

Changes in Multiparametric Magnetic Resonance Imaging and Plasma Amyloid-Beta Protein in Subjective Cognitive Decline
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Study Design and Participants
2.3. Clinical Data Collection and Neuropsychological Scale Assessment
2.4. Plasma Aβ42 and Aβ40 Detection
2.5. APOE Genotyping
2.6. MRI Scanning and Processing
- (1)
- Structural MRI scanning: T1-weighted 3D-SPGR sequence imaging. TR = 9.5 ms, TE = 3.9 ms, TI = 450 ms, flip angle = 20°, and matrix size = 512 × 512.
- (2)
- rs-fMRI scanning: SE-EPI sequence imaging. TR = 2 ms, TE = 30 ms, FOV = 240 × 240 mm, flip angle = 90°, matrix size = 64 × 64, slice thickness = 4 mm, and slice gap = 0.6 mm.
2.7. Preprocessing of fMRI Data and the Network Topology Feature
2.8. Preprocessing of Structural MRI Data
2.9. Statistical Analyses
3. Results
3.1. Comparison of General Clinical Data
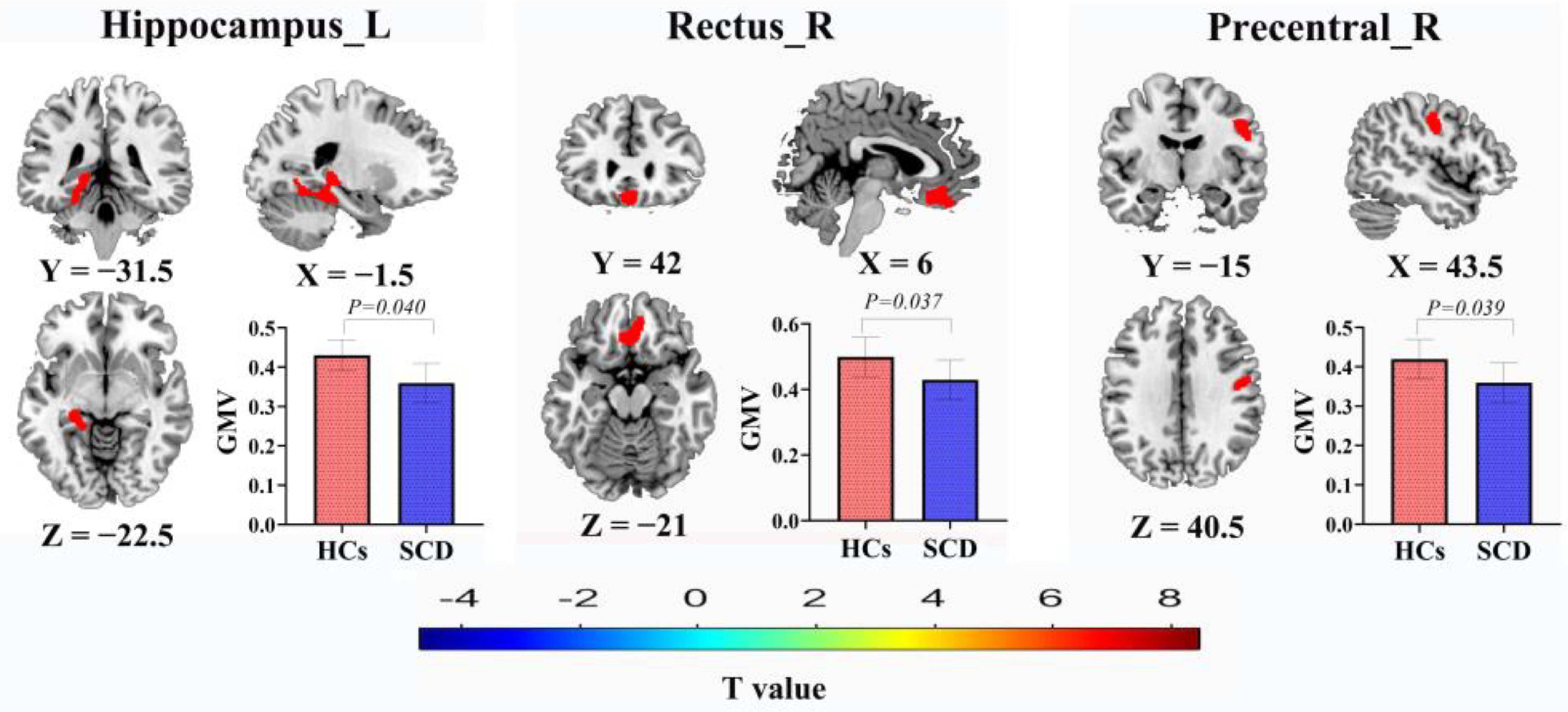
3.2. GMV Results
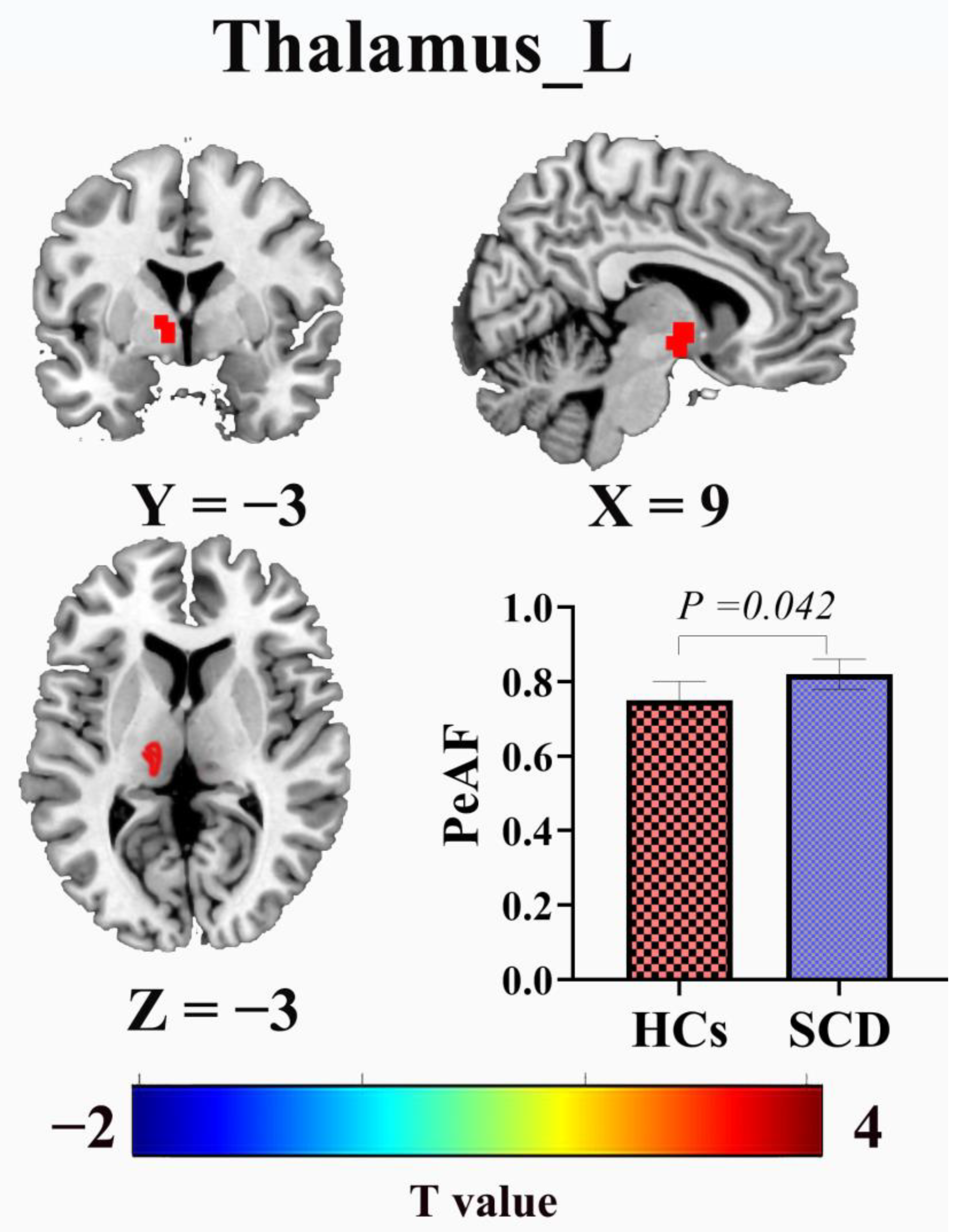
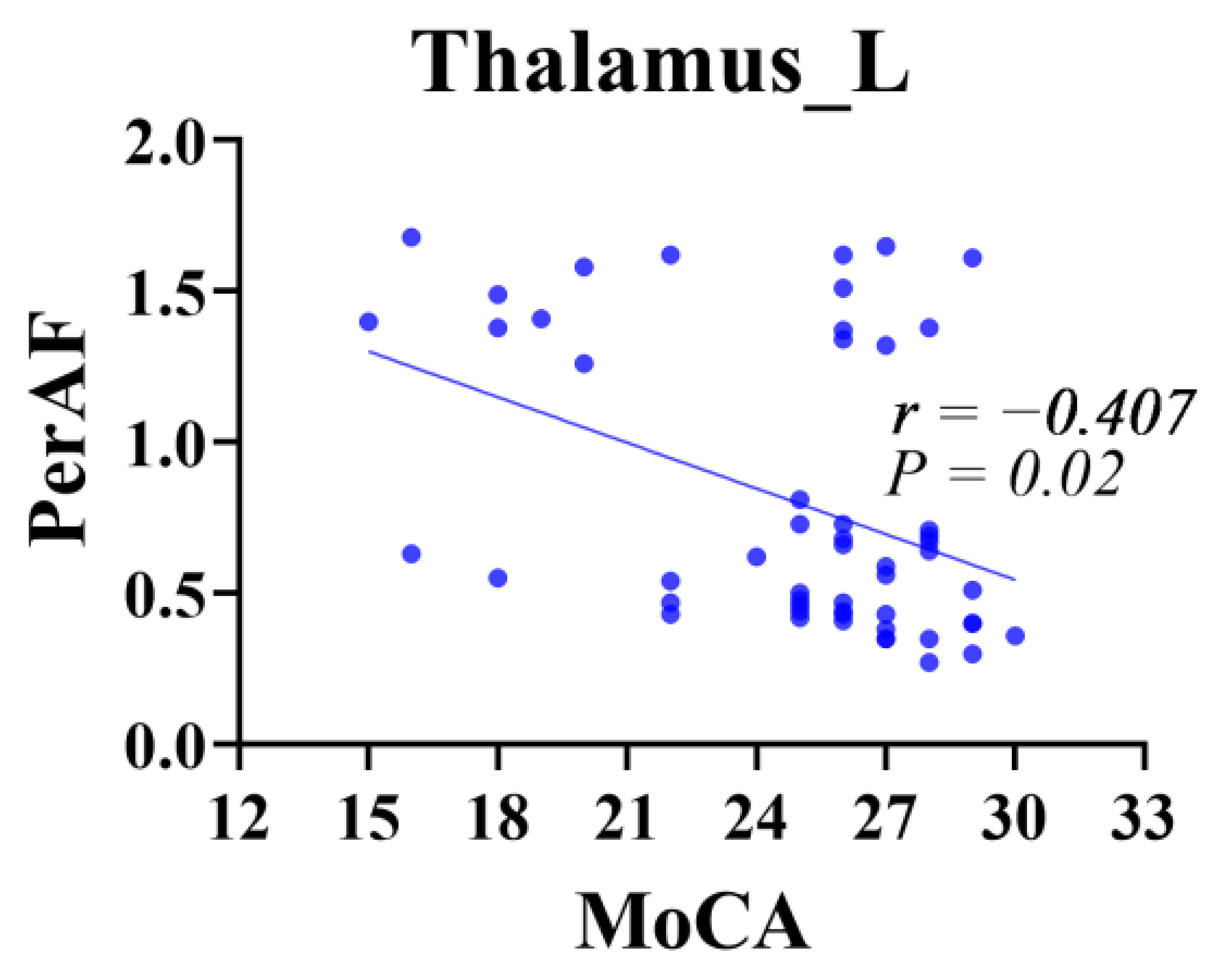
3.3. PerAF Comparison Results
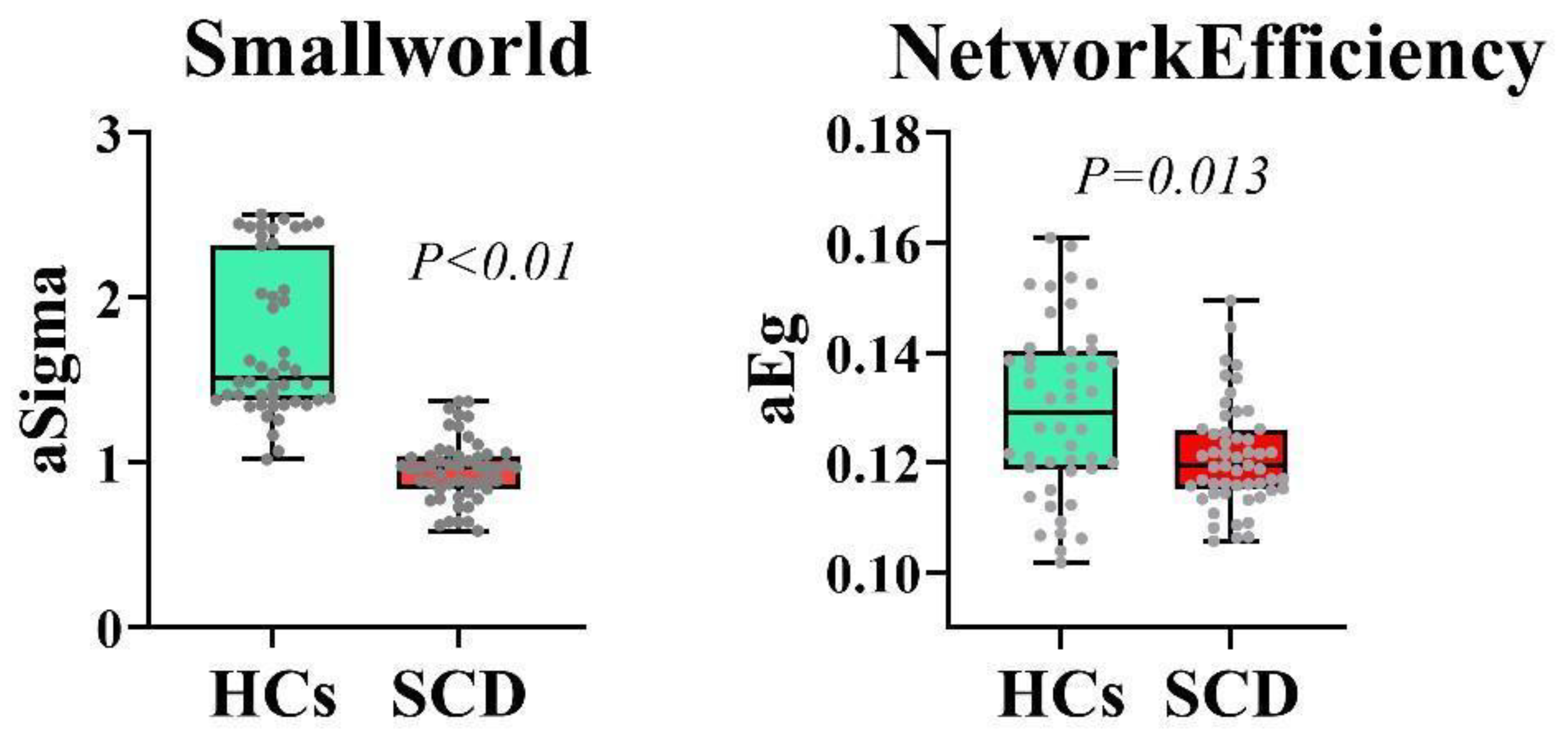
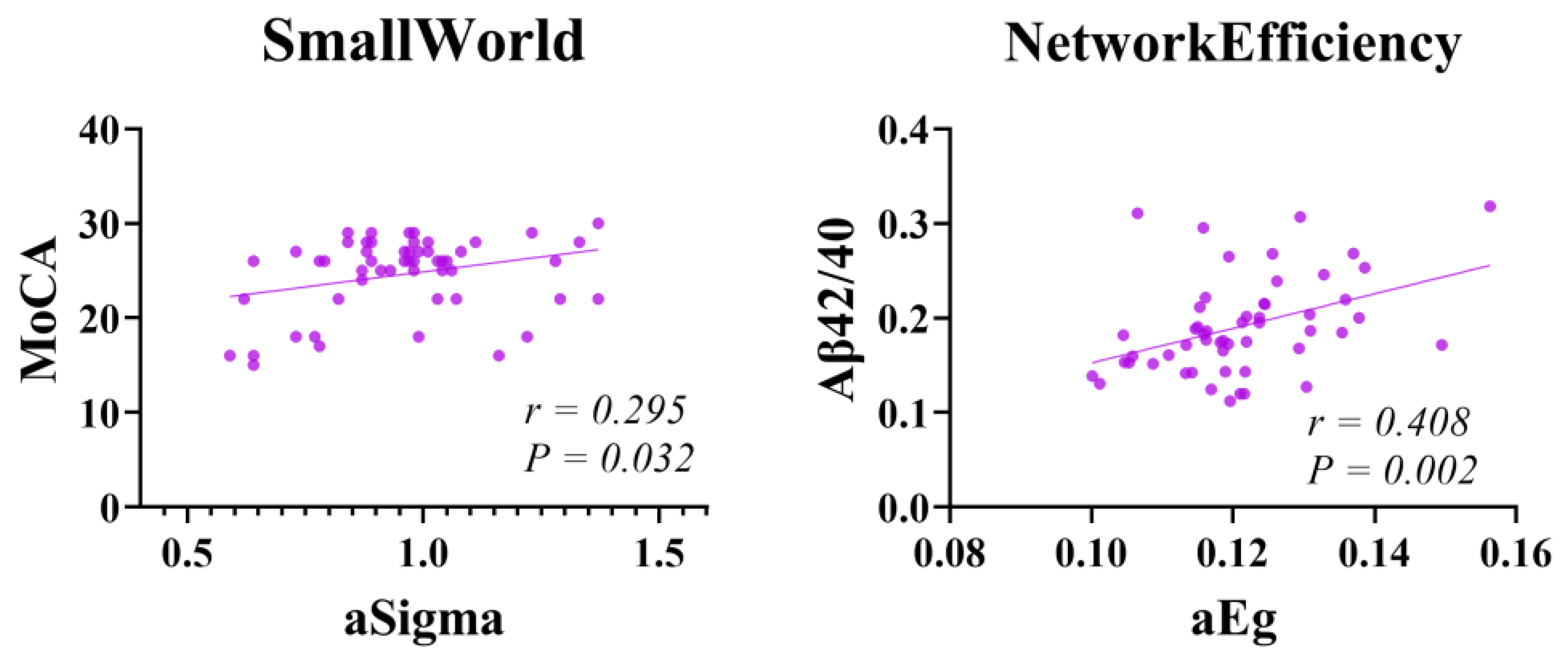
3.4. Graph Theory Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slot, R.E.R.; Sikkes, S.A.M.; Berkhof, J.; Brodaty, H.; Buckley, R.; Cavedo, E.; Dardiotis, E.; Guillo-Benarous, F.; Hampel, H.; Kochan, N.A.; et al. Subjective cognitive decline and rates of incident Alzheimer’s disease and non-Alzheimer’s disease dementia. Alzheimer’s Dement. 2019, 15, 465–476. [Google Scholar] [CrossRef]
- Pike, K.E.; Cavuoto, M.G.; Li, L.; Wright, B.J.; Kinsella, G.J. Subjective cognitive decline: Level of risk for future dementia and mild cognitive impairment, a meta-analysis of longitudinal studies. Neuropsychol. Rev. 2022, 32, 703–735. [Google Scholar] [CrossRef]
- Janssen, O.; Jansen, W.J.; Vos, S.J.B.; Boada, M.; Parnetti, L.; Gabryelewicz, T.; Fladby, T.; Molinuevo, J.L.; Villeneuve, S.; Hort, J.; et al. Characteristics of subjective cognitive decline associated with amyloid positivity. Alzheimer’s Dement. 2022, 18, 1832–1845. [Google Scholar] [CrossRef] [PubMed]
- Rabin, L.A.; Smart, C.M.; Amariglio, R.E. Subjective cognitive decline in preclinical Alzheimer’s disease. Annu. Rev. Clin. Psychol. 2017, 13, 369–396. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.S.; DeKosky, S.T.; Dickson, D.; Dubois, B.; Feldman, H.H.; Fox, N.C.; Gamst, A.; Holtzman, D.M.; Jagust, W.J.; Petersen, R.C.; et al. The diagnosis of mild cognitive impairment due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimer’s Dement. 2011, 7, 270–279. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Jessen, F.; Amariglio, R.E.; van Boxtel, M.; Breteler, M.; Ceccaldi, M.; Chételat, G.; Dubois, B.; Dufouil, C.; Ellis, K.A.; van der Flier, W.M.; et al. A conceptual framework for research on subjective cognitive decline in preclinical Alzheimer’s disease. Alzheimer’s Dement. 2014, 10, 844–852. [Google Scholar] [CrossRef]
- Jessen, F.; Amariglio, R.E.; Buckley, R.F.; van der Flier, W.M.; Han, Y.; Molinuevo, J.L.; Rabin, L.; Rentz, D.M.; Rodriguez-Gomez, O.; Saykin, A.J.; et al. The characterisation of subjective cognitive decline. Lancet Neurol. 2020, 19, 271–278. [Google Scholar] [CrossRef]
- Fan, L.Y.; Lai, Y.M.; Chen, T.F.; Hsu, Y.C.; Chen, P.Y.; Huang, K.Z.; Cheng, T.W.; Tseng, W.I.; Hua, M.S.; Chen, Y.F.; et al. Diminution of context association memory structure in subjects with subjective cognitive decline. Hum. Brain Mapp. 2018, 39, 2549–2562. [Google Scholar] [CrossRef]
- Ohlhauser, L.; Parker, A.F.; Smart, C.M.; Gawryluk, J.R.; Alzheimer’s Disease Neuroimaging, Initiative. White matter and its relationship with cognition in subjective cognitive decline. Alzheimer’s Dement. 2019, 11, 28–35. [Google Scholar] [CrossRef]
- Viviano, R.P.; Hayes, J.M.; Pruitt, P.J.; Fernandez, Z.J.; van Rooden, S.; van der Grond, J.; Rombouts, S.A.R.B.; Damoiseaux, J.S. Aberrant memory system connectivity and working memory performance in subjective cognitive decline. Neuroimage 2019, 185, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, K.; Kitamura, S.; Taoka, T.; Yasuno, F.; Tanimura, M.; Matsuoka, K.; Ikawa, D.; Toritsuka, M.; Hashimoto, K.; Makinodan, M.; et al. Gray and white matter changes in subjective cognitive impairment, amnestic mild cognitive impairment and Alzheimer’s disease: A voxel-based analysis study. PLoS ONE 2014, 9, e104007. [Google Scholar] [CrossRef]
- Rogne, S.; Vangberg, T.; Eldevik, P.; Wikran, G.; Mathiesen, E.B.; Schirmer, H. Magnetic resonance volumetry: Prediction of subjective memory complaints and mild cognitive impairment, and associations with genetic and cardiovascular risk factors. Dement. Geriatr. Cogn. Dis. Extra 2016, 6, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Striepens, N.; Scheef, L.; Wind, A.; Popp, J.; Spottke, A.; Cooper-Mahkorn, D.; Suliman, H.; Wagner, M.; Schild, H.H.; Jessen, F. Volume loss of the medial temporal lobe structures in subjective memory impairment. Dement. Geriatr. Cogn. Disord. 2010, 29, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Wang, T.; Wang, J.; Li, G.; Wang, J.; Li, X.; Li, W.; Hu, M.; Xiao, S. Asymmetry of hippocampus and amygdala defect in subjective cognitive decline among the community dwelling Chinese. Front. Psychiatry 2018, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Pennanen, C.; Kivipelto, M.; Tuomainen, S.; Hartikainen, P.; Hänninen, T.; Laakso, M.P.; Hallikainen, M.; Vanhanen, M.; Nissinen, A.; Helkala, E.L.; et al. Hippocampus and entorhinal cortex in mild cognitive impairment and early AD. Neurobiol. Aging 2004, 25, 303–310. [Google Scholar] [CrossRef]
- Teipel, S.J.; Pruessner, J.C.; Faltraco, F.; Born, C.; Rocha-Unold, M.; Evans, A.; Möller, H.J.; Hampel, H. Comprehensive dissection of the medial temporal lobe in AD: Measurement of hippocampus, amygdala, entorhinal, perirhinal and parahippocampal cortices using MRI. J. Neurol. 2006, 253, 794–800. [Google Scholar] [CrossRef]
- Ryu, S.Y.; Lim, E.Y.; Na, S.; Shim, Y.S.; Cho, J.H.; Yoon, B.; Hong, Y.J.; Yang, D.W. Hippocampal and entorhinal structures in subjective memory impairment: A combined MRI volumetric and DTI study. Int. Psychogeriatr. 2017, 29, 785–792. [Google Scholar] [CrossRef]
- Sun, Y.; Dai, Z.; Li, Y.; Sheng, C.; Li, H.; Wang, X.; Chen, X.; He, Y.; Han, Y. Subjective cognitive decline: Mapping functional and structural brain changes-A combined resting-state functional and structural MR imaging study. Radiology 2016, 281, 185–192. [Google Scholar] [CrossRef]
- Cantero, J.L.; Atienza, M.; Sanchez-Juan, P.; Rodriguez-Rodriguez, E.; Vazquez-Higuera, J.L.; Pozueta, A.; Gonzalez-Suarez, A.; Vilaplana, E.; Pegueroles, J.; Montal, V.; et al. Cerebral changes and disrupted gray matter cortical networks in asymptomatic older adults at risk for Alzheimer’s disease. Neurobiol. Aging 2018, 64, 58–67. [Google Scholar] [CrossRef]
- Greicius, M.D.; Srivastava, G.; Reiss, A.L.; Menon, V. Default-mode network activity distinguishes Alzheimer’s disease from healthy aging: Evidence from functional MRI. Proc. Natl. Acad. Sci. USA 2004, 101, 4637–4642. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef]
- Verfaillie, S.C.J.; Pichet Binette, A.; Vachon-Presseau, E.; Tabrizi, S.; Savard, M.; Bellec, P.; Ossenkoppele, R.; Scheltens, P.; van der Flier, W.M.; Breitner, J.C.S.; et al. Subjective cognitive decline is associated with altered default mode network connectivity in individuals with a family history of Alzheimer’s disease. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Dillen, K.N.H.; Jacobs, H.I.L.; Kukolja, J.; Richter, N.; von Reutern, B.; Onur, Ö.A.; Langen, K.J.; Fink, G.R. Functional disintegration of the default mode network in prodromal Alzheimer’s disease. J. Alzheimer’s Dis. 2017, 59, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yan, Y.; Wang, Y.; Hu, X.; Lu, J.; Chan, P.; Yan, T.; Han, Y. Gradual disturbances of the amplitude of low-frequency fluctuations (ALFF) and fractional ALFF in Alzheimer spectrum. Front. Neurosci. 2018, 12, 975. [Google Scholar] [CrossRef]
- Xu, K.; Wei, Y.; Zhang, S.; Zhao, L.; Geng, B.; Mai, W.; Li, P.; Liang, L.; Chen, D.; Zeng, X.; et al. Percentage amplitude of fluctuation and structural covariance changes of subjective cognitive decline in patients: A multimodal imaging study. Front. Neurosci. 2022, 16, 888174. [Google Scholar] [CrossRef]
- Jia, X.Z.; Sun, J.W.; Ji, G.J.; Liao, W.; Lv, Y.T.; Wang, J.; Wang, Z.; Zhang, H.; Liu, D.Q.; Zang, Y.F. Percent amplitude of fluctuation: A simple measure for resting-state fMRI signal at single voxel level. PLoS ONE 2020, 15, e0227021. [Google Scholar] [CrossRef]
- De Rojas, I.; Romero, J.; Rodríguez-Gomez, O.; Pesini, P.; Sanabria, A.; Pérez-Cordon, A.; Abdelnour, C.; Hernández, I.; Rosende-Roca, M.; Mauleón, A.; et al. Correlations between plasma and PET beta-amyloid levels in individuals with subjective cognitive decline: The Fundació ACE Healthy Brain Initiative (FACEHBI). Alzheimer’s Res. Ther. 2018, 10, 119. [Google Scholar] [CrossRef]
- Youn, Y.C.; Kang, S.; Suh, J.; Park, Y.H.; Kang, M.J.; Pyun, J.M.; Choi, S.H.; Jeong, J.H.; Park, K.W.; Lee, H.W.; et al. Blood amyloid-β oligomerization associated with neurodegeneration of Alzheimer’s disease. Alzheimer’s Res. Ther. 2019, 11, 40. [Google Scholar] [CrossRef]
- Lindeboom, J.; Ter Horst, R.; Hooyer, C.; Dinkgreve, M.; Jonker, C. Some psychometric properties of the CAMCOG. Psychol. Med. 1993, 23, 213–219. [Google Scholar] [CrossRef]
- Maslov, S.; Sneppen, K. Specificity and stability in topology of protein networks. Science 2002, 296, 910–913. [Google Scholar] [CrossRef]
- Chandra, A.; Valkimadi, P.E.; Pagano, G.; Cousins, O.; Dervenoulas, G.; Politis, M.; Alzheimer’s Disease Neuroimaging Initiative. Applications of amyloid, tau, and neuroinflammation PET imaging to Alzheimer’s disease and mild cognitive impairment. Hum. Brain Mapp. 2019, 40, 5424–5442. [Google Scholar] [CrossRef]
- Andreasson, U.; Blennow, K.; Zetterberg, H. Update on ultrasensitive technologies to facilitate research on blood biomarkers for central nervous system disorders. Alzheimer’s Dement. 2016, 3, 98–102. [Google Scholar] [CrossRef]
- Ovod, V.; Ramsey, K.N.; Mawuenyega, K.G.; Bollinger, J.G.; Hicks, T.; Schneider, T.; Sullivan, M.; Paumier, K.; Holtzman, D.M.; Morris, J.C.; et al. Amyloid β concentrations and stable isotope labeling kinetics of human plasma specific to central nervous system amyloidosis. Alzheimer’s Dement. 2017, 13, 841–849. [Google Scholar] [CrossRef]
- Nakamura, A.; Kaneko, N.; Villemagne, V.L.; Kato, T.; Doecke, J.; Doré, V.; Fowler, C.; Li, Q.X.; Martins, R.; Rowe, C.; et al. High performance plasma amyloid-β biomarkers for Alzheimer’s disease. Nature 2018, 554, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Schindler, S.E.; Bollinger, J.G.; Ovod, V.; Mawuenyega, K.G.; Li, Y.; Gordon, B.A.; Holtzman, D.M.; Morris, J.C.; Benzinger, T.L.S.; Xiong, C.; et al. High-precision plasma β-amyloid 42/40 predicts current and future brain amyloidosis. Neurology 2019, 93, e1647–e1659. [Google Scholar] [CrossRef]
- Schindler, S.E.; Gray, J.D.; Gordon, B.A.; Xiong, C.; Batrla-Utermann, R.; Quan, M.; Wahl, S.; Benzinger, T.L.S.; Holtzman, D.M.; Morris, J.C.; et al. Cerebrospinal fluid biomarkers measured by Elecsys assays compared to amyloid imaging. Alzheimer’s Dement. 2018, 14, 1460–1469. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, S.; Zhang, Y.; Guan, Y.; Wang, X.; Zhao, L.; Shi, Z.; Yue, W.; Zhang, Y.; Liu, S.; et al. Apolipoprotein E e4 allele is associated with subjective cognitive decline: A meta-analysis. Neuroepidemiology 2017, 49, 165–173. [Google Scholar] [CrossRef]
- Pavel, N.A.; Paun, M.R.; Matei, P.V.; Dutu, I.; Tudose, C. APOE4 status and cognitive function in middle-aged and elderly people. Balkan J. Med. Genet. 2022, 25, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Grau, S.; Rodríguez-Gómez, O.; Sanabria, Á.; Pérez-Cordón, A.; Sánchez-Ruiz, D.; Abdelnour, C.; Valero, S.; Hernández, I.; Rosende-Roca, M.; Mauleón, A.; et al. Exploring APOE genotype effects on Alzheimer’s disease risk and amyloid β burden in individuals with subjective cognitive decline: The FundacioACE Healthy Brain Initiative (FACEHBI) study baseline results. Alzheimer’s Dement. 2018, 14, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, X.; Wang, Y.; Dong, H.; Lu, J.; Scheininger, T.; Ewers, M.; Jessen, F.; Zuo, X.N.; Han, Y. Anxiety correlates with cortical surface area in subjective cognitive decline: APOE ε4 carriers versus APOE ε4 non-carriers. Alzheimer’s Res. Ther. 2019, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Tan, L.; Bi, Y.L.; Xu, W.; Tan, L.; Shen, X.N.; Hou, X.H.; Ma, Y.H.; Dong, Q.; Yu, J.T. Association between methylation of BIN1 promoter in peripheral blood and preclinical Alzheimer’s disease. Transl. Psychiatry 2021, 11, 89. [Google Scholar] [CrossRef] [PubMed]
- Soto-Rojas, L.O.; Pacheco-Herrero, M.; Martínez-Gómez, P.A.; Campa-Córdoba, B.B.; Apátiga-Pérez, R.; Villegas-Rojas, M.M.; Harrington, C.R.; de la Cruz, F.; Garcés-Ramírez, L.; Luna-Muñoz, J. The neurovascular unit dysfunction in Alzheimer’s disease. Int. J. Mol. Sci. 2021, 22, 2022. [Google Scholar] [CrossRef]
- Attems, J. Sporadic cerebral amyloid angiopathy: Pathology, clinical implications, and possible pathomechanisms. Acta Neuropathol. 2005, 110, 345–359. [Google Scholar] [CrossRef]
- Botella Lucena, P.; Vanherle, S.; Lodder, C.; Gutiérrez de Ravé, M.; Stancu, I.C.; Lambrichts, I.; Vangheluwe, R.; Bruffaerts, R.; Dewachter, I. Blood-based Aβ42 increases in the earliest pre-pathological stage before decreasing with progressive amyloid pathology in preclinical models and human subjects: Opening new avenues for prevention. Acta Neuropathol. 2022, 144, 489–508. [Google Scholar] [CrossRef]
- Bayram, E.; Caldwell, J.Z.K.; Banks, S.J. Current understanding of magnetic resonance imaging biomarkers and memory in Alzheimer’s disease. Alzheimer’s Dement. 2018, 4, 395–413. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.; Scheef, L.; Abdulkadir, A.; Boecker, H.; Heneka, M.; Wagner, M.; Koppara, A.; Klöppel, S.; Jessen, F.; Alzheimer’s Disease Neuroimaging Initiative. Gray matter atrophy pattern in elderly with subjective memory impairment. Alzheimer’s Dement. 2014, 10, 99–108. [Google Scholar] [CrossRef]
- Huang, W.; Li, X.; Li, X.; Kang, G.; Han, Y.; Shu, N. Combined support vector machine classifier and brain structural network features for the individual classification of amnestic mild cognitive impairment and subjective cognitive decline patients. Front. Aging Neurosci. 2021, 13, 687927. [Google Scholar] [CrossRef]
- Evangelista, N.D.; O’Shea, A.; Kraft, J.N.; Hausman, H.K.; Boutzoukas, E.M.; Nissim, N.R.; Albizu, A.; Hardcastle, C.; Van Etten, E.J.; Bharadwaj, P.K.; et al. Independent contributions of dorsolateral prefrontal structure and function to working memory in healthy older adults. Cereb Cortex 2021, 31, 1732–1743. [Google Scholar] [CrossRef]
- Henri-Bhargava, A.; Stuss, D.T.; Freedman, M. Clinical assessment of prefrontal lobe functions. Continuum 2018, 24, 704–726. [Google Scholar] [CrossRef]
- Buchy, L.; Barbato, M.; MacMaster, F.P.; Bray, S.; Clark, D.; Deighton, S.; Addington, J. Cognitive insight is associated with cortical thickness in first-episode psychosis. Schizophr. Res. 2016, 172, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Romund, L.; Golde, S.; Lorenz, R.C.; Raufelder, D.; Pelz, P.; Gleich, T.; Heinz, A.; Beck, A. Neural correlates of the self-concept in adolescence-A focus on the significance of friends. Hum. Brain Mapp. 2017, 38, 987–996. [Google Scholar] [CrossRef]
- Bi, X.A.; Li, L.; Xu, R.; Xing, Z. Pathogenic factors identification of brain imaging and gene in late mild cognitive impairment. Interdiscip. Sci. 2021, 13, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Burks, J.D.; Conner, A.K.; Bonney, P.A.; Glenn, C.A.; Baker, C.M.; Boettcher, L.B.; Briggs, R.G.; O’Donoghue, D.L.; Wu, D.H.; Sughrue, M.E. Anatomy and white matter connections of the orbitofrontal gyrus. J. Neurosurg. 2018, 128, 1865–1872. [Google Scholar] [CrossRef]
- Li, W.; Lou, W.; Zhang, W.; Tong, R.K.; Jin, R.; Peng, W. Gyrus rectus asymmetry predicts trait alexithymia, cognitive empathy, and social function in neurotypical adults. Cereb Cortex 2023, 33, 1941–1954. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Benavides, G.; Grau-Rivera, O.; Suárez-Calvet, M.; Minguillon, C.; Cacciaglia, R.; Gramunt, N.; Falcon, C.; ALFA Study; Gispert, J.D.; Molinuevo, J.L. Brain and cognitive correlates of subjective cognitive decline-plus features in a population-based cohort. Alzheimer’s Res. Ther. 2018, 10, 123. [Google Scholar] [CrossRef]
- Thomas, T.; Thomas, G.; McLendon, C.; Sutton, T.; Mullan, M. Beta-amyloid-mediated vasoactivity and vascular endothelial damage. Nature 1996, 380, 168–171. [Google Scholar] [CrossRef]
- Niwa, K.; Porter, V.A.; Kazama, K.; Cornfield, D.; Carlson, G.A.; Iadecola, C. A beta-peptides enhance vasoconstriction in cerebral circulation. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H2417–H2424. [Google Scholar] [CrossRef]
- Bennett, S.A.; Pappas, B.A.; Stevens, W.D.; Davidson, C.M.; Fortin, T.; Chen, J. Cleavage of amyloid precursor protein elicited by chronic cerebral hypoperfusion. Neurobiol. Aging 2000, 21, 207–214. [Google Scholar] [CrossRef]
- Janelidze, S.; Stomrud, E.; Palmqvist, S.; Zetterberg, H.; van Westen, D.; Jeromin, A.; Song, L.; Hanlon, D.; Tan Hehir, C.A.; Baker, D.; et al. Plasma beta-amyloid in Alzheimer’s disease and vascular disease. Sci. Rep. 2016, 6, 26801. [Google Scholar] [CrossRef]
- Park, D.C.; Reuter-Lorenz, P. The adaptive brain: Aging and neurocognitive scaffolding. Annu. Rev. Psychol. 2009, 60, 173–196. [Google Scholar] [CrossRef] [PubMed]
- Viviano, R.P.; Damoiseaux, J.S. Functional neuroimaging in subjective cognitive decline: Current status and a research path forward. Alzheimer’s Res. Ther. 2020, 12, 23. [Google Scholar] [CrossRef]
- Morcom, A.M.; Henson, R.N.A. Increased prefrontal activity with aging reflects nonspecific neural responses Rather than compensation. J. Neurosci. 2018, 38, 7303–7313. [Google Scholar] [CrossRef]
- Alexander-Bloch, A.; Giedd, J.N.; Bullmore, E. Imaging structural co-variance between human brain regions. Nat. Rev. Neurosci. 2013, 14, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Humphries, M.D.; Gurney, K. Network ‘small-world-ness’: A quantitative method for determining canonical network equivalence. PLoS ONE 2008, 3, e0002051. [Google Scholar] [CrossRef]
- Ten Kate, M.; Visser, P.J.; Bakardjian, H.; Barkhof, F.; Sikkes, S.A.M.; van der Flier, W.M.; Scheltens, P.; Hampel, H.; Habert, M.O.; Dubois, B.; et al. Gray matter network disruptions and regional amyloid beta in cognitively normal adults. Front. Aging Neurosci. 2018, 10, 67. [Google Scholar] [CrossRef]
- Verfaillie, S.C.J.; Slot, R.E.R.; Dicks, E.; Prins, N.D.; Overbeek, J.M.; Teunissen, C.E.; Scheltens, P.; Barkhof, F.; van der Flier, W.M.; Tijms, B.M. A more randomly organized grey matter network is associated with deteriorating language and global cognition in individuals with subjective cognitive decline. Hum. Brain Mapp. 2018, 39, 3143–3151. [Google Scholar] [CrossRef]
- Tijms, B.M.; Ten Kate, M.; Gouw, A.A.; Borta, A.; Verfaillie, S.; Teunissen, C.E.; Scheltens, P.; Barkhof, F.; van der Flier, W.M. Gray matter networks and clinical progression in subjects with predementia Alzheimer’s disease. Neurobiol. Aging 2018, 61, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Perrotin, A.; La Joie, R.; de La Sayette, V.; Barré, L.; Mézenge, F.; Mutlu, J.; Guilloteau, D.; Egret, S.; Eustache, F.; Chételat, G. Subjective cognitive decline in cognitively normal elders from the community or from a memory clinic: Differential affective and imaging correlates. Alzheimer’s Dement. 2017, 13, 550–560. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HCs (n = 53) | SCD (n = 46) | χ2/t/U-Value | p-Value | |
|---|---|---|---|---|
| Male sex (n/%) | 31/31.1 | 36/35.9 | 0.03 a | 0.955 |
| Age (years) | 55.85 ± 6.80 | 55.83 ± 6.49 | 0.013 b | 0.990 |
| Education (years) | 8.10 ± 4.05 | 7.472 ± 3.16 | 0.283 b | 0.754 |
| Hypertension (n/%) | 6/7.0 | 9/8.0 | 0.297 a | 0.86 |
| Diabetes (n/%) | 1/0.9% | 1/1.1% | 0.010 a | 0.919 |
| Hyperlipidemia (n/%) | 3/4.2% | 6/4.8 | 0.68 a | 0.407 |
| Smoking (n/%) | 6/17.6 | 14/38.9 | 5.653 a | 0.059 |
| Drinking (n/%) | 9/26.5 | 17/47.2 | 4.350 a | 0.114 |
| MMSE (score) | 29 (27, 30) | 29 (27, 30) | −0.670 c | 0.503 |
| MoCA (score) | 25 (23, 27) | 24 (20, 26) | −1.966 c | 0.049 |
| ADL (score) | 20 (20, 20) | 20 (20, 20) | 1.504 c | 0.259 |
| CAMCOG (score) | 93 (87, 101) | 91 (84, 98) | 1.313 c | 0.294 |
| CDR (score) | 0 (0, 0) | 0 (0, 0) | −1.871 c | 0.061 |
| Aβ42 (pg/mL) | 57.50 ± 9.82 | 61.43 ± 11.13 | 2.186 b | 0.037 |
| Aβ40 (pg/mL) | 293.55 ± 54.16 | 313.25 ± 62.56 | 1.135 b | 0.205 |
| Aβ42/40 | 0.19 (0.15, 0.24) | 0.12 (0.11, 0.12) | −1.024 c | 0.306 |
| HC (n = 43) | SCD (n = 51) | χ2 | p | |
|---|---|---|---|---|
| E2 | 14/7.0 | 17/17.0 | 0.013 | 0.993 |
| E3 | 57/5 6.8 | 69/69.2 | ||
| E4 | 11/11.3 | 14/13.7 |
| HC (n = 43) | SCD (n = 51) | χ2 | p | |
|---|---|---|---|---|
| E2/E2 | 3/2.7 | 3/3.3 | 2.250 | 0.134 |
| E2/E3 | 5/5.3 | 7/6.7 | ||
| E3/E3 | 22/21.8 | 27/27.2 | ||
| E2/E4 | 3/3.1 | 4/3.9 | ||
| E3/E4 | 8/8.0 | 10/10.0 | ||
| E4/E4 | 0/0 | 0/0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Yang, J.; Cheng, F.; Ning, Z.; Xi, C.; Sun, Z. Changes in Multiparametric Magnetic Resonance Imaging and Plasma Amyloid-Beta Protein in Subjective Cognitive Decline. Brain Sci. 2023, 13, 1624. https://doi.org/10.3390/brainsci13121624
Xu Q, Yang J, Cheng F, Ning Z, Xi C, Sun Z. Changes in Multiparametric Magnetic Resonance Imaging and Plasma Amyloid-Beta Protein in Subjective Cognitive Decline. Brain Sciences. 2023; 13(12):1624. https://doi.org/10.3390/brainsci13121624
Chicago/Turabian StyleXu, Qiaoqiao, Jiajia Yang, Fang Cheng, Zhiwen Ning, Chunhua Xi, and Zhongwu Sun. 2023. "Changes in Multiparametric Magnetic Resonance Imaging and Plasma Amyloid-Beta Protein in Subjective Cognitive Decline" Brain Sciences 13, no. 12: 1624. https://doi.org/10.3390/brainsci13121624