
Low-Intensity Focused Ultrasound-Mediated Attenuation of Acute Seizure Activity Based on EEG Brain Functional Connectivity

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
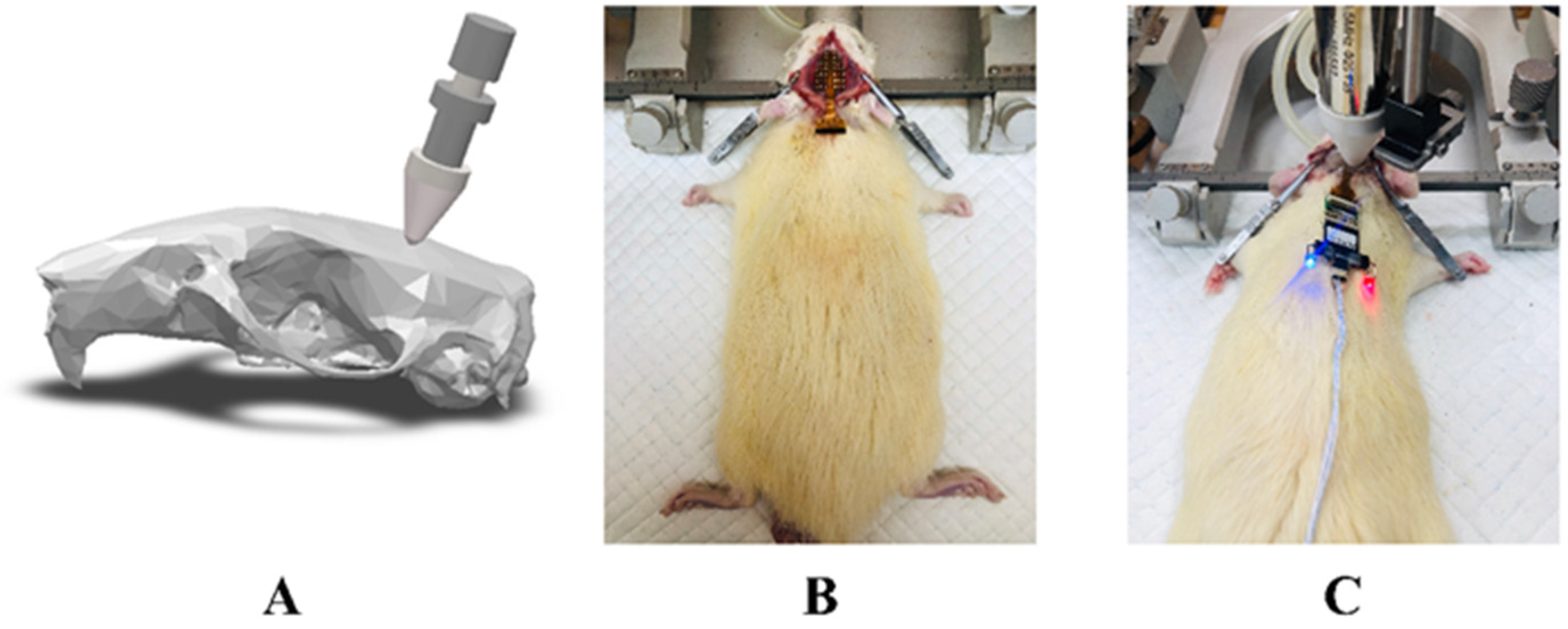
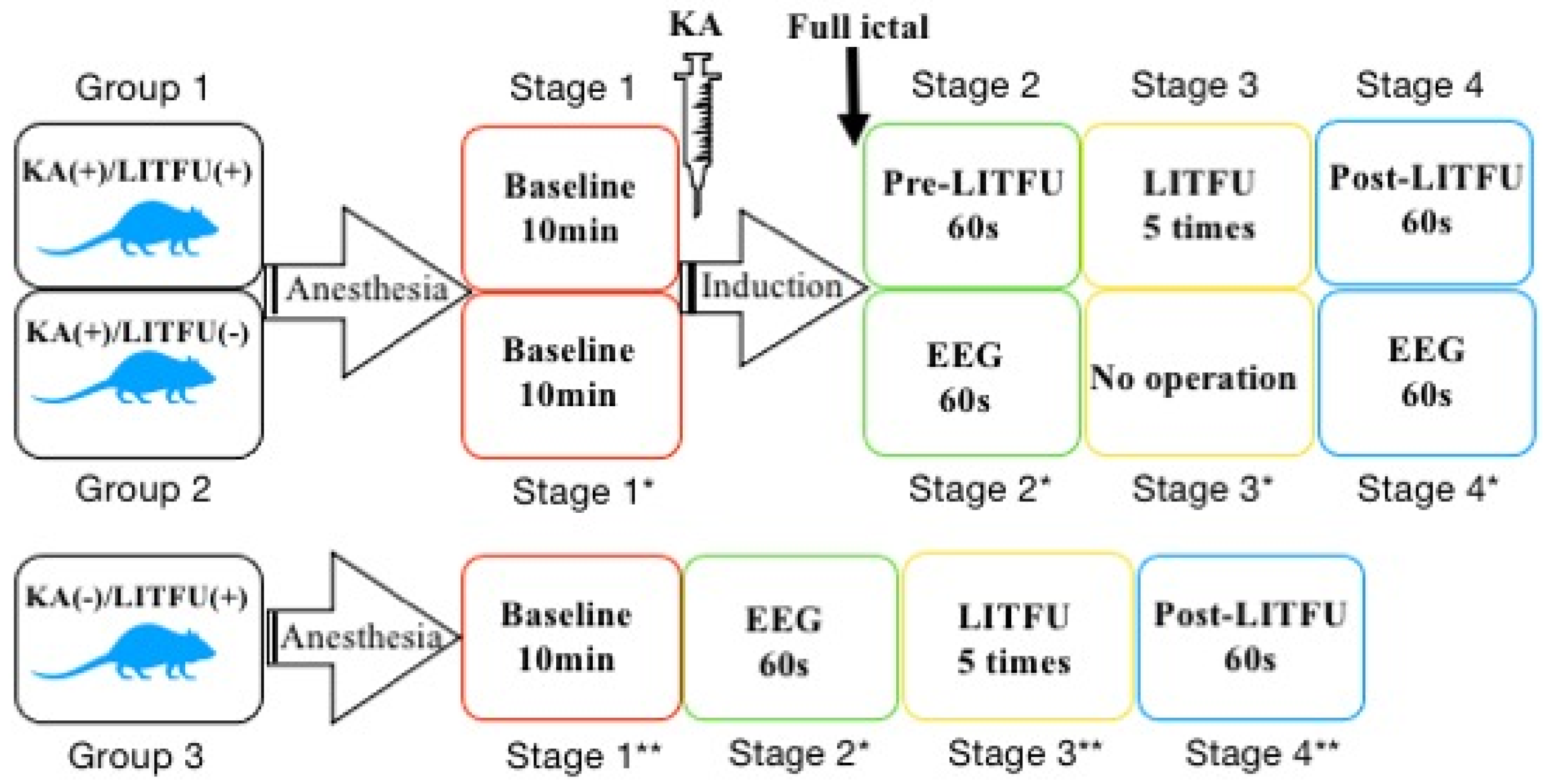
2.1. Animal Preparation and Grouping
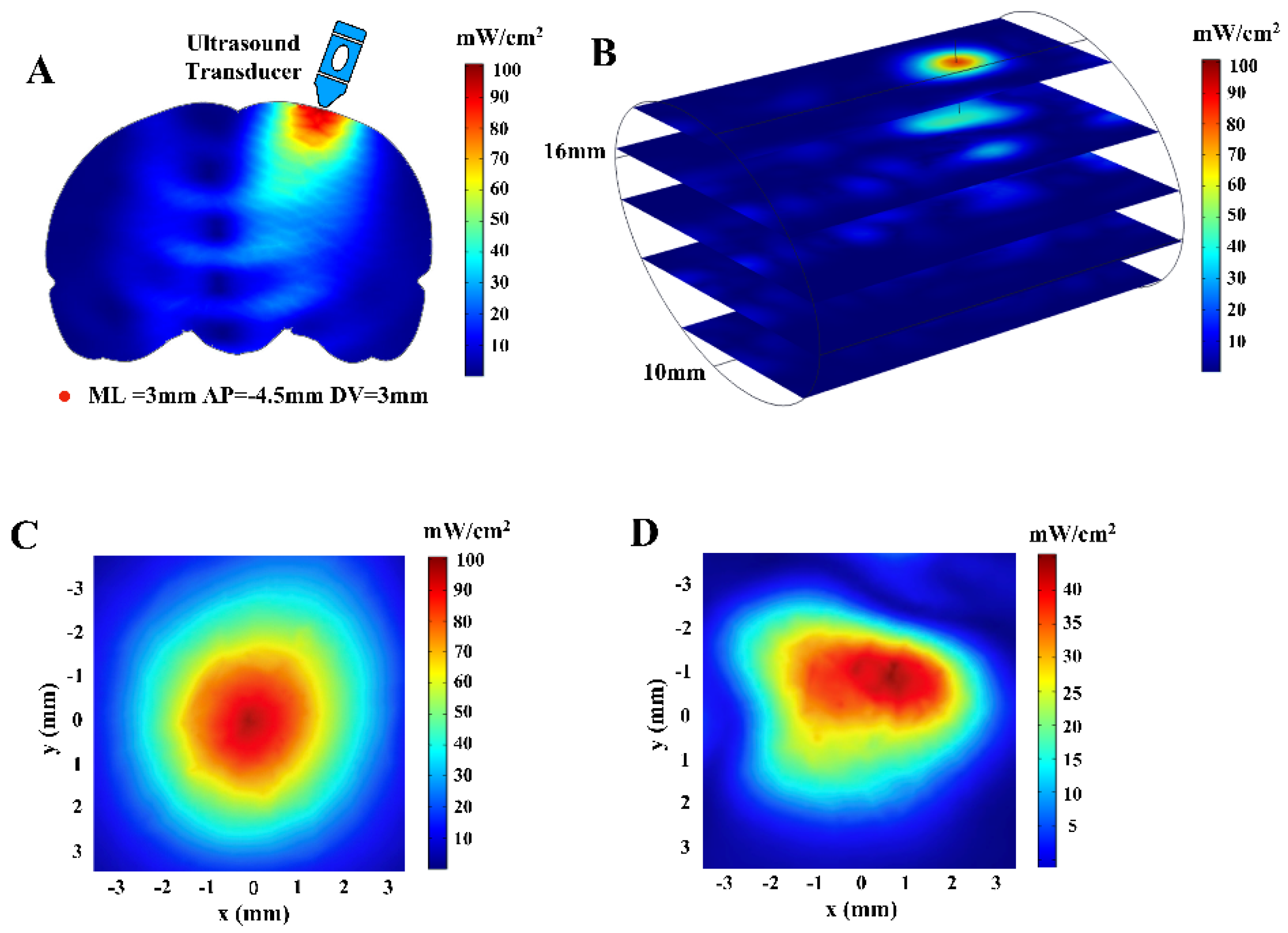
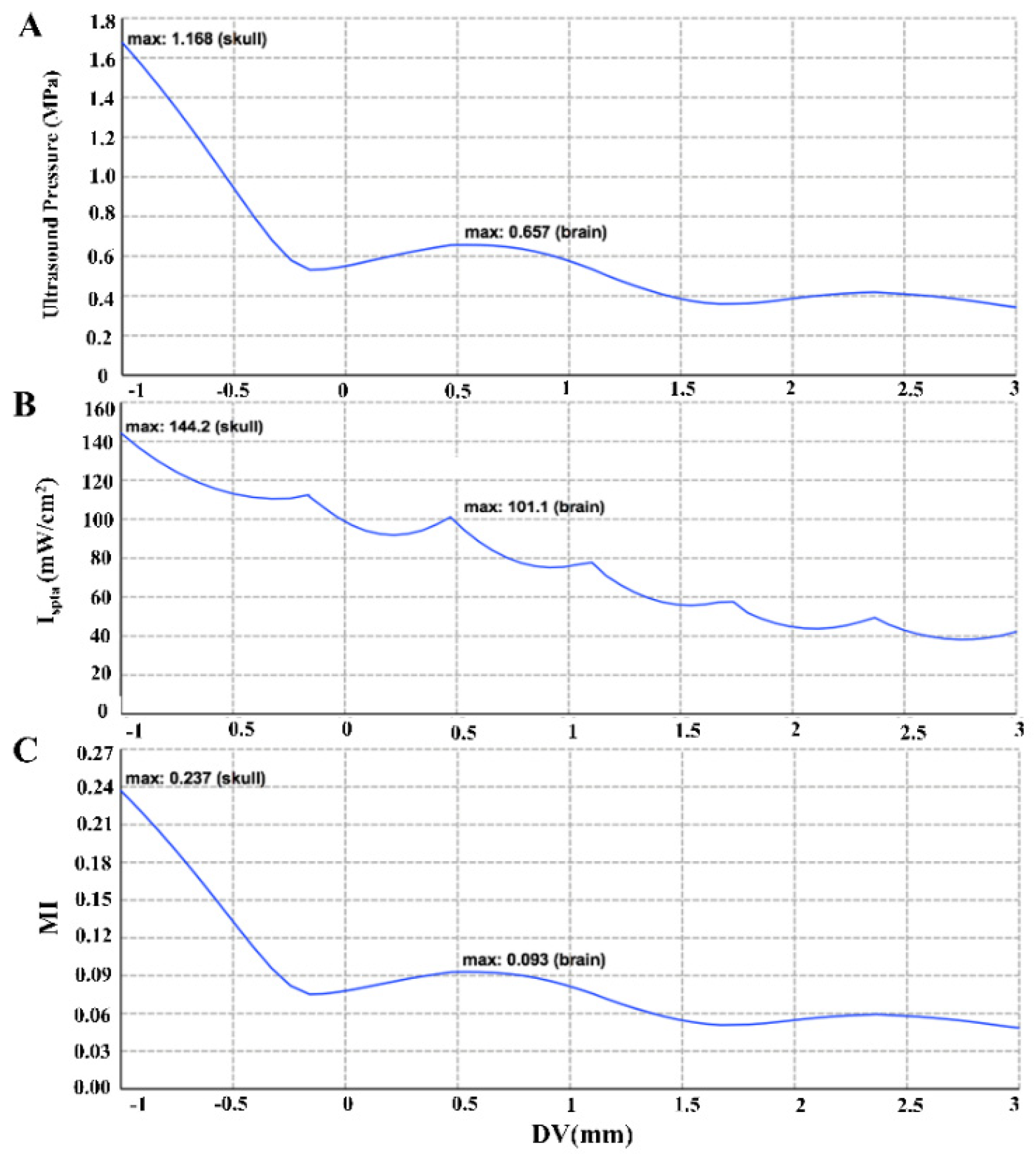
2.2. Computational Simulation Modeling and Ultrasonic Field Distribution
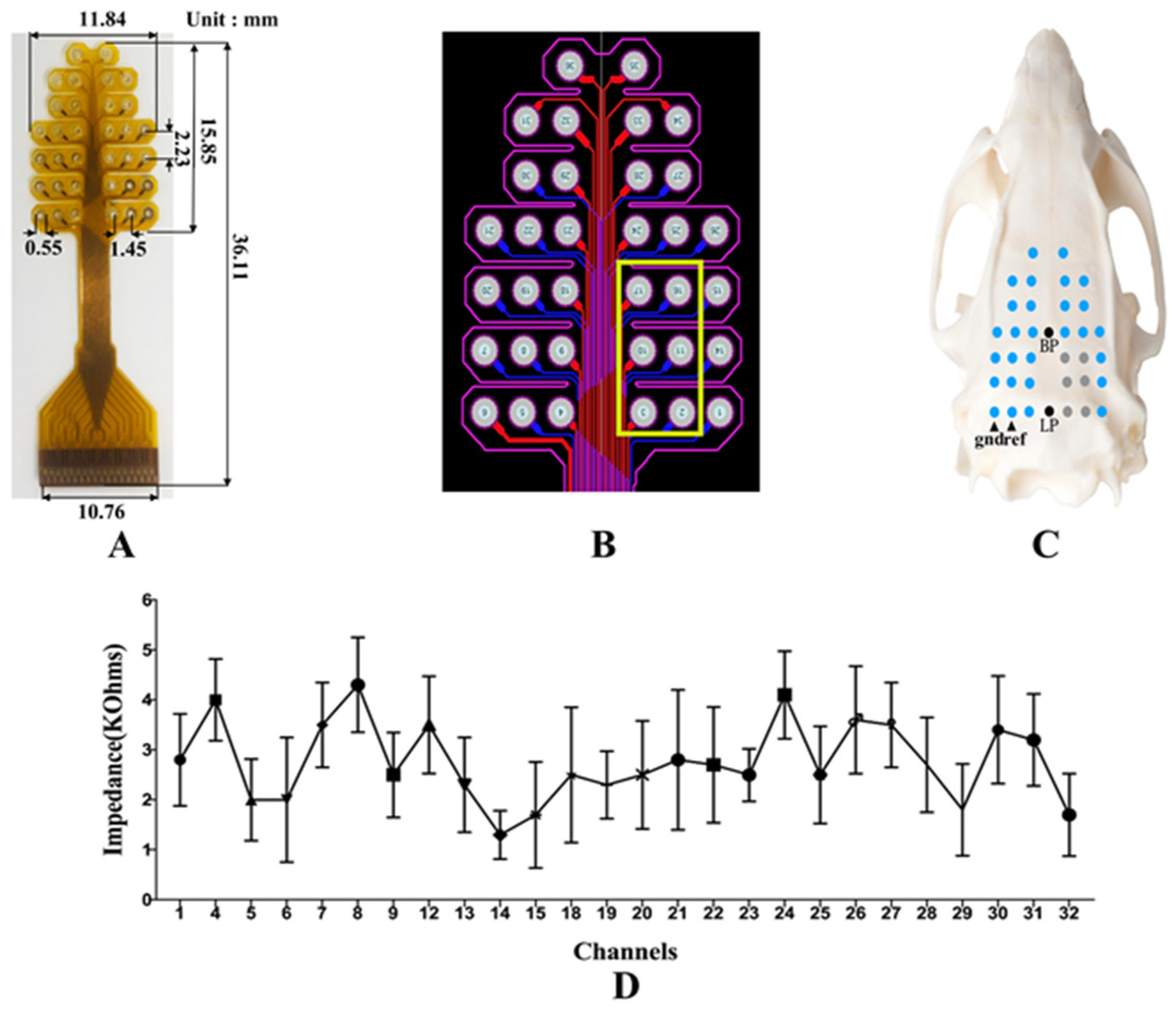
2.3. Electrode Positioning and Implantation
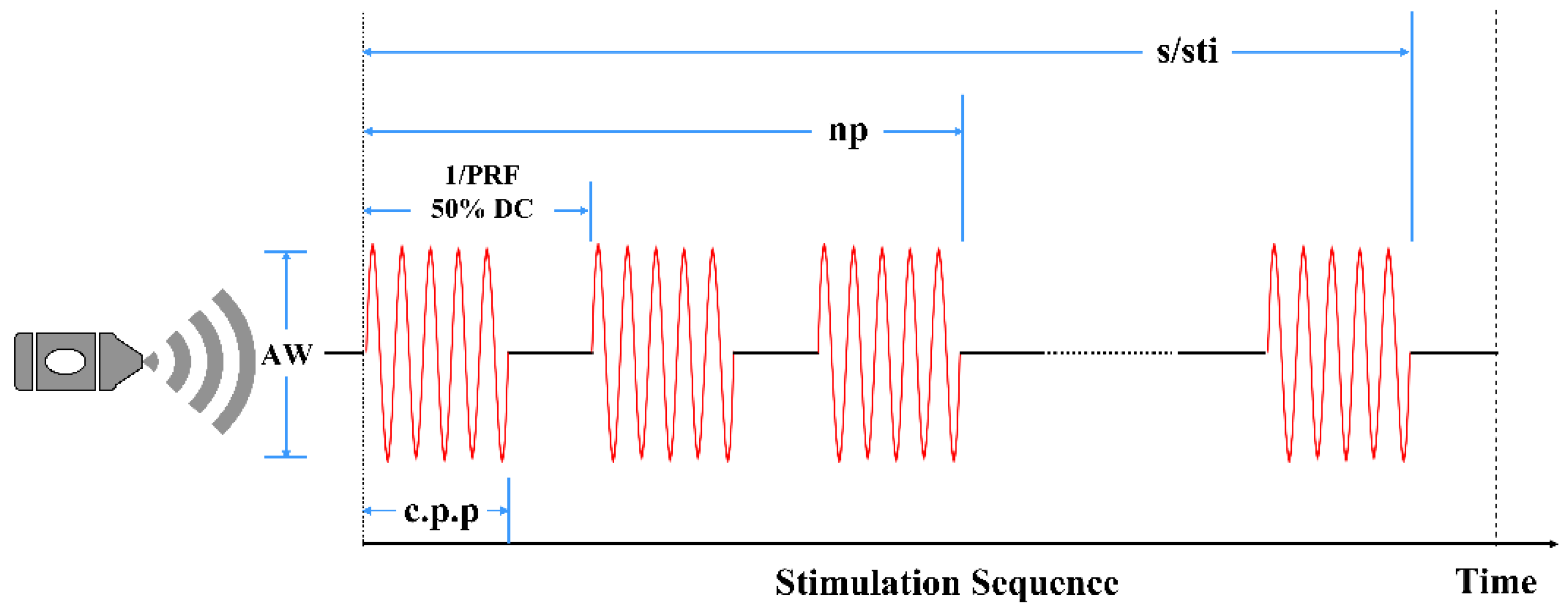
2.4. LIFU Sonication and Induction of Epilepsy
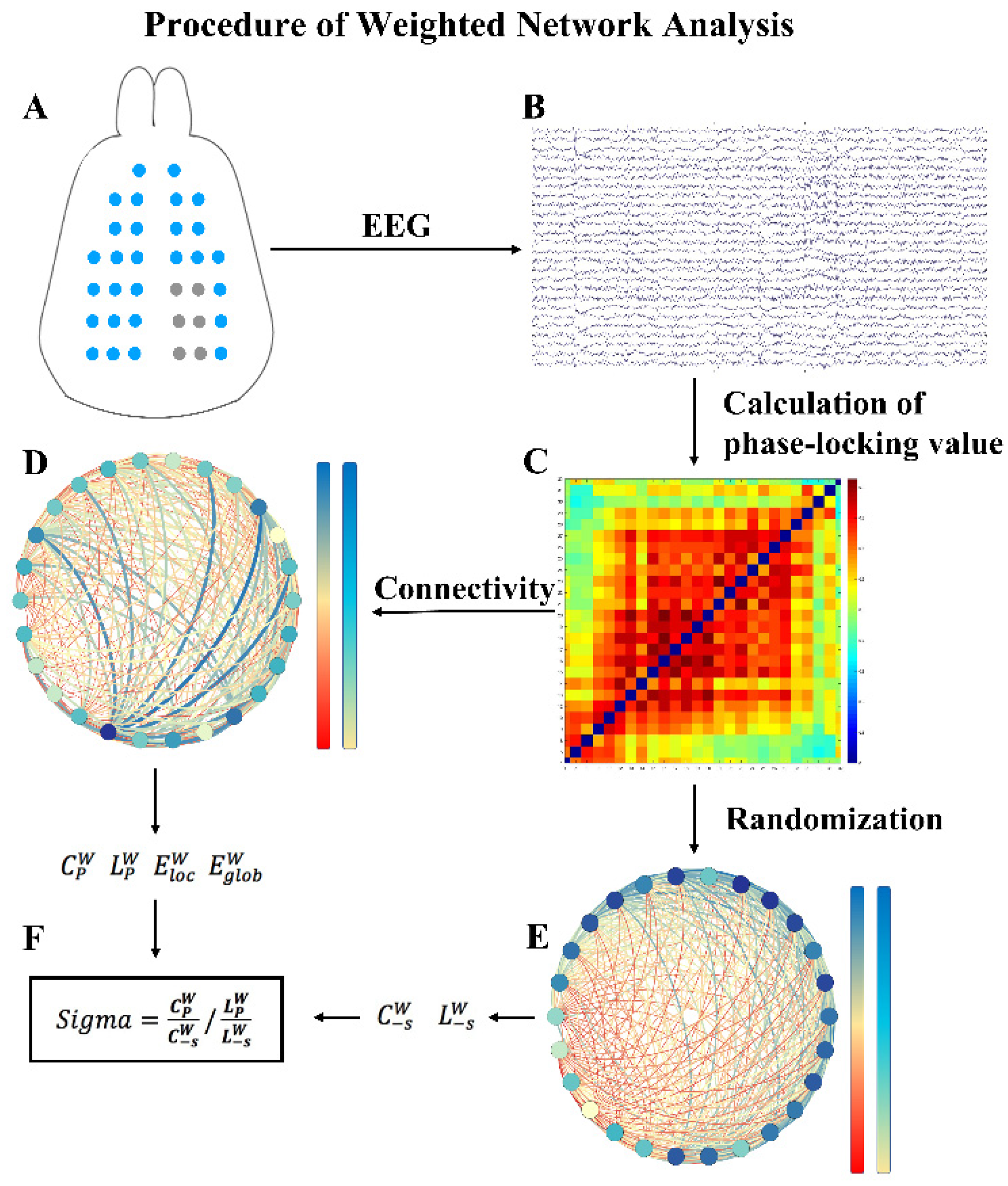
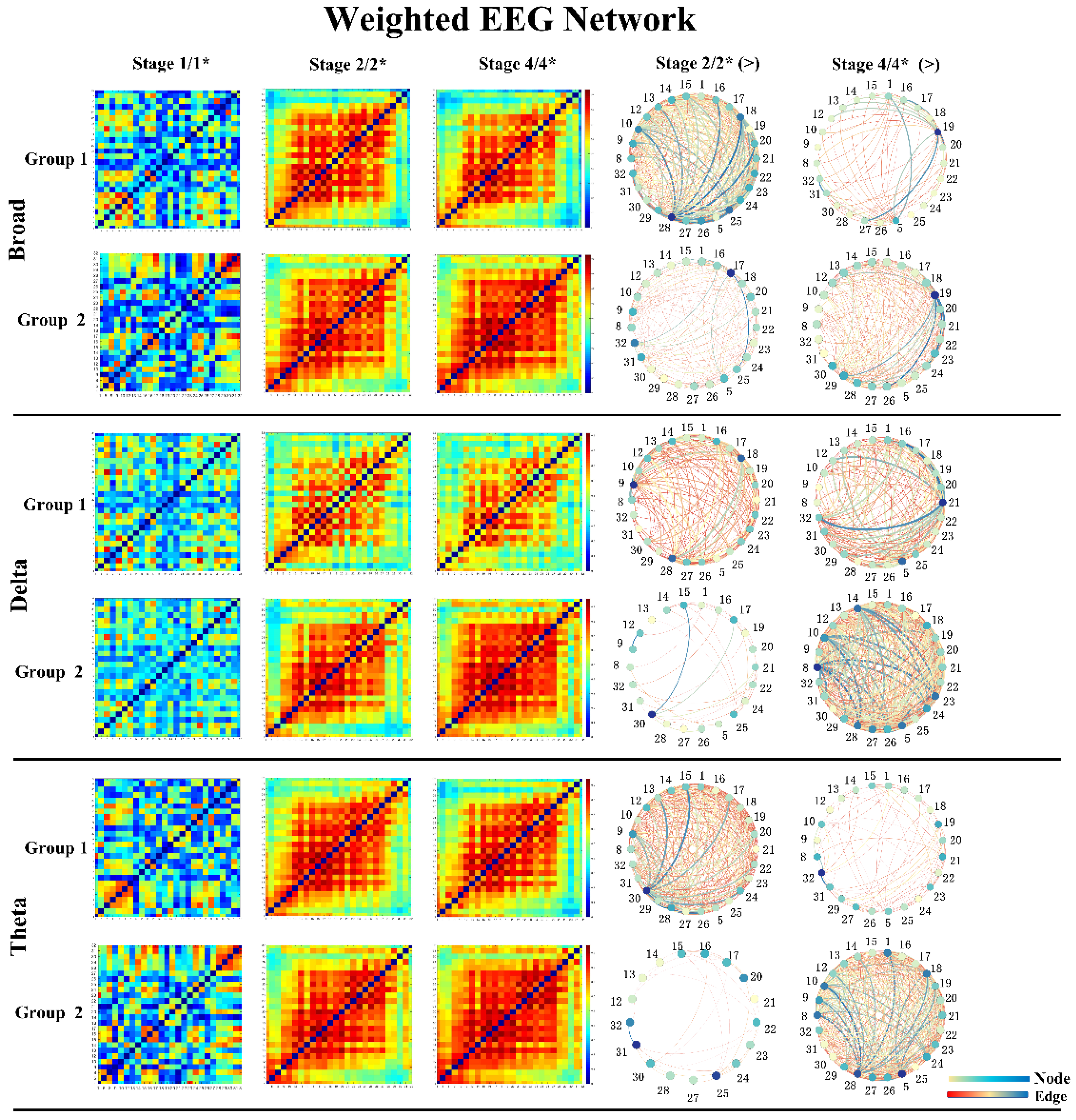
2.5. Brain Network Construction and Graph Theoretical Analysis
3. Results
3.1. Simulated Sound Field Distribution and Line Graph
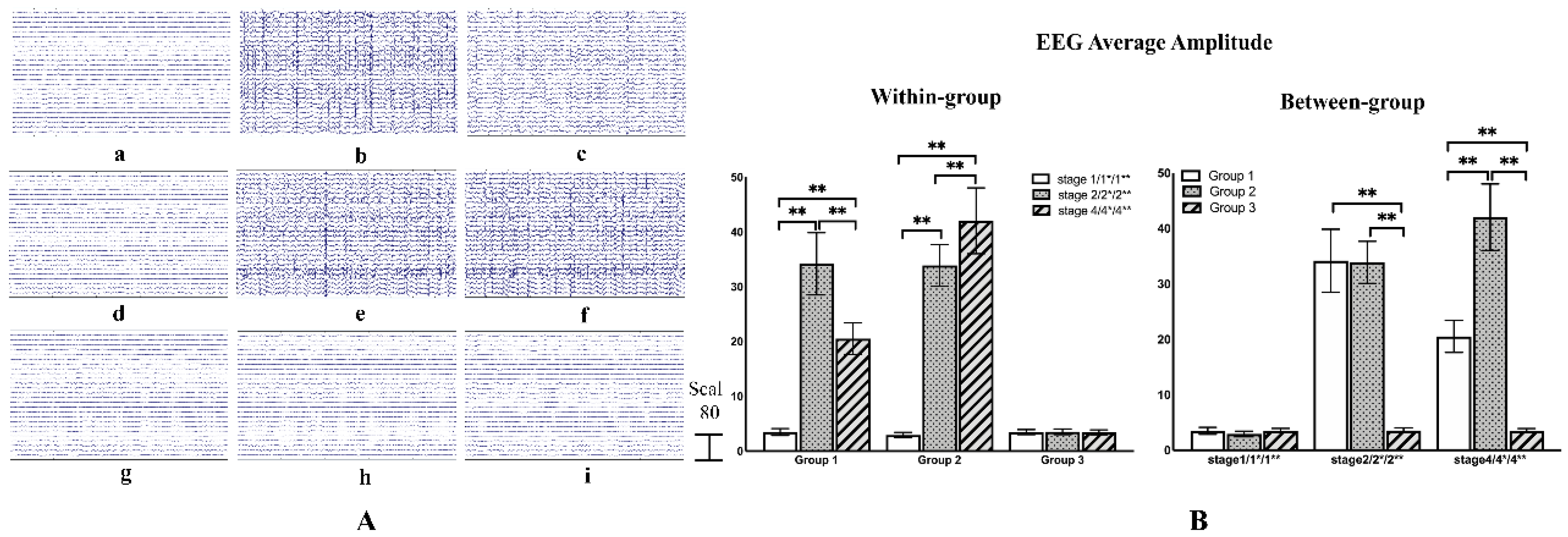
3.2. Epileptic Raw EEG Analysis
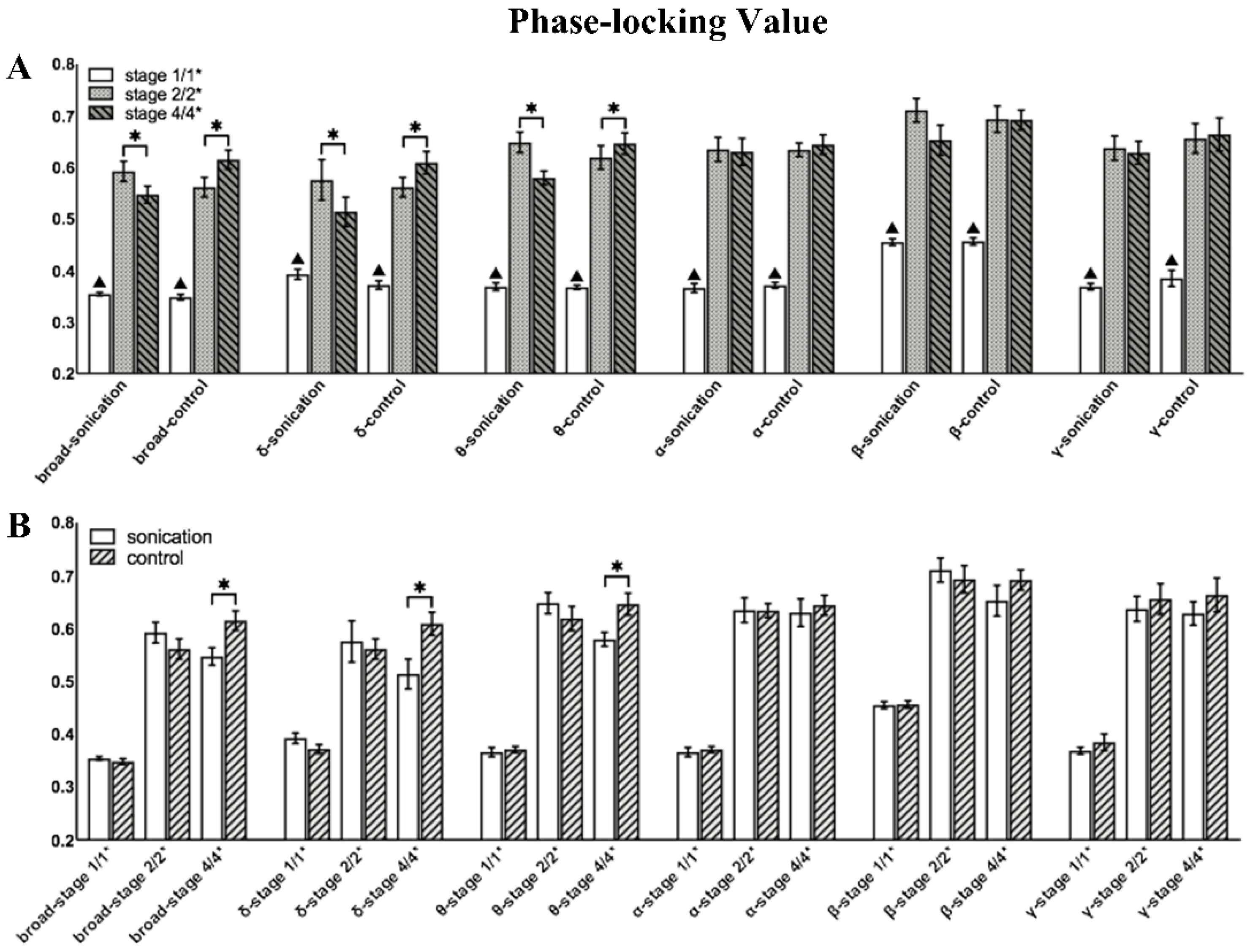
3.3. Phase-Locking Value Changes before and after Sonication
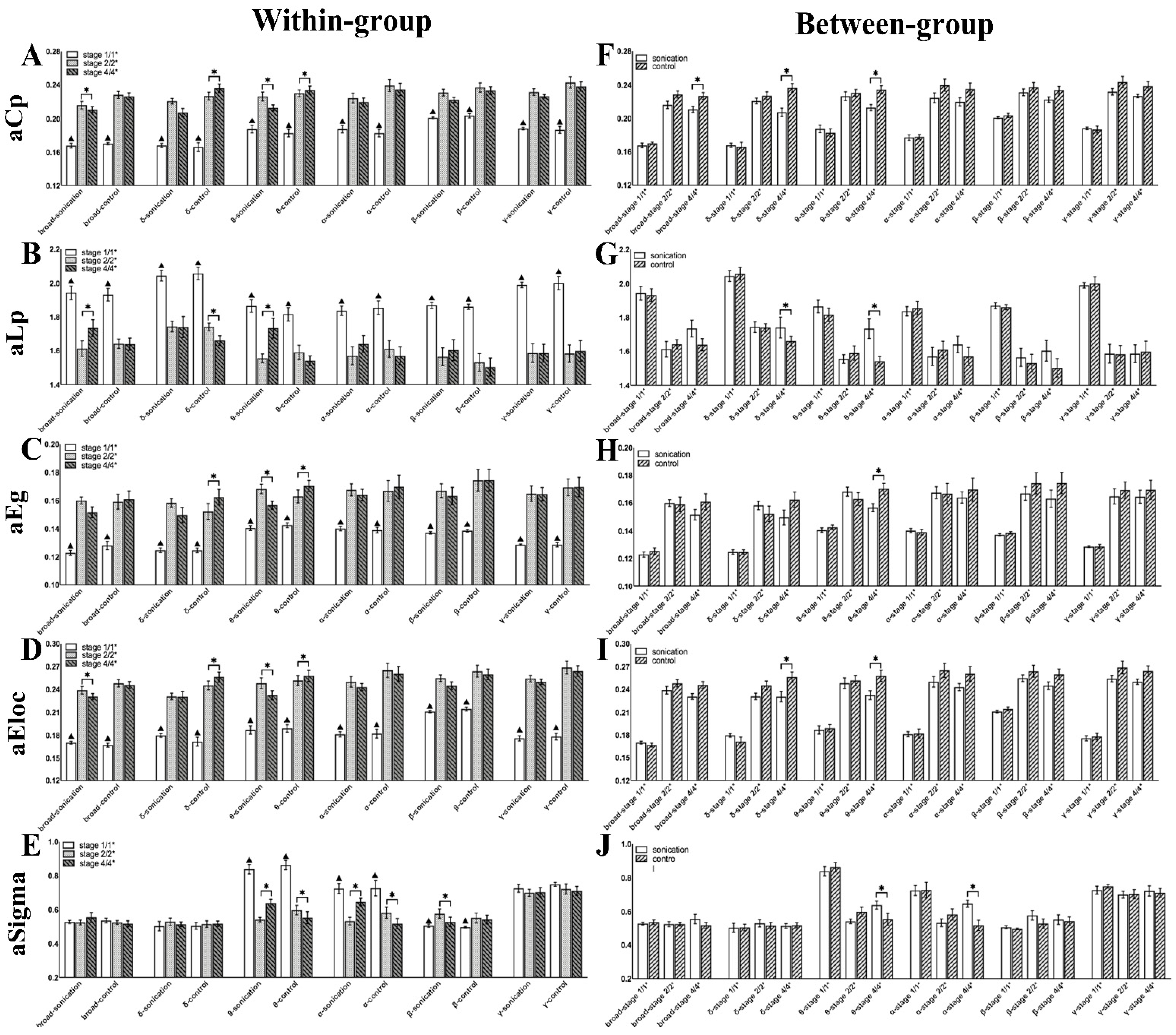
3.4. Brain Network Indicators before and after Sonication
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tyler, W.J.; Tufail, Y.; Finsterwald, M.; Tauchmann, M.L.; Olson, E.J.; Majestic, C. Remote excitation of neuronal circuits using low-intensity, low-frequency ultrasound. PLoS ONE 2008, 3, e3511. [Google Scholar] [CrossRef] [PubMed]
- Tufail, Y.; Matyushov, A.; Baldwin, N.; Tauchmann, M.L.; Georges, J.; Yoshihiro, A.; Tillery, S.I.H.; Tyler, W.J. Transcranial Pulsed Ultrasound Stimulates Intact Brain Circuits. Neuron 2010, 66, 681–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, K.; Sohrabpour, A.; He, B. Electrophysiological Source Imaging of Brain Networks Perturbed by Low-Intensity Transcranial Focused Ultrasound. IEEE Trans. Biomed. Eng. 2016, 63, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.F.; Phipps, M.A.; Newton, A.T.; Chaplin, V.; Gore, J.C.; Caskey, C.F.; Chen, L.M. Neuromodulation of sensory networks in monkey brain by focused ultrasound with MRI guidance and detection. Sci. Rep. 2018, 8, 7993. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhao, Y.; Jia, H.; Liu, M.; Hu, S.; Li, Y.; Li, X. Cortical hemodynamic responses under focused ultrasound stimulation using real-time laser speckle contrast imaging. Front. Neurosci. 2018, 12, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Dong, Y.; Hu, S.; Zheng, T.; Du, D.; Du, J.; Liu, L. Reduced apparent diffusion coefficient in various brain areas following low-intensity transcranial ultrasound stimulation. Front. Neurosci. 2017, 11, 562. [Google Scholar] [CrossRef] [Green Version]
- Min, B.K.; Bystritsky, A.; Jung, K.I.; Fischer, K.; Zhang, Y.; Maeng, L.S.; Park, S.; Chung, Y.A.; Jolesz, F.A.; Yoo, S.S. Focused ultrasound-mediated suppression of chemically-induced acute epileptic EEG activity. BMC Neurosci. 2011, 12, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakimova, H.; Kim, S.; Chu, K.; Lee, S.K.; Jeong, B.; Jeon, D. Ultrasound stimulation inhibits recurrent seizures and improves behavioral outcome in an experimental model of mesial temporal lobe epilepsy. Epilepsy Behav. 2015, 49, 26–32. [Google Scholar] [CrossRef]
- Li, X.; Yang, H.; Wang, X.; Li, X.; Yi, Y. Low-Intensity Pulsed Ultrasound Stimulation Modulates the Nonlinear Dynamics of Local Field Potentials in Temporal Lobe Epilepsy. Front. Neurosci. 2019, 13, 287. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Alonso, J.; Bellas-Lamas, P. Surgical Treatment for Drug-Resistant Epilepsy. Jama 2015, 313, 1572. [Google Scholar] [CrossRef]
- Sasaki, D. Temporal Lobe Epilepsy Surgery:Surgical Complications; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Nadler, J.V.; Spencer, D.D. What Is a Seizure Focus? Adv. Exp. Med. Biol. 2014, 813, 55–62. [Google Scholar] [PubMed]
- Bertram, E.H. Temporal lobe epilepsy: Where do the seizures really begin? Epilepsy Behav. 2009, 14 (Suppl. 1), 32–37. [Google Scholar] [CrossRef] [Green Version]
- Strogatz, S.H. Exploring complex networks. Nature 2001, 410, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Varela, F.; Lachaux, J.P.; Rodriguez, E.; Martinerie, J. The brainweb: Phase synchronization and large-scale integration. Nat. Rev. Neurosci. 2001, 2, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Stam, C.J.; Reijneveld, J.C. Graph theoretical analysis of complex networks in the brain. Nonlinear Biomed. Phys. 2007, 1, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ponten, S.C.; Bartolomei, F.; Stam, C.J. Small-world networks and epilepsy: Graph theoretical analysis of intracerebrally recorded mesial temporal lobe seizures. Clin. Neurophysiol. 2007, 118, 918–927. [Google Scholar] [CrossRef]
- Cymerblit-Sabba, A.; Schiller, Y. Network dynamics during development of pharmacologically induced epileptic seizures in rats in vivo. J. Neurosci. 2010, 30, 1619–1630. [Google Scholar] [CrossRef] [Green Version]
- Schwob, J.E.; Fuller, T.; Price, J.L.; Olney, J.W. Widespread patterns of neuronal damage following systemic or intracerebral injections of kainic acid: A histological study. Neuroscience 1980, 5, 991–1014. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academic Press: Washington, DC, USA, 2010. [Google Scholar]
- Mueller, J.; Legon, W.; Opitz, A.; Sato, T.F.; Tyler, W.J. Transcranial focused ultrasound modulates intrinsic and evoked EEG dynamics. Brain Stimul. 2014, 7, 900–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichardo, S.; Sin, V.W.; Hynynen, K. Multi-frequency characterization of the speed of sound and attenuation coefficient for longitudinal transmission of freshly excised human skulls. Phys. Med. Biol. 2011, 56, 219–250. [Google Scholar] [CrossRef] [Green Version]
- Goss, S.A.; Johnston, R.L.; Dunn, F. Comprehensive compilation of empirical ultrasonic properties of mammalian tissues. J. Acoust. Soc. Am. 1978, 64, 423–457. [Google Scholar] [CrossRef]
- Li, B.; Liu, S.; Hu, D.; Li, G.; Tang, R.; Song, D.; Lang, Y.; He, J. Electrocortical activity in freely walking rats varies with environmental conditions. Brain Res. 2021, 1751. [Google Scholar] [CrossRef] [PubMed]
- Teplan, M. Fundamentals of EEG Measurement. Meas. Sci. Rev. 2002, 2, 1–11. [Google Scholar]
- Zhang, M.; Tang, R.; Lang, Y.; He, J. Construction of a focused ultrasound neuromodulation system for the treatment of epileptic seizure. In Proceedings of the 2019 International Conference on Intelligent Informatics and Biomedical Sciences (ICIIBMS), Shanghai, China, 21–24 November 2019. [Google Scholar] [CrossRef]
- Winrow, V.R.; Winyard, P.G.; Morris, C.J.; Blake, D.R. Free radicals in inflammation: Second messengers and mediators of tissue destruction. Br. Med. Bull. 1993, 49, 506–522. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation: Cortical areas. Electroencephalogr. Clin. Neurophysiol. 1975, 38, 1–12. [Google Scholar] [CrossRef]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of “small-world” network. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef]
- Sporns, O.; Tononi, G.; Edelman, G.M. Connectivity and complexity: The relationship between neuroanatomy and brain dynamics. Neural Netw. 2000, 13, 909–922. [Google Scholar] [CrossRef]
- Salvador, R.; Suckling, J.; Coleman, M.R.; Pickard, J.D.; Menon, D.; Bullmore, E. Neurophysiological architecture of functional magnetic resonance images of human brain. Cereb. Cortex 2005, 15, 1332–2342. [Google Scholar] [CrossRef] [Green Version]
- Achard, S.; Salvador, R.; Whitcher, B.; Suckling, J.; Bullmore, E. A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs. J. Neurosci. 2006, 26, 63–72. [Google Scholar] [CrossRef]
- Micheloyannis, S.; Pachou, E.; Stam, C.J.; Vourkas, M.; Erimaki, S.; Tsirka, V. Using graph theoretical analysis of multi channel EEG to evaluate the neural efficiency hypothesis. Neurosci. Lett. 2006, 402, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Netoff, T.I.; Clewley, R.; Arno, S.; Keck, T.; White, J.A. Epilepsy in small-world networks. J. Neurosci. 2004, 24, 8075–8083. [Google Scholar] [CrossRef] [Green Version]
- Chavez, M.; Hwang, D.U.; Amann, A.; Boccaletti, S. Synchronizing weighted complex networks. Chaos 2006, 16, 015106. [Google Scholar] [CrossRef]
- Rao, M.S.; Hattiangady, B.; Reddy, D.S.; Shetty, A.K. Hippocampal neurodegenetation spontaneous seizures, and mossy fiber sprouting in the F344 rat model of temporal lobe epilepsy. J. Neurosci. Res. 2006, 83, 1088–1105. [Google Scholar] [CrossRef]
- Sanon, N.; Carmant, L.; Emond, M.; Congar, P.; Lacaille, J.C. Short-term effects of kainic acid on CA1 hippocampal interneurons differentially vulnerable to excitotoxicity. Epilepsia 2005, 46, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Douw, L.; van Dellen, E.; de Groot, M.; Heimans, J.J.; Klein, M.; Stam, C.J.; Reijneveld, J.C. Epilepsy is related to theta band brain connectivity and network topology in brain tumor patients. BMC Neurosci. 2010, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavrilov, L.R.; Gersuni, G.V.; Ilyinsky, O.B.; Sirotyuk, M.G.; Tsirulnikov, E.M.; Shchekanov, E.E. The Effect of Focused Ultrasound on the Skin and Deep Nerve Structures of Man and Animal. Prog. Brain Res. 1976, 43, 279–292. [Google Scholar] [CrossRef]
- Sundaram, J.; Mellein, B.R.; Mitragotri, S. An Experimental and Theoretical Analysis of Ultrasound-Induced Permeabilization of Cell Membranes. Biophys. J. 2003, 84, 3087–3101. [Google Scholar] [CrossRef] [Green Version]
- Dinno, M.A.; Dyson, M.; Young, S.R.; Mortimer, A.J.; Hart, J.; Crum, L.A. The significance of membrane changes in the safe and effective use of therapeutic and diagnostic ultrasound. Phys. Med. Biol. 1989, 34, 1543–1552. [Google Scholar] [CrossRef]
- Fisher, R.S.; Van Emde Boas, W.; Blume, W.; Elger, C.; Genton, P.; Lee, P.; Engel, J. Epileptic Seizures and Epilepsy: Definitions Proposed by the International League Against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia 2005, 46, 470–472. [Google Scholar] [CrossRef]
- Borrelli, M.J.; Bailey, K.I.; Dunn, F. Early ultrasonic effects upon mammalian CNS structures (chemical synapses). J. Acoust. Soc. Am. 1981, 69, 1514–1516. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Lyu, K.; Meng, L.; Li, C.; Zhang, H.; Niu, L.; Lin, Z.; Zheng, H.; Tang, J. Low-Intensity Ultrasound Causes Direct Excitation of Auditory Cortical Neurons. Neural Plast. 2021, 2021, 1–10. [Google Scholar] [CrossRef]
- Ye, P.P.; Brown, J.R.; Pauly, K.B. Frequency dependence of ultrasound neurostimulation in the mouse brain. Ultrasound Med. Biol. 2016, 42, 1512–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimsley, J.M.; Galazyuk, A. V An improved approach to separating startle data from noise. J. Neurosci. Methods 2015, 1–12. [Google Scholar] [CrossRef] [Green Version]
- King, R.L.; Brown, J.R.; Newsome, W.T.; Pauly, K.B. Effective parameters for ultrasound-induced in vivo neurostimulation. Ultrasound Med. Biol. 2013, 39, 312–331. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Shapiro, M.G.; Tsao, D.Y. Ultrasonic Neuromodulation Causes Widespread Cortical Activation via an Indirect Auditory Mechanism. Neuron 2018, 98, 1031–1041.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruters, K.G.; Groh, J.M. Sounds and beyond: Multisensory and other non-auditory signals in the inferior colliculus. Front. Neural Circuits 2012, 6, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, V.S.; Altschuler, E.L. The use of visual feedback, in particular mirror visual feedback, in restoring brain function. Brain 2009, 132, 1693–1710. [Google Scholar] [CrossRef] [Green Version]
- Schofield, B.R.; Motts, S.D.; Mellott, J.G. Cholinergic cells of the pontomesencephalic tegmentum: Connections with auditory structures from cochlear nucleus to cortex. Hear. Res. 2011, 279, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korn, M.; Borges, S.S.; Maia, P.R.M.; Lima, J.L.F.C.; Lapa, R.A.S. Reagent generation for chemical analysis assisted by ultrasonic irradiation. Ultrasonics 2004, 42, 585–590. [Google Scholar] [CrossRef]
- Mihran, R.T.; Barnes, F.S.; Wachtel, H. Temporally-specific modification of myelinated axon excitability in vitro following a single ultrasound pulse. Ultrasound Med. Biol. 1990, 16, 297–309. [Google Scholar] [CrossRef]
- Menz, M.D.; Oralkan, Ö.; Khuri-Yakub, P.T.; Baccus, S.A. Precise neural stimulation in the retina using focused ultrasound. J. Neurosci. 2013, 33, 4550–4560. [Google Scholar] [CrossRef]
- Tufail, Y.; Yoshihiro, A.; Pati, S.; Li, M.M.; Tyler, W.J. Ultrasonic neuromodulation by brain stimulation with transcranial ultrasound. Nat. Protoc. 2011, 6, 1453–1470. [Google Scholar] [CrossRef]
- Yoo, S.S.; Bystritsky, A.; Lee, J.H.; Zhang, Y.; Fischer, K.; Min, B.K.; McDannold, N.J.; Pascual-Leone, A.; Jolesz, F.A. Focused ultrasound modulates region-specific brain activity. Neuroimage 2011, 56, 1267–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Garra, B. How to Interpret the Ultrasound Output Display Standard for Higher Acoustic Output Diagnostic Ultrasound Devices: Version 2. J. Ultrasound Med. 2004, 23, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Niu, X.; Krook-magnuson, E.; He, B. Intrinsic functional neuron-type selectivity of transcranial focused ultrasound neuromodulation. Nat. Commun. 2021. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Character | Description |
|---|---|---|
| Weighted clustering coefficient | The extent of local clustering or cliquishness of a network | |
| Shorted weighted path length | The extent of the overall routing efficiency of a network | |
| Sigma | The small-worldness indicating the extent of a network regarding randomness and order | |
| Weighted local efficiency | How efficiently information is propagated to the direct neighbors of a node | |
| Weighted global efficiency | How efficiently information is propagated through the whole network |
| p-Value | Group 1 | Group 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frequency Band | PLV | aCp | aLp | aEg | aEloc | aSigma | PLV | aCp | aLp | aEg | aEloc | aSigma |
| 0.1–48 Hz | 0.1018 | 0.018 | 0.043 | - | 0.018 | - | 0.018 | - | - | - | - | - |
| 0.1–4 Hz | 0.028 | - | - | - | - | - | 0.018 | 0.018 | 0.018 | 0.018 | 0.028 | - |
| 4–8 Hz | 0.018 | 0.018 | - | 0.018 | 0.018 | 0.018 | 0.018 | 0.018 | — | 0.028 | 0.018 | 0.028 |
| 8–12 Hz | - | - | - | - | - | 0.018 | - | - | - | - | - | 0.018 |
| 12–30 Hz | - | - | - | - | - | 0.018 | - | - | - | - | - | - |
| 30–48 Hz | - | - | - | - | - | - | - | - | - | - | - | - |
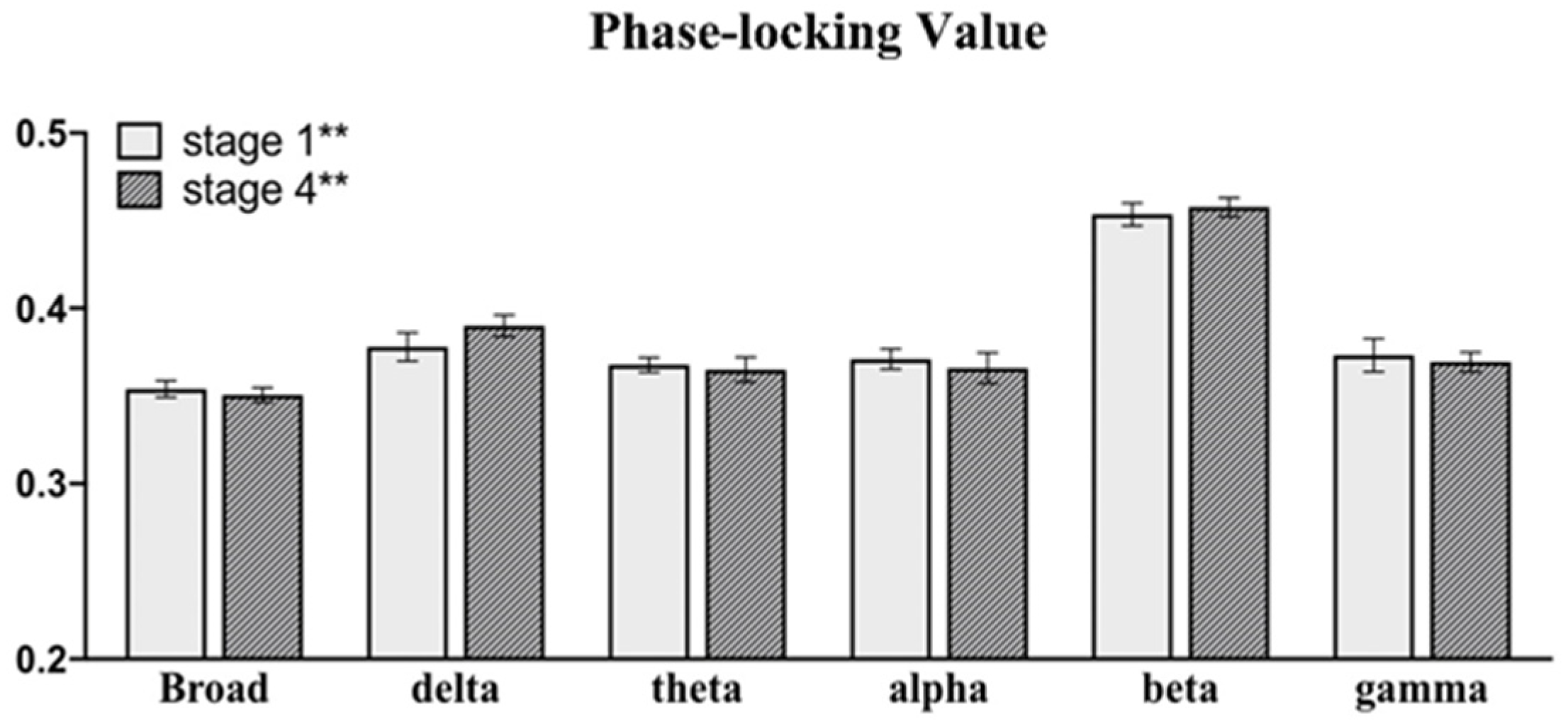
| p-Value | Stage 4/4* | |||||
|---|---|---|---|---|---|---|
| Frequency | PLV | aCp | aLp | aEg | aEloc | aSigma |
| 0.1–48 Hz | 0.048 | 0.035 | - | - | - | - |
| 0.1–4 Hz | 0.018 | 0.011 | - | - | 0.017 | - |
| 4–8 Hz | 0.048 | 0.006 | 0.013 | 0.015 | 0.048 | 0.048 |
| 8–12 Hz | - | - | - | - | 0.018 | |
| 12–30 Hz | - | - | - | - | - | |
| 30–48 Hz | - | - | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Li, B.; Lv, X.; Liu, S.; Liu, Y.; Tang, R.; Lang, Y.; Huang, Q.; He, J. Low-Intensity Focused Ultrasound-Mediated Attenuation of Acute Seizure Activity Based on EEG Brain Functional Connectivity. Brain Sci. 2021, 11, 711. https://doi.org/10.3390/brainsci11060711
Zhang M, Li B, Lv X, Liu S, Liu Y, Tang R, Lang Y, Huang Q, He J. Low-Intensity Focused Ultrasound-Mediated Attenuation of Acute Seizure Activity Based on EEG Brain Functional Connectivity. Brain Sciences. 2021; 11(6):711. https://doi.org/10.3390/brainsci11060711
Chicago/Turabian StyleZhang, Minjian, Bo Li, Xiaodong Lv, Sican Liu, Yafei Liu, Rongyu Tang, Yiran Lang, Qiang Huang, and Jiping He. 2021. "Low-Intensity Focused Ultrasound-Mediated Attenuation of Acute Seizure Activity Based on EEG Brain Functional Connectivity" Brain Sciences 11, no. 6: 711. https://doi.org/10.3390/brainsci11060711