Enhanced Detection of Estrogen-like Compounds by Genetically Engineered Yeast Sensor Strains

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
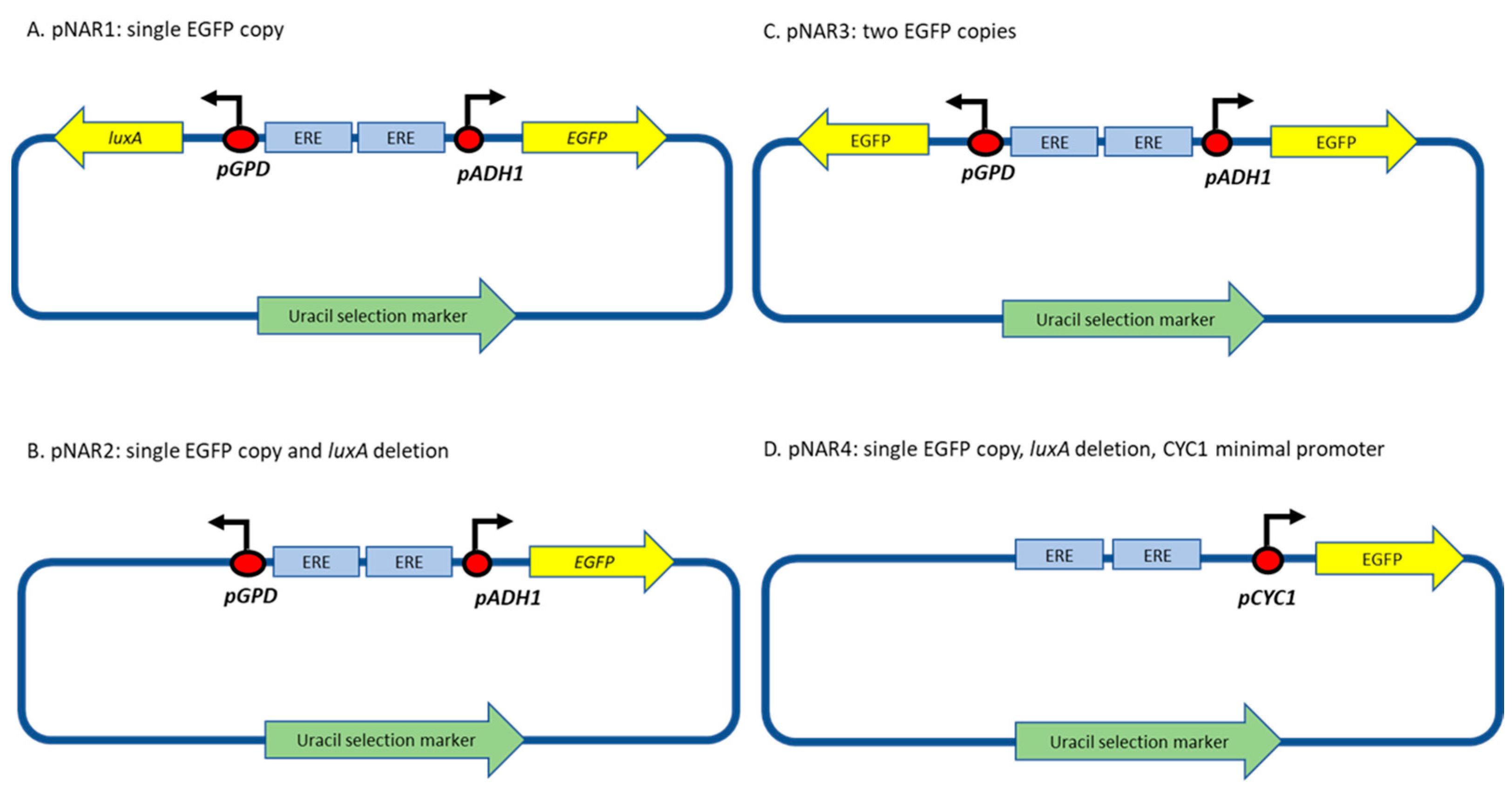
2.2. Yeast Strains, Plasmids and Growth Conditions
- Deleting the luxA gene, yielding plasmid pNAR2 (Figure 1B). Plasmid NAR1 was digested with NotI and SalI restriction enzymes, and a Klenow fragment (New England Biolabs (Ipswich, MA, USA)) was used to create blunt ends, allowing the ligation of the complementary sequence.
- Substituting both the luxA and luxB genes with identical EGFP genes, to produce plasmid pNAR3 (Figure 1C), using restriction enzymes followed by ligation to a respective insert fragment.
- Replacing the ADH1 promoter with the minimal cytochrome C promoter (CYCmin; Plasmid pNAR4, Figure 1D). CYCmin was amplified from plasmid Prsii426-ERE-yNLucP [6], a kind gift from Prof. E. Michelini and Dr. A. Lopreside (University of Bologna, Bologna, Italy), and inserted instead of the ADH1 promoter using the Gibson assembly technique (NEBuilder HiFi DNA Assembly Cloning kit, New England Bio Lab) [18], employing the primers listed in Table S1.
2.3. Deletion of Three Plasma Membrane ABC Cassette Transporter Genes
2.4. Estrogenicity Assay, 96-Well Plates
2.5. Estrogenicity Assay, HPTLC Plate
2.6. Wastewater Treatment Plant Sample Preparation
2.7. Calculations
3. Results
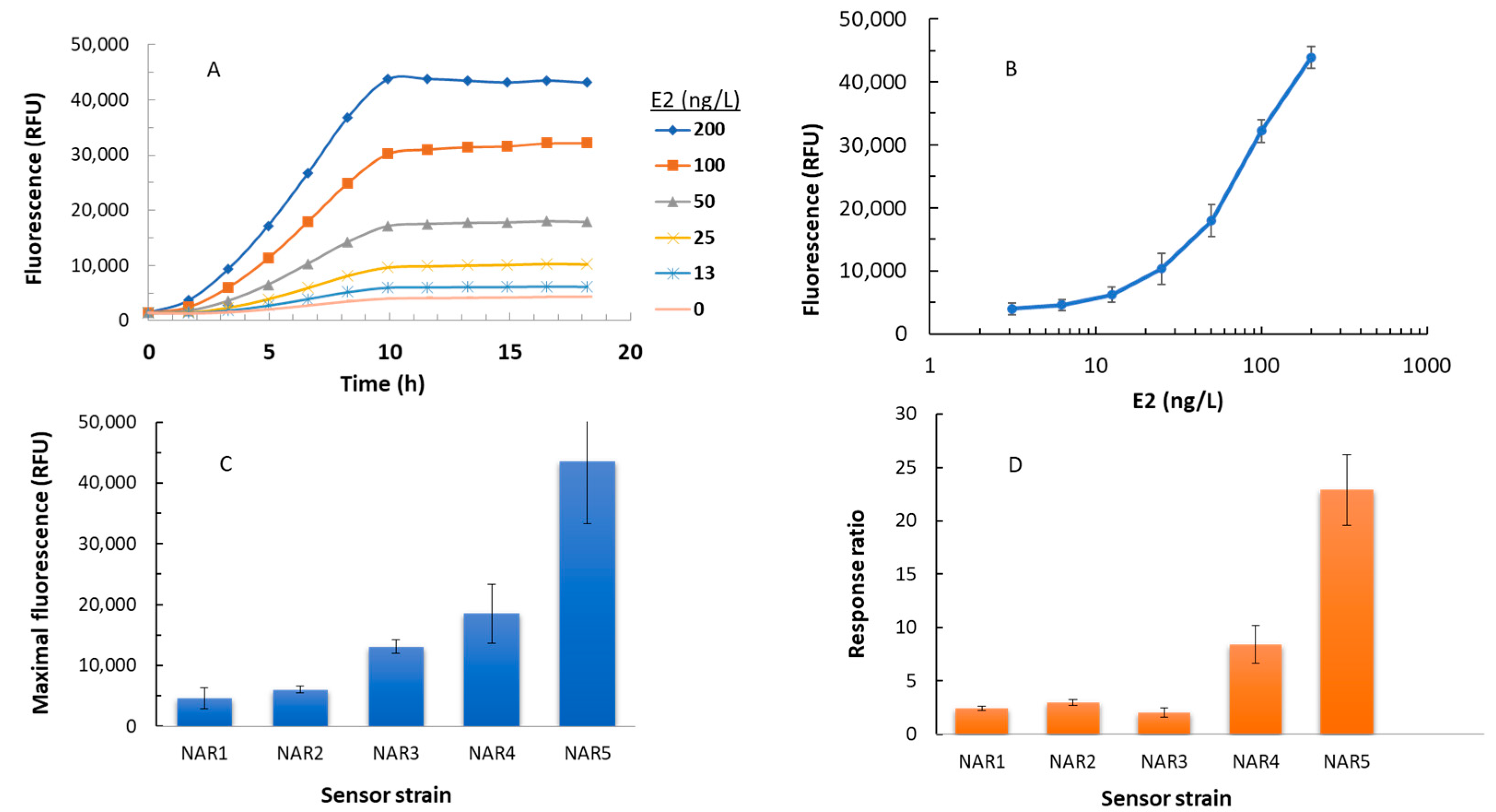
3.1. EGFP Synthesis in Response to Model Estrogenic Compounds in a 96-Well Plate Assay
3.2. Estrogenicity Assay on a TLC Surface
3.3. Detection of Estrogenic Activities in Wastewaters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Campbell, C.G.; Borglin, S.E.; Green, F.B.; Grayson, A.; Wozei, E.; Stringfellow, W.T. Biologically Directed Environmental Monitoring, Fate, and Transport of Estrogenic Endocrine Disrupting Compounds in Water: A Review. Chemosphere 2006, 65, 1265–1280. [Google Scholar] [CrossRef] [PubMed]
- Purdom, C.E.; Hardiman, P.A.; Bye, V.V.J.; Eno, N.C.; Tyler, C.R.; Sumpter, J.P. Estrogenic Effects of Effluents from Sewage Treatment Works. Chem. Ecol. 1994, 8, 275–285. [Google Scholar] [CrossRef]
- Sosa-Ferrera, Z.; Mahugo-Santana, C.; Santana-Rodríguez, J.J. Analytical Methodologies for the Determination of Endocrine Disrupting Compounds in Biological and Environmental Samples. BioMed Res. Int. 2013, 2013, 674838. [Google Scholar] [CrossRef]
- Dotan, P.; Godinger, T.; Odeh, W.; Groisman, L.; Al-Khateeb, N.; Rabbo, A.A.; Tal, A.; Arnon, S. Occurrence and Fate of Endocrine Disrupting Compounds in Wastewater Treatment Plants in Israel and the Palestinian West Bank. Chemosphere 2016, 155, 86–93. [Google Scholar] [CrossRef]
- Adeel, M.; Song, X.; Wang, Y.; Francis, D.; Yang, Y. Environmental Impact of Estrogens on Human, Animal and Plant Life: A Critical Review. Environ. Int. 2017, 99, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Cevenini, L.; Lopreside, A.; Calabretta, M.M.; D’Elia, M.; Simoni, P.; Michelini, E.; Roda, A. A Novel Bioluminescent NanoLuc Yeast-Estrogen Screen Biosensor (NanoYES) with a Compact Wireless Camera for Effect-Based Detection of Endocrine-Disrupting Chemicals. Anal. Bioanal. Chem. 2018, 410, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Moscovici, L.; Riegraf, C.; Abu-Rmailah, N.; Atias, H.; Shakibai, D.; Buchinger, S.; Reifferscheid, G.; Belkin, S. Yeast-Based Fluorescent Sensors for the Simultaneous Detection of Estrogenic and Androgenic Compounds, Coupled with High-Performance Thin Layer Chromatography. Biosensors 2020, 10, 169. [Google Scholar] [CrossRef] [PubMed]
- Gregório, B.J.R.; Ramos, I.I.; Marques, S.S.; Barreiros, L.B.; Magalhães, L.M.; Schneider, R.J.; Segundo, M.A. Microcarrier-based Fluorescent Yeast Estrogen Screen Assay for Fast Determination of Endocrine Disrupting Compounds. Talanta 2024, 271, 125665. [Google Scholar] [CrossRef] [PubMed]
- Wahid, E.; Ocheja, O.B.; Marsili, E.; Guaragnella, C.; Guaragnella, N. Biological and Technical Challenges for Implementation of Yeast-based Biosensors. Microb. Biotechnol. 2023, 16, 54–66. [Google Scholar] [CrossRef]
- Riegraf, C.; Bell, A.M.; Ohlig, M.; Reifferscheid, G.; Buchinger, S. Planar Chromatography-Bioassays for the Parallel and Sensitive Detection of Androgenicity, Anti-androgenicity and Cytotoxicity. J. Chromatogr. A 2022, 1684, 463582. [Google Scholar] [CrossRef]
- Meyer, D.; Marin-Kuan, M.; Mayrhofer, E.; Kirchnawy, C.; Debon, E.; Latado, H.; Patin, A.; Schilter, B.; Morlock, G. Effect-detection by Planar SOS-Umu-C Genotoxicity Bioassay and Chemical Identification of Genotoxins in Packaging Migrates, Proven by Microtiter Plate Assays SOS-Umu-C and Ames-MPF. Food Control 2023, 147, 109546. [Google Scholar] [CrossRef]
- Cruz, G.; Foster, W.; Paredes, A.; Yi, K.D.; Uzumcu, M. Long-term Effects of Early-life Exposure to Environmental Oestrogens on Ovarian Function: Role of Epigenetics. J. Neuroendocrinol. 2014, 26, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Sanseverino, J.; Gupta, R.K.; Layton, A.C.; Patterson, S.S.; Ripp, S.A.; Saidak, L.; Simpson, M.L.; Schultz, T.W.; Sayler, G.S. Use of Saccharomyces Cerevisiae BLYES Expressing Bacterial Bioluminescence for Rapid, Sensitive Detection of Estrogenic Compounds. Appl. Environ. Microbiol. 2005, 71, 4455–4460. [Google Scholar] [CrossRef]
- Baetz, N.; Rothe, L.; Wirzberger, V.; Sures, B.; Schmidt, T.C.; Tuerk, J. High-Performance Thin-Layer Chromatography in Combination with a Yeast-Based Multi-Effect Bioassay to Determine Endocrine Effects in Environmental Samples. Anal. Bioanal. Chem. 2021, 413, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural Diversity of ABC Transporters. J. Gen. Physiol. 2014, 143, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Paumi, C.M.; Chuk, M.; Snider, J.; Stagljar, I.; Michaelis, S. ABC Transporters in Saccharomyces Cerevisiae and Their Interactors: New Technology Advances the Biology of the ABCC (MRP) Subfamily. Microbiol. Mol. Biol. Rev. 2009, 73, 577–593. [Google Scholar] [CrossRef]
- Sipos, G.; Kuchler, K. Fungal ATP-Binding Cassette (ABC) Transporters in Drug Resistance & Detoxification. Curr. Drug Targets 2006, 7, 471–481. [Google Scholar]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic Assembly of DNA Molecules up to Several Hundred Kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Kuchler, K.; Valachovic, M. ABC Proteins in Yeast and Fungal Pathogens. Essays Biochem. 2011, 50, 101. [Google Scholar]
- Güldener, U.; Heck, S.; Fiedler, T.; Beinhauer, J.; Hegemann, J.H. A New Efficient Gene Disruption Cassette for Repeated Use in Budding Yeast. Nucleic Acids Res. 1996, 24, 2519–2524. [Google Scholar] [CrossRef]
- Shakibai, D.; Riegraf, C.; Moscovici, L.; Reifferscheid, G.; Buchinger, S.; Belkin, S. Coupling High-Performance Thin-Layer Chromatography with Bacterial Genotoxicity Bioreporters. Environ. Sci. Technol. 2019, 53, 6410–6419. [Google Scholar] [CrossRef] [PubMed]
- Riegraf, C.; Reifferscheid, G.; Belkin, S.; Moscovici, L.; Shakibai, D.; Hollert, H.; Buchinger, S. Combination of Yeast-Based in vitro Screens with High-Performance Thin-Layer Chromatography as a Novel Tool for the Detection of Hormonal and Dioxin-like Compounds. Anal. Chim. Acta 2019, 1081, 218–230. [Google Scholar] [CrossRef]
- Könemann, S.; Kase, R.; Simon, E.; Swart, K.; Buchinger, S.; Schlüsener, M.; Hollert, H.; Escher, B.I.; Werner, I.; Aït-Aïssa, S.; et al. Effect-based and Chemical Analytical Methods to Monitor Estrogens Under the European Water Framework Directive. Trends Anal. Chem. 2018, 102, 225–235. [Google Scholar] [CrossRef]
- Brack, W.; Dulio, V.; Ågerstrand, M.; Allan, I.; Altenburger, R.; Brinkmann, M.; Bunke, D.; Burgess, R.M.; Cousins, I.; Escher, B.I.; et al. Towards the Review of the European Union Water Framework Directive: Recommendations for More Efficient Assessment and Management of Chemical Contamination in European Surface Water Resources. Sci. Total Environ. 2017, 576, 720–737. [Google Scholar] [CrossRef] [PubMed]
- Verordnung über Anforderungen an das Einleiten von Abwasser in Gewässer (Abwasserverordnung—AbwV) neugefasst durch B. v. 17.06.2004 BGBl. I S. 1108, 2625; zuletzt geändert durch Artikel 1 V. v. 27.02.2024 BGBl. 2024 I Nr. 66. Available online: https://www.gesetze-im-internet.de/abwv/ (accessed on 9 April 2024).
- Hasenbrink, G.; Sievernich, A.; Wildt, L.; Ludwig, J.; Lichtenberg-Fraté, H.; Hasenbrink, G.; Sievernich, A.; Wildt, L.; Ludwig, J.; Lichtenberg-Fraté, H. Estrogenic Effects of Natural and Synthetic Compounds Including Tibolone Assessed in Saccharomyces Cerevisiae Expressing the Human Estrogen α and β Receptors. FASEB J. 2006, 20, 1552–1554. [Google Scholar] [CrossRef]
- Sievernich, A.; Wildt, L.; Lichtenberg-Fraté, H. In Vitro Bioactivity of 17α-Estradiol. J. Steroid Biochem. Mol. Biol. 2004, 92, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Bovee, T.F.; Helsdingen, R.J.; Koks, P.D.; Kuiper, H.A.; Hoogenboom, R.L.; Keijer, J. Development of a Rapid Yeast Estrogen Bioassay, Based on the Expression of Green Fluorescent Protein. Gene 2004, 325, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Chamas, A.; Pham, H.T.M.; Jähne, M.; Hettwer, K.; Gehrmann, L.; Tuerk, J.; Uhlig, S.; Simon, K.; Baronian, K.; Kunze, G. Separation and Identification of Hormone-Active Compounds Using a Combination of Chromatographic Separation and Yeast-Based Reporter Assay. Sci. Total Environ. 2017, 605, 507–513. [Google Scholar] [CrossRef]
- Jaber, M.; Jähne, M.; Oberle, M.; Morlock, G.E. Screening Bisphenols in Complex Samples via a Planar Arxula adeninivorans bioluminescence bioassay. Anal Bioanal Chem. 2023, 415, 5193–5204. [Google Scholar] [CrossRef]
- Arnold, S.F.; Robinson, M.K.; Notides, A.C.; Guillette, L.J., Jr.; McLachlan, J.A. A Yeast Estrogen Screen for Examining the Relative Exposure of Cells to Natural and Xenoestrogens. Environ. Health Perspect. 1996, 104, 544–548. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development (OECD). Test No. 455: The Stably Transfected Human Estrogen Receptor-alpha Transcriptional Activation Assay for Detection of Estrogenic Agonist-Activity of Chemicals; OECD Publishing: Paris, France, 2009. [Google Scholar]
- Schreiner, T.; Ronzheimer, A.; Friz, M.; Morlock, G.E. Multiplex Planar Bioassay with Reduced Diffusion on Normal Phase, Identifying Androgens, Verified Antiandrogens and Synergists in Botanicals via 12D Hyphenation. Food Chem. 2022, 395, 133610. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Host | Plasmid | Comment |
|---|---|---|---|
| NAR1 | WT | pNAR1 | Baseline plasmid [7] |
| NAR2 | WT | pNAR2 | luxA deletion |
| NAR3 | WT | pNAR3 | luxA deletion, EGFPx2 |
| NAR4 | WT | pNAR4 | luxA deletion, CYCmin promotor |
| NAR5 | ΔPDR5, ΔSNQ2, ΔYOR1 | pNAR4 | luxA deletion, CYCmin promotor, 3 transporter mutations |
| Sensor Strain: | NAR1 | NAR2 | NAR3 | NAR4 | NAR5 |
|---|---|---|---|---|---|
| Maximal fluorescence * ** | 4600 ± 17.15 | 6050 ± 610 | 13,100 ± 1052 | 18,550 ± 4830 | 43,570 ± 10,230 |
| Maximal response ratio * | 2.4 ± 0.2 | 3 ± 0.3 | 2 ± 0.4 | 8.4 ± 1.8 | 22.9 ± 3 |
| LOD (ng/L) | 29 ± 3 | 34 ± 7 | 38 ± 4 | 12 ± 1.5 | 8 ± 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abu-Rmailah, N.; Moscovici, L.; Riegraf, C.; Atias, H.; Buchinger, S.; Reifferscheid, G.; Belkin, S. Enhanced Detection of Estrogen-like Compounds by Genetically Engineered Yeast Sensor Strains. Biosensors 2024, 14, 193. https://doi.org/10.3390/bios14040193
Abu-Rmailah N, Moscovici L, Riegraf C, Atias H, Buchinger S, Reifferscheid G, Belkin S. Enhanced Detection of Estrogen-like Compounds by Genetically Engineered Yeast Sensor Strains. Biosensors. 2024; 14(4):193. https://doi.org/10.3390/bios14040193
Chicago/Turabian StyleAbu-Rmailah, Nidaa, Liat Moscovici, Carolin Riegraf, Hadas Atias, Sebastian Buchinger, Georg Reifferscheid, and Shimshon Belkin. 2024. "Enhanced Detection of Estrogen-like Compounds by Genetically Engineered Yeast Sensor Strains" Biosensors 14, no. 4: 193. https://doi.org/10.3390/bios14040193