
The Interplay of Oxytocin and Attachment in Schizophrenic Patients: An fMRI Study

 ,
,
Abstract
:1. Introduction
1.1. Neurobiology of Attachment and Mentalizing in Psychosis: A Developmental Perspective
1.2. Hypotheses
2. Materials and Methods
2.1. Sample Characteristics
2.2. Measures
2.2.1. Attachment Measure as the Basis for the fMRI Paradigm
2.2.2. Clinical Measures
2.2.3. Oxytocin Measurement
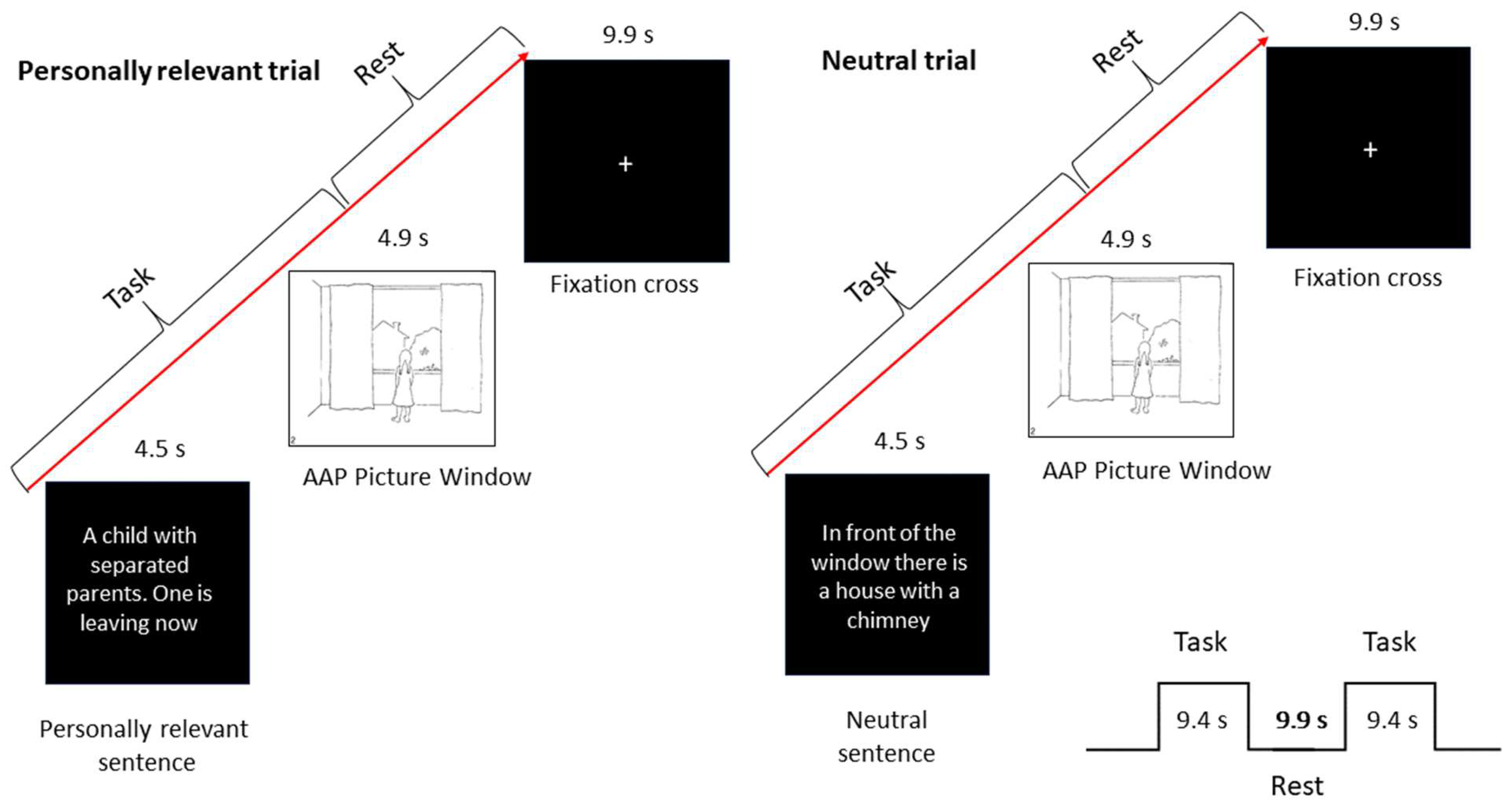
2.2.4. Experimental Design
2.2.5. Data Aquisition
2.2.6. Statistical Data Analysis
3. Results
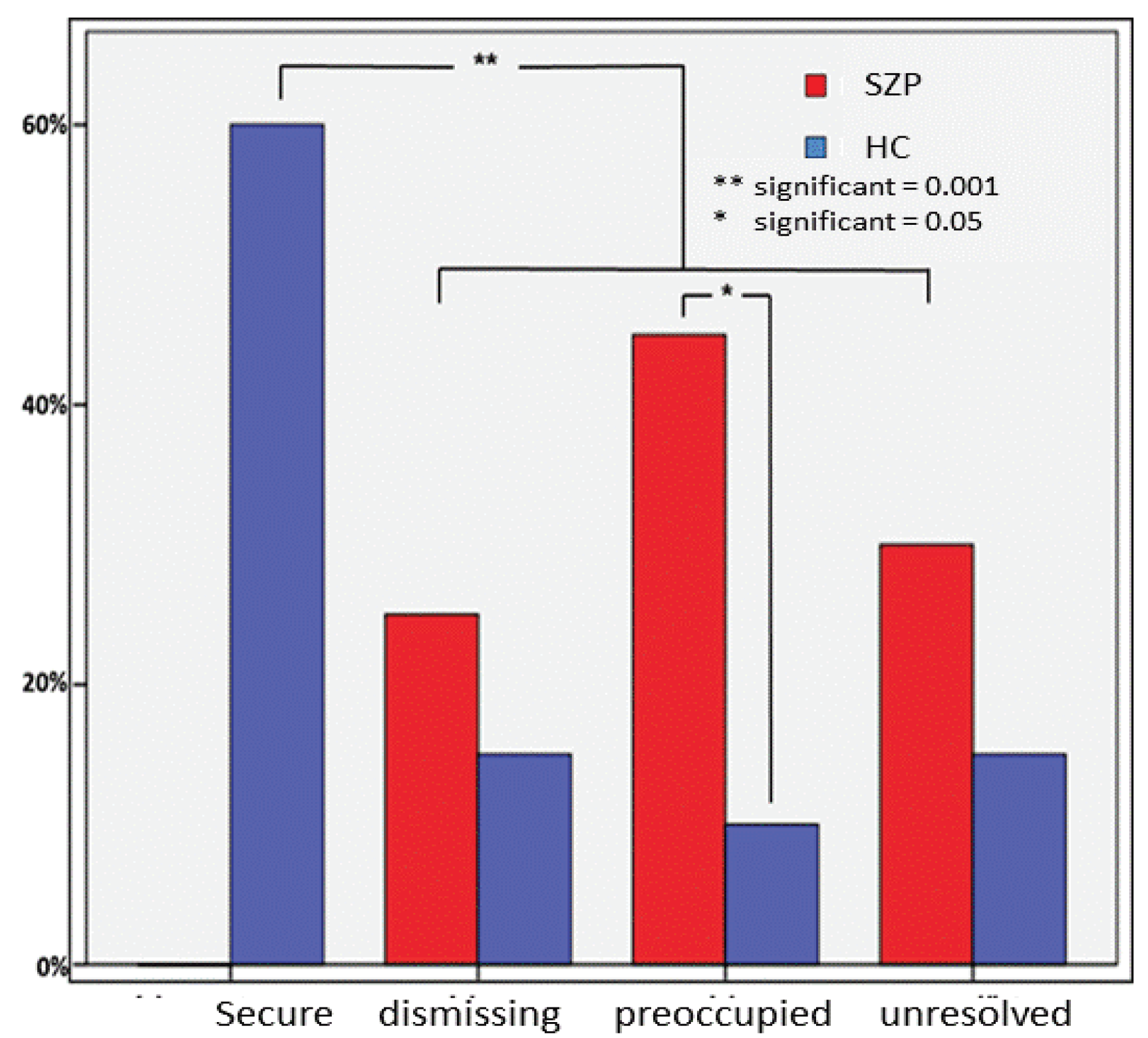
3.1. Attachment Classifications
3.2. Endocrinological Data
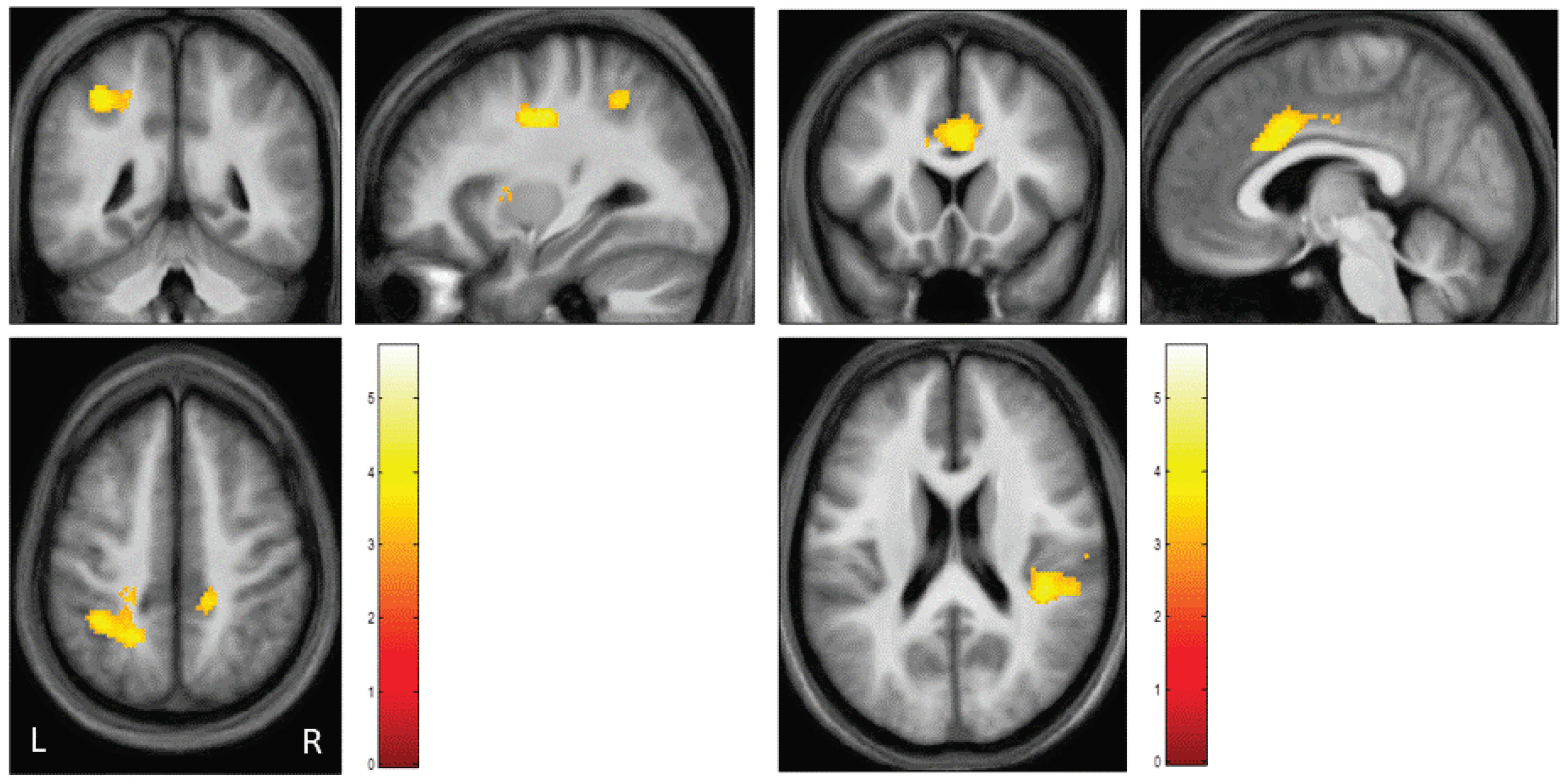
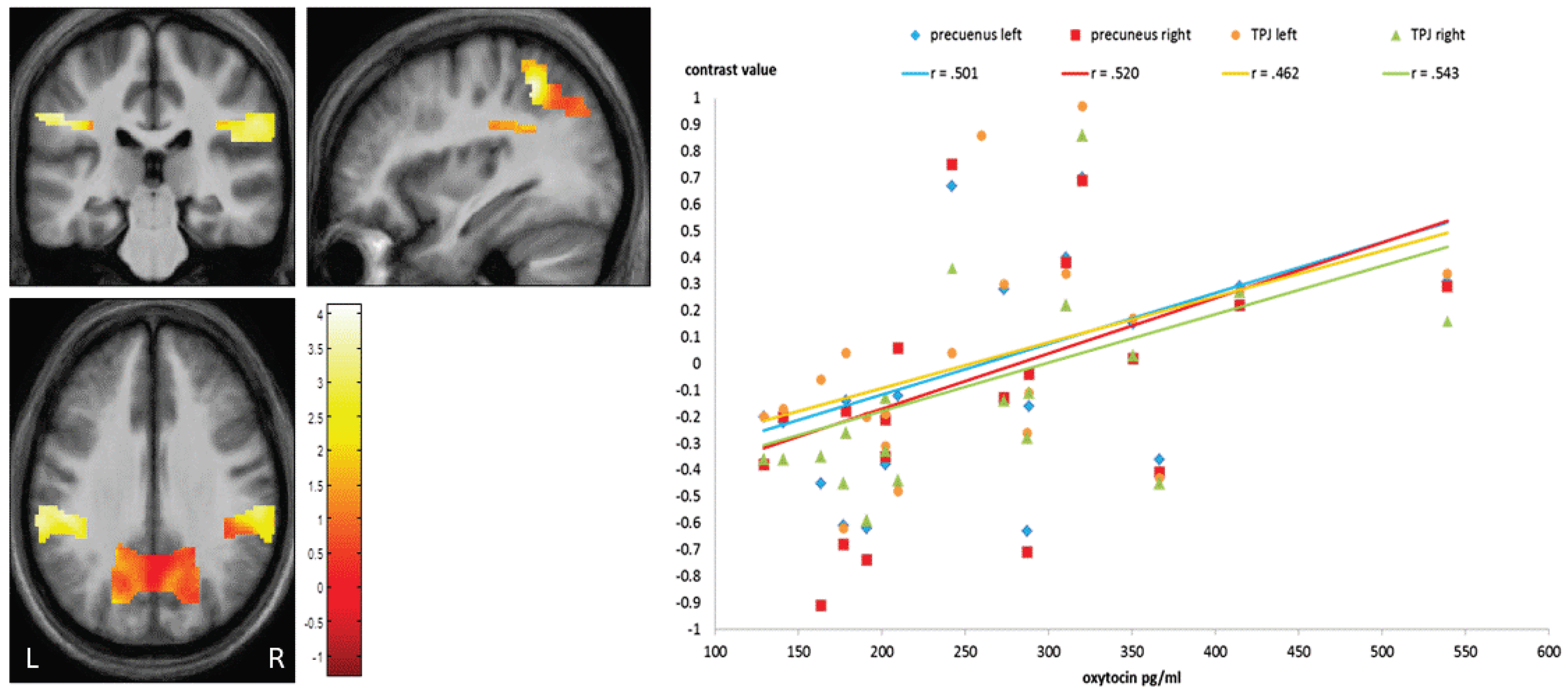
3.3. Neuroimaging Data
4. Discussion
4.1. Discussion on Oxytocin Level
4.2. Discussion on Attachment Pattern in Relation to Oxytocin
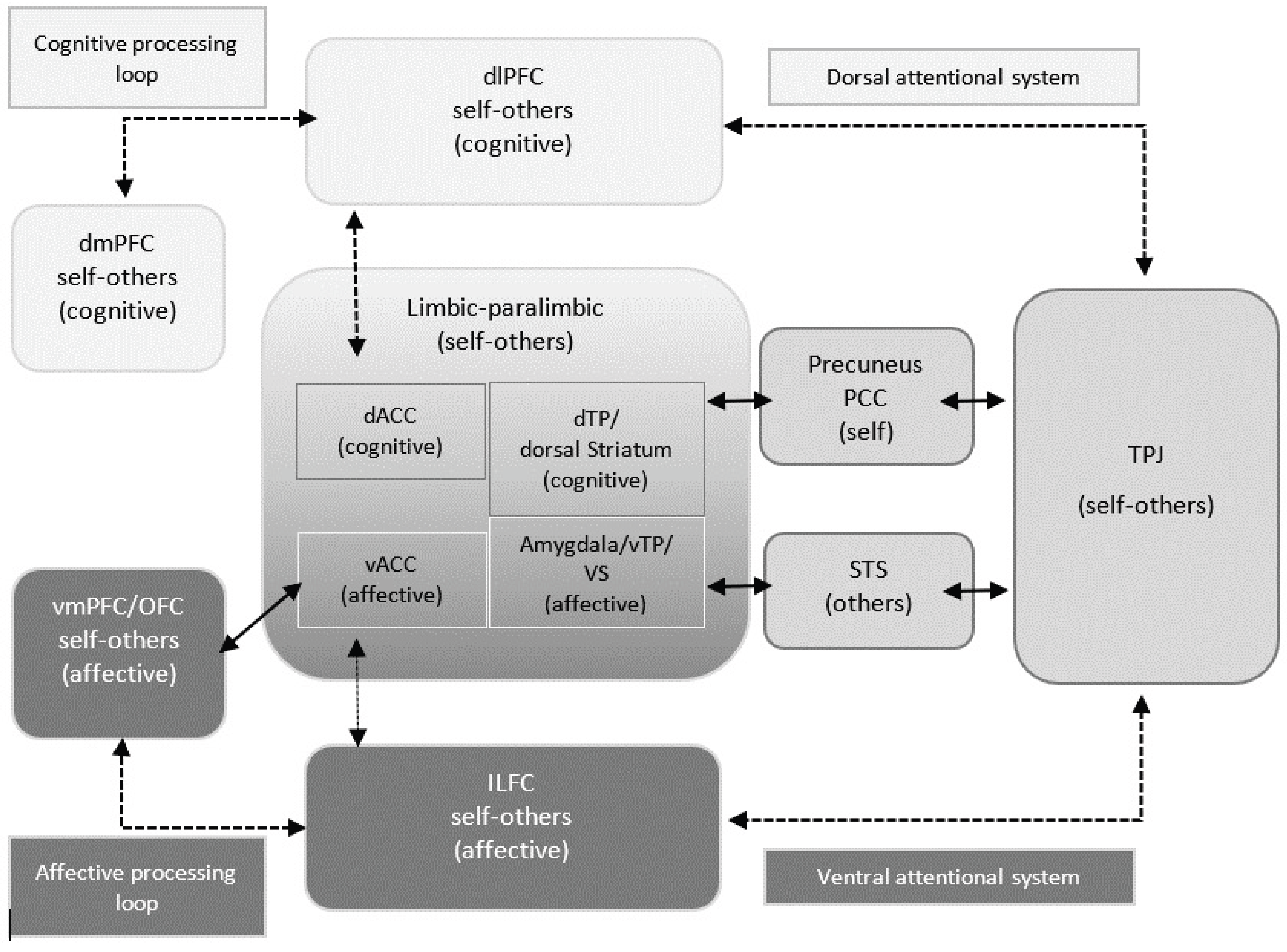
4.3. Discussion on fMRI Data
4.3.1. Stimuli-Based Discussion of Brain Activation
4.3.2. Group Comparison
4.3.3. OXT-Based Discussion of Brain Activation
4.3.4. Attachment-Based Discussion of Brain Activation
4.3.5. Psychosis-Based Discussion of Brain Activation
4.3.6. General Discussion
4.4. Limitations
- (a)
- With regard to psychotic symptomatology: The mean PANSS total score was about 60 points, therefore the patient sample can be considered psychopathological stable. According to Leucht and colleagues [93], scores between 43 and 61 correspond to a clinical global impression (CGI, [167]) of three or mildly ill ([93], p. 234). The highest value corresponded to a CGI of four, i.e., moderately ill ([93], p. 235), and the lowest to a CGI of two, respectively on the borderline of mental illness ([93], p. 234).
- (b)
- With regard to medication: The intake of atypical antipsychotic drugs might have affected OXT plasma levels. With the exception of one patient, all patients were medicated, so that, as expected, there was little positive symptomatology such as hallucinations and delusions, but a certain degree of negative symptomatology such as social and emotional withdrawal and flattening of affect. A point of criticism to be discussed in this context is the fact that one patient was not receiving neuroleptic medication at the time of data collection. This is a potential confounding variable. However, the non-medicated patient in our sample did not show any abnormal results compared to the other medicated patients with respect to his neuropsychological functioning level or the pattern of his brain activation in the two fMRI paradigms.
- (c)
- Sample characteristics: Our findings are limited by sample size, since in both groups only 20 subjects could be finally included for statistical analysis. Further studies have to be conducted to provide more evidence in larger samples and for differentiating between different subtypes of psychosis and different states of disease and separating by considering the endurance of the disease since the first episode. Our results might be gender-related since we included only male patients and cannot be transferred to female patients. The results therefore cannot be generalized to a general schizophrenic population. Nevertheless, our findings indicate that an unresolved attachment style in schizophrenia is related to less concentrated OXT and to a cluster of increased brain activation within the mentalizing neural network.
- (d)
- Since a cross-sectional design is used, behavioral and neurophysiological parameters cannot be analyzed over time. To conclude cause- and effect relationships is difficult because the data are based on a one-time measurement of both the alleged cause and effect.
- (e)
- With regard to attachment representations a limited generalizability can be assumed and has to be addressed in further studies. However, we selected one of the most widely accepted, well-validated methods of assessing individual differences in attachment representations in adolescence and adulthood.
5. Conclusions
5.1. Conclusion with Regard to Developmentals Aspects
5.2. Conclusions with Regard to Prevention and Psychotherapy
5.3. With Regard to a Key Contemporary Attachment Concept: Mentalising
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holmes, J.; Slade, A. The neuroscience of attachment: Implications for psychological therapies. Br. J. Psychiatry 2019, 214, 318–319. [Google Scholar] [CrossRef]
- Cassidy, J.; Shaver, P. Handbook of Attachment: Theory, Research, and Clinical Applications; Guilford Press: New York, NY, USA, 2016. [Google Scholar]
- Bentall, R.P.; Fernyhough, C.; Morrison, A.P.; Lewis, S.; Corcoran, R. Prospects for a cognitive-developmental account of psychotic experiences. Br. J. Clin. Psychol. 2007, 46, 155–173. [Google Scholar] [CrossRef]
- Berry, K.; Barrowclough, C.; Wearden, A. Attachment theory: A framework for understanding symptoms and interpersonal relationships in psychosis. Behav. Res. Ther. 2008, 46, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Pos, K.; Bartels-Velthuis, A.A.; Simons, C.J.; Korver-Nieberg, N.; Meijer, C.J.; de Haan, L.; GROUP. Theory of Mind and attachment styles in people with psychotic disorders, their siblings, and controls. Aust. N. Z. J. Psychiatry 2015, 49, 171–180. [Google Scholar] [CrossRef]
- Rutten, B.P.; Hammels, C.; Geschwind, N.; Menne-Lothmann, C.; Pishva, E.; Schruers, K.; van den Hove, D.; Kenis, G.; van Os, J.; Wichers, M. Resilience in mental health: Linking psychological and neurobiological perspectives. Acta Psychiatr. Scand. 2013, 128, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Bowlby, J. Attachment and Loss: Volume 1. Attachment; Basic Books: New York, NY, USA, 1969. [Google Scholar]
- Bowlby, J. A Secure Base: Parent-Child Attachment and Healthy Human Development; Basic Books: New York, NY, USA, 1988. [Google Scholar]
- Buchheim, A.; George, C.; Gündel, H. Editorial: Neuroscience of human attachment volume II. Front. Hum. Neurosci. 2022, 16, 1018528. [Google Scholar] [CrossRef]
- Buchheim, A.; George, C.; Gündel, H.; Viviani, R. Editorial: Neuroscience of Human Attachment. Front. Hum. Neurosci. 2017, 11, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.F.; Olivier, B.; Crawley, J.N.; Penn, D.L.; Silverstein, S. Social Cognition in Schizophrenia: Recommendations from the Measurement and Treatment Research to Improve Cognition in Schizophrenia New Approaches Conference. Schizophr. Bull. 2005, 31, 882–887. [Google Scholar] [CrossRef] [Green Version]
- Lavin, R.; Bucci, S.; Varese, F.; Berry, K. The relationship between insecure attachment and paranoia in psychosis: A systematic literature review. Br. J. Clin. Psychol. 2020, 59, 39–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, T.; Patrick, D. Social functioning as an outcome measure in schizophrenia studies. Acta Psychiatr. Scand. 2007, 116, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Juckel, G.; Morosini, P.L. The new approach: Psychosocial functioning as a necessary outcome criterion for therapeutic success in schizophrenia. Curr. Opin. Psychiatry 2008, 21, 630–639. [Google Scholar] [CrossRef]
- McCutcheon, R.A.; Keefe, R.S.E.; McGuire, P.K. Cognitive impairment in schizophrenia: Aetiology, pathophysiology, and treatment. Mol. Psychiatry 2023. [Google Scholar] [CrossRef]
- Green, M.F.; Bearden, C.E.; Cannon, T.D.; Fiske, A.P.; Hellemann, G.S.; Horan, W.P.; Kee, K.; Kern, R.S.; Lee, J.; Sergi, M.J.; et al. Social Cognition in Schizophrenia, Part 1: Performance Across Phase of Illness. Schizophr. Bull. 2011, 38, 854–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.F.; Horan, W.P.; Lee, J. Social cognition in schizophrenia. Nat. Rev. Neurosci. 2015, 16, 620–631. [Google Scholar] [CrossRef]
- Derntl, B.; Finkelmeyer, A.; Toygar, T.K.; Hülsmann, A.; Schneider, F.; Falkenberg, D.I.; Habel, U. Generalized deficit in all core components of empathy in schizophrenia. Schizophr. Res. 2009, 108, 197–206. [Google Scholar] [CrossRef]
- Kohler, C.G.; Walker, J.B.; Martin, E.A.; Healey, K.M.; Moberg, P.J. Facial Emotion Perception in Schizophrenia: A Meta-analytic Review. Schizophr. Bull. 2009, 36, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Achim, A.M.; Ouellet, R.; Roy, M.-A.; Jackson, P.L. Assessment of empathy in first-episode psychosis and meta-analytic comparison with previous studies in schizophrenia. Psychiatry Res. 2011, 190, 3–8. [Google Scholar] [CrossRef]
- Amminger, G.P.; Schäfer, M.R.; Klier, C.M.; Schlögelhofer, M.; Mossaheb, N.; Thompson, A.; Bechdolf, A.; Allott, K.; McGorry, P.D.; Nelson, B. Facial and vocal affect perception in people at ultra-high risk of psychosis, first-episode schizophrenia and healthy controls. Early Interv. Psychiatry 2012, 6, 450–454. [Google Scholar] [CrossRef]
- Amminger, G.P.; Schäfer, M.R.; Papageorgiou, K.; Klier, C.M.; Schlögelhofer, M.; Mossaheb, N.; Werneck-Rohrer, S.; Nelson, B.; McGorry, P.D. Emotion Recognition in Individuals at Clinical High-Risk for Schizophrenia. Schizophr. Bull. 2012, 38, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.; Papas, A.; Bartholomeusz, C.; Allott, K.; Amminger, G.P.; Nelson, B.; Wood, S.; Yung, A. Social cognition in clinical “at risk” for psychosis and first episode psychosis populations. Schizophr. Res. 2012, 141, 204–209. [Google Scholar] [CrossRef]
- Thompson, A.D.; Bartholomeusz, C.; Yung, A.R. Social cognition deficits and the ‘ultra high risk’ for psychosis population: A review of literature. Early Interv. Psychiatry 2011, 5, 192–202. [Google Scholar] [CrossRef]
- Brüne, M.; Abdel-Hamid, M.; Lehmkämper, C.; Sonntag, C. Mental state attribution, neurocognitive functioning, and psychopathology: What predicts poor social competence in schizophrenia best? Schizophr. Res. 2007, 92, 151–159. [Google Scholar] [CrossRef]
- Smith, M.J.; Horan, W.P.; Karpouzian, T.M.; Abram, S.V.; Cobia, D.J.; Csernansky, J.G. Self-reported empathy deficits are uniquely associated with poor functioning in schizophrenia. Schizophr. Res. 2012, 137, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Lysaker, P.H.; Gagen, E.; Wright, A.; Vohs, J.L.; Kukla, M.; Yanos, P.T.; Hasson-Ohayon, I. Metacognitive Deficits Predict Impaired Insight in Schizophrenia Across Symptom Profiles: A Latent Class Analysis. Schizophr. Bull. 2018, 45, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Dozier, M.; Stovall-McClough, K.C.; Albus, K.E. Attachment and Psychopathology in Adulthood. In Handbook of Attachment: Theory, Research, and Clinical Applications; Cassidy, J., Shaver, P., Eds.; The Guilford Press: New York, NY, USA, 2008; pp. 718–744. [Google Scholar]
- Debbané, M.; Salaminios, G.; Luyten, P.; Badoud, D.; Armando, M.; Solida Tozzi, A.; Fonagy, P.; Brent, B.K. Attachment, Neurobiology, and Mentalizing along the Psychosis Continuum. Front. Hum. Neurosci. 2016, 10, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, C.; Kaplan, N.; Main, M. The Adult Attachment Interview; University of California: Berkeley, CA, USA, 1985. [Google Scholar]
- George, C.; West, M. The Adult Attachment Projective: Measuring individual differences in attachment security using projective methodology. In Objective and Projective Assessment of Personality and Psychopathology; Hilsenroth, M., Segal, D., Eds.; John Wiley & Sons: New York, NY, USA, 2003; Volume 2. [Google Scholar]
- George, C.; West, M. The Adult Attachment Projective Picture System: Attachment Theory and Assessment in Adults; Guilford Press: New York, NY, USA, 2012. [Google Scholar]
- Tyrrell, C.L.; Dozier, M.; Teague, G.B.; Fallot, R.D. Effective treatment relationships for persons with serious psychiatric disorders: The importance of attachment states of mind. J. Consult. Clin. Psychol. 1999, 67, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Carnelley, K.B.; Newman-Taylor, K. How does insecure attachment lead to paranoia? A systematic critical review of cognitive, affective, and behavioural mechanisms. Br. J. Clin. Psychol. 2022, 61, 781–815. [Google Scholar] [CrossRef]
- Korver-Nieberg, N.; Berry, K.; Meijer, C.J.; de Haan, L. Adult attachment and psychotic phenomenology in clinical and non-clinical samples: A systematic review. Psychol. Psychother. 2014, 87, 127–154. [Google Scholar] [CrossRef]
- Gumley, A.I.; Taylor, H.E.F.; Schwannauer, M.; MacBeth, A. A systematic review of attachment and psychosis: Measurement, construct validity and outcomes. Acta Psychiatr. Scand. 2014, 129, 257–274. [Google Scholar] [CrossRef] [Green Version]
- Berry, K.; Roberts, N.; Danquah, A.; Davies, L. An exploratory study of associations between adult attachment, health service utilisation and health service costs. Psychosis 2014, 6, 355–358. [Google Scholar] [CrossRef]
- Ponizovsky, A.M.; Vitenberg, E.; Baumgarten-Katz, I.; Grinshpoon, A. Attachment styles and affect regulation among outpatients with schizophrenia: Relationships to symptomatology and emotional distress. Psychol. Psychother. 2013, 86, 164–182. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.; Emsley, R.; Berry, K. Attachment in psychosis: A latent profile analysis of attachment styles and association with symptoms in a large psychosis cohort. Psychiatry Res. 2017, 247, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, F.; Bernasconi, A.; Bosia, M.; Cavallaro, R.; Dallaspezia, S.; Falini, A.; Poletti, S.; Radaelli, D.; Riccaboni, R.; Scotti, G.; et al. Functional and structural brain correlates of theory of mind and empathy deficits in schizophrenia. Schizophr. Res. 2009, 114, 154–160. [Google Scholar] [CrossRef]
- Herold, R.; Feldmann, Á.; Simon, M.; Tényi, T.; Kövér, F.; Nagy, F.; Varga, E.; Fekete, S. Regional gray matter reduction and theory of mind deficit in the early phase of schizophrenia: A voxel-based morphometric study. Acta Psychiatr. Scand. 2009, 119, 199–208. [Google Scholar] [CrossRef]
- Koelkebeck, K.; Hirao, K.; Miyata, J.; Kawada, R.; Saze, T.; Dannlowski, U.; Ubukata, S.; Ohrmann, P.; Bauer, J.; Pedersen, A.; et al. Impact of gray matter reductions on theory of mind abilities in patients with schizophrenia. Soc. Neurosci. 2013, 8, 631–639. [Google Scholar] [CrossRef]
- Marjoram, D.; Job, D.E.; Whalley, H.C.; Gountouna, V.-E.; McIntosh, A.M.; Simonotto, E.; Cunningham-Owens, D.; Johnstone, E.C.; Lawrie, S. A visual joke fMRI investigation into Theory of Mind and enhanced risk of schizophrenia. Neuroimage 2006, 31, 1850–1858. [Google Scholar] [CrossRef]
- Kronbichler, L.; Tschernegg, M.; Martin, A.I.; Schurz, M.; Kronbichler, M. Abnormal Brain Activation During Theory of Mind Tasks in Schizophrenia: A Meta-Analysis. Schizophr. Bull. 2017, 43, 1240–1250. [Google Scholar] [CrossRef] [Green Version]
- Vucurovic, K.; Caillies, S.; Kaladjian, A. Neural correlates of theory of mind and empathy in schizophrenia: An activation likelihood estimation meta-analysis. J. Psychiatr. Res. 2020, 120, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Herold, R.; Varga, E.; Hajnal, A.; Hamvas, E.; Berecz, H.; Tóth, B.; Tényi, T. Altered Neural Activity during Irony Comprehension in Unaffected First-Degree Relatives of Schizophrenia Patients—An fMRI Study. Front. Psychol. 2018, 8, 2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberger, D.R. Implications of Normal Brain Development for the Pathogenesis of Schizophrenia. Arch. Gen. Psychiatry 1987, 44, 660–669. [Google Scholar] [CrossRef]
- Selemon, L.D.; Zecevic, N. Schizophrenia: A tale of two critical periods for prefrontal cortical development. Transl. Psychiatry 2015, 5, e623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogtay, N. Cortical Brain Development in Schizophrenia: Insights from Neuroimaging Studies in Childhood-Onset Schizophrenia. Schizophr. Bull. 2007, 34, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander-Bloch, A.F.; Reiss, P.T.; Rapoport, J.; McAdams, H.; Giedd, J.N.; Bullmore, E.T.; Gogtay, N. Abnormal Cortical Growth in Schizophrenia Targets Normative Modules of Synchronized Development. Biol. Psychiatry 2014, 76, 438–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampis, D.; Parise, E.; Csibra, G.; Kovács, Á.M. Neural signatures for sustaining object representations attributed to others in preverbal human infants. Proc. Biol. Sci. 2015, 282, 20151683. [Google Scholar] [CrossRef]
- Grosse Wiesmann, C.; Friederici, A.D.; Singer, T.; Steinbeis, N. Two systems for thinking about others’ thoughts in the developing brain. Proc. Natl. Acad. Sci. USA 2020, 117, 6928–6935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, D.C.; Simon, C.E.; Ting, F.; Nikolaeva, J.I. Functional Organization of the Temporal–Parietal Junction for Theory of Mind in Preverbal Infants: A Near-Infrared Spectroscopy Study. J. Neurosci. 2018, 38, 4264. [Google Scholar] [CrossRef] [Green Version]
- Braehler, C.; Holowka, D.; Brunet, A.; Beaulieu, S.; Baptista, T.; Debruille, J.-B.; Walker, C.-D.; King, S. Diurnal cortisol in schizophrenia patients with childhood trauma. Schizophr. Res. 2005, 79, 353–354. [Google Scholar] [CrossRef]
- Heim, C.; Young, L.J.; Newport, D.J.; Mletzko, T.; Miller, A.H.; Nemeroff, C.B. Lower CSF oxytocin concentrations in women with a history of childhood abuse. Mol. Psychiatry 2009, 14, 954–958. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.C.M.; Sharifi, N.; Condren, R.; Thakore, J.H. Evidence of basal pituitary–adrenal overactivity in first episode, drug naïve patients with schizophrenia. Psychoneuroendocrinology 2004, 29, 1065–1070. [Google Scholar] [CrossRef]
- Mondelli, V.; Pariante, C.M.; Navari, S.; Aas, M.; D’Albenzio, A.; Di Forti, M.; Handley, R.; Hepgul, N.; Marques, T.R.; Taylor, H.; et al. Higher cortisol levels are associated with smaller left hippocampal volume in first-episode psychosis. Schizophr. Res. 2010, 119, 75–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teicher, M.H.; Andersen, S.L.; Polcari, A.; Anderson, C.M.; Navalta, C.P.; Kim, D.M. The neurobiological consequences of early stress and childhood maltreatment. Neurosci. Biobehav. Rev. 2003, 27, 33–44. [Google Scholar] [CrossRef] [PubMed]
- McCrory, E.; De Brito, S.A.; Viding, E. The impact of childhood maltreatment: A review of neurobiological and genetic factors. Front. Psychiatry 2011, 2, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnsten, A.F. Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 2009, 10, 410–422. [Google Scholar] [CrossRef]
- Brent, B.K.; Seidman, L.J.; Thermenos, H.W.; Holt, D.J.; Keshavan, M.S. Self-disturbances as a possible premorbid indicator of schizophrenia risk: A neurodevelopmental perspective. Schizophr. Res. 2014, 152, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Tas, C.; Brown, E.C.; Gonzalez, C.; Brüne, M. Experimental Usage of Oxytocin to Combat Deficits in Social Cognition in Schizophrenia. In Social Cognition and Metacognition in Schizophrenia; Lysaker, P.H., Dimaggio, G., Brüne, M., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 301–312. [Google Scholar]
- Rubin, L.H.; Carter, C.S.; Drogos, L.; Pournajafi-Nazarloo, H.; Sweeney, J.A.; Maki, P.M. Peripheral oxytocin is associated with reduced symptom severity in schizophrenia. Schizophr. Res. 2010, 124, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasayama, D.; Hattori, K.; Teraishi, T.; Hori, H.; Ota, M.; Yoshida, S.; Arima, K.; Higuchi, T.; Amano, N.; Kunugi, H. Negative correlation between cerebrospinal fluid oxytocin levels and negative symptoms of male patients with schizophrenia. Schizophr. Res. 2012, 139, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer-Shofty, M.; Shamay-Tsoory, S.; Levkovitz, Y. Characterization of the effects of oxytocin on fear recognition in patients with schizophrenia and in healthy controls. Front. Neurosci. 2013, 7, 127. [Google Scholar] [CrossRef] [Green Version]
- Fischer-Shofty, M.; Shamay-Tsoory, S.G.; Harari, H.; Levkovitz, Y. The effect of intranasal administration of oxytocin on fear recognition. Neuropsychologia 2010, 48, 179–184. [Google Scholar] [CrossRef]
- Marsh, A.A.; Yu, H.H.; Pine, D.S.; Blair, R.J. Oxytocin improves specific recognition of positive facial expressions. Psychopharmacology 2010, 209, 225–232. [Google Scholar] [CrossRef]
- Lischke, A.; Berger, C.; Prehn, K.; Heinrichs, M.; Herpertz, S.C.; Domes, G. Intranasal oxytocin enhances emotion recognition from dynamic facial expressions and leaves eye-gaze unaffected. Psychoneuroendocrinology 2011, 37, 475–481. [Google Scholar] [CrossRef]
- Lischke, A.; Gamer, M.; Berger, C.; Grossmann, A.; Hauenstein, K.; Heinrichs, M.; Herpertz, S.C.; Domes, G. Oxytocin increases amygdala reactivity to threatening scenes in females. Psychoneuroendocrinology 2012, 37, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Van IJzendoorn, M.H.; Bakermans-Kranenburg, M.J. A sniff of trust: Meta-analysis of the effects of intranasal oxytocin administration on face recognition, trust to in-group, and trust to out-group. Psychoneuroendocrinology 2012, 37, 438–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domes, G.; Heinrichs, M.; Michel, A.; Berger, C.; Herpertz, S.C. Oxytocin Improves “Mind-Reading” in Humans. Biol. Psychiatry 2007, 61, 731–733. [Google Scholar] [CrossRef] [PubMed]
- Bartz, J.A.; Zaki, J.; Bolger, N.; Hollander, E.; Ludwig, N.N.; Kolevzon, A.; Ochsner, K.N. Oxytocin selectively improves empathic accuracy. Psychol. Sci. 2010, 21, 1426–1428. [Google Scholar] [CrossRef] [PubMed]
- Krueger, F.; Parasuraman, R.; Moody, L.; Twieg, P.; de Visser, E.; McCabe, K.; O’Hara, M.; Lee, M.R. Oxytocin selectively increases perceptions of harm for victims but not the desire to punish offenders of criminal offenses. Soc. Cogn. Affect. Neurosci. 2012, 8, 494–498. [Google Scholar] [CrossRef]
- Kosfeld, M.; Heinrichs, M.; Zak, P.J.; Fischbacher, U.; Fehr, E. Oxytocin increases trust in humans. Nature 2005, 435, 673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoridou, A.; Rowe, A.C.; Penton-Voak, I.S.; Rogers, P.J. Oxytocin and social perception: Oxytocin increases perceived facial trustworthiness and attractiveness. Horm. Behav. 2009, 56, 128–132. [Google Scholar] [CrossRef]
- Averbeck, B.B.; Bobin, T.; Evans, S.; Shergill, S.S. Emotion recognition and oxytocin in patients with schizophrenia. Psychol. Med. 2012, 42, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, C.A.; Gibson, C.M.; Rau, S.W.; Salimi, K.; Smedley, K.L.; Casey, R.L.; Leserman, J.; Jarskog, L.F.; Penn, D.L. Intranasal oxytocin reduces psychotic symptoms and improves Theory of Mind and social perception in schizophrenia. Schizophr. Res. 2011, 132, 50–53. [Google Scholar] [CrossRef]
- Walss-Bass, C.; Fernandes, J.M.; Roberts, D.L.; Service, H.; Velligan, D. Differential correlations between plasma oxytocin and social cognitive capacity and bias in schizophrenia. Schizophr. Res. 2013, 147, 387–392. [Google Scholar] [CrossRef]
- Goldman, M.; Marlow-O’Connor, M.; Torres, I.; Carter, C.S. Diminished plasma oxytocin in schizophrenic patients with neuroendocrine dysfunction and emotional deficits. Schizophr. Res. 2008, 98, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Kéri, S.; Kiss, I.; Kelemen, O. Sharing secrets: Oxytocin and trust in schizophrenia. Soc. Neurosci. 2009, 4, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Jobst, A.; Dehning, S.; Ruf, S.; Notz, T.; Buchheim, A.; Henning-Fast, K.; Meißner, D.; Meyer, S.; Bondy, B.; Müller, N.; et al. Oxytocin and vasopressin levels are decreased in the plasma of male schizophrenia patients. Acta Neuropsychiatr. 2014, 26, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. Sniffing around oxytocin: Review and meta-analyses of trials in healthy and clinical groups with implications for pharmacotherapy. Transl. Psychiatry 2013, 3, e258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broniarczyk-Czarniak, M.; Szemraj, J.; Smigielski, J.; Galecki, P. The Role of OXT, OXTR, AVP, and AVPR1a Gene Expression in the Course of Schizophrenia. Curr. Issues Mol. Biol. 2022, 44, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.P.; De Luca, V.; Meltzer, H.Y.; Lieberman, J.A.; Kennedy, J.L. Schizophrenia severity and clozapine treatment outcome association with oxytocinergic genes. Int. J. Neuropsychopharmacol. 2010, 13, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Montag, C.; Dziobek, I.; Richter, I.S.; Neuhaus, K.; Lehmann, A.; Sylla, R.; Heekeren, H.R.; Heinz, A.; Gallinat, J. Different aspects of theory of mind in paranoid schizophrenia: Evidence from a video-based assessment. Psychiatry Res. 2011, 186, 203–209. [Google Scholar] [CrossRef]
- Do, K.Q.; Cabungcal, J.H.; Frank, A.; Steullet, P.; Cuenod, M. Redox dysregulation, neurodevelopment, and schizophrenia. Curr. Opin. Neurobiol. 2009, 19, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Takahashi, T.; Sumitani, K.; Takatsu, H.; Urano, S. Glucocorticoid generates ROS to induce oxidative injury in the hippocampus, leading to impairment of cognitive function of rats. J. Clin. Biochem. Nutr. 2010, 47, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Do, K.Q.; Trabesinger, A.H.; Kirsten-Krüger, M.; Lauer, C.J.; Dydak, U.; Hell, D.; Holsboer, F.; Boesiger, P.; Cuénod, M. Schizophrenia: Glutathione deficit in cerebrospinal fluid and prefrontal cortex in vivo. Eur. J. Neurosci. 2000, 12, 3721–3728. [Google Scholar] [CrossRef]
- Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. The first 10,000 Adult Attachment Interviews: Distributions of adult attachment representations in clinical and non-clinical groups. Attach. Hum. Dev. 2009, 11, 223–263. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-IV; American Psychiatric Association: Washington, DC, USA, 1994. [Google Scholar]
- Wittchen, H.U.; Zaudig, M.; Fydrich, T. SKID. Strukturiertes Klinisches Interview für DSM-IV. Achse I und II.; Hogrefe: Göttingen, Germany, 1997. [Google Scholar]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The Positive and Negative Syndrome Scale (PANSS) for Schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Leucht, S.; Kane, J.M.; Kissling, W.; Hamann, J.; Etschel, E.; Engel, R.R. What does the PANSS mean? Schizophr. Res. 2005, 79, 231–238. [Google Scholar] [CrossRef]
- Buchheim, A.; Viviani, R.; Kessler, H.; Kächele, H.; Cierpka, M.; Roth, G.; George, C.; Kernberg, O.F.; Bruns, G.; Taubner, S. Changes in Prefrontal-Limbic Function in Major Depression after 15 Months of Long-Term Psychotherapy. PLoS ONE 2012, 7, e33745. [Google Scholar] [CrossRef] [PubMed]
- George, C.; West, M. The development and preliminary validation of a new measure of adult attachment: The adult attachment projective. Attach. Hum. Dev. 2001, 3, 30–61. [Google Scholar] [CrossRef] [PubMed]
- Buchheim, A.; Labek, K.; Taubner, S.; Kessler, H.; Pokorny, D.; Kachele, H.; Cierpka, M.; Roth, G.; Pogarell, O.; Karch, S. Modulation of Gamma Band Activity and Late Positive Potential in Patients with Chronic Depression after Psychodynamic Psychotherapy. Psychother. Psychosom. 2018, 87, 252–254. [Google Scholar] [CrossRef]
- Gander, M.; Karabatsiakis, A.; Nuderscher, K.; Bernheim, D.; Doyen-Waldecker, C.; Buchheim, A. Secure Attachment Representation in Adolescence Buffers Heart-Rate Reactivity in Response to Attachment-Related Stressors. Front. Hum. Neurosci. 2022, 16, 806987. [Google Scholar] [CrossRef]
- Bernheim, D.; Buchheim, A.; Domin, M.; Mentel, R.; Lotze, M. Neural Correlates of Attachment Representation in Patients with Borderline Personality Disorder Using a Personalized Functional Magnet Resonance Imaging Task. Front. Hum. Neurosci. 2022, 16, 810417. [Google Scholar] [CrossRef]
- Buchheim, A.; George, C. Attachment disorganization in borderline personality disorder and anxiety disorder. In Disorganization of Attachment and Caregiving; Solomon, J., George, V., Eds.; Guilford Press: New York, NY, USA, 2011; pp. 343–383. [Google Scholar]
- Hautzinger, M.; Bailer, M.; Worall, H.; Keller, F. Beck-Depressions-Inventar (BDI). Testhandbuch (2. überarbeitete Auflage); Huber: Bern, Switzerland, 1995. [Google Scholar]
- Beck, A.T.; Steer, R.A.; Brown, G.K. Manual for the Beck Depression Inventory-II; Psychological Corporation: San Antonio, TX, USA, 1996. [Google Scholar]
- Laux, L.; Glanzmann, P.; Schaffner, P.; Spielberger, C.D. Das State-Trait-Angstinventar (Testmappe mit Handanweisung, Fragebogen STAI–G Form X 1 und Fragebogen STAI–G Form X 2); Beltz: Weinheim, Germany, 1981. [Google Scholar]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and Validation of Brief Measures of Positive and Negative Affect: The PANAS Scales. J. Pers. Soc. Psychol. 1988, 54, 1063. [Google Scholar] [CrossRef]
- Schmidt, K.H.; Metzler, P. Wortschatztest (WST); Beltz Test GmbH: Göttingen, Germany, 1992. [Google Scholar]
- Chapman, L.J.; Chapman, J.P. The measurement of handedness. Brain Cogn. 1987, 6, 175–183. [Google Scholar] [CrossRef]
- Zimmermann, P.; Fimm, B. Testbatterie zur Aufmerksamkeitsprüfung (TAP), Version 2.1; Psytest Psychologische Testsysteme: Herzogenrath, Germany, 2007. [Google Scholar]
- Von Aster, M.; Neubauer, A.; Horn, R. Wechsler Intelligenztest für Erwachsene (Wechsler Intelligence Scale for Adults); Hogrefe: Göttingen, Germany, 2006. [Google Scholar]
- Haralanova, E.; Haralanov, S.; Beraldi, A.; Möller, H.-J.; Hennig-Fast, K. Subjective emotional over-arousal to neutral social scenes in paranoid schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2012, 262, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Carter, C.S.; Ying, J.; Bellugi, U.; Pournajafi-Nazarloo, H.; Korenberg, J.R. Oxytocin and Vasopressin Are Dysregulated in Williams Syndrome, a Genetic Disorder Affecting Social Behavior. PLoS ONE 2012, 7, e38513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisman, O.; Zagoory-Sharon, O.; Schneiderman, I.; Gordon, I.; Feldman, R. Plasma oxytocin distributions in a large cohort of women and men and their gender-specific associations with anxiety. Psychoneuroendocrinology 2013, 38, 694–701. [Google Scholar] [CrossRef]
- Abu-Akel, A. Impaired theory of mind in schizophrenia. Pragmat. Cogn. 1999, 7, 247–282. [Google Scholar] [CrossRef]
- Abu-Akel, A.; Bailey, A.L. The possibility of different forms of theory of mind impairment in psychiatric and developmental disorders. Psychol. Med. 2000, 30, 735–738. [Google Scholar] [CrossRef]
- Abu-Akel, A.; Shamay-Tsoory, S. Neuroanatomical and neurochemical bases of theory of mind. Neuropsychologia 2011, 49, 2971–2984. [Google Scholar] [CrossRef] [PubMed]
- Fonagy, P.; Gergely, G.; Target, M. The parent–infant dyad and the construction of the subjective self. J. Child Psychol. Psychiatry 2007, 48, 288–328. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, R.; Cahill, C.; Frith, C.D. The appreciation of visual jokes in people with schizophrenia: A study of ‘mentalizing’ ability. Schizophr. Res. 1997, 24, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Frith, C.D.; Corcoran, R. Exploring ‘theory of mind’ in people with schizophrenia. Psychol. Med. 1996, 26, 521–530. [Google Scholar] [CrossRef]
- Sarfati, Y.; Hardy-Baylé, M.-C.; Brunet, E.; Widlöcher, D. Investigating theory of mind in schizophrenia: Influence of verbalization in disorganized and non-disorganized patients. Schizophr. Res. 1999, 37, 183–190. [Google Scholar] [CrossRef]
- Bora, E.; Pantelis, C. Theory of mind impairments in first-episode psychosis, individuals at ultra-high risk for psychosis and in first-degree relatives of schizophrenia: Systematic review and meta-analysis. Schizophr. Res. 2013, 144, 31–36. [Google Scholar] [CrossRef]
- Pierrehumbert, B.; Torrisi, R.; Ansermet, F.; Borghini, A.; Halfon, O. Adult attachment representations predict cortisol and oxytocin responses to stress. Attach. Hum. Dev. 2012, 14, 453–476. [Google Scholar] [CrossRef]
- De Dreu, C.K.W. Oxytocin modulates cooperation within and competition between groups: An integrative review and research agenda. Horm. Behav. 2012, 61, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Colonnello, V.; Chen, F.S.; Panksepp, J.; Heinrichs, M. Oxytocin sharpens self-other perceptual boundary. Psychoneuroendocrinology 2013, 38, 2996–3002. [Google Scholar] [CrossRef] [PubMed]
- Goh, K.K.; Lu, M.-L. Relationship between the domains of theory of mind, social dysfunction, and oxytocin in schizophrenia. J. Psychiatr. Res. 2022, 155, 420–429. [Google Scholar] [CrossRef]
- Abu-Akel, A. A neurobiological mapping of theory of mind. Brain Res. Rev. 2003, 43, 29–40. [Google Scholar] [CrossRef]
- Nakata, H.; Sakamoto, K.; Ferretti, A.; Gianni Perrucci, M.; Del Gratta, C.; Kakigi, R.; Luca Romani, G. Somato-motor inhibitory processing in humans: An event-related functional MRI study. Neuroimage 2008, 39, 1858–1866. [Google Scholar] [CrossRef] [PubMed]
- Blumenfeld, R.S.; Parks, C.M.; Yonelinas, A.P.; Ranganath, C. Putting the Pieces Together: The Role of Dorsolateral Prefrontal Cortex in Relational Memory Encoding. J. Cogn. Neurosci. 2011, 23, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Murray, L.J.; Ranganath, C. The Dorsolateral Prefrontal Cortex Contributes to Successful Relational Memory Encoding. J. Neurosci. 2007, 27, 5515. [Google Scholar] [CrossRef] [Green Version]
- Wylie, K.P.; Tregellas, J.R. The role of the insula in schizophrenia. Schizophr. Res. 2010, 123, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Devue, C.; Collette, F.; Balteau, E.; Degueldre, C.; Luxen, A.; Maquet, P.; Brédart, S. Here I am: The cortical correlates of visual self-recognition. Brain Res. 2007, 1143, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Van Veluw, S.J.; Chance, S.A. Differentiating between self and others: An ALE meta-analysis of fMRI studies of self-recognition and theory of mind. Brain Imaging Behav. 2014, 8, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, K.N.; Zaki, J.; Hanelin, J.; Ludlow, D.H.; Knierim, K.; Ramachandran, T.; Glover, G.H.; Mackey, S.C. Your pain or mine? Common and distinct neural systems supporting the perception of pain in self and other. Soc. Cogn. Affect. Neurosci. 2008, 3, 144–160. [Google Scholar] [CrossRef] [Green Version]
- Singer, T. The neuronal basis and ontogeny of empathy and mind reading: Review of literature and implications for future research. Neurosci. Biobehav. Rev. 2006, 30, 855–863. [Google Scholar] [CrossRef]
- Posner, M.I.; Rothbart, M.K.; Sheese, B.E.; Tang, Y. The anterior cingulate gyrus and the mechanism of self-regulation. Cogn. Affect. Behav. Neurosci. 2007, 7, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapin, H.; Bagarinao, E.; Mackey, S. Real-time fMRI applied to pain management. Neurosci. Lett. 2012, 520, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Han, S. Attention and reality constraints on the neural processes of empathy for pain. Neuroimage 2007, 36, 256–267. [Google Scholar] [CrossRef]
- Etkin, A.; Egner, T.; Peraza, D.M.; Kandel, E.R.; Hirsch, J. Resolving Emotional Conflict: A Role for the Rostral Anterior Cingulate Cortex in Modulating Activity in the Amygdala. Neuron 2006, 51, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apps, M.A.J.; Balsters, J.H.; Ramnani, N. The anterior cingulate cortex: Monitoring the outcomes of others’ decisions. Soc. Neurosci. 2012, 7, 424–435. [Google Scholar] [CrossRef]
- D’Argembeau, A.; Ruby, P.; Collette, F.; Degueldre, C.; Balteau, E.; Luxen, A.; Maquet, P.; Salmon, E. Distinct Regions of the Medial Prefrontal Cortex Are Associated with Self-referential Processing and Perspective Taking. J. Cogn. Neurosci. 2007, 19, 935–944. [Google Scholar] [CrossRef]
- Sajonz, B.; Kahnt, T.; Margulies, D.S.; Park, S.Q.; Wittmann, A.; Stoy, M.; Ströhle, A.; Heinz, A.; Northoff, G.; Bermpohl, F. Delineating self-referential processing from episodic memory retrieval: Common and dissociable networks. Neuroimage 2010, 50, 1606–1617. [Google Scholar] [CrossRef]
- Cavanna, A.E.; Trimble, M.R. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 2006, 129, 564–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, J.A.; Garrison, K.A.; Whitfield-Gabrieli, S. What about the “Self” is Processed in the Posterior Cingulate Cortex? Front. Hum. Neurosci. 2013, 7, 647. [Google Scholar] [CrossRef] [Green Version]
- Brody, A.L.; Mandelkern, M.A.; Olmstead, R.E.; Jou, J.; Tiongson, E.; Allen, V.; Scheibal, D.; London, E.D.; Monterosso, J.R.; Tiffany, S.T.; et al. Neural Substrates of Resisting Craving During Cigarette Cue Exposure. Biol. Psychiatry 2007, 62, 642–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippi, C.L.; Tranel, D.; Duff, M.; Rudrauf, D. Damage to the default mode network disrupts autobiographical memory retrieval. Soc. Cogn. Affect. Neurosci. 2014, 10, 318–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews-Hanna, J.R.; Reidler, J.S.; Sepulcre, J.; Poulin, R.; Buckner, R.L. Functional-Anatomic Fractionation of the Brain’s Default Network. Neuron 2010, 65, 550–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissman, D.H.; Roberts, K.C.; Visscher, K.M.; Woldorff, M.G. The neural bases of momentary lapses in attention. Nat. Neurosci. 2006, 9, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Liu, Y.; Yao, L.; Ding, M. Top-Down Regulation of Default Mode Activity in Spatial Visual Attention. J. Neurosci. 2013, 33, 6444. [Google Scholar] [CrossRef] [Green Version]
- Sowden, S.; Catmur, C. The Role of the Right Temporoparietal Junction in the Control of Imitation. Cereb. Cortex 2013, 25, 1107–1113. [Google Scholar] [CrossRef] [Green Version]
- Nolte, T.; Bolling, D.; Hudac, C.; Fonagy, P.; Mayes, L.; Pelphrey, K. Brain mechanisms underlying the impact of attachment-related stress on social cognition. Front. Hum. Neurosci. 2013, 7, 816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kober, H.; Barrett, L.F.; Joseph, J.; Bliss-Moreau, E.; Lindquist, K.; Wager, T.D. Functional grouping and cortical–subcortical interactions in emotion: A meta-analysis of neuroimaging studies. Neuroimage 2008, 42, 998–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wager, T.D.; Barrett, L.F.; Bliss-Moreau, E.; Lindquist, K.A.; Duncan, S.; Kober, H.; Joseph, J.; Davidson, M.; Mize, J. The Neuroimaging of Emotion. In Handbook of Emotions, 3rd ed.; Lewis, M., Haviland-Jones, J.M., Barrett, L.F., Eds.; The Guilford Press: New York, NY, USA, 2008; pp. 249–271. [Google Scholar]
- Freund, W.; Stuber, G.; Wunderlich, A.P.; Schmitz, B. Cortical correlates of perception and suppression of electrically induced pain. Somatosens. Mot. Res. 2007, 24, 203–212. [Google Scholar] [CrossRef]
- Nielsen, F.Å.; Balslev, D.; Hansen, L.K. Mining the posterior cingulate: Segregation between memory and pain components. Neuroimage 2005, 27, 520–532. [Google Scholar] [CrossRef] [Green Version]
- Buchheim, A.; Erk, S.; George, C.; Kachele, H.; Kircher, T.; Martius, P.; Pokorny, D.; Ruchsow, M.; Spitzer, M.; Walter, H. Neural correlates of attachment trauma in borderline personality disorder: A functional magnetic resonance imaging study. Psychiatry Res. 2008, 163, 223–235. [Google Scholar] [CrossRef]
- Baskak, B.; Kir, Y.; Sedes, N.; Kuşman, A.; Türk, E.G.; Baran, Z.; Gönüllü, I.; Artar, M.; Munir, K. Attachment Style Predicts Cortical Activity in Temporoparietal Junction (TPJ): An fNIRS Study Using a Theory of Mind (ToM) Task in Healthy University Students. J. Psychophysiol. 2020, 34, 99–109. [Google Scholar] [CrossRef]
- Chan, R.C.K.; Di, X.; McAlonan, G.M.; Gong, Q.-y. Brain Anatomical Abnormalities in High-Risk Individuals, First-Episode, and Chronic Schizophrenia: An Activation Likelihood Estimation Meta-analysis of Illness Progression. Schizophr. Bull. 2009, 37, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison-Wright, I.; Glahn, D.C.; Laird, A.R.; Thelen, S.M.; Bullmore, E. The Anatomy of First-Episode and Chronic Schizophrenia: An Anatomical Likelihood Estimation Meta-Analysis. Am. J. Psychiatry 2008, 165, 1015–1023. [Google Scholar] [CrossRef]
- Kuperberg, G.R.; Broome, M.R.; McGuire, P.K.; David, A.S.; Eddy, M.; Ozawa, F.; Goff, D.; West, W.C.; Williams, S.C.R.; van der Kouwe, A.J.W.; et al. Regionally Localized Thinning of the Cerebral Cortex in Schizophrenia. Arch. Gen. Psychiatry 2003, 60, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Nesvåg, R.; Lawyer, G.; Varnäs, K.; Fjell, A.M.; Walhovd, K.B.; Frigessi, A.; Jönsson, E.G.; Agartz, I. Regional thinning of the cerebral cortex in schizophrenia: Effects of diagnosis, age and antipsychotic medication. Schizophr. Res. 2008, 98, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Sheffield, J.M.; Rogers, B.P.; Blackford, J.U.; Heckers, S.; Woodward, N.D. Insula functional connectivity in schizophrenia. Schizophr. Res. 2020, 220, 69–77. [Google Scholar] [CrossRef]
- Yao, B.; Gu, P.; Lasagna, C.A.; Peltier, S.; Taylor, S.F.; Tso, I.F.; Thakkar, K.N. Structural connectivity of an interoception network in schizophrenia. Neuroimaging 2023, 331, 111636. [Google Scholar] [CrossRef]
- Gebhardt, S.; Nasrallah, H.A. The role of the insula in cognitive impairment of schizophrenia. Schizophr. Res. Cogn. 2023, 32, 100277. [Google Scholar] [CrossRef] [PubMed]
- Tran The, J.; Magistretti, P.J.; Ansermet, F. Interoception Disorder and Insular Cortex Abnormalities in Schizophrenia: A New Perspective Between Psychoanalysis and Neuroscience. Front. Psychol. 2021, 12, 628355. [Google Scholar] [CrossRef] [PubMed]
- Jardri, R.; Pouchet, A.; Pins, D.; Thomas, P. Cortical Activations During Auditory Verbal Hallucinations in Schizophrenia: A Coordinate-Based Meta-Analysis. Am. J. Psychiatry 2011, 168, 73–81. [Google Scholar] [CrossRef]
- Palaniyappan, L.; Liddle, P.F. Does the salience network play a cardinal role in psychosis? An emerging hypothesis of insular dysfunction. J. Psychiatry Neurosci. 2012, 37, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, J.; Fosse, R.; Moskowitz, A.; Perry, B. The traumagenic neurodevelopmental model of psychosis revisited. Neuropsychiatry 2014, 4, 65–79. [Google Scholar] [CrossRef]
- Brent, B.K.; Holt, D.J.; Keshavan, M.S.; Seidman, L.J.; Fonagy, P. Mentalization-based Treatment for Psychosis: Linking an Attachment-based Model to the Psychotherapy for Impaired Mental State Understanding in People with Psychotic Disorders. Isr. J. Psychiatry Relat. Sci. 2014, 51, 17–24. [Google Scholar] [PubMed]
- Brent, B.K.; Fonagy, P. A mentalization-based treatment approach to disturbances of social understanding in schizophrenia. In Social Cognition and Metacognition in Schizophrenia; Lysaker, P., Dimaggio, G., Brune, M., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 245–259. [Google Scholar]
- Guy, W. ECDEU Assessment Manual for Psychopharmacology Revised (DHEW Publ. No. ADM 76-338); National Institute of Mental Health: Rockville, MD, USA, 1976. [Google Scholar]
- Briones, T.L.; Klintsova, A.Y.; Greenough, W.T. Stability of synaptic plasticity in the adult rat visual cortex induced by complex environment exposure. Brain Res. 2004, 1018, 130–135. [Google Scholar] [CrossRef]
- Rincon-Cortes, M.; Sullivan, R.M. Early life trauma and attachment: Immediate and enduring effects on neurobehavioral and stress axis development. Front. Endocrinol. 2014, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.S.; Hayes, S.C.; Biglan, A.; Embry, D.D. Evolving the future: Toward a science of intentional change. Behav. Brain Sci. 2014, 37, 395–416. [Google Scholar] [CrossRef]
- Beebe, B.; Lachmann, F. The Origins of Attachment: Infant Research and Adult Treatment; Routledge: London, UK, 2013. [Google Scholar]
- Holmes, J.; Slade, A. Attachment in Therapeutic Practice; Sage: London, UK, 2017. [Google Scholar]
- Slade, A. Imagining Fear: Attachment, Threat, and Psychic Experience. Psychoanal. Dialogues 2014, 24, 253–266. [Google Scholar] [CrossRef]
- Brown, C.; Coan, J.A. The social regulation of neural threat responding. In The Social Neuroscience of Human-Animal Interaction; Freund, L.S., McCune, S., Esposito, F., Gee, N.R., McCardle, P., Eds.; American Psychological Association: Washington, DC, USA, 2016; pp. 127–146. [Google Scholar]
- Csulak, T.; Hajnal, A.; Kiss, S.; Dembrovszky, F.; Varjú-Solymár, M.; Sipos, Z.; Kovács, M.A.; Herold, M.; Varga, E.; Hegyi, P.; et al. Implicit Mentalizing in Patients with Schizophrenia: A Systematic Review and Meta-Analysis. Front. Psychol. 2022, 13, 790494. [Google Scholar] [CrossRef]
- Sprong, M.; Schothorst, P.; Vos, E.; Hox, J.; Van Engeland, H. Theory of mind in schizophrenia: Meta-analysis. Br. J. Psychiatry 2007, 191, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, A.K.; Robinson, G.; Dzafic, I.; Reutens, D.; Mowry, B. Theory of mind and the social brain: Implications for understanding the genetic basis of schizophrenia. Genes Brain Behav. 2014, 13, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Bora, E.; Yucel, M.; Pantelis, C. Theory of mind impairment in schizophrenia: Meta-analysis. Schizophr. Res. 2009, 109, 1–9. [Google Scholar] [CrossRef]
- Herold, R.; Tényi, T.; Lénárd, K.; Trixler, M. Theory of mind deficit in people with schizophrenia during remission. Psychol. Med. 2002, 32, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.M.; Penn, D.L.; Perkins, D.; Woods, S.W.; Addington, J. Theory of mind and social judgments in people at clinical high risk of psychosis. Schizophr. Res. 2013, 150, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Davidson, C.A.; Piskulic, D.; Addington, J.; Cadenhead, K.S.; Cannon, T.D.; Cornblatt, B.A.; McGlashan, T.H.; Perkins, D.O.; Seidman, L.J.; Tsuang, M.T.; et al. Age-related trajectories of social cognition in youth at clinical high risk for psychosis: An exploratory study. Schizophr. Res. 2018, 201, 130–136. [Google Scholar] [CrossRef]
- Tikka, D.L.; Singh, A.R.; Tikka, S.K. Social cognitive endophenotypes in schizophrenia: A study comparing first episode schizophrenia patients and, individuals at clinical- and familial- ‘at-risk’ for psychosis. Schizophr. Res. 2020, 215, 157–166. [Google Scholar] [CrossRef]
- Velthorst, E.; Fett, A.-K.J.; Reichenberg, A.; Perlman, G.; van Os, J.; Bromet, E.J.; Kotov, R. The 20-Year Longitudinal Trajectories of Social Functioning in Individuals with Psychotic Disorders. Am. J. Psychiatry 2017, 174, 1075–1085. [Google Scholar] [CrossRef]
- Langdon, R.; Flynn, M.; Connaughton, E.; Brüne, M. Impairments of spontaneous and deliberative mentalizing co-occur, yet dissociate, in schizophrenia. Br. J. Clin. Psychol. 2017, 56, 372–387. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients (N = 20) | Controls (N = 20) | Statistics | Significance | |
|---|---|---|---|---|
| Age (Mean, SD) | 24.3 (3.7) | 25.3 (3.2) | 0.8 2 | 0.370 (n.s.) |
| (range: 18–30) | (range: 19–30) | |||
| Years of education | 12.2 (1.5) | 12.6 (1.6) | 0.5 2 | 0.482 (n.s.) |
| Handedness | right:17 | right:17 | 2.57 3 | 0.349 (n.s.) |
| left: 1 | left: 3 | |||
| ambydexter: 2 | ambydexter: 0 | |||
| VIQ (WST) | 106.3 (9.8) | 111.3 (11.7) | 2.2 2 | 0.150 (n.s.) |
| STAI-T | 40.7 (11.2) 1 | 36.5 (8.2) 1 | 1.8 2 | 0.189 (n.s.) |
| STAI-S | 39.4 (7.5) 1 | 40.1 (8.3) 1 | 0.1 2 | 0.781 (n.s.) |
| PANAS | ||||
| PA | 30.2 (6.3) 1 | 34.9 (4.8) 1 | 7.0 2 | 0.012 (*) |
| NA | 19.6 (4.8) 1 | 18.8 (6.7) 1 | 0.2 2 | 0.666 (n.s.) |
| Statistics (Between Diagnosis Group Difference) | Significance | ||
|---|---|---|---|
| Diagnosis/ | paranoid: 13× | ||
| schizophrenia | disorganized: 5× | ||
| subtype | undifferentiated: 2× | ||
| Duration of illness since 1. SPA | 3.96 (3.18) | 0.4 1 | 0.701 (n. s.) |
| PANSS total positive negative | 55.9 (13.4) 9.75 (2.5) 18.6 (5.5) | 3.95 2 3.84 2 2.19 2 | 0.139 (n. s.) 0.147 (n. s.) 0.334 (n. s.) |
| Patients (N = 20) | Controls (N = 20) | Statistics | Significance | |
|---|---|---|---|---|
| Oxytocin (1st measurement) | 266.5 (106.7) pg/mL | 386.6 (221.4) pg/mL | −2.0 1 | 0.043 (*) |
| Oxytocin (2nd measurement) | 258.1 (105.6) pg/mL | 385.8 (200.4) pg/mL | −2.2 1 | 0.028 (*) |
| Oxytocin (mean) | 262.3 (102.0) pg/mL | 386.2 (208.3) pg/mL | −2.1 1 | 0.033 (*) |
| Brodmann Area | Hemisphere | Z-Scores | Cluster-Size | MNI-Coordinates (x,y,z) | |||
|---|---|---|---|---|---|---|---|
| Cingulate gyrus | BA24 BA31 BA32 | L | 4.56 | 2129 | −16 | −10 | 38 |
| R | 4.10 | 6 | 10 | 34 | |||
| L | 5.18 | −16 | −18 | 38 | |||
| R | 3.80 | 18 | −36 | 44 | |||
| L | 3.30 | −10 | 20 | 32 | |||
| R | 3.85 | 4 | 22 | 28 | |||
| Precuneus | BA7 | L | 3.82 | 2129 | −20 | −52 | 48 |
| R | 3.10 | 12 | −38 | 54 | |||
| TPJ | BA40 | L | 3.89 | 2129 | −36 | −46 | 48 |
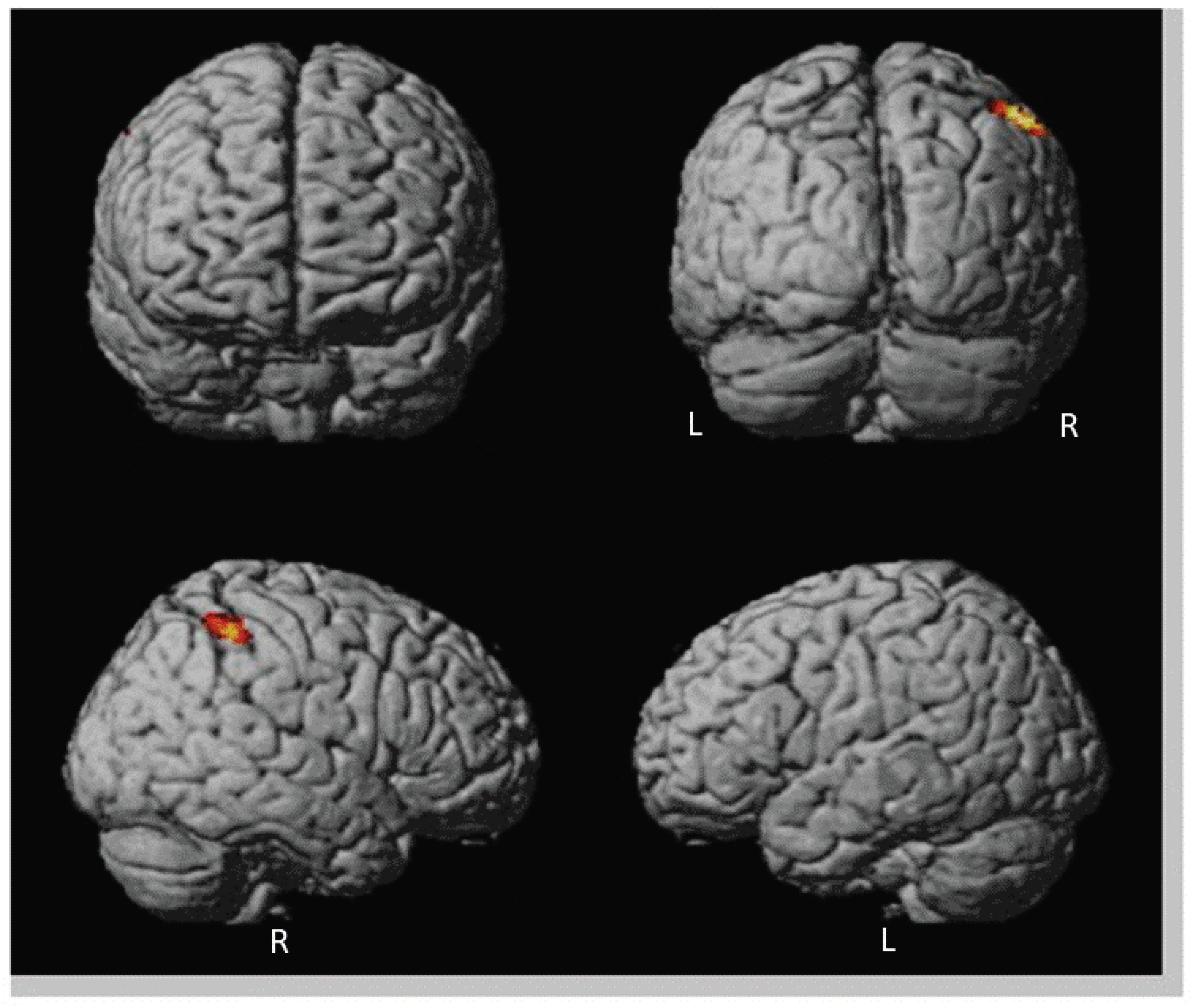
| R | 3.21 | 565 | 54 | −30 | 28 | ||
| Precentral gyrus | BA6 | L | 4.36 | 2129 | −32 | −10 | 38 |
| BA4 | L | 4.16 | −30 | −14 | 40 | ||
| Postcentral gyrus | BA2 | R | 4.01 | 565 | 60 | −18 | 26 |
| Insula | BA13 | L | 3.69 | 402 | −48 | −16 | 14 |
| R | 4.06 | 565 | 42 | −32 | 20 | ||
| Claustrum | - | L | 4.57 | 2129 | −38 | −4 | 6 |
| Brodmann Area | Hemisphere | Z-Scores | Cluster-Size | MNI-Coordinates (x,y,z) | |||
|---|---|---|---|---|---|---|---|
| Posterior cingulate gyrus | 23/31 | L | 4.60 | 137 | −6 | −50 | 24 |
| Brodmann Area | Hemisphere | Z-Scores | Cluster-Size | MNI-Coordinates (x,y,z) | |||
|---|---|---|---|---|---|---|---|
| TPJ | 40 | R | 3.98 | 92 | 50 | −48 | 56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hennig-Fast, K.; Meissner, D.; Steuwe, C.; Dehning, S.; Blautzik, J.; Eilert, D.W.; Zill, P.; Müller, N.; Meindl, T.; Reiser, M.; et al. The Interplay of Oxytocin and Attachment in Schizophrenic Patients: An fMRI Study. Brain Sci. 2023, 13, 1125. https://doi.org/10.3390/brainsci13081125
Hennig-Fast K, Meissner D, Steuwe C, Dehning S, Blautzik J, Eilert DW, Zill P, Müller N, Meindl T, Reiser M, et al. The Interplay of Oxytocin and Attachment in Schizophrenic Patients: An fMRI Study. Brain Sciences. 2023; 13(8):1125. https://doi.org/10.3390/brainsci13081125
Chicago/Turabian StyleHennig-Fast, Kristina, Dominik Meissner, Carolin Steuwe, Sandra Dehning, Janusch Blautzik, Dirk W. Eilert, Peter Zill, Norbert Müller, Thomas Meindl, Maximilian Reiser, and et al. 2023. "The Interplay of Oxytocin and Attachment in Schizophrenic Patients: An fMRI Study" Brain Sciences 13, no. 8: 1125. https://doi.org/10.3390/brainsci13081125