
Chronic Intermittent Ethanol Administration during Adolescence Produces Sex Dependent Impairments in Behavioral Flexibility and Survivability

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
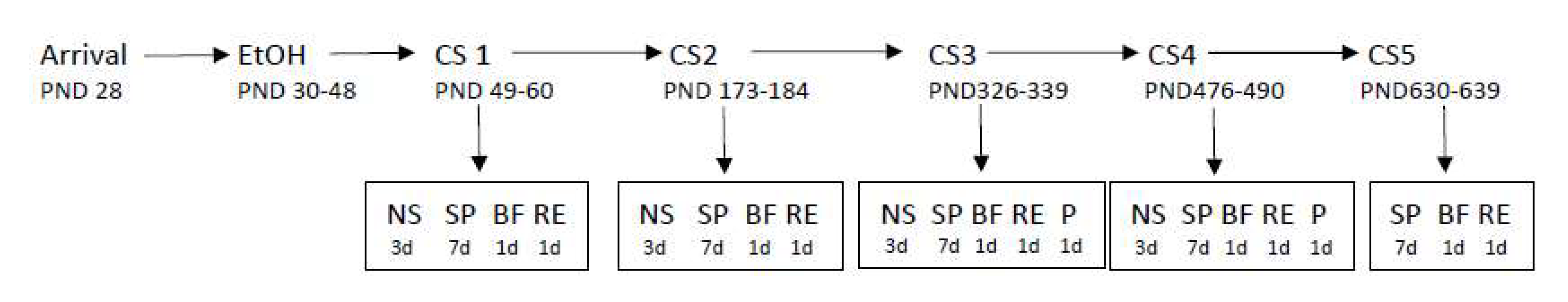
2.2. Chronic Intermittent Ethanol Exposure via Gavage (CIEg)
2.3. Blood Ethanol Concentration
2.4. Impact of CIE during Adolescence on Non-Spatial and Spatial Learning
2.5. Non-Spatial, Cue-Based, Learning (for Cognitive Sessions 1–4)
2.6. Spatial, Location-Based, Learning (for Cognitive Sessions 1–5)
2.7. Probe Trials
2.8. Statistical Analysis
3. Results
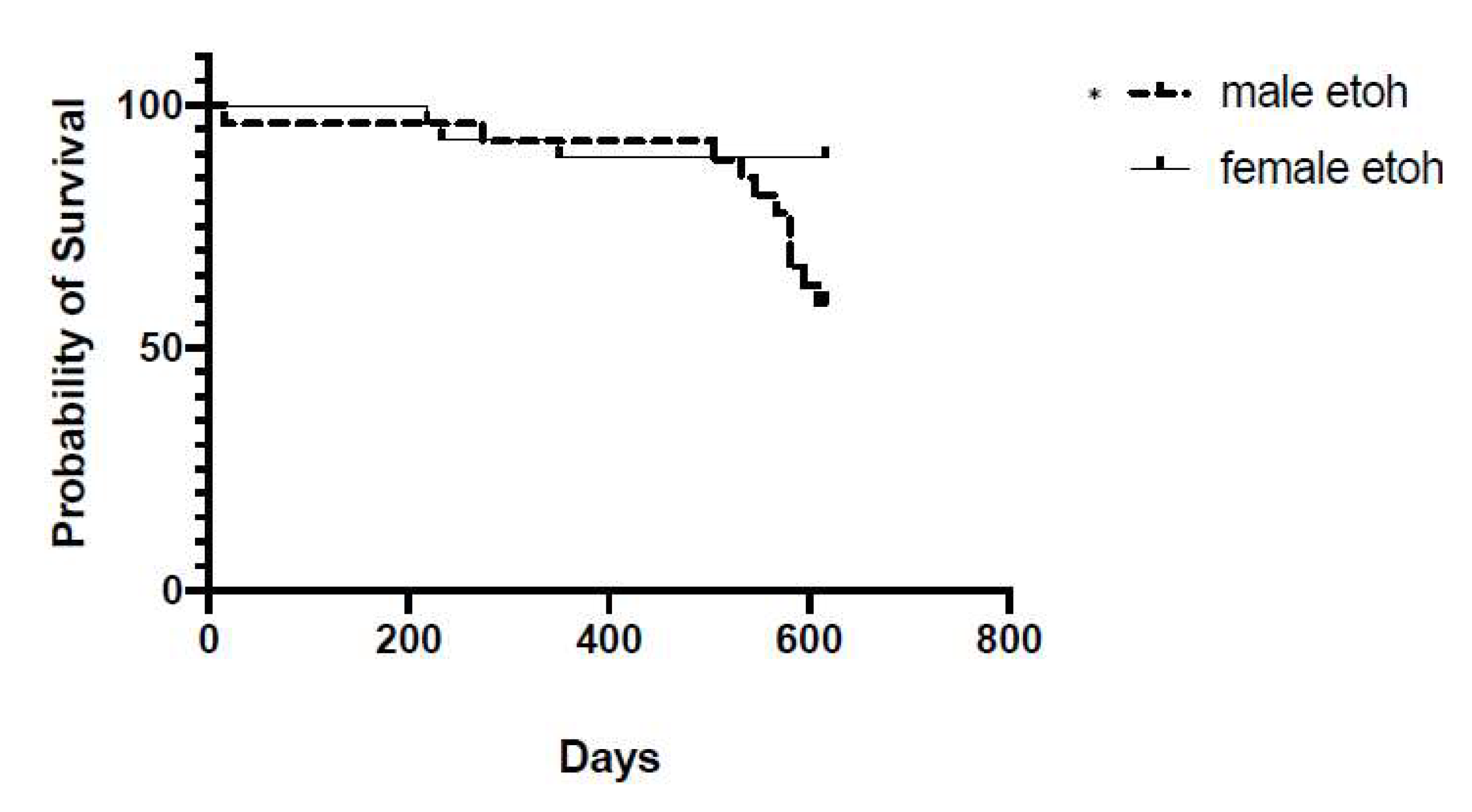
3.1. Impact of Chronic Intermittent Ethanol Exposure on Survival
3.2. Impact of Chronic Intermittent Ethanol Exposure on Bodyweight
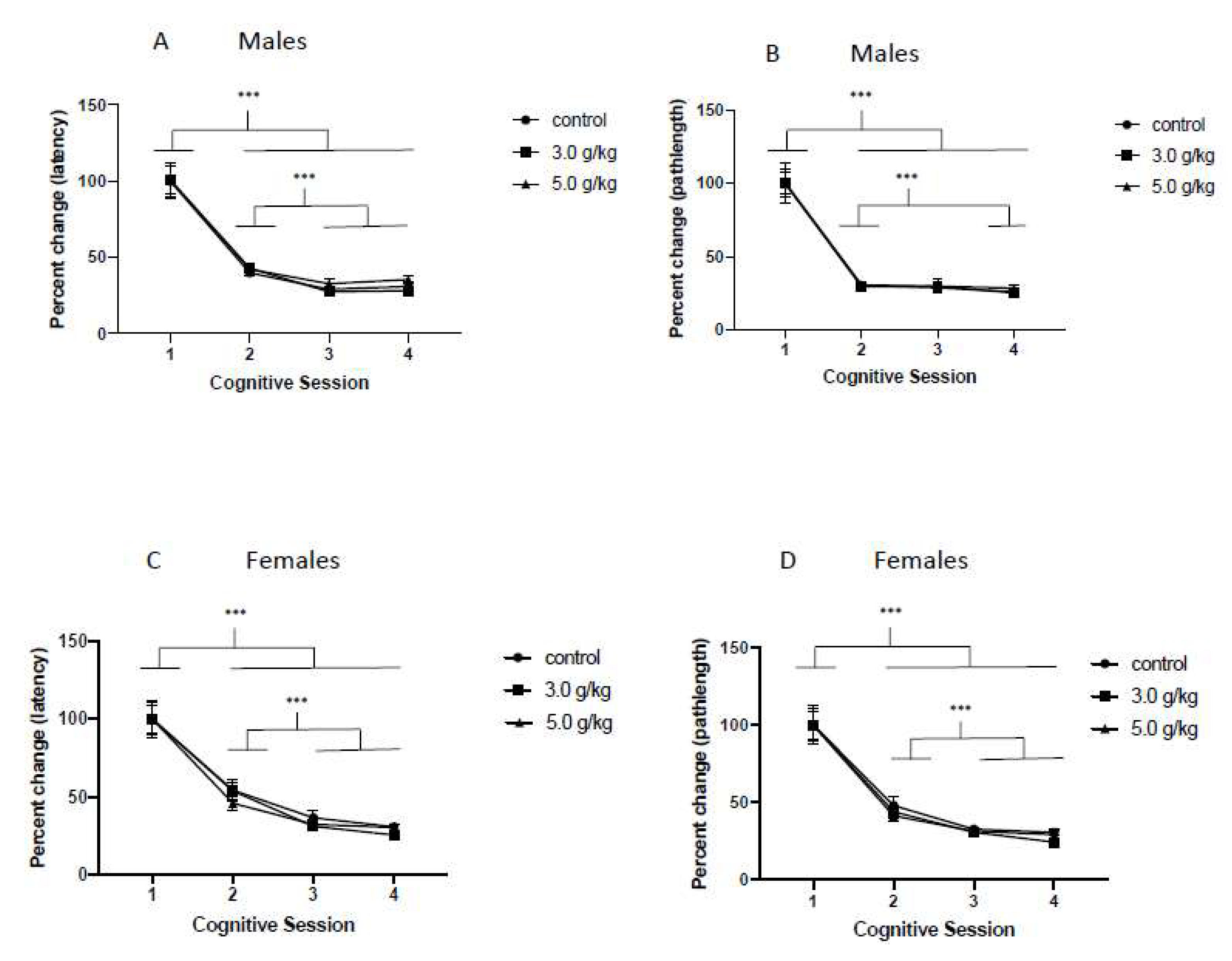
3.3. Impact of Chronic Intermittent Ethanol Exposure on Non-Spatial Learning
3.4. Impact of Chronic Intermittent Ethanol Exposure during Adolescence on Spatial Learning
3.5. Impact of Chronic Intermittent Ethanol Exposure during Adolescence on Probe Trials
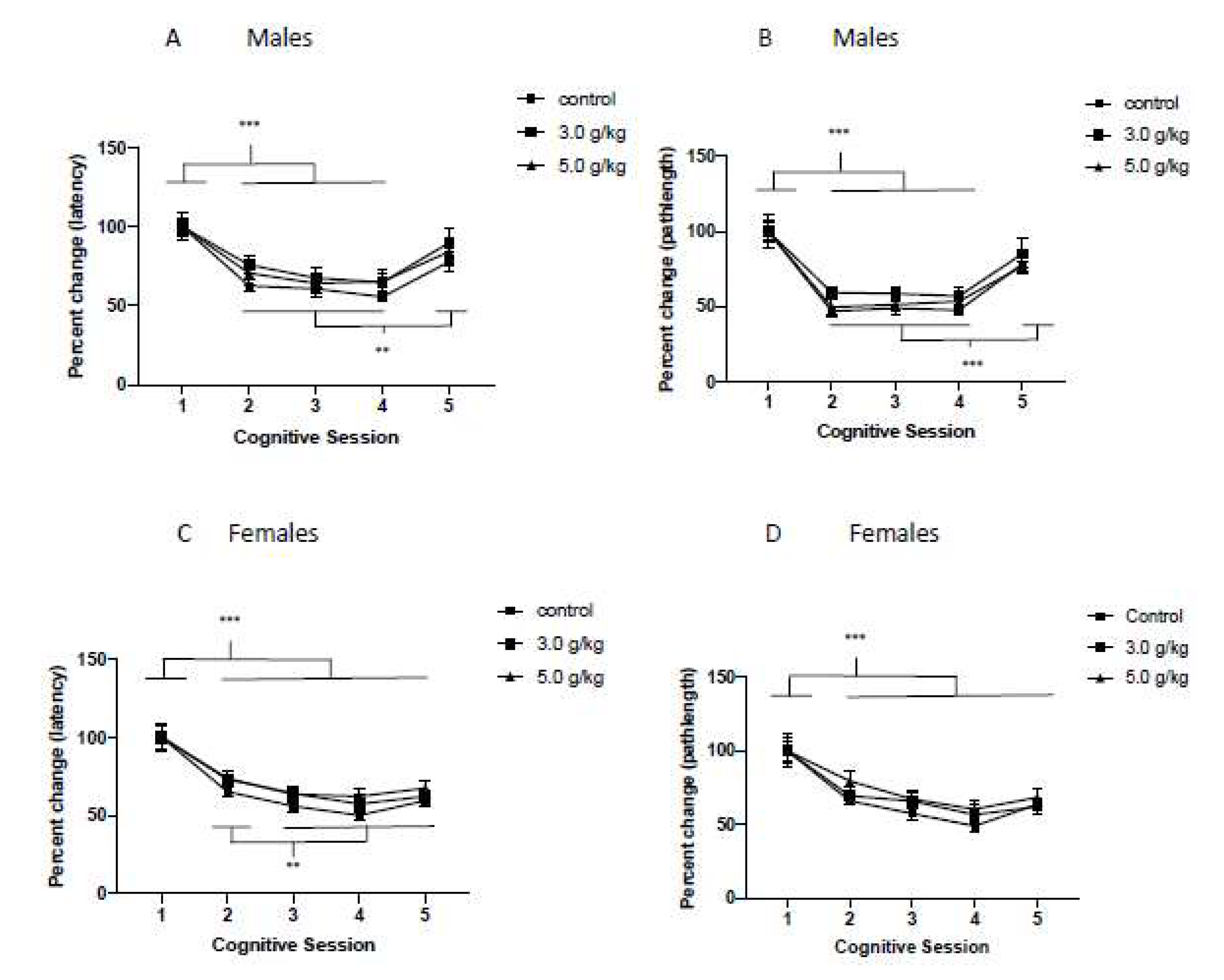
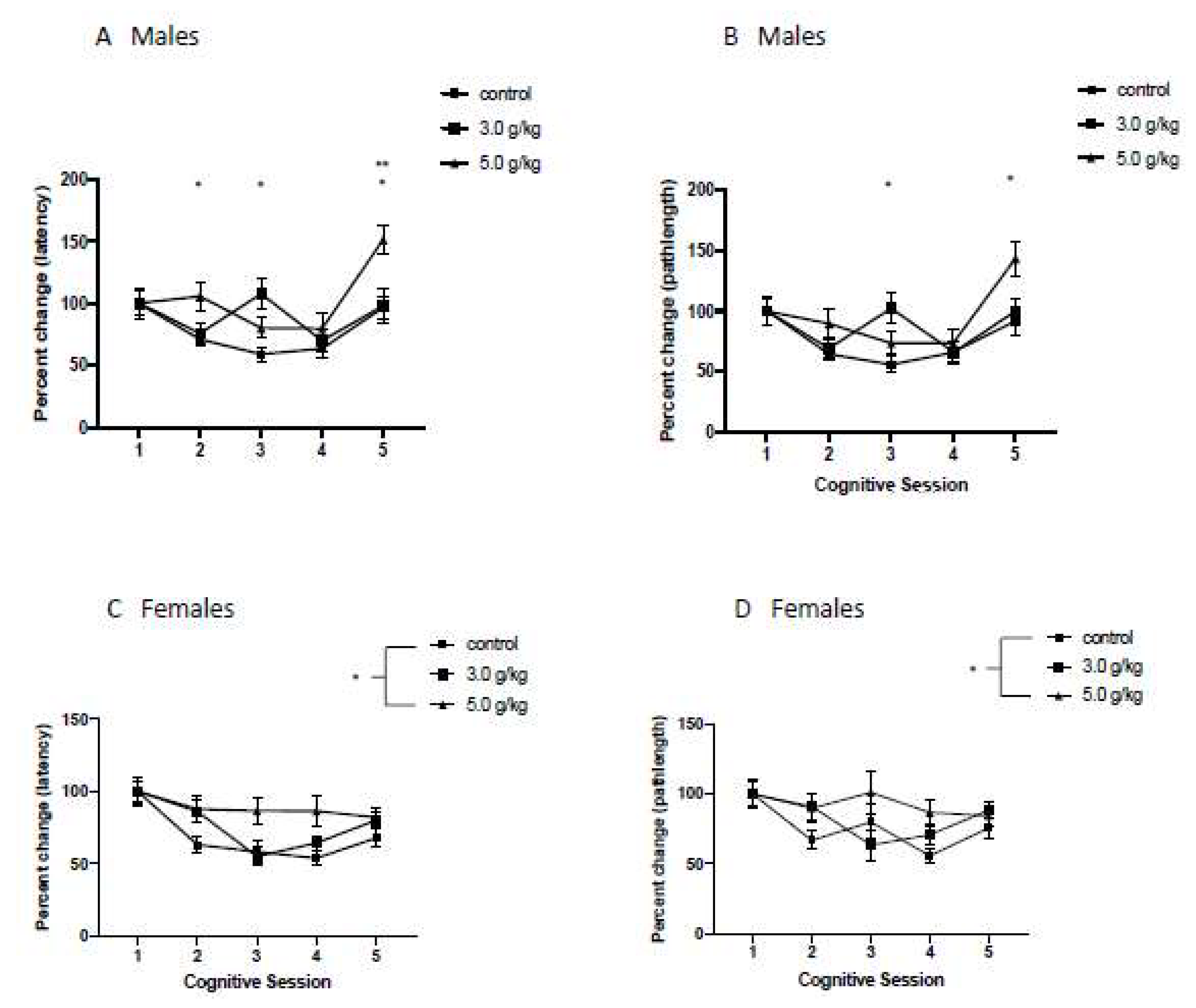
3.6. Impact of Chronic Intermittent Ethanol Exposure during Adolescence on Behavioral Flexibility
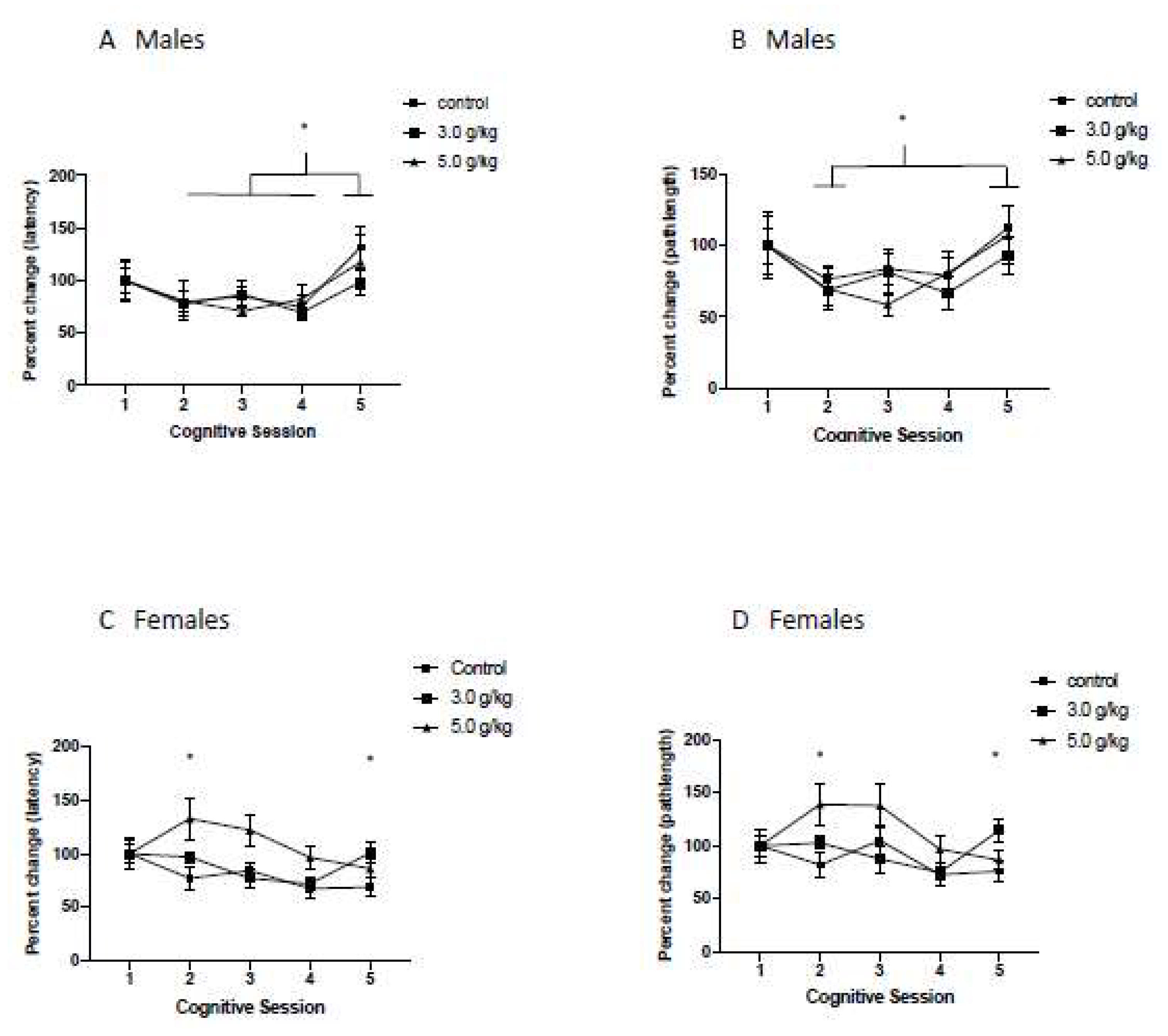
3.7. Impact of Chronic Intermittent Ethanol Exposure during Adolescence on Relearning
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferreira, M.P.; Willoughby, D. Alcohol consumption: The good, the bad, and the indifferent. Appl. Physiol. Nutr. Metab. 2008, 33, 12–20. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 Alcohol Collaborators. Alcohol use and burden for 195 countries and territories, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2018, 392, 1015–1035. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Health and Human Services SAaMHSA, Center for Behavioral health Statistics and Quality. National Survey on Drug Use and Health, 2019; U.S. Department of Health and Human Services SAaMHSA, Center for Behavioral Health Statistics and Quality: Bethesda, MD, USA, 2019. [Google Scholar]
- Haighton, C.; Wilson, G.; Ling, J.; McCabe, K.; Crosland, A.; Kaner, E. A Qualitative Study of Service Provision for Alcohol Related Health Issues in Mid to Later Life. PLoS ONE 2016, 11, e0148601. [Google Scholar] [CrossRef] [PubMed]
- Center for Disease Control and Prevention. Alcohol: Key Facts. In Fact Sheet; Center for Disease Control and Prevention: Atlanta, GA, USA, 2018; pp. 1–8. [Google Scholar]
- Varlinskaya, E.I.; Spear, L.P. Acute effects of ethanol on social behavior of adolescent and adult rats: Role of familiarity of the test situation. Alcohol. Clin. Exp. Res. 2002, 26, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.B.; Mittleman, G. Age-dependent effects of chronic intermittent ethanol treatment: Gross motor behavior and body weight in aged, adult and adolescent rats. Neurosci. Lett. 2017, 657, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, R.L.; Spear, L.P. Ontogeny of ethanol-induced motor impairment following acute ethanol: Assessment via the negative geotaxis reflex in adolescent and adult rats. Pharmacol. Biochem. Behav. 2010, 95, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Van Skike, C.E.; Botta, P.; Chin, V.S.; Tokunaga, S.; McDaniel, J.M.; Venard, J.; Diaz-Granados, J.L.; Valenzuela, C.F.; Matthews, D.B. Behavioral effects of ethanol in cerebellum are age dependent: Potential system and molecular mechanisms. Alcohol Clin. Exp. Res. 2010, 34, 2070–2080. [Google Scholar] [CrossRef] [Green Version]
- White, A.M.; Bae, J.G.; Truesdale, M.C.; Ahmad, S.; Wilson, W.A.; Swartzwelder, H.S. Chronic-intermittent ethanol exposure during adolescence prevents normal developmental changes in sensitivity to ethanol-induced motor impairments. Alcohol Clin. Exp. Res. 2002, 26, 960–968. [Google Scholar] [CrossRef]
- Ornelas, L.C.; Novier, A.; Van Skike, C.E.; Diaz-Granados, J.L.; Matthews, D.B. The effects of acute alcohol on motor impairments in adolescent, adult, and aged rats. Alcohol 2015, 49, 121–126. [Google Scholar] [CrossRef]
- Perkins, A.E.; Vore, A.S.; Lovelock, D.; Varlinskaya, E.; Deak, T. Late aging alters behavioral sensitivity to ethanol in a sex-specific manner in Fischer 344 rats. Pharmacol. Biochem. Behav. 2018, 175, 1–9. [Google Scholar] [CrossRef]
- Swartzwelder, H.S.; Richardson, R.C.; Markwiese-Foerch, B.; Wilson, W.A.; Little, P.J. Developmental differences in the acquisition of tolerance to ethanol. Alcohol 1998, 15, 311–314. [Google Scholar] [CrossRef]
- Silveri, M.M.; Spear, L.P. Ontogeny of ethanol elimination and ethanol-induced hypothermia. Alcohol 2000, 20, 45–53. [Google Scholar] [CrossRef]
- Watson, M.R.; James, K.; Mittleman, G.; Matthews, D.B. Impact of acute ethanol exposure on body temperatures in aged, adult and adolescent male rats. Alcohol 2020, 82, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.B.; Imhoff, B.M. Age modifies the effect of ethanol on behavior: Investigations in adolescent, adult and aged rats. Int. Rev. Neurobiol. 2021, 161, 251–275. [Google Scholar]
- Coleman, L.G.; Jr Liu, W.; Oguz, I.; Styner, M.; Crews, F.T. Adolescent binge ethanol treatment alters adult brain regional volumes, cortical extracellular matrix protein and behavioral flexibility. Pharmacol. Biochem. Behav. 2014, 116, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Kokare, D.M.; Kyzar, E.J.; Zhang, H.; Sakharkar, A.J.; Pandey, S.C. Adolescent Alcohol Exposure-Induced Changes in Alpha-Melanocyte Stimulating Hormone and Neuropeptide Y Pathways via Histone Acetylation in the Brain During Adulthood. Int. J. Neuropsychopharmacol. 2017, 20, 758–768. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Coehlo, M.A.; Solton, N.R.; Szumlinski, K.K. Negative Affect and Excessive Alcohol Intake Incubate during Protracted Withdrawal from Binge-Drinking in Adolescent, But Not Adult, Mice. Front. Psychol. 2017, 8, 1128. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.C.; Sakharkar, A.J.; Tang, L.; Zhang, H. Potential role of adolescent alcohol exposure-induced amygdaloid histone modifications in anxiety and alcohol intake during adulthood. Neurobiol. Dis. 2015, 82, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Van Skike, C.E.; Diaz-Granados, J.L.; Matthews, D.B. Chronic Intermittent Ethanol Exposure Produces Persistent Anxiety in Adolescent and Adult Rats. Alcohol. Clin. Exp. Res. 2015, 39, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Varlinskaya, E.I.; Truxell, E.M.; Spear, L.P. Ethanol intake under social circumstances or alone in sprague-dawley rats: Impact of age, sex, social activity, and social anxiety-like behavior. Alcohol Clin. Exp. Res. 2015, 39, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Vetreno, R.P.; Broadwater, M.; Liu, W.; Spear, L.P.; Crews, F.T. Adolescent, but not adult, binge ethanol exposure leads to persistent global reductions of choline acetyltransferase expressing neurons in brain. PLoS ONE 2014, 9, e113421. [Google Scholar] [CrossRef] [Green Version]
- Vetreno, R.P.; Yaxley, R.; Paniagua, B.; Crews, F.T. Diffusion tensor imaging reveals adolescent binge ethanol-induced brain structural integrity alterations in adult rats that correlate with behavioral dysfunction. Addict. Biol. 2016, 21, 939–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, D.B.; Tinsley, K.L.; Diaz-Granados, J.L.; Tokunaga, S.; Silvers, J.A. Chronic intermittent exposure to ethanol during adolescence produces tolerance to the hypnotic effects of ethanol in male rats: A dose-dependent analysis. Alcohol 2008, 42, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Silvers, J.M.; Tokunaga, S.; Mittleman, G.; Matthews, D.B. Chronic intermittent injections of high-dose ethanol during adolescence produce metabolic, hypnotic, and cognitive tolerance in rats. Alcohol Clin. Exp. Res. 2003, 27, 1606–1612. [Google Scholar] [CrossRef] [PubMed]
- Silvers, J.M.; Tokunaga, S.; Mittleman, G.; O’Buckley, T.; Morrow, A.L.; Matthews, D.B. Chronic intermittent ethanol exposure during adolescence reduces the effect of ethanol challenge on hippocampal allopregnanolone levels and Morris water maze task performance. Alcohol 2006, 39, 151–158. [Google Scholar] [CrossRef]
- Barker, J.M.; Bryant, K.G.; Osborne, J.I.; Chandler, L.J. Age and Sex Interact to Mediate the Effects of Intermittent, High-Dose Ethanol Exposure on Behavioral Flexibility. Front. Pharmacol. 2017, 8, 450. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, G.M.; Lew, B.J.; Vedder, L.C.; Savage, L.M. Chronic intermittent ethanol exposure leads to alterations in brain-derived neurotrophic factor within the frontal cortex and impaired behavioral flexibility in both adolescent and adult rats. Neuroscience 2017, 348, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Sey, N.Y.A.; Gomez, A.A.; Madayag, A.C.; Boettiger, C.A.; Robinson, D.L. Adolescent intermittent ethanol impairs behavioral flexibility in a rat foraging task in adulthood. Behav. Brain Res. 2019, 373, 112085. [Google Scholar] [CrossRef]
- Gass, J.T.; Glen, W.B.; Jr McGonigal, J.T.; Trantham-Davidson, H.; Lopez, M.F.; Randall, P.K.; Yaxley, R.; Floresco, S.B.; Chandler, L.J. Adolescent alcohol exposure reduces behavioral flexibility, promotes disinhibition, and increases resistance to extinction of ethanol self-administration in adulthood. Neuropsychopharmacology 2014, 39, 2570–2583. [Google Scholar] [CrossRef]
- Contreras, A.; Polin, E.; Miguens, M.; Perez-Garcia, C.; Perez, V.; Ruiz-Gayo, M.; Morales, L.; Del Olmo, N. Intermittent-Excessive and Chronic-Moderate Ethanol Intake during Adolescence Impair Spatial Learning, Memory and Cognitive Flexibility in the Adulthood. Neuroscience 2019, 418, 205–217. [Google Scholar] [CrossRef]
- Macht, V.; Elchert, N.; Crews, F. Adolescent Alcohol Exposure Produces Protracted Cognitive-Behavioral Impairments in Adult Male and Female Rats. Brain Sci. 2020, 10, 785. [Google Scholar] [CrossRef] [PubMed]
- Sjoerds, Z.; de Wit, S.; van den Brink, W.; Robbins, T.W.; Beekman, A.T.; Penninx, B.W.; Veltman, D.J. Behavioral and neuroimaging evidence for overreliance on habit learning in alcohol-dependent patients. Transl. Psychiatry 2013, 3, e337. [Google Scholar] [CrossRef] [PubMed]
- Shnitko, T.A.; Gonzales, S.W.; Grant, K.A. Low cognitive flexibility as a risk for heavy alcohol drinking in non-human primates. Alcohol 2019, 74, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Corbit, L.H.; Nie, H.; Janak, P.H. Habitual alcohol seeking: Time course and the contribution of subregions of the dorsal striatum. Biol. Psychiatry 2012, 72, 389–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangieri, R.A.; Cofresi, R.U.; Gonzales, R.A. Ethanol Seeking by Long Evans Rats Is Not Always a Goal-Directed Behavior. PLoS ONE 2012, 7, e42886. [Google Scholar] [CrossRef]
- Renteria, R.; Baltz, E.T.; Gremel, C.M. Chronic alcohol exposure disrupts top-down control over basal ganglia action selection to produce habits. Nat. Commun. 2018, 9, 211. [Google Scholar] [CrossRef] [Green Version]
- Sallet, J.; Noonan, M.P.; Thomas, A.; O’Reilly, J.X.; Anderson, J.; Papageorgiou, G.K.; Neubert, F.X.; Ahmed, B.; Smith, J.; Bell, A.H.; et al. Behavioral flexibility is associated with changes in structure and function distributed across a frontal cortical network in macaques. PLoS Biol. 2020, 18, e3000605. [Google Scholar] [CrossRef]
- Gremel, C.M.; Lovinger, D.M. Associative and sensorimotor cortico-basal ganglia circuit roles in effects of abused drugs. Genes Brain Behav. 2017, 16, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Van Skike, C.E.; Goodlett, C.; Matthews, D.B. Acute Alcohol and Cognition: Remembering What It Causes Us to Forget. Alcohol 2019, 79, 105–125. [Google Scholar] [CrossRef]
- Scheidt, L.; Fries, G.R.; Stertz, L.; Cabral, J.C.; Kapczinski, F.; de Almeida, R.M. Ethanol during adolescence decreased the BDNF levels in the hippocampus in adult male Wistar rats, but did not alter aggressive and anxiety-like behaviors. Trends Psychiatry Psychother. 2015, 37, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Vetreno, R.P.; Crews, F.T. Binge ethanol exposure during adolescence leads to a persistent loss of neurogenesis in the dorsal and ventral hippocampus that is associated with impaired adult cognitive functioning. Front. Neurosci. 2015, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, D.B.; Novier, A.; Diaz-Granados, J.L.; Van Skike, C.E.; Ornelas, L.; Mittleman, G. Impact of adolescent alcohol use across the lifespan: Long-lasting tolerance to high-dose alcohol coupled with potentiated spatial memory impairments to moderate-dose alcohol. Alcohol 2017, 61, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Risher, M.L.; Fleming, R.L.; Risher, W.C.; Miller, K.M.; Klein, R.C.; Wills, T.; Acheson, S.K.; Moore, S.D.; Wilson, W.A.; Eroglu, C.; et al. Adolescent Intermittent Alcohol Exposure: Persistence of Structural and Functional Hippocampal Abnormalities into Adulthood. Alcohol. Clin. Exp. Res. 2015, 39, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.M.; Ehlers, C.L. Appetitive motivational experience during adolescence results in enhanced alcohol consumption during adulthood. Behav. Neurosci. 2009, 123, 926–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, R.G.M.; Garrud, P.; Rawlins, J.N.P.; Okeefe, J. Place Navigation Impaired in Rats with Hippocampal-Lesions. Nature 1982, 297, 681–683. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Value of water mazes for assessing spatial and egocentric learning and memory in rodent basic research and regulatory studies. Neurotoxicol. Teratol. 2014, 45, 75–90. [Google Scholar] [CrossRef]
- Barton, E.A.; Baker, C.; Leasure, J.L. Investigation of Sex Differences in the Microglial Response to Binge Ethanol and Exercise. Brain Sci. 2017, 7, 139. [Google Scholar] [CrossRef] [Green Version]
- Beaudet, G.; Valable, S.; Bourgine, J.; Lelong-Boulouard, V.; Lanfumey, L.; Freret, T.; Boulouard, M.; Paizanis, E. Long-Lasting Effects of Chronic Intermittent Alcohol Exposure in Adolescent Mice on Object Recognition and Hippocampal Neuronal Activity. Alcohol Clin. Exp. Res. 2016, 40, 2591–2603. [Google Scholar] [CrossRef]
- Becker, J.B.; Koob, G.F. Sex Differences in Animal Models: Focus on Addiction. Pharmacol. Rev. 2016, 68, 242–263. [Google Scholar] [CrossRef] [Green Version]
- Goodman, J.; Packard, M.G. Memory Systems and the Addicted Brain. Front. Psychiatry 2016, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Packard, M.G.; Hirsh, R.; White, N.M. Differential-Effects of Fornix and Caudate-Nucleus Lesions on 2 Radial Maze Tasks—Evidence for Multiple Memory-Systems. J. Neurosci. 1989, 9, 1465–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, C.N.; Gremel, C.M. Review of Orbitofrontal Cortex in Alcohol Dependence: A Disrupted Cognitive Map? Alcohol Clin. Exp. Res. 2020, 44, 1952–1964. [Google Scholar] [CrossRef] [PubMed]
- Shnitko, T.A.; Gonzales, S.W.; Newman, N.; Grant, K.A. Behavioral Flexibility in Alcohol-Drinking Monkeys: The Morning After. Alcohol Clin. Exp. Res. 2020, 44, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Santin, L.J.; Rubio, S.; Begega, A.; Arias, J.L. Effects of chronic alcohol consumption on spatial reference and working memory tasks. Alcohol 2000, 20, 149–159. [Google Scholar] [CrossRef]
- Crews, F.T.; Vetreno, R.P.; Broadwater, M.A.; Robinson, D.L. Adolescent Alcohol Exposure Persistently Impacts Adult Neurobiology and Behavior. Pharmacol. Rev. 2016, 68, 1074–1109. [Google Scholar] [CrossRef] [Green Version]
- De Falco, E.; White, S.M.; Morningstar, M.D.; Ma, B.; Nkurunziza, L.T.; Ahmed-Dilibe, A.; Wellman, C.L.; Lapish, C.C. Impaired cognitive flexibility and heightened urgency are associated with increased alcohol consumption in rodent models of excessive drinking. Addict. Biol. 2021, 26, e13004. [Google Scholar] [CrossRef]
- Flores-Bonilla, A.; Richardson, H.N. Sex Differences in the Neurobiology of Alcohol Use Disorder. Alcohol Res. 2020, 40, 04. [Google Scholar] [CrossRef]
- Finn, D.A.; Sinnott, R.S.; Ford, M.M.; Long, S.L.; Tanchuck, M.A.; Phillips, T.J. Sex differences in the effect of ethanol injection and consumption on brain allopregnanolone levels in C57BL/6 mice. Neuroscience 2004, 123, 813–819. [Google Scholar] [CrossRef]
- VanDoren, M.J.; Matthews, D.B.; Janis, G.C.; Grobin, A.C.; Devaud, L.L.; Morrow, A.L. Neuroactive steroid 3alpha-hydroxy-5alpha-pregnan-20-one modulates electrophysiological and behavioral actions of ethanol. J. Neurosci. 2000, 20, 1982–1989. [Google Scholar] [CrossRef] [Green Version]
- Chin, V.S.; Van Skike, C.E.; Berry, R.B.; Kirk, R.E.; Diaz-Granados, J.; Matthews, D.B. Effect of acute ethanol and acute allopregnanolone on spatial memory in adolescent and adult rats. Alcohol 2011, 45, 473–483. [Google Scholar] [CrossRef]
- Matthews, D.B.; Morrow, A.L.; Tokunaga, S.; McDaniel, J.R. Acute ethanol administration and acute allopregnanolone administration impair spatial memory in the Morris water task. Alcohol. Clin. Exp. Res. 2002, 26, 1747–1751. [Google Scholar] [CrossRef] [PubMed]
- Sharrett-Field, L.; Butler, T.R.; Reynolds, A.R.; Berry, J.N.; Prendergast, M.A. Sex differences in neuroadaptation to alcohol and withdrawal neurotoxicity. Pflugers. Arch. 2013, 465, 643–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salling, M.C.; Skelly, M.J.; Avegno, E.; Regan, S.; Zeric, T.; Nichols, E.; Harrison, N.L. Alcohol Consumption during Adolescence in a Mouse Model of Binge Drinking Alters the Intrinsic Excitability and Function of the Prefrontal Cortex through a Reduction in the Hyperpolarization-Activated Cation Current. J. Neurosci. 2018, 38, 6207–6222. [Google Scholar] [CrossRef] [PubMed]
- White, A.M.; Ghia, A.J.; Levin, E.D.; Swartzwelder, H.S. Binge pattern ethanol exposure in adolescent and adult rats: Differential impact on subsequent responsiveness to ethanol. Alcohol Clin. Exp. Res. 2000, 24, 1251–1256. [Google Scholar] [CrossRef]
- Squeglia, L.M.; Schweinsburg, A.D.; Pulido, C.; Tapert, S.F. Adolescent binge drinking linked to abnormal spatial working memory brain activation: Differential gender effects. Alcohol Clin. Exp. Res. 2011, 35, 1831–1841. [Google Scholar] [CrossRef]
- Squeglia, L.M.; Spadoni, A.D.; Infante, M.A.; Myers, M.G.; Tapert, S.F. Initiating moderate to heavy alcohol use predicts changes in neuropsychological functioning for adolescent girls and boys. Psychol. Addict. Behav. 2009, 23, 715–722. [Google Scholar] [CrossRef] [Green Version]
- McQuail, J.A.; Dunn, A.R.; Stern, Y.; Barnes, C.A.; Kempermann, G.; Rapp, P.R.; Kaczorowski, C.C.; Foster, T.C. Cognitive reserve in model systems for mechanistic discovery: The importance of longitudinal studies. Front. Aging Neurosci. 2021, 12, 607685. [Google Scholar] [CrossRef]
- Huurre, T.; Lintonen, T.; Kaprio, J.; Pelkonen, M.; Marttunen, M.; Aro, H. Adolescent risk factors for excessive alcohol use at age 32 years. A 16-year prospective follow-up study. Soc. Psychiatry Psychiatr. Epidemiol. 2010, 45, 125–134. [Google Scholar] [CrossRef]
- Towner, T.T.; Varlinskaya, E.I. Adolescent Ethanol Exposure: Anxiety-Like Behavioral Alterations, Ethanol Intake, and Sensitivity. Front Behav. Neurosci. 2020, 14, 45. [Google Scholar] [CrossRef]
- Amodeo, L.R.; Wills, D.N.; Sanchez-Alavez, M.; Nguyen, W.; Conti, B.; Ehlers, C.L. Intermittent voluntary ethanol consumption combined with ethanol vapor exposure during adolescence increases drinking and alters other behaviors in adulthood in female and male rats. Alcohol 2018, 73, 57–66. [Google Scholar] [CrossRef]
- Broadwater, M.; Spear, L.P. Consequences of ethanol exposure on cued and contextual fear conditioning and extinction differ depending on timing of exposure during adolescence or adulthood. Behav. Brain Res. 2013, 256, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodd-Henricks, Z.A.; Bell, R.L.; Kuc, K.A.; Murphy, J.M.; McBride, W.J.; Lumeng, L.; Li, T.K. Effects of ethanol exposure on subsequent acquisition and extinction of ethanol self-administration and expression of alcohol-seeking behavior in adult alcohol-preferring (P) rats: II. Adult exposure. Alcohol Clin. Exp. Res. 2002, 26, 1642–1652. [Google Scholar] [CrossRef] [PubMed]
- Toalston, J.E.; Deehan, G.A.; Jr Hauser, S.R.; Engleman, E.A.; Bell, R.L.; Murphy, J.M.; McBride, W.J.; Rodd, Z.A. The reinforcing properties of ethanol are quantitatively enhanced in adulthood by peri-adolescent ethanol, but not saccharin, consumption in female alcohol-preferring (P) rats. Alcohol 2015, 49, 513–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyrer-Jackson, J.M.; Hood, L.E.; Olive, M.F. Drugs of Abuse Differentially Alter the Neuronal Excitability of Prefrontal Layer V Pyramidal Cell Subtypes. Front. Cell Neurosci. 2021, 15, 703655. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Cabrera-Garcia, D.; Salling, M.C.; Au, E.; Yang, G.; Harrison, N.L. Alcohol reduces the activity of somatostatin interneurons in the mouse prefrontal cortex: A neural basis for its disinhibitory effect? Neuropharmacology 2021, 188, 108501. [Google Scholar] [CrossRef]
- Nimitvilai, S.; Lopez, M.F.; Mulholland, P.J.; Woodward, J.J. Chronic Intermittent Ethanol Exposure Enhances the Excitability and Synaptic Plasticity of Lateral Orbitofrontal Cortex Neurons and Induces a Tolerance to the Acute Inhibitory Actions of Ethanol. Neuropsychopharmacology 2016, 41, 1112–1127. [Google Scholar] [CrossRef] [Green Version]
- Hughes, B.A.; Crofton, E.J.; O’Buckley, T.K.; Herman, M.A.; Morrow, A.L. Chronic ethanol exposure alters prelimbic prefrontal cortical Fast-Spiking and Martinotti interneuron function with differential sex specificity in rat brain. Neuropharmacology 2020, 162, 107805. [Google Scholar] [CrossRef]
- McGuier, N.S.; Padula, A.E.; Lopez, M.F.; Woodward, J.J.; Mulholland, P.J. Withdrawal from chronic intermittent alcohol exposure increases dendritic spine density in the lateral orbitofrontal cortex of mice. Alcohol 2015, 49, 21–27. [Google Scholar] [CrossRef] [Green Version]
- White, A.M. Gender Differences in the Epidemiology of Alcohol Use and Related Harms in the United States. Alcohol. Res. 2020, 40, 01. [Google Scholar] [CrossRef]
- Novier, A.; Van Skike, C.E.; Diaz-Granados, J.L.; Mittleman, G.; Matthews, D.B. Acute alcohol produces ataxia and cognitive impairments in aged animals: A comparison between young adult and aged rats. Alcohol Clin. Exp. Res. 2013, 37, 1317–1324. [Google Scholar] [CrossRef]
- Matthews, D.B.; Scaletty, S.; Schreiber, A.; Trapp, S. Acute ethanol administration produces larger spatial and nonspatial memory impairments in 29-33 month old rats compared to adult and 18-24 month old rats. Pharmacol. Biochem. Behav. 2020, 199, 173074. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.B.; Watson, M.R.; James, K.; Kastner, A.; Schneider, A.; Mittleman, G. The impact of low to moderate chronic intermittent ethanol exposure on behavioral endpoints in aged, adult, and adolescent rats. Alcohol 2019, 78, 33–42. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matthews, D.B.; Scaletty, S.; Trapp, S.; Kastner, A.; Schneider, A.M.; Schreiber, A.; Rossmann, G. Chronic Intermittent Ethanol Administration during Adolescence Produces Sex Dependent Impairments in Behavioral Flexibility and Survivability. Brain Sci. 2022, 12, 606. https://doi.org/10.3390/brainsci12050606
Matthews DB, Scaletty S, Trapp S, Kastner A, Schneider AM, Schreiber A, Rossmann G. Chronic Intermittent Ethanol Administration during Adolescence Produces Sex Dependent Impairments in Behavioral Flexibility and Survivability. Brain Sciences. 2022; 12(5):606. https://doi.org/10.3390/brainsci12050606
Chicago/Turabian StyleMatthews, Douglas B., Samantha Scaletty, Sarah Trapp, Abigail Kastner, Amelia M. Schneider, Areonna Schreiber, and Gillian Rossmann. 2022. "Chronic Intermittent Ethanol Administration during Adolescence Produces Sex Dependent Impairments in Behavioral Flexibility and Survivability" Brain Sciences 12, no. 5: 606. https://doi.org/10.3390/brainsci12050606