
Distinct Effects of Brain Activation Using tDCS and Observational Practice: Implications for Motor Rehabilitation
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
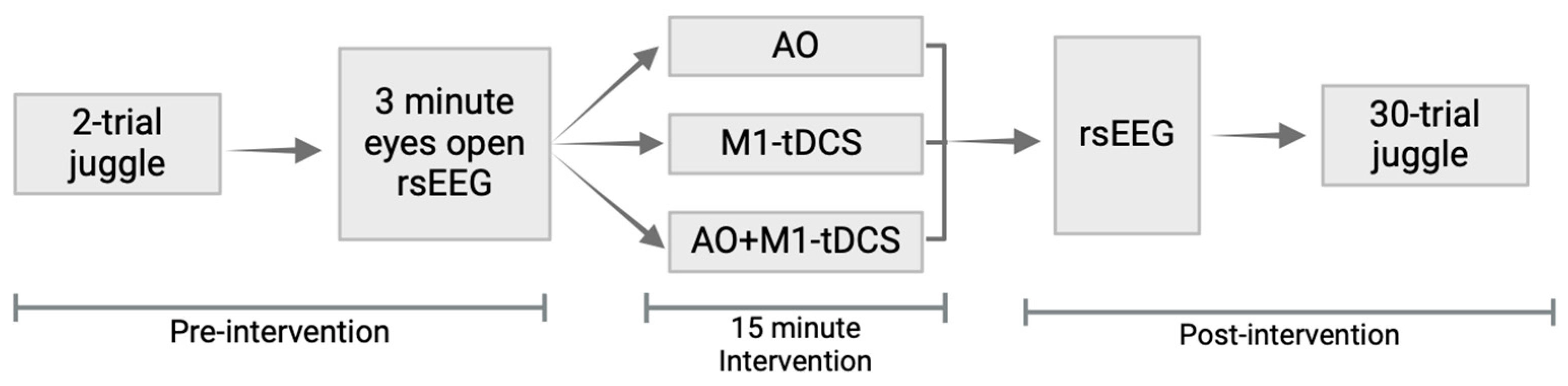
2.2. Study Design
2.3. Juggling Task
2.4. Interventions
2.4.1. Video Observation (AO)
2.4.2. M1-tDCS
2.5. EEG Recording and Preprocessing
2.6. Effective Connectivity
3. Results
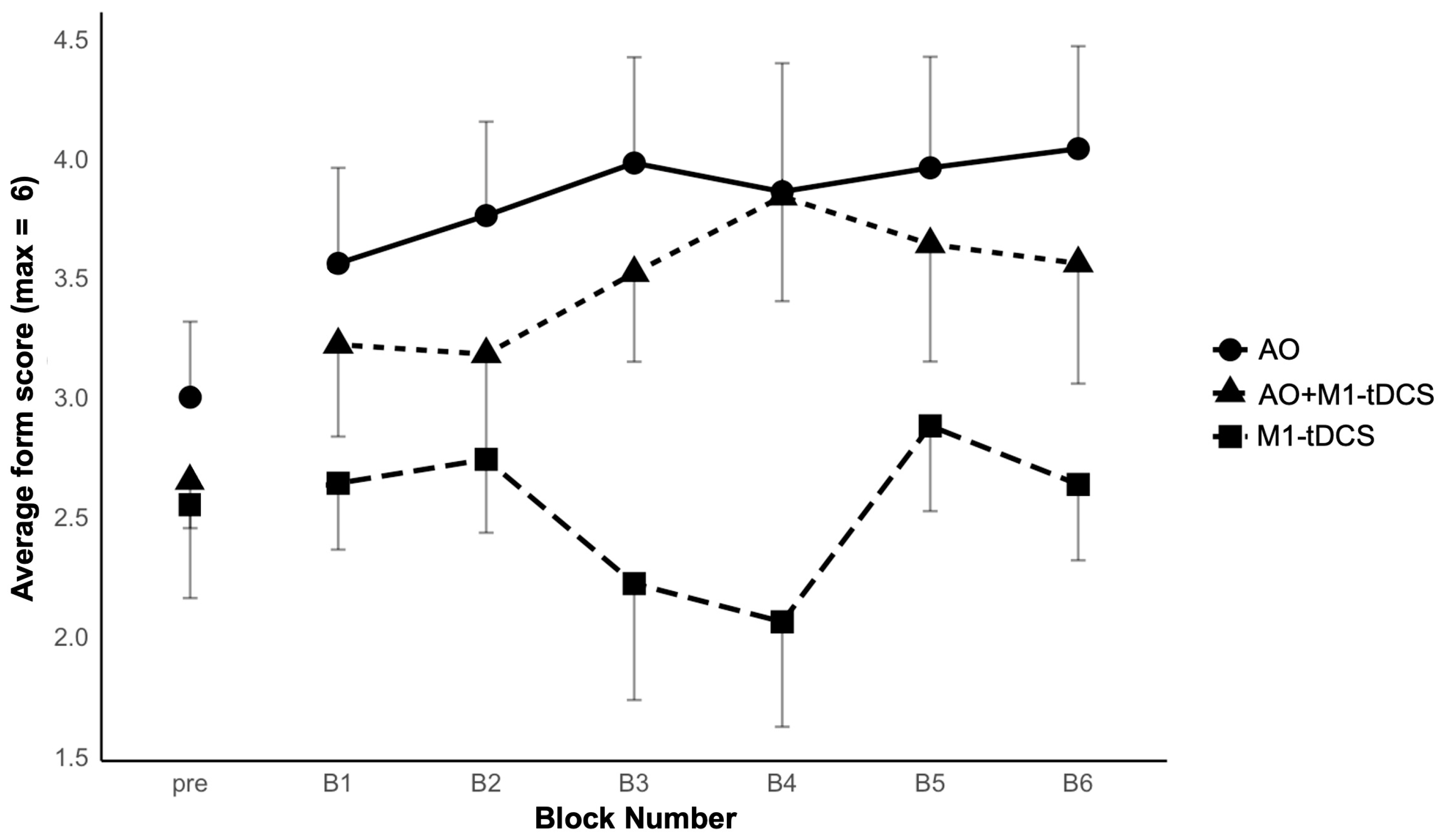
3.1. Juggling
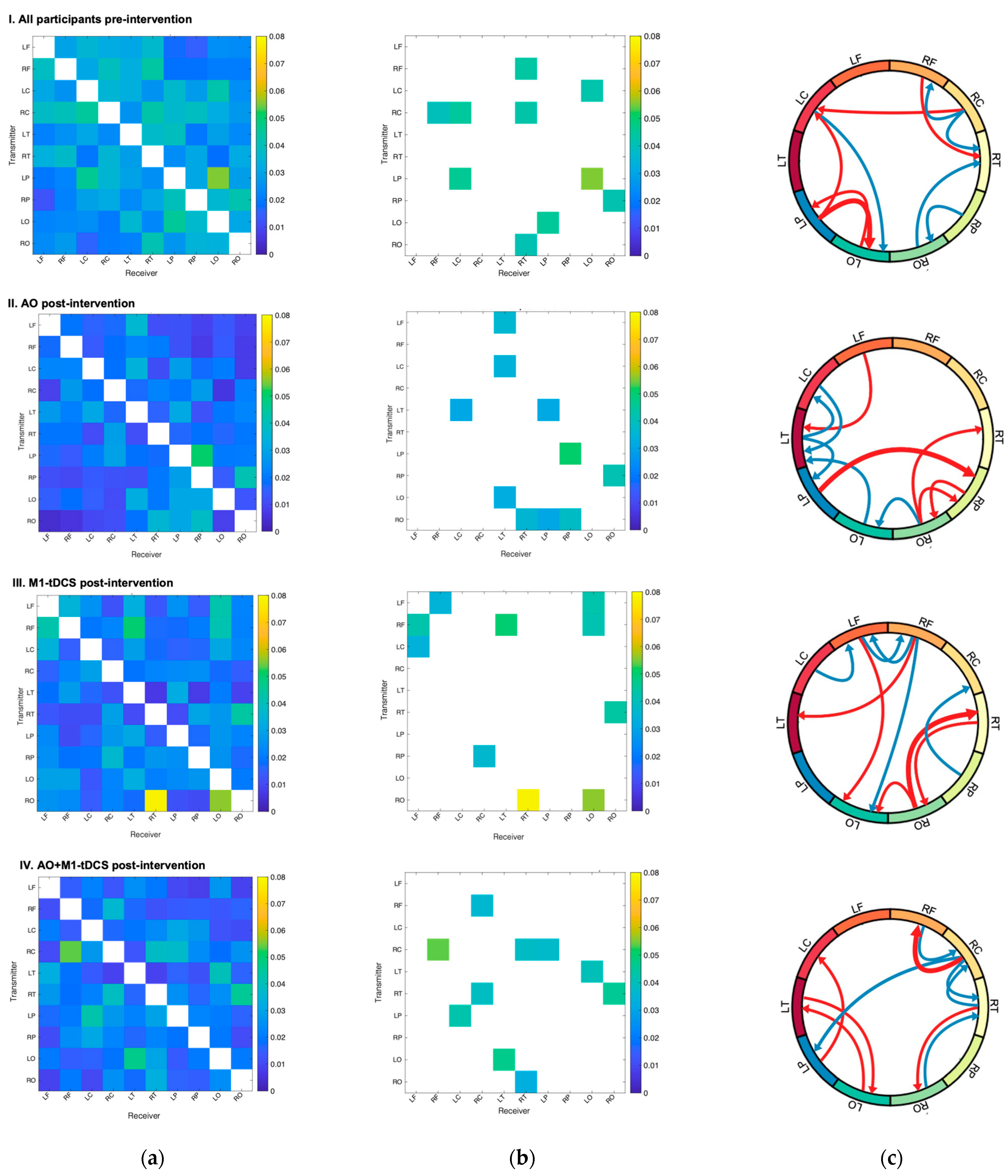
3.2. Qualitative Comparison of Effective Connectivity Patterns
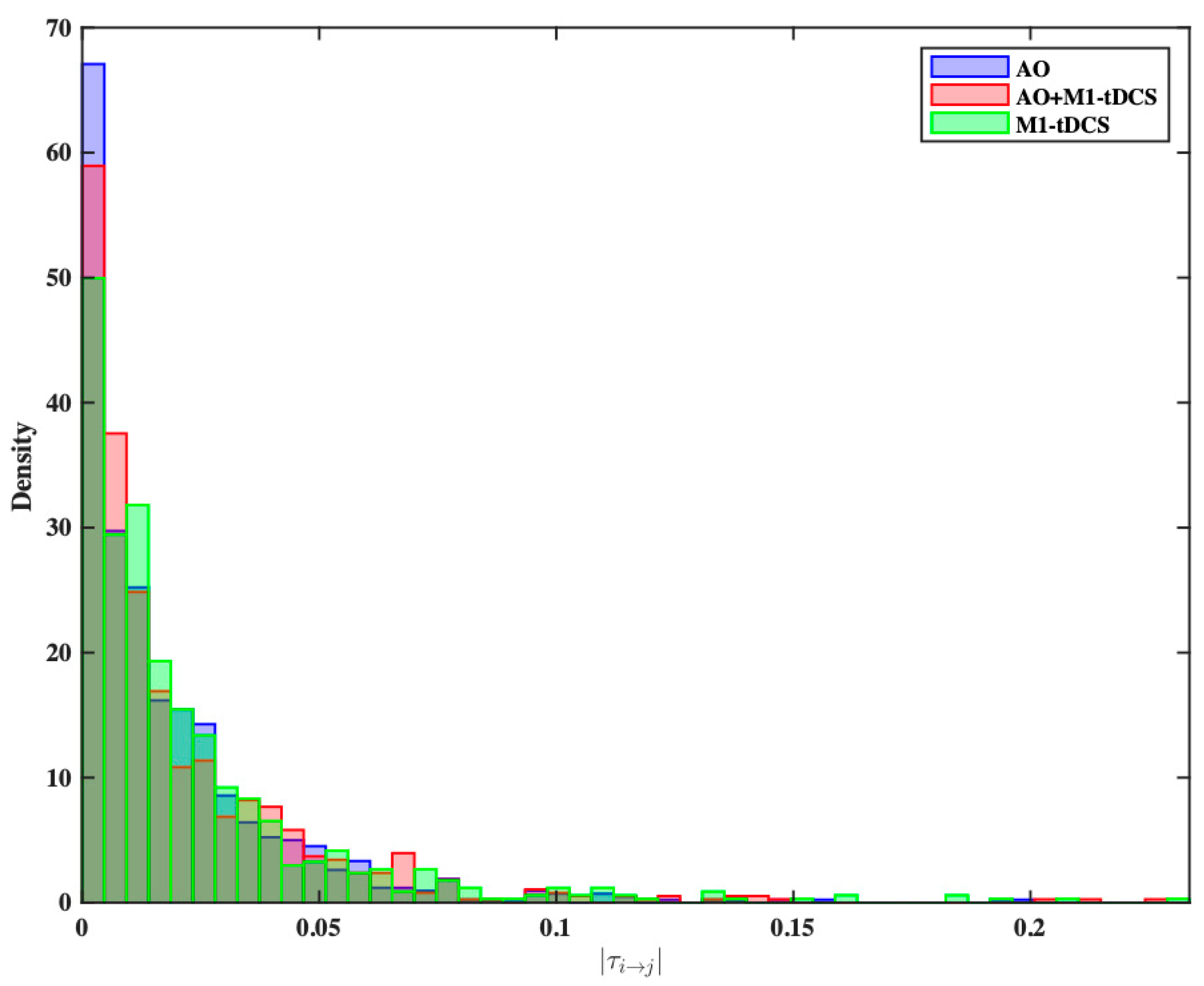
3.3. Statistical Comparison of Effective Connectivity Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hodges, N.J. Observations on Action-Observation Research: An Autobiographical Retrospective Across the Past Two Decades. Kinesiol. Rev. 2017, 6, 240–260. [Google Scholar] [CrossRef]
- Hodges, N.J.; Williams, A.M.; Hayes, S.J.; Breslin, G. What is modelled during observational learning? J. Sports Sci. 2007, 25, 531–545. [Google Scholar] [CrossRef]
- Ramsey, R.; Kaplan, D.M.; Cross, E.S. Watch and Learn: The Cognitive Neuroscience of Learning from Others’ Actions. Trends Neurosci. 2021, 44, 478–491. [Google Scholar] [CrossRef]
- Calvo-Merino, B.; Grèzes, J.; Glaser, D.E.; Passingham, R.E.; Haggard, P. Seeing or Doing? Influence of Visual and Motor Familiarity in Action Observation. Curr. Biol. 2006, 16, 1905–1910. [Google Scholar] [CrossRef]
- Grèzes, J.; Armony, J.L.; Rowe, J.; Passingham, R.E. Activations related to “mirror” and “canonical” neurones in the human brain: An fMRI study. NeuroImage 2003, 18, 928–937. [Google Scholar] [CrossRef]
- Virji-Babul, N.; Moiseev, A.; Cheung, T.; Weeks, D.; Cheyne, D.; Ribary, U. Spatial-temporal dynamics of cortical activity underlying reaching and grasping. Hum. Brain Mapp. 2009, 31, 160–171. [Google Scholar] [CrossRef]
- Alhajri, N.; Hodges, N.J.; Zwicker, J.G.; Virji-Babul, N. Mu Suppression Is Sensitive to Observational Practice but Results in Different Patterns of Activation in Comparison with Physical Practice. Neural Plast. 2018, 2018, e8309483. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527 Pt 3, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Paulus, W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef] [PubMed]
- Bersani, F.S.; Minichino, A.; Bernabei, L.; Spagnoli, F.; Corrado, A.; Vergnani, L.; Mannarelli, D.; Pauletti, C.; Fattapposta, F.; Biondi, M.; et al. Prefronto-cerebellar tDCS enhances neurocognition in euthymic bipolar patients. Findings from a placebo-controlled neuropsychological and psychophysiological investigation. J. Affect. Disord. 2017, 209, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Bikson, M.; Bulow, P.; Stiller, J.W.; Datta, A.; Battaglia, F.; Karnup, S.V.; Postolache, T.T. Transcranial direct current stimulation for major depression: A general system for quantifying transcranial electrotherapy dosage. Curr. Treat. Options Neurol. 2008, 10, 377–385. [Google Scholar] [CrossRef]
- Kim, W.S.; Lee, K.; Kim, S.; Cho, S.; Paik, N.-J. Transcranial direct current stimulation for the treatment of motor impairment following traumatic brain injury. J. NeuroEng. Rehabil. 2019, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- Karok, S.; Witney, A.G. Enhanced Motor Learning Following Task-Concurrent Dual Transcranial Direct Current Stimulation. PLoS ONE 2013, 8, e85693. [Google Scholar] [CrossRef]
- Rumpf, J.-J.; Wegscheider, M.; Hinselmann, K.; Fricke, C.; King, B.R.; Weise, D.; Klann, J.; Binkofski, F.; Buccino, G.; Karni, A.; et al. Enhancement of motor consolidation by post-training transcranial direct current stimulation in older people. Neurobiol. Aging 2017, 49, 1–8. [Google Scholar] [CrossRef]
- Ehsani, F.; Bakhtiary, A.H.; Jaberzadeh, S.; Talimkhani, A.; Hajihasani, A. Differential effects of primary motor cortex and cerebellar transcranial direct current stimulation on motor learning in healthy individuals: A randomized double-blind sham-controlled study. Neurosci. Res. 2016, 112, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Goodwill, A.; Reynolds, J.; Daly, R.; Kidgell, D. Formation of cortical plasticity in older adults following tDCS and motor training. Front. Aging Neurosci. 2013, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Pixa, N.H.; Pollok, B. Effects of tDCS on Bimanual Motor Skills: A Brief Review. Front. Behav. Neurosci. 2018, 12, 63. [Google Scholar] [CrossRef]
- Friston, K.J. Functional and Effective Connectivity: A Review. Brain Connect. 2011, 1, 13–36. [Google Scholar] [CrossRef]
- Calzolari, S.; Jalali, R.; Fernández-Espejo, D. Characterising stationary and dynamic effective connectivity changes in the motor network during and after tDCS. NeuroImage 2023, 269, 119915. [Google Scholar] [CrossRef]
- Hristopulos, D.T.; Babul, A.; Babul, S.; Brucar, L.R.; Virji-Babul, N. Disrupted Information Flow in Resting-State in Adolescents with Sports Related Concussion. Front. Hum. Neurosci. 2019, 13, 419. [Google Scholar] [CrossRef]
- Liang, X.S. The Liang-Kleeman Information Flow: Theory and Applications. Entropy 2013, 15, 327–360. [Google Scholar] [CrossRef]
- Berchicci, M.; Quinzi, F.; Dainese, A.; Di Russo, F. Time-source of neural plasticity in complex bimanual coordinative tasks: Juggling. Behav. Brain Res. 2017, 328, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Hodges, N.J.; Coppola, T. What we think we learn from watching others: The moderating role of ability on perceptions of learning from observation. Psychol. Res. 2015, 79, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Zentgraf, K.; Munzert, J. Effects of attentional-focus instructions on movement kinematics. Psychol. Sport Exerc. 2009, 10, 520–525. [Google Scholar] [CrossRef]
- Tadel, F.; Baillet, S.; Mosher, J.C.; Pantazis, D.; Leahy, R.M. Brainstorm: A User-Friendly Application for MEG/EEG Analysis. Comput. Intell. Neurosci. 2011, 2011, e879716. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.S. Unraveling the cause-effect relation between time series. Phys. Rev. E 2014, 90, 052150. [Google Scholar] [CrossRef]
- Chong, T.T.J.; Cunnington, R.; Williams, M.A.; Kanwisher, N.; Mattingley, J.B. fMRI adaptation reveals mirror neurons in human inferior parietal cortex. Curr. Biol. 2008, 18, 1576–1580. [Google Scholar] [CrossRef]
- Hamilton, A.F.d.C.; Grafton, S.T. Goal representation in human anterior intraparietal sulcus. J. Neurosci. 2006, 26, 1133–1137. [Google Scholar] [CrossRef]
- Urgen, B.A.; Orban, G.A. The unique role of parietal cortex in action observation: Functional organization for communicative and manipulative actions. NeuroImage 2021, 237, 118220. [Google Scholar] [CrossRef]
- Pitcher, D.; Ungerleider, L.G. Evidence for a Third Visual Pathway Specialized for Social Perception. Trends Cogn. Sci. 2021, 25, 100–110. [Google Scholar] [CrossRef]
- Lang, N.; Siebner, H.R.; Ward, N.S.; Lee, L.; Nitsche, M.A.; Paulus, W.; Rothwell, J.C.; Lemon, R.N.; Frackowiak, R.S. How does transcranial DC stimulation of the primary motor cortex alter regional neuronal activity in the human brain? Eur. J. Neurosci. 2005, 22, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.T.; Miller, L.M.; Rao, A.A.; D’Esposito, M. Functional Connectivity of Cortical Networks Involved in Bimanual Motor Sequence Learning. Cereb. Cortex 2007, 17, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Leone, A.; Nguyet, D.; Cohen, L.G.; Brasil-Neto, J.P.; Cammarota, A.; Hallett, M. Modulation of muscle responses evoked by transcranial magnetic stimulation during the acquisition of new fine motor skills. J. Neurophysiol. 1995, 74, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Nitsche, M.A.; Ren, X.; Wang, D.; Wang, L. Top-down and bottom-up stimulation techniques combined with action observation treatment in stroke rehabilitation: A perspective. Front. Neurol. 2023, 14, 1156987. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Source Region |
|---|---|
| LF | Frontal, left hemisphere |
| RF | Frontal, right hemisphere |
| LC | Central, left hemisphere |
| RC | Central, right hemisphere |
| LT | Temporal, left hemisphere |
| RT | Temporal, right hemisphere |
| LP | Parietal, left hemisphere |
| RP | Parietal, right hemisphere |
| LO | Occipital, left hemisphere |
| RO | Occipital, right hemisphere |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McLeod, J.; Chavan, A.; Lee, H.; Sattari, S.; Kurry, S.; Wake, M.; Janmohamed, Z.; Hodges, N.J.; Virji-Babul, N. Distinct Effects of Brain Activation Using tDCS and Observational Practice: Implications for Motor Rehabilitation. Brain Sci. 2024, 14, 175. https://doi.org/10.3390/brainsci14020175
McLeod J, Chavan A, Lee H, Sattari S, Kurry S, Wake M, Janmohamed Z, Hodges NJ, Virji-Babul N. Distinct Effects of Brain Activation Using tDCS and Observational Practice: Implications for Motor Rehabilitation. Brain Sciences. 2024; 14(2):175. https://doi.org/10.3390/brainsci14020175
Chicago/Turabian StyleMcLeod, Julianne, Anuj Chavan, Harvey Lee, Sahar Sattari, Simrut Kurry, Miku Wake, Zia Janmohamed, Nicola Jane Hodges, and Naznin Virji-Babul. 2024. "Distinct Effects of Brain Activation Using tDCS and Observational Practice: Implications for Motor Rehabilitation" Brain Sciences 14, no. 2: 175. https://doi.org/10.3390/brainsci14020175