
How Posture and Previous Sensorimotor Experience Influence Muscle Activity during Gait Imagery in Young Healthy Individuals
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Instrumentation
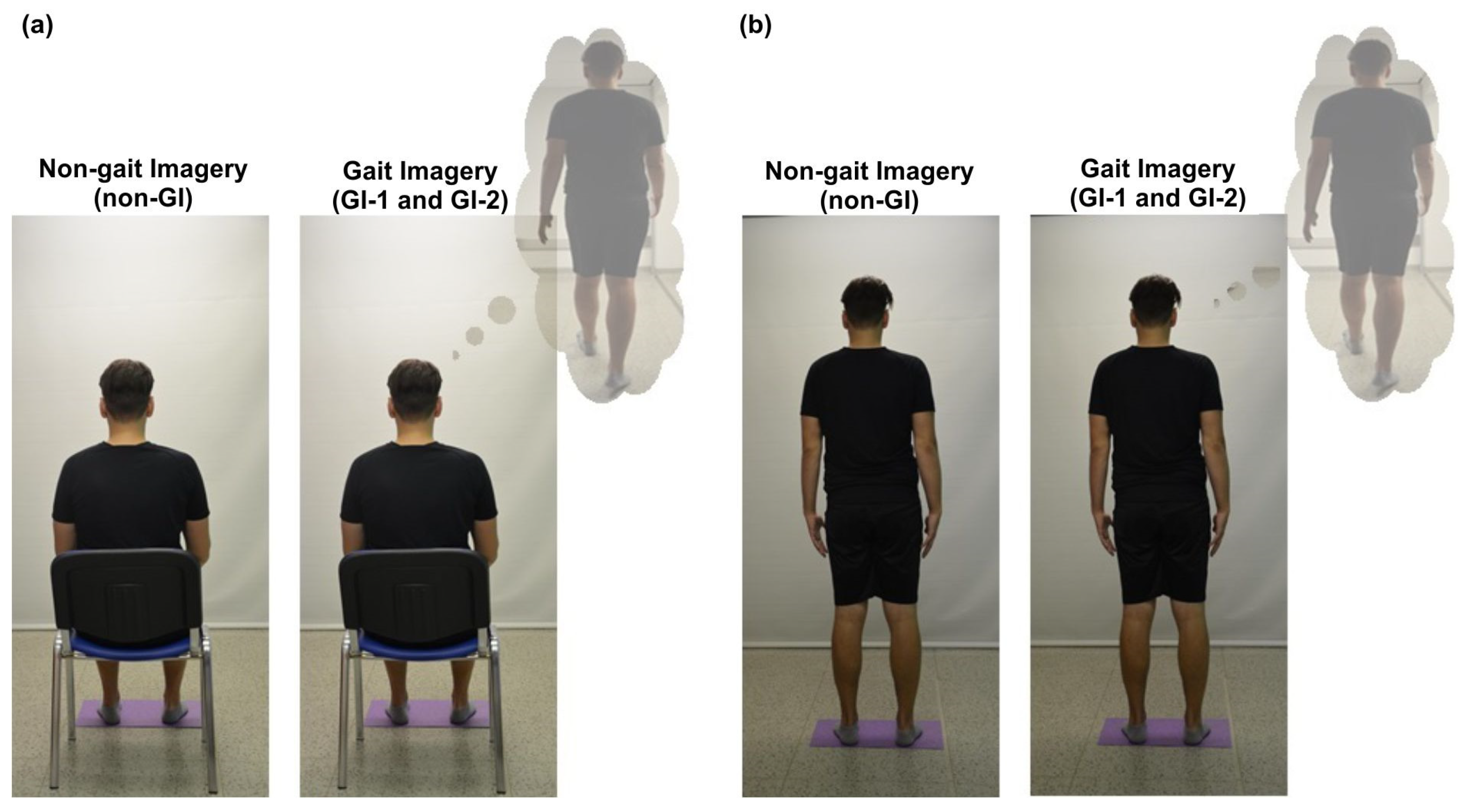
2.3. Experiment Protocol
2.4. Subjective Assessment of GI
2.5. Data Processing
2.6. Data Analysis
3. Results
4. Discussion
4.1. Electromyographic Activity during Motor Imagery
4.2. Factors That May Contribute to the Character of Electromyographic Activity When Imagining
4.2.1. Imagination of Gait
4.2.2. Default Posture during Gait Imagination
4.2.3. Previous Sensorimotor Experience with Real Walking
4.3. Study Limitations
4.4. Implications for Clinical Practice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mulder, T.H. Motor imagery and action observation: Cognitive tools for rehabilitation. J. Neural Transm. 2007, 114, 1265–1278. [Google Scholar] [CrossRef]
- Decety, J.; Grezes, J. Neural mechanisms subserving the perception of human actions. Trends Cogn. Sci. 1999, 3, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Jeannerod, M. Neural simulation of action: A unifying mechanism for motor cognition. NeuroImage 2001, 14, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M.; Fieldman, J.; Cohen, L.G.; Sadato, N.; Pascual-Leone, A. Involvement of primary motor cortex in motor imagery and mental practice. Behav. Brain Sci. 1994, 17, 210. [Google Scholar] [CrossRef]
- Nicholson, V.P.; Keogh, J.W.L.; Low Choy, N.L. Can a single session of motor imagery promote motor learning of locomotion in older adults? A randomized controlled trial. Clin. Interv. Aging 2018, 13, 713–722. [Google Scholar] [CrossRef]
- Malouin, F.; Richards, C.L.; Jackson, P.L.; Dumas, F.; Doyon, J. Brain activations during motor imagery of locomotor-related tasks: A PET study. Hum. Brain. Mapp. 2003, 19, 47–62. [Google Scholar] [CrossRef]
- Fleury, L.; Dreyer, L.; El Makkaoui, R.; Leroy, E.; Rossetti, Y.; Collet, C. Inter-Task Transfer of Prism Adaptation through Motor Imagery. Brain Sci. 2023, 13, 114. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Oh, D.-W.; Kim, S.-Y.; Choi, J.-D. Visual and kinesthetic locomotor imagery training integrated with auditory step rhythm for walking performance of patients with chronic stroke. Clin. Rehabil. 2011, 25, 134–145. [Google Scholar] [CrossRef]
- Bovonsunthonchai, S.; Aung, N.; Hiengkaew, V.; Tretriluxana, J. A randomized controlled trial of motor imagery combined with structured progressive circuit class therapy on gait in stroke survivors. Sci. Rep. 2020, 10, 6945. [Google Scholar] [CrossRef] [PubMed]
- Oostra, K.M.; Oomen, A.; Vanderstraeten, G.; Vingerhoets, G. Influence of motor imagery training on gait rehabilitation in sub-acute stroke: A randomized controlled trial. J. Rehabil. Med. 2015, 47, 204–209. [Google Scholar] [CrossRef]
- Zaparolli, L.; Sacheli, L.M.; Seghezzi, S.; Preto, M.; Stucovitz, E.; Negrini, F.; Paulesu, E. Motor imagery training speeds up gait recovery and decreases the risk of falls in patients submitted to total knee arthroplasty. Sci. Rep. 2020, 10, 8917. [Google Scholar] [CrossRef]
- Stippich, C.; Ochmann, H.; Sartor, K. Somatotopic mapping of the human primary sensorimotor cortex during motor imagery and motor execution by functional magnetic resonance imaging. Neurosci. Lett. 2002, 331, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Ehrsson, H.H.; Geyer, S.; Naito, E. Imagery of voluntary movement of fingers, toes, and tongue activates corresponding body-part-specific motor representations. J. Neurophysiol. 2003, 90, 3304–3316. [Google Scholar] [CrossRef] [PubMed]
- Hétu, S.; Grégoire, M.; Saimpont, A.; Coll, M.-P.; Eugène, F.; Michon, P.-E.; Jackson, P.L. The neural network of motor imagery: An ALE meta-analysis. Neurosci. Biobehav. Rev. 2013, 37, 930–949. [Google Scholar] [CrossRef] [PubMed]
- Vrana, A.; Hotz-Boendermaker, S.; Stämpfli, P.; Hänggi, J.; Seifritz, E.; Humphreys, B.K.; Meier, M.L. Differential Neural Processing during Motor Imagery of Daily Activities in Chronic Low Back Pain Patients. PLoS ONE 2015, 10, e0142391. [Google Scholar] [CrossRef] [PubMed]
- van der Meulen, M.; Allali, G.; Rieger, S.W.; Assal, F.; Vuilleumier, P. The influence of individual motor imagery ability on cerebral recruitment during gait imagery. Hum. Brain Mapp. 2014, 35, 455–470. [Google Scholar] [CrossRef]
- Sharma, N.; Simmons, L.H.; Jones, P.S.; Day, D.J.; Carpenter, T.A.; Pomeroy, V.M.; Baron, J.C. Motor imagery after subcortical stroke: A functional magnetic resonance imaging study. Stroke 2009, 40, 1315–1324. [Google Scholar] [CrossRef]
- Bonnet, M.; Decety, J.; Jeannerod, M.; Requin, J. Mental simulation of an action modulates the excitability of spinal reflex pathways in man. Cogn. Brain Res. 1997, 5, 221–228. [Google Scholar] [CrossRef]
- Guillot, A.; Di Rienzo, F.; MacIntyre, T.; Moran, A.; Collet, C. Imagining is Not Doing but Involves Specific Motor Commands: A Review of Experimental Data Related to Motor Inhibition. Front. Hum. Neurosci. 2012, 6, 247. [Google Scholar] [CrossRef]
- Roosink, M.; Zijdewind, I. Corticospinal excitability during observation and imagery of simple and complex hand tasks: Implications for motor rehabilitation. Behav. Brain Res. 2010, 213, 35–41. [Google Scholar] [CrossRef]
- Solodkin, A.; Hlustik, P.; Chen, E.E.; Small, S.L. Fine modulation in network activation during motor execution and motor imagery. Cereb. Cortex 2004, 14, 1246–1255. [Google Scholar] [CrossRef]
- Dietz, V.; Duysens, J. Significance of load receptor input during locomotion: A review. Gait Posture 2000, 11, 102–110. [Google Scholar] [CrossRef]
- Dietz, V. Spinal cord pattern generators for locomotion. Clin. Neurophysiol. 2003, 114, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Dietz, V. Behavior of spinal neurons deprived of supraspinal input. Nat. Rev. Neurol. 2010, 6, 167–174. [Google Scholar] [CrossRef] [PubMed]
- MacKay-Lyons, M. Central pattern generation of locomotion: A review of the evidence. Phys. Ther. 2002, 82, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Bakker, M.; de Lange, F.P.; Stevens, J.A.; Toni, I.; Bloem, B.R. Motor imagery of gait: A quantitative approach. Exp. Brain Res. 2007, 179, 497–504. [Google Scholar] [CrossRef]
- Mizuguchi, N.; Sakamoto, M.; Muraoka, T.; Moriyama, N.; Nakagawa, K.; Nakata, H.; Kanosue, K. Influence of somatosensory input on corticospinal excitability during motor imagery. Neurosci. Lett. 2012, 514, 127–130. [Google Scholar] [CrossRef]
- Mizuguchi, N.; Sakamoto, M.; Muraoka, T.; Kanosue, K. Influence of touching an object on corticospinal excitability during motor imagery. Exp. Brain Res. 2009, 196, 529–535. [Google Scholar] [CrossRef]
- Guillot, A.; Lebon, F.; Rouffet, D.; Champely, S.; Doyon, J.; Collet, C. Muscular responses during motor imagery as a function of muscle contraction types. Int. J. Psychophysiol. 2007, 66, 18–27. [Google Scholar] [CrossRef]
- Vargas, C.D.; Olivier, E.; Craighero, L.; Fadiga, L.; Duhamel, J.R.; Sirigu, A. The influence of hand posture on corticospinal excitability during motor imagery: A transcranial magnetic stimulation study. Cereb. Cortex 2004, 14, 1200–1206. [Google Scholar] [CrossRef]
- Nakazawa, K.; Kawashima, N.; Obata, H.; Yamanaka, K.; Nozaki, D.; Akai, M. Facilitation of both stretch reflex and corticospinal pathways of the tibialis anterior muscle during standing in humans. Neurosci. Lett. 2003, 338, 53–56. [Google Scholar] [CrossRef]
- Yao, W.X.; Ranganathan, V.K.; Allexandre, D.; Siemionow, V.; Yue, G.H. Kinesthetic imagery training of forceful muscle contractions increases brain signal and muscle strength. Front. Hum. Neurosci. 2013, 7, 561. [Google Scholar] [CrossRef]
- Dos Anjos, T.; Guillot, A.; Kerautret, Y.; Daligault, S.; Di Rienzo, F. Corticomotor Plasticity Underlying Priming Effects of Motor Imagery on Force Performance. Brain Sci. 2022, 12, 1537. [Google Scholar] [CrossRef]
- Kobelt, M.; Wirth, B.; Schuster-Amft, C. Muscle Activation During Grasping With and Without Motor Imagery in Healthy Volunteers and Patients After Stroke or With Parkinson’s Disease. Front. Psychol. 2018, 9, 597. [Google Scholar] [CrossRef]
- Geiger, D.E.; Behrendt, F.; Schuster-Amft, C. EMG Muscle Activation Pattern of Four Lower Extremity Muscles during Stair Climbing, Motor Imagery, and Robot-Assisted Stepping: A Cross-Sectional Study in Healthy Individuals. BioMed Res. Int. 2019, 2019, 9351689. [Google Scholar] [CrossRef]
- Lemos, T.; Souza, N.S.; Horsczaruk, C.H.; Nogueira-Campos, A.A.; de Oliveira, L.A.; Vargas, C.D.; Rodrigues, E.C. Motor imagery modulation of body sway is task-dependent and relies on imagery ability. Front. Hum. Neurosci. 2014, 8, 290. [Google Scholar] [CrossRef]
- Kolářová, B.; Krobot, A.; Polehlová, K.; Hluštík, P.; Richards, J. Effect of Gait Imagery Tasks on Lower Limb Muscle Activity With Respect to Body Posture. Percept. Mot. Skills 2016, 122, 411–431. [Google Scholar] [CrossRef]
- Bussel, B.; Roby-Brami, A.; Neris, O.R.; Yakovleff, A. Evidence for a spinal stepping generator in man: An electrophysiological study. Acta Neurobiol. Exp. 1996, 56, 465–468. [Google Scholar]
- Harkema, S.J.; Hurley, S.L.; Patel, U.K.; Requejo, P.S.; Dobkin, B.H.; Edgerton, V.R. The human lumbosacral spinal cord interprets loading during stepping. J. Neurophysiol. 1997, 77, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Takakusaki, K.; Habaguchi, T.; Ohtinata-Sugimoto, J.; Saitoh, K.; Sakamoto, T. Basal ganglia efferents to the brainstem centers controlling postural muscle tone and locomotion: A new concept for understanding motor disorders in basal ganglia dysfunction. Neuroscience 2003, 119, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Saimpont, A.; Malouin, F.; Tousignant, B.; Jackson, P.L. The influence of body configuration on motor imagery of walking in younger and older adults. Neuroscience 2012, 222, 49–57. [Google Scholar] [CrossRef]
- Shimba, S.; Kawashima, N.; Ohta, Y.; Yamamoto, S.; Nakazawa, K. Enhanced stretch reflex excitability in the soleus muscle during passive standing posture in humans. J. Electromyogr. Kinesiol. 2010, 20, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Beauchet, O.; Launay, C.; Sekhon, H. Body position and motor imagery strategy effects on imagining gait in healthy adults: Results from a cross-sectional study. PLoS ONE 2018, 13, e0191513. [Google Scholar] [CrossRef]
- Horslen, B.C.; Inglis, J.T.; Blouin, J.S.; Carpenter, M.G. Both standing and postural threat decrease Achilles’ tendon reflex inhibition from tendon electrical stimulation. J. Physiol. 2017, 595, 4493–4506. [Google Scholar] [CrossRef] [PubMed]
- McCrea, D.A. Spinal circuitry of sensorimotor control of locomotion. J. Physiol. 2001, 533, 41–50. [Google Scholar] [CrossRef]
- Mayer, W.P.; Murray, A.J.; Brenner-Morton, S.; Jessell, T.M.; Tourtellotte, W.G.; Akay, T. Role of muscle spindle feedback in regulating muscle activity strength during walking at different speeds in mice. J. Neurophysiol. 2018, 120, 2484–2497. [Google Scholar] [CrossRef] [PubMed]
- Saimpont, A.; Malouin, F.; Durand, A.; Mercier, C.; di Rienzo, F.; Saruco, E.; Collet, C.; Guillot, A.; Jackson, P.L. The effects of body position and actual execution on motor imagery of locomotor tasks in people with a lower-limb amputation. Sci. Rep. 2021, 11, 13788. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.R.; Martin, K.A. Measuring movement imagery abilities: A revision of the Movement Imagery Questionnaire. J. Ment. Imag. 1997, 21, 143–154. [Google Scholar]
- Monsma, E.; Short, S.; Hall, C.; Gregg, M.; Sullivan, P. Psychometric Properties of the Revised Movement Imagery Questionnaire (MIQ-R). J. Imag. Res. Sport Phys. Act. 2009, 4. [Google Scholar] [CrossRef]
- Stegeman, D.; Hermens, H. Standards for surface electromyography: The European project Surface EMG for non-invasive assessment of muscles (SENIAM). J. Appl. Biomech. 2007, 108–112. [Google Scholar]
- Farina, D.; Merletti, R.; Enoka, R.M. The extraction of neural strategies from the surface EMG: An update. J. Appl. Physiol. 2014, 117, 1215–1230. [Google Scholar] [CrossRef] [PubMed]
- Heckman, C.J.; Enoka, R.M. Motor units. Compr. Physiol. 2012, 2, 2629–2682. [Google Scholar] [CrossRef]
- Heckman, C.J.; Mottram, C.; Quinlan, K.; Theiss, R.; Schuster, J. Motoneuron excitability: The importance of neuromodulatory inputs. Clin. Neurophysiol. 2009, 120, 2040–2054. [Google Scholar] [CrossRef]
- Aoyama, T.; Kaneko, F.; Ohashi, Y.; Kohno, Y. Dissociation between cortical and spinal excitability of the antagonist muscle during combined motor imagery and action observation. Sci. Rep. 2019, 9, 13120. [Google Scholar] [CrossRef] [PubMed]
- Faist, M.; Hoefer, C.; Hodapp, M.; Dietz, V.; Berger, W.; Duysens, J. In humans Ib facilitation depends on locomotion while suppression of Ib inhibition requires loading. Brain Res. 2006, 1076, 87–92. [Google Scholar] [CrossRef]
- Sheik, A.A.; Korn, E.R. Imagery in Sports and Physical Performance; Baywood Publishing Company, Inc.: Amityville, NY, USA, 1994. [Google Scholar]
- Lotze, M.; Cohen, L.G. Volition and imagery in neurorehabilitation. Cogn. Behav. Neurol. 2006, 19, 135–140. [Google Scholar] [CrossRef]
- Kolářová, B.; Richards, J.; Haltmar, H.; Lippertová, K.; Connell, L.; Chohan, A. The effect of motor imagery on quality of movement when performing reaching tasks in healthy subjects: A proof of concept. J. Bodyw. Mov. Ther. 2022, 29, 161–166. [Google Scholar] [CrossRef]
- Vigotsky, A.D.; Halperin, I.; Lehman, G.J.; Trajano, G.S.; Vieira, T.M. Interpreting Signal Amplitudes in Surface Electromyography Studies in Sport and Rehabilitation Sciences. Front. Physiol. 2018, 8, 985. [Google Scholar] [CrossRef] [PubMed]
- Milton, J.; Small, S.L.; Solodkin, A. Imaging motor imagery: Methodological issues related to expertise. Methods 2008, 45, 336–341. [Google Scholar] [CrossRef]
- Gentili, R.; Papaxanthis, C.; Pozzo, T. Improvement and generalization of arm motor performance through motor imagery practice. Neuroscience 2006, 137, 761–772. [Google Scholar] [CrossRef]
- Muñoz, M.R.; González-Sánchez, M.; Cuesta-Vargas, A.I. Tibialis anterior analysis from a functional and architectural perspective during isometric foot dorsiflexion: A cross-sectional study of repeated measures. J. Foot Ankle Res. 2015, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, S.; Gravel, D.; Arsenault, A.B.; Bourbonnais, D. Plantarflexor weakness as a limiting factor of gait speed in stroke subjects and the compensating role of hip flexors. Clin. Biomech. 1999, 14, 125–135. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Non-Gait Imagery Condition (non-GI) | Gait Imagery before Walking (GI-1) | Gait Imagery after Walking (GI-2) | Friedman’s ANOVA | Wilcoxon Post hoc Test | ||||
|---|---|---|---|---|---|---|---|---|
| non-GI vs. GI-1 | non-GI vs. GI-2 | GI-1 vs. GI-2 | ||||||
| Mean (SD) | Mean (SD) | Mean (SD) | p | p | p | p | ||
| SITTING | Right TA | 5.85 (5.33) | 5.96 (5.41) | 4.67 (2.44) | 0.024 | 0.775 | 0.029 | 0.008 * |
| Left TA | 5.35 (3.29) | 5.38 (3.69) | 4.36 (1.98) | 0.017 | 0.948 | 0.004 * | 0.009 * | |
| Right GM | 4.39 (2.56) | 4.47 (3.3) | 3.98 (2.35) | 0.002 | 0.836 | 0.038 | 0.066 | |
| Left GM | 4.22 (2.57) | 4.26 (2.94) | 3.76 (1.85) | 0.048 | 0.824 | 0.124 | 0.118 | |
| STANDING | Right TA | 5.44 (3.42) | 6.06 (5.46) | 4.54 (2.87) | 0.042 | 0.966 | 0.059 | 0.01 * |
| Left TA | 5.56 (2.46) | 6.47 (7.3) | 4.73 (2.32) | 0.249 | 0.726 | 0.085 | 0.413 | |
| Right GM | 6.89 (4.1) | 7.69 (5.93) | 7.46 (4.67) | 0.663 | 0.13 | 0.431 | 0.844 | |
| Left GM | 7.71 (4.32) | 8.74 (4.96) | 7.85 (5.62) | 0.134 | 0.379 | 0.342 | 0.042 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolářová, B.; Tomsa, M.; Kolář, P.; Haltmar, H.; Diatelová, T.; Janura, M. How Posture and Previous Sensorimotor Experience Influence Muscle Activity during Gait Imagery in Young Healthy Individuals. Brain Sci. 2023, 13, 1605. https://doi.org/10.3390/brainsci13111605
Kolářová B, Tomsa M, Kolář P, Haltmar H, Diatelová T, Janura M. How Posture and Previous Sensorimotor Experience Influence Muscle Activity during Gait Imagery in Young Healthy Individuals. Brain Sciences. 2023; 13(11):1605. https://doi.org/10.3390/brainsci13111605
Chicago/Turabian StyleKolářová, Barbora, Marek Tomsa, Petr Kolář, Hana Haltmar, Tereza Diatelová, and Miroslav Janura. 2023. "How Posture and Previous Sensorimotor Experience Influence Muscle Activity during Gait Imagery in Young Healthy Individuals" Brain Sciences 13, no. 11: 1605. https://doi.org/10.3390/brainsci13111605