ohmichi, M. Hepatocyte Growth Factor Secreted by Ovarian Cancer Cells Stimulates Peritoneal Implantation via the Mesothelial-Mesenchymal Transition of the Peritoneum. Gynecol. Oncol. 2015, 139, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Santin, A.D.; Bellone, S.; Ravaggi, A.; Roman, J.; Smith, C.V.; Pecorelli, S.; Cannon, M.J.; Parham, G.P. Increased Levels of Interleukin-10 and Transforming Growth Factor-β in the Plasma and Ascitic Fluid of Patients with Advanced Ovarian Cancer. Br. J. Obstet. Gynaecol. 2001, 108, 804–808. [Google Scholar]
- Moradi, M.M.; Carson, L.F.; Weinberg, B.; Haney, A.F.; Twiggs, L.B.; Ramakrishnan, S. Serum and Ascitic Fluid Levels of Interleukin-1, Interleukin-6, and Tumor Necrosis Factor-Alpha in Patients with Ovarian Epithelial Cancer. Cancer 1993, 72, 2433–2440. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Lara-Pezzi, E.; Selgas, R.; Ramírez-Huesca, M.; Domínguez-Jiménez, C.; Jiménez-Heffernan, J.A.; Aguilera, A.; Sánchez-Tomero, J.A.; Bajo, M.A.; Alvarez, V.; et al. Peritoneal Dialysis and Epithelial-to-Mesenchymal Transition of Mesothelial Cells. N. Engl. J. Med. 2003, 348, 403–413. [Google Scholar] [CrossRef]
- Aroeira, L.S.; Aguilera, A.; Sánchez-Tomero, J.A.; Bajo, M.A.; del Peso, G.; Jiménez-Heffernan, J.A.; Selgas, R.; López-Cabrera, M. Epithelial to Mesenchymal Transition and Peritoneal Membrane Failure in Peritoneal Dialysis Patients: Pathologic Significance and Potential Therapeutic Interventions. J. Am. Soc. Nephrol. 2007, 18, 2004–2013. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Bajo, M.A.; del Peso, G.; Yu, X.; Selgas, R. Preventing peritoneal membrane fibrosis in peritoneal dialysis patients. Kidney Int. 2016, 90, 515–524. [Google Scholar] [CrossRef]
- Sluiter, N.; de Cuba, E.; Kwakman, R.; Kazemier, G.; Meijer, G.; Te Velde, E.A. Adhesion Molecules in Peritoneal Dissemination: Function, Prognostic Relevance and Therapeutic Options. Clin. Exp. Metastasis 2016, 33, 401–416. [Google Scholar] [CrossRef]
- Weidle, U.H.; Birzele, F.; Kollmorgen, G.; Rueger, R. Mechanisms and Targets Involved in Dissemination of Ovarian Cancer. Cancer Genom. Proteom. 2016, 13, 407–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casey, R.C.; Oegema, T.R.; Skubitz, K.M.; Pambuccian, S.E.; Grindle, S.M.; Skubitz, A.P. Cell Membrane Glycosylation Mediates the Adhesion, Migration, and Invasion of Ovarian Carcinoma Cells. Clin. Exp. Metastasis 2003, 20, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Heyman, L.; Kellouche, S.; Fernandes, J.; Dutoit, S.; Poulain, L.; Carreiras, F. Vitronectin and Its Receptors Partly Mediate Adhesion of Ovarian Cancer Cells to Peritoneal Mesothelium In Vitro. Tumor Biol. 2008, 29, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.J.; Catterall, J.B.; Jones, L.M.H.; Turner, G.A. Human Ovarian Tumour Cells Can Bind Hyaluronic Acid via Membrane CD44: A Possible Step in Peritoneal Metastasis. Clin. Exp. Metastasis 1996, 14, 325–334. [Google Scholar] [CrossRef]
- Aruffo, A.; Stamenkovic, I.; Melnick, M.; Underhill, C.B.; Seed, B. CD44 Is the Principal Cell Surface Receptor for Hyaluronate. Cell 1990, 61, 1303–1313. [Google Scholar] [CrossRef]
- Jalkanen, S.; Jalkanen, M. Lymphocyte CD44 Binds the COOH-Terminal Heparin-Binding Domain of Fibronectin. J. Cell Biol. 1992, 116, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Lessan, K.; Aguiar, D.J.; Oegema, T.; Siebenson, L.; Skubitz, A.P.N. CD44 and β1 Integrin Mediate Ovarian Carcinoma Cell Adhesion to Peritoneal Mesothelial Cells. Am. J. Pathol. 1999, 154, 1525–1537. [Google Scholar] [CrossRef]
- Moos, M.; Tacke, R.; Scherer, H.; Teplow, D.; Früh, K.; Schachner, M. Neural Adhesion Molecule Ll as a Member of the Immunoglobulin Superfamily with Binding Domains Similar to Fibronectin. Nature 1988, 334, 701–703. [Google Scholar] [CrossRef]
- Stoeck, A.; Schlich, S.; Issa, Y.; Gschwend, V.; Wenger, T.; Herr, I.; Marmé, A.; Bourbie, S.; Altevogt, P.; Gutwein, P. L1 on Ovarian Carcinoma Cells Is a Binding Partner for Neuropilin-1 on Mesothelial Cells. Cancer Lett. 2006, 239, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Arlt, M.J.E.; Novak-Hofer, I.; Gast, D.; Gschwend, V.; Moldenhauer, G.; Grünberg, J.; Honer, M.; Schubiger, P.A.; Altevogt, P.; Krüger, A. Efficient Inhibition of Intra-Peritoneal Tumor Growth and Dissemination of Human Ovarian Carcinoma Cells in Nude Mice by Anti-L1-Cell Adhesion Molecule Monoclonal Antibody Treatment. Cancer Res. 2006, 66, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.A.; Artemenko, M.; Tang, M.K.S.; Wong, A.S.T. Selectins: An Important Family of Glycan-Binding Cell Adhesion Molecules in Ovarian Cancer. Cancers 2020, 12, 2238. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Ip, C.K.M.; Tang, M.Y.H.; Tang, M.K.S.; Tong, Y.; Zhang, J.; Hassan, A.A.; Mak, A.S.C.; Yung, S.; Chan, T.M.; et al. Sialyl Lewisx-P-Selectin Cascade Mediates Tumor–Mesothelial Adhesion in Ascitic Fluid Shear Flow. Nat. Commun. 2019, 10, 2406. [Google Scholar] [CrossRef] [Green Version]
- Carroll, M.J.; Fogg, K.C.; Patel, H.A.; Krause, H.B.; Mancha, A.S.; Patankar, M.S.; Weisman, P.S.; Barroilhet, L.; Kreeger, P.K. Alternatively-Activated Macrophages Upregulate Mesothelial Expression of P-Selectin to Enhance Adhesion of Ovarian Cancer Cells. Cancer Res. 2018, 78, 3560–3573. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Rooper, L.; Xie, J.; Kajdacsy-Balla, A.A.; Barbolina, M.V. Fractalkine Receptor CX(3)CR1 Is Expressed in Epithelial Ovarian Carcinoma Cells and Required for Motility and Adhesion to Peritoneal Mesothelial Cells. Mol. Cancer Res. 2012, 10, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasui, H.; Kajiyama, H.; Tamauchi, S.; Suzuki, S.; Peng, Y.; Yoshikawa, N.; Sugiyama, M.; Nakamura, K.; Kikkawa, F. CCL2 Secreted From Cancer-Associated Mesothelial Cells Promotes Peritoneal Metastasis of Ovarian Cancer Cells Through the P38-MAPK Pathway. Clin. Exp. Metastasis 2020, 37, 145–158. [Google Scholar] [CrossRef]
- Westermann, A.M.; Havik, E.; Postma, F.R.; Beijnen, J.H.; Dalesio, O.; Moolenaar, W.H.; Rodenhuis, S. Malignant effusions contain lysophosphatidic acid (LPA)-like activity. Ann. Oncol. 1998, 9, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.J.; Schwartz, B.; Washington, M.; Kennedy, A.; Webster, K.; Belinson, J.; Xu, Y. Electrospray ionization mass spectrometry analysis of lysophospholipids in human ascitic fluids: Comparison of the lysophospholipid contents in malignant vs nonmalignant ascitic fluids. Anal. Biochem. 2001, 290, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Xiao, Y.J.; Singh, L.S.; Zhao, X.; Zhao, Z.; Feng, L.; Rose, T.M.; Prestwich, G.D.; Xu, Y. Lysophosphatidic acid is constitutively produced by human peritoneal mesothelial cells and enhances adhesion, migration, and invasion of ovarian cancer cells. Cancer Res. 2006, 66, 3006–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Said, N.A.; Najwer, I.; Socha, M.J.; Fulton, D.J.; Mok, S.C.; Motamed, K. SPARC inhibits LPA-mediated mesothelial-ovarian cancer cell crosstalk. Neoplasia 2007, 9, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwanicki, M.P.; Davidowitz, R.A.; Ng, M.R.; Besser, A.; Muranen, T.; Merritt, M.; Danuser, G.; Ince, T.A.; Brugge, J.S. Ovarian Cancer Spheroids Use Myosin-Generated Force to Clear the Mesothelium. Cancer Discov. 2011, 1, 144–157. [Google Scholar] [CrossRef] [Green Version]
- Kenny, H.A.; Nieman, K.M.; Mitra, A.K.; Lengyel, E. The First Line of Intra-Abdominal Metastatic Attack: Breaching the Mesothelial Cell Layer. Cancer Discov. 2011, 1, 100–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, M.; Yamakita, Y.; Kajiyama, H.; Senga, T.; Koya, Y.; Yamashita, M.; Nawa, A.; Kikkawa, F. Filopodia Play an Important Role in the Trans-Mesothelial Migration of Ovarian Cancer Cells. Exp. Cell Res. 2020, 392, 112011. [Google Scholar] [CrossRef]
- Kalluri, R. The Biology and Function of Fibroblasts in Cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Satoyoshi, R.; Aiba, N.; Yanagihara, K.; Yashiro, M.; Tanaka, M. Tks5 Activation in Mesothelial Cells Creates Invasion Front of Peritoneal Carcinomatosis. Oncogene 2015, 34, 3176–3187. [Google Scholar] [CrossRef]
- Givel, A.M.; Kieffer, Y.; Scholer-Dahirel, A.; Sirven, P.; Cardon, M.; Pelon, F.; Magagna, I.; Gentric, G.; Costa, A.; Bonneau, C.; et al. MiR200-Regulated CXCL12β Promotes Fibroblast Heterogeneity and Immunosuppression in Ovarian Cancers. Nat. Commun. 2018, 9, 1056. [Google Scholar] [CrossRef]
- Santiskulvong, C.; Konecny, G.E.; Fekete, M.; Chen, K.Y.; Karam, A.; Mulholland, D.; Eng, C.; Wu, H.; Song, M.; Dorigo, O. Dual Targeting of Phosphoinositide 3-Kinase and Mammalian Target Of Rapamycin Using NVP-BEZ235 as a Novel Therapeutic Approach in Human Ovarian Carcinoma. Clin. Cancer Res. 2011, 17, 2373–2384. [Google Scholar] [CrossRef] [Green Version]
- Westfall, S.D.; Skinner, M.K. Inhibition of Phosphatidylinositol 3-Kinase Sensitizes Ovarian Cancer Cells to Carboplatin and Allows Adjunct Chemotherapy Treatment. Mol. Cancer Ther. 2005, 4, 1764–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasano, T.; Mabuchi, S.; Kuroda, H.; Kawano, M.; Matsumoto, Y.; Takahashi, R.; Hisamatsu, T.; Sawada, K.; Hashimoto, K.; Isobe, A.; et al. Preclinical Efficacy for AKT Targeting in Clear Cell Carcinoma of the Ovary. Mol. Cancer Res. 2015, 13, 795–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Sawah, E.; Chen, X.; Marchion, D.C.; Xiong, Y.; Ramirez, I.J.; Abbasi, F.; Bou Zgheib, N.; Chon, H.S.; Wenham, R.M.; Apte, S.M.; et al. Perifosine, An AKT Inhibitor, Modulates Ovarian Cancer Cell Line Sensitivity to Cisplatin-Induced Growth Arrest. Gynecol. Oncol. 2013, 131, 207–212. [Google Scholar] [CrossRef]
- Ricciardelli, C.; Lokman, N.A.; Ween, M.P.; Oehler, M.K. WOMEN IN CANCER THEMATIC REVIEW: Ovarian Cancer-Peritoneal Cell Interactions Promote Extracellular Matrix Processing. Endocr. Relat. Cancer 2016, 23, T155–T168. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.; Weng, D.; Chen, G.; Tao, W.; Zhu, T.; Yang, X.; Meng, L.; Wang, S.; Lu, Y.; Ma, D. Activation of Fibronectin/PI-3K/Akt2 Leads to Chemoresistance to Docetaxel by Regulating Survivin Protein Expression in Ovarian and Breast Cancer Cells. Cancer Lett. 2008, 261, 108–119. [Google Scholar] [CrossRef]
- Yu, Q.; Xue, Y.; Liu, J.; Xi, Z.; Li, Z.; Liu, Y. Fibronectin Promotes the Malignancy of Glioma Stem-Like Cells via Modulation of Cell Adhesion, Differentiation, Proliferation and Chemoresistance. Front. Mol. Neurosci. 2018, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.; Tinker, A.V.; Friedlander, M. “Platinum Resistant” Ovarian Cancer: What Is It, Who to Treat and How to Measure Benefit? Gynecol. Oncol. 2014, 133, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ma, Q.; Liu, Q.; Yu, H.; Zhao, L.; Shen, S.; Yao, J. Blockade of SDF-1/CXCR4 signalling inhibits pancreatic cancer progression in vitro via inactivation of canonical Wnt pathway. Br. J. Cancer 2008, 99, 1695–1703. [Google Scholar] [CrossRef] [PubMed]
- Froeling, F.E.M.; Feig, C.; Chelala, C.; Dobson, R.; Mein, C.E.; Tuveson, D.A.; Clevers, H.; Hart, I.R.; Kocher, H.M. Retinoic acid-induced pancreatic stellate cell quiescence reduces paracrine Wntβ-catenin signaling to slow tumor progression. Gastroenterology 2011, 141, 1486–1497. [Google Scholar] [CrossRef]
- Sherman, M.H.; Yu, R.T.; Engle, D.D.; Ding, N.; Atkins, A.R.; Tiriac, H.; Collisson, E.A.; Connor, F.; Van Dyke, T.; Kozlov, S.; et al. Vitamin D receptor-mediated stromal reprogramming suppresses pancreatitis and enhances pancreatic cancer therapy. Cell 2014, 159, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Ferrer-Mayorga, G.; Gómez-López, G.; Barbáchano, A.; Fernández-Barral, A.; Peña, C.; Pisano, D.G.; Cantero, R.; Rojo, F.; Muñoz, A.; Larriba, M.J. Vitamin D receptor expression and associated gene signature in tumour stromal fibroblasts predict clinical outcome in colorectal cancer. Gut 2017, 66, 1449–1462. [Google Scholar] [CrossRef]
- Freedman, J.D.; Duffy, M.R.; Lei-Rossmann, J.; Muntzer, A.; Scott, E.M.; Hagel, J.; Campo, L.; Bryant, R.J.; Verrill, C.; Lambert, A.; et al. An Oncolytic Virus Expressing a T-cell Engager Simultaneously Targets Cancer and Immunosuppressive Stromal Cells. Cancer Res. 2018, 78, 6852–6865. [Google Scholar] [CrossRef] [Green Version]
- De Sostoa, J.; Fajardo, C.A.; Moreno, R.; Ramos, M.D.; Farrera-Sal, M.; Alemany, R. Targeting the tumor stroma with an oncolytic adenovirus secreting a fibroblast activation protein-targeted bispecific T-cell engager. J. Immunother. Cancer 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Kajiyama, H.; Suzuki, S.; Yoshihara, M.; Nishino, K.; Yoshikawa, N.; Utsumi, F.; Niimi, K.; Mizuno, M.; Kawai, M.; Oguchi, H.; et al. Possible Existence of Occult Metastasis in Patients with Ovarian Clear-Cell Carcinoma Who Underwent Complete Resection without Any Residual Tumors. Oncotarget 2018, 9, 6298–6307. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Markers | Location | Articles |
|---|---|---|
| Calretinin | Intracellular, Cytosol | [36] |
| Podoplanin | Plasma membrane | [39,40] |
| WT1 | Intracellular, Nucleoplasm | [37,38] |
| Mesothelin | Intracellular, Vesicles and Nucleoplasm | [41,42,43,44] |
| Desmin | Intracellular, Intermediate filaments | [46] |
| Cytokeratin 5/6 | Intracellular, Intermediate filaments | [46] |
| N-cadherin | Plasma membrane | [46] |
| LRRN4 | Intracellular, Nucleoli | [45] |
| UPK3B | Plasma membrane, Intracellular, Cytosol | [45] |
| Ligands | Receptors | References | |
|---|---|---|---|
| ECM | |||
| Laminin, fibronectin, vitronectin, collagen I, and IV | Integrins | [16,20,51,60,61] | |
| Vitronectin | uPAR | [63] | |
| Hyaluronan | CD44 | [62,64,65,66,67] | |
| Membrane protein | |||
| VCAM1 | Integrins | [60,61,68] | |
| Neuropilin-1 | L1CAM | [60,61,69,70] | |
| Mesothelin | MUC16 | [43,44] | |
| P-selectin | sialyl-Lewisx (sLex)CD24 | [71,72,73] | |
| Chemokine | |||
| SDF-1 | CXCR4 | [15] | |
| CX3CL1 | CXCR1 | [74] | |
| CCL2 | CCR2 | [75] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mogi, K.; Yoshihara, M.; Iyoshi, S.; Kitami, K.; Uno, K.; Tano, S.; Koya, Y.; Sugiyama, M.; Yamakita, Y.; Nawa, A.; et al. Ovarian Cancer-Associated Mesothelial Cells: Transdifferentiation to Minions of Cancer and Orchestrate Developing Peritoneal Dissemination. Cancers 2021, 13, 1352. https://doi.org/10.3390/cancers13061352
Mogi K, Yoshihara M, Iyoshi S, Kitami K, Uno K, Tano S, Koya Y, Sugiyama M, Yamakita Y, Nawa A, et al. Ovarian Cancer-Associated Mesothelial Cells: Transdifferentiation to Minions of Cancer and Orchestrate Developing Peritoneal Dissemination. Cancers. 2021; 13(6):1352. https://doi.org/10.3390/cancers13061352
Chicago/Turabian StyleMogi, Kazumasa, Masato Yoshihara, Shohei Iyoshi, Kazuhisa Kitami, Kaname Uno, Sho Tano, Yoshihiro Koya, Mai Sugiyama, Yoshihiko Yamakita, Akihiro Nawa, and et al. 2021. "Ovarian Cancer-Associated Mesothelial Cells: Transdifferentiation to Minions of Cancer and Orchestrate Developing Peritoneal Dissemination" Cancers 13, no. 6: 1352. https://doi.org/10.3390/cancers13061352