Neuronal Electrophysiological Activities Detection of Defense Behaviors Using an Implantable Microelectrode Array in the Dorsal Periaqueductal Gray


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Fabrication and Nanomodification of the MEA
2.3. Surgery
2.4. Behavioral Recording Procedures
2.5. Histology
2.6. Statistics
3. Results
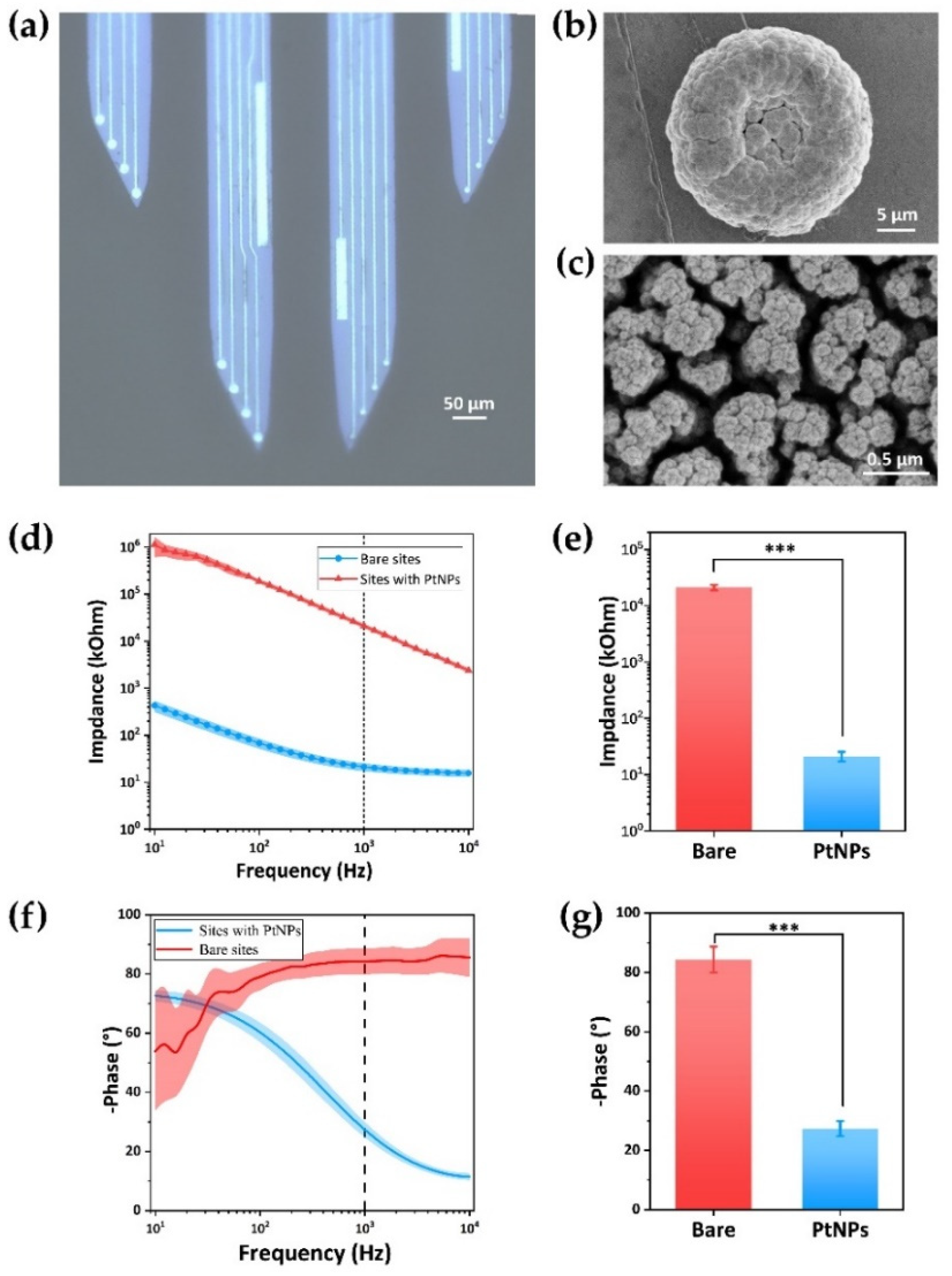
3.1. Characterization of MEAs
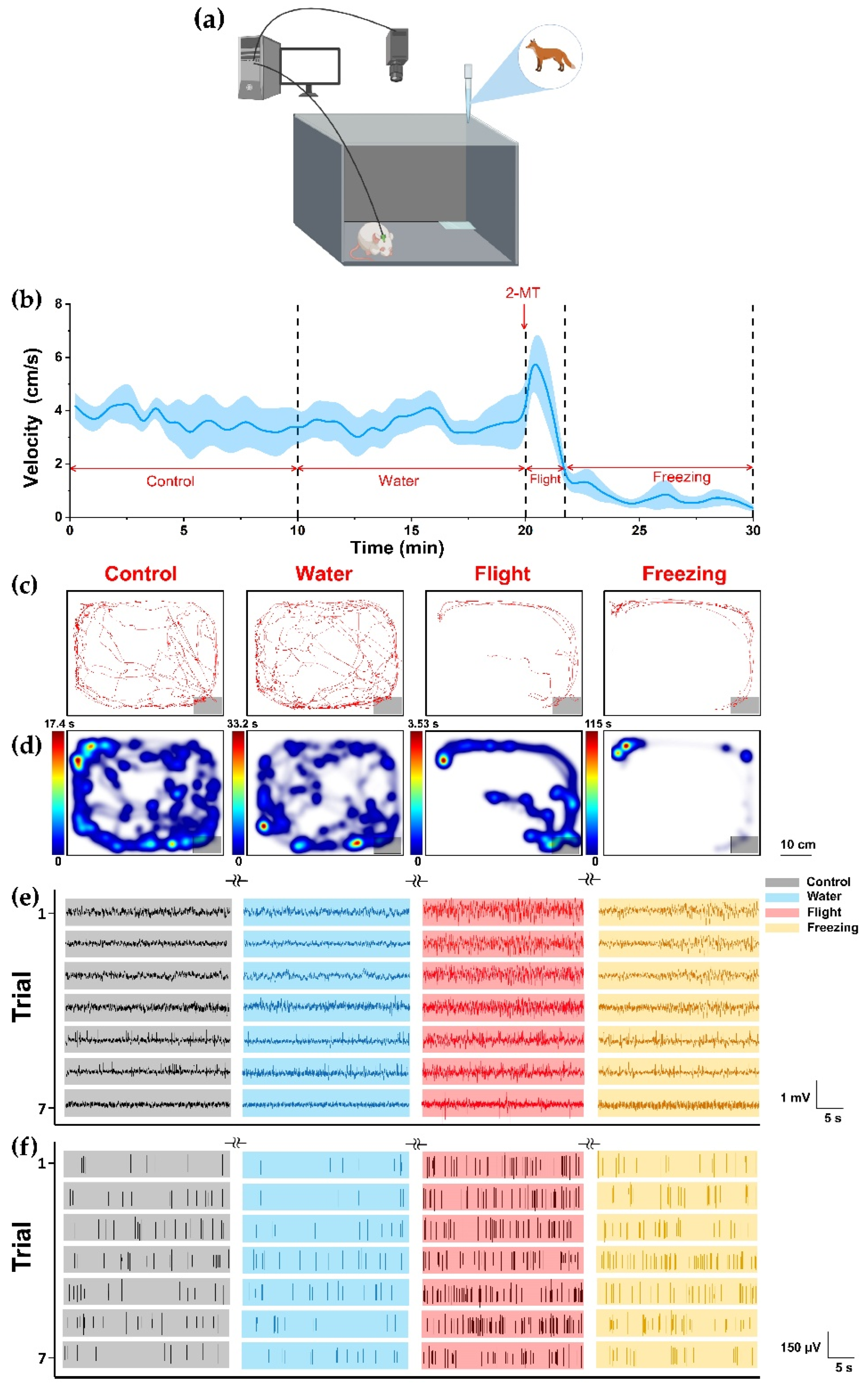
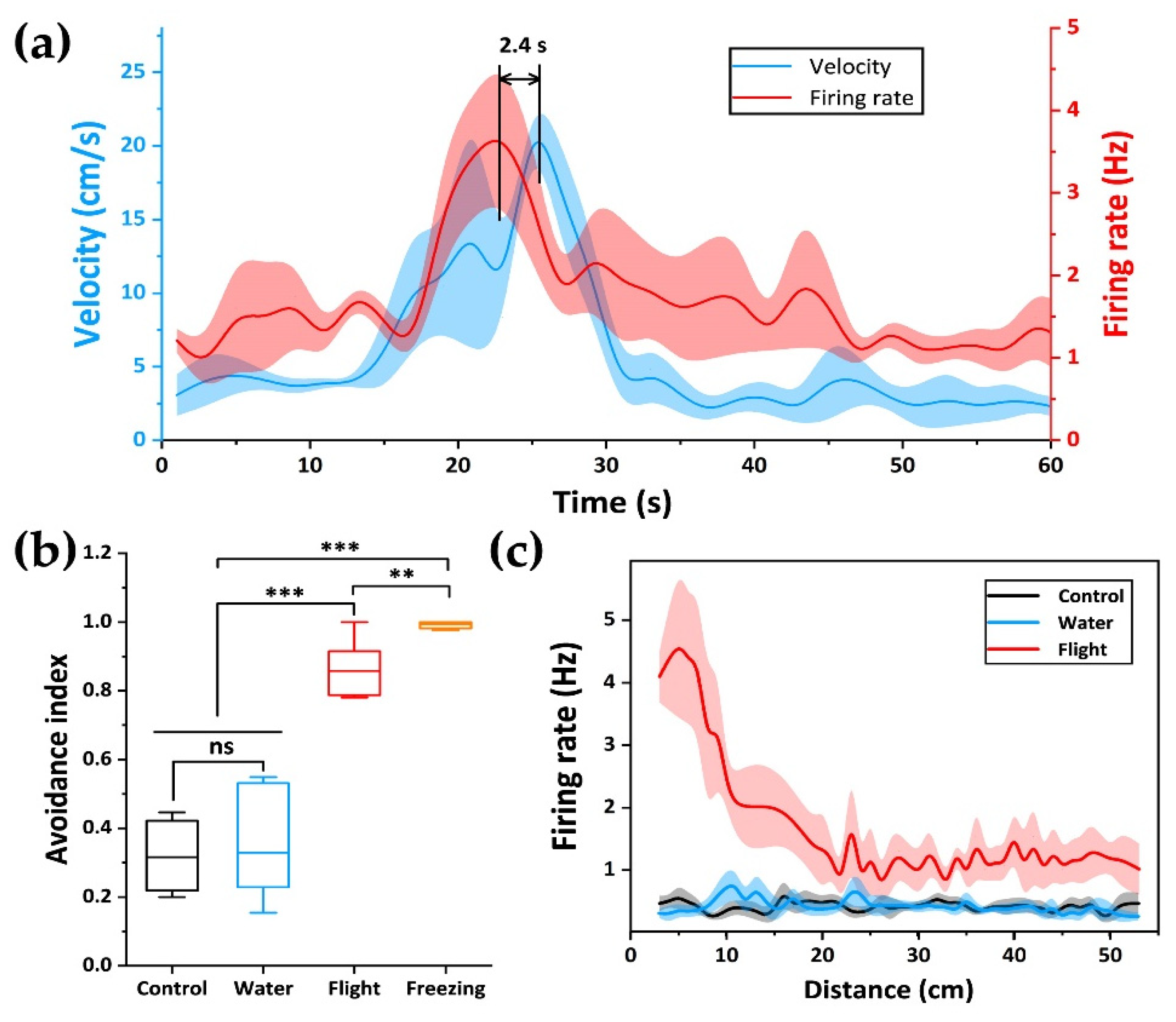
3.2. Behavioral and Electrophysiological Recording of Pedator Odor Induced Innate Fear
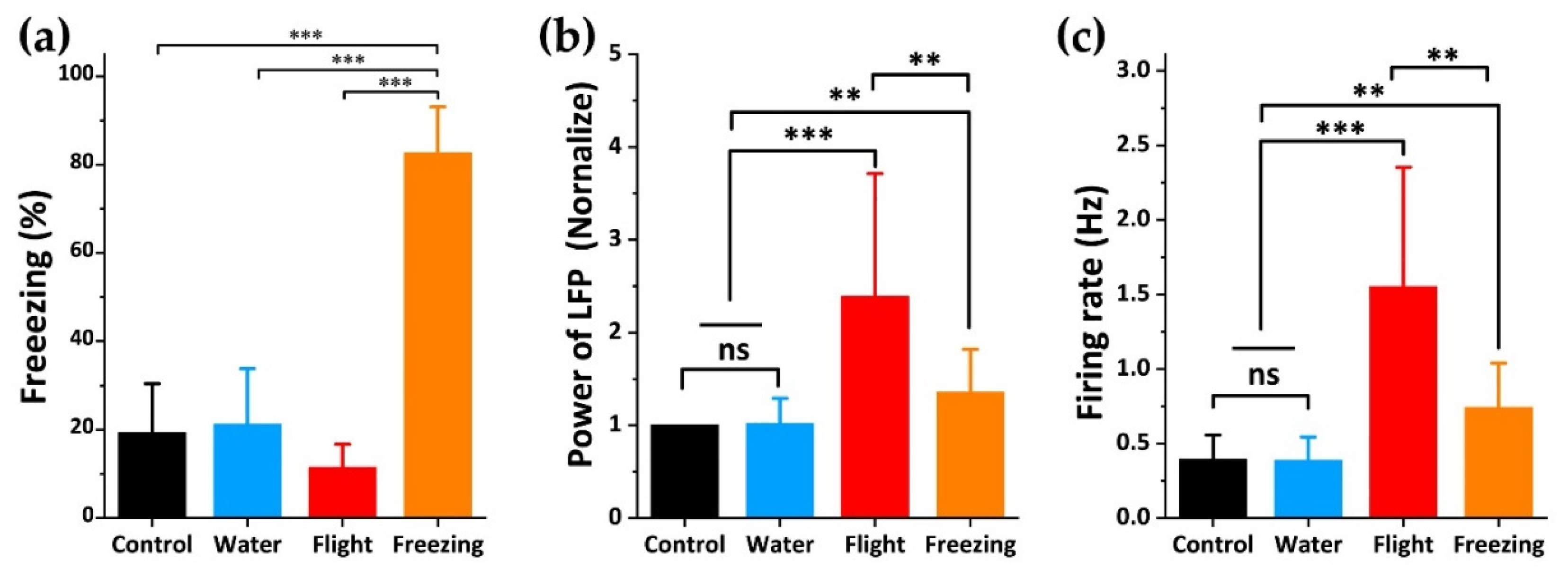
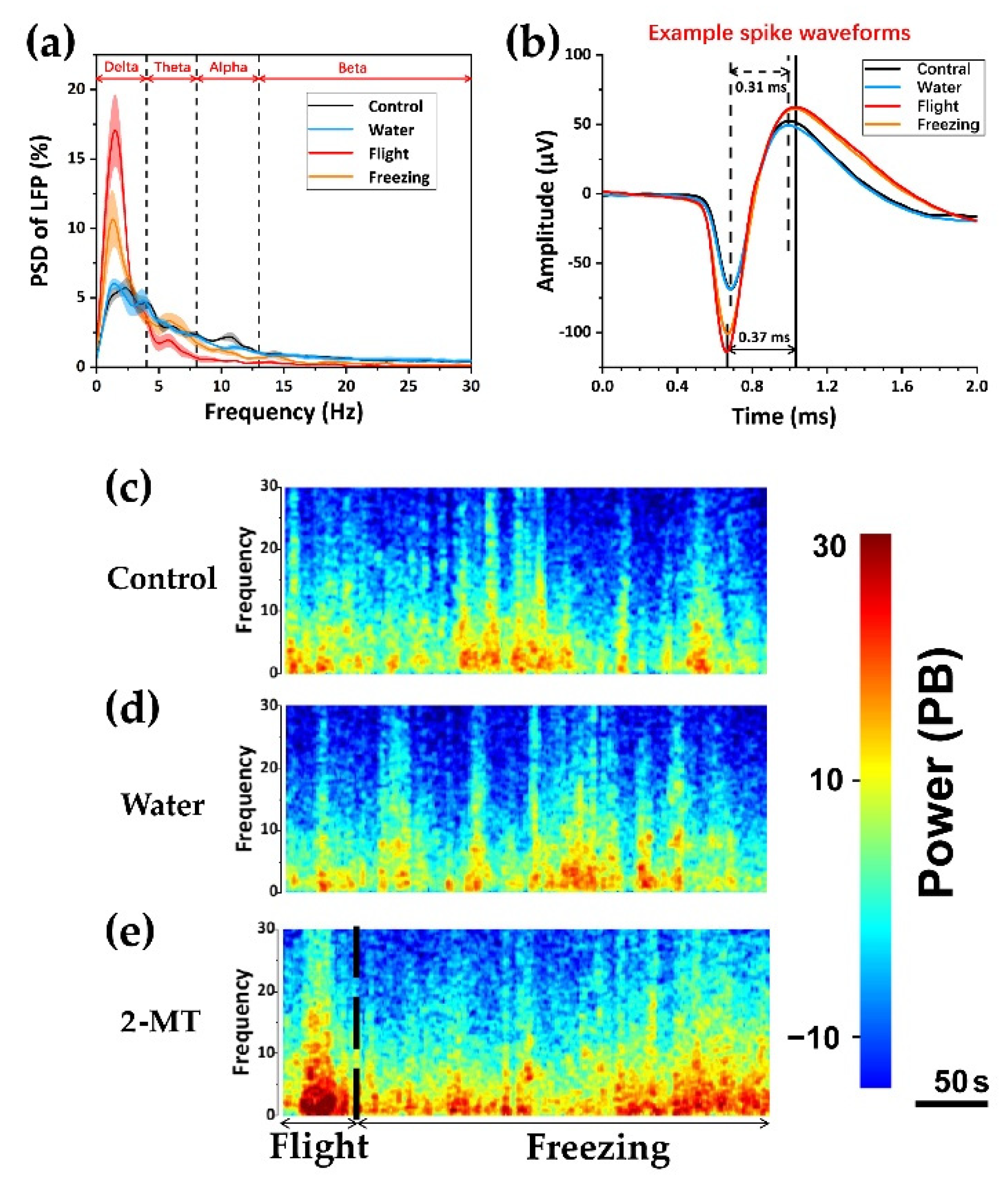
3.3. Relationship between dPAG Activity and Flight Behavior during Exposure to Predator Odor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blanchard, D.C.; Blanchard, R.J. Ethoexperimental approaches to the biology of emotion. Annu. Rev. Psychol. 1988, 39, 43–68. [Google Scholar] [CrossRef] [PubMed]
- Mobbs, D.; Petrovic, P.; Marchant, J.L.; Hassabis, D.; Weiskopf, N.; Seymour, B.; Dolan, R.J.; Frith, C.D. When fear is near: Threat imminence elicits prefrontal-periaqueductal gray shifts in humans. Science 2007, 317, 1079–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, J.B.; Pagani, J.H.; Rolla, K.L.; Davis, C. Analysis of behavioral constraints and the neuroanatomy of fear to the predator odor trimethylthiazoline: A model for animal phobias. Neurosci. Biobehav. Rev. 2008, 32, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Parsons, R.G.; Ressler, K.J. Implications of memory modulation for post-traumatic stress and fear disorders. Nat. Neurosci. 2013, 16, 146–153. [Google Scholar] [CrossRef]
- Pellman, B.A.; Kim, J.J. What Can Ethobehavioral Studies Tell Us about the Brain’s Fear System? Trends Neurosci. 2016, 39, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, L.K.; Nakashima, B.R.; Hong, H.; Watanabe, K. The smell of danger: A behavioral and neural analysis of predator odor-induced fear. Neurosci. Biobehav. Rev. 2005, 29, 1157–1167. [Google Scholar] [CrossRef]
- Apfelbach, R.; Blanchard, C.D.; Blanchard, R.J.; Hayes, R.A.; McGregor, I.S. The effects of predator odors in mammalian prey species: A review of field and laboratory studies. Neurosci. Biobehav. Rev. 2005, 29, 1123–1144. [Google Scholar] [CrossRef]
- Gross, C.T.; Canteras, N.S. The many paths to fear. Nat. Rev. Neurosci. 2012, 13, 651–658. [Google Scholar] [CrossRef]
- Saito, H.; Nishizumi, H.; Suzuki, S.; Matsumoto, H.; Ieki, N.; Abe, T.; Kiyonari, H.; Morita, M.; Yokota, H.; Hirayama, N.; et al. Immobility responses are induced by photoactivation of single glomerular species responsive to fox odour TMT. Nat. Commun. 2017, 8, 16011. [Google Scholar] [CrossRef]
- Chen, W.B.; Pan, H.Q.; He, Y.; Wang, X.H.; Zhang, W.H.; Pan, B.X. Rap1b but not Rap1a in the forebrain is required for learned fear. Cell Biosci. 2020, 10, 107. [Google Scholar] [CrossRef]
- Isosaka, T.; Matsuo, T.; Yamaguchi, T.; Funabiki, K.; Nakanishi, S.; Kobayakawa, R.; Kobayakawa, K. Htr2a-Expressing Cells in the Central Amygdala Control the Hierarchy between Innate and Learned Fear. Cell 2015, 163, 1153–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Cao, L.; Lee, C.Y.; Matsuo, T.; Wu, K.; Asher, G.; Tang, L.; Saitoh, T.; Russell, J.; Klewe-Nebenius, D.; et al. Large-scale forward genetics screening identifies Trpa1 as a chemosensor for predator odor-evoked innate fear behaviors. Nat. Commun. 2018, 9, 2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhang, J.; Lee, H.; Kang, M.S.; Lee, H.S.; Park, H.; Han, J.H. Anterior cingulate cortex and its input to the basolateral amygdala control innate fear response. Nat. Commun. 2018, 9, 2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandão, M.L.; Anseloni, V.Z.; Pandóssio, J.E.; De Araújo, J.E.; Castilho, V.M. Neurochemical mechanisms of the defensive behavior in the dorsal midbrain. Neurosci. Biobehav. Rev. 1999, 23, 863–875. [Google Scholar] [CrossRef]
- Silva, C.; McNaughton, N. Are periaqueductal gray and dorsal raphe the foundation of appetitive and aversive control? A comprehensive review. Prog. Neurobiol. 2019, 177, 33–72. [Google Scholar] [CrossRef]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Fadok, J.P.; Krabbe, S.; Markovic, M.; Courtin, J.; Xu, C.; Massi, L.; Botta, P.; Bylund, K.; Muller, C.; Kovacevic, A.; et al. A competitive inhibitory circuit for selection of active and passive fear responses. Nature 2017, 542, 96–100. [Google Scholar] [CrossRef]
- Motta, S.C.; Carobrez, A.P.; Canteras, N.S. The periaqueductal gray and primal emotional processing critical to influence complex defensive responses, fear learning and reward seeking. Neurosci. Biobehav. Rev. 2017, 76, 39–47. [Google Scholar] [CrossRef]
- Fanselow, M.S.; Kim, J.J.; Young, S.L.; Calcagnetti, D.J.; DeCola, J.P.; Helmstetter, F.J.; Landeira-Fernandez, J. Differential effects of selective opioid peptide antagonists on the acquisition of Pavlovian fear conditioning. Peptides 1991, 12, 1033–1037. [Google Scholar] [CrossRef] [Green Version]
- Frontera, J.L.; Baba Aissa, H.; Sala, R.W.; Mailhes-Hamon, C.; Georgescu, I.A.; Lena, C.; Popa, D. Bidirectional control of fear memories by cerebellar neurons projecting to the ventrolateral periaqueductal grey. Nat. Commun. 2020, 11, 5207. [Google Scholar] [CrossRef]
- Bittencourt, A.S.; Nakamura-Palacios, E.M.; Mauad, H.; Tufik, S.; Schenberg, L.C. Organization of electrically and chemically evoked defensive behaviors within the deeper collicular layers as compared to the periaqueductal gray matter of the rat. Neuroscience 2005, 133, 873–892. [Google Scholar] [CrossRef] [PubMed]
- Tovote, P.; Esposito, M.S.; Botta, P.; Chaudun, F.; Fadok, J.P.; Markovic, M.; Wolff, S.B.; Ramakrishnan, C.; Fenno, L.; Deisseroth, K.; et al. Midbrain circuits for defensive behaviour. Nature 2016, 534, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Johansen, J.P.; Tarpley, J.W.; LeDoux, J.E.; Blair, H.T. Neural substrates for expectation-modulated fear learning in the amygdala and periaqueductal gray. Nat. Neurosci. 2010, 13, 979–986. [Google Scholar] [CrossRef]
- Chou, X.L.; Wang, X.; Zhang, Z.G.; Shen, L.; Zingg, B.; Huang, J.; Zhong, W.; Mesik, L.; Zhang, L.I.; Tao, H.W. Inhibitory gain modulation of defense behaviors by zona incerta. Nat. Commun. 2018, 9, 1151. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.R.; Liang, F.; Zingg, B.; Ji, X.Y.; Ibrahim, L.A.; Tao, H.W.; Zhang, L.I. Auditory cortex controls sound-driven innate defense behaviour through corticofugal projections to inferior colliculus. Nat. Commun. 2015, 6, 7224. [Google Scholar] [CrossRef] [Green Version]
- LeDoux, J.E. Emotion circuits in the brain. Annu. Rev. Neurosci. 2000, 23, 155–184. [Google Scholar] [CrossRef]
- Herry, C.; Ciocchi, S.; Senn, V.; Demmou, L.; Muller, C.; Luthi, A. Switching on and off fear by distinct neuronal circuits. Nature 2008, 454, 600–606. [Google Scholar] [CrossRef]
- Duvarci, S.; Popa, D.; Pare, D. Central amygdala activity during fear conditioning. J. Neurosci. 2011, 31, 289–294. [Google Scholar] [CrossRef]
- Shin, H.; Son, Y.; Chae, U.; Kim, J.; Choi, N.; Lee, H.J.; Woo, J.; Cho, Y.; Yang, S.H.; Lee, C.J.; et al. Multifunctional multi-shank neural probe for investigating and modulating long-range neural circuits in vivo. Nat. Commun. 2019, 10, 3777. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Song, Y.; Wang, M.; Zhang, Z.; Fan, X.; Song, X.; Zhuang, P.; Yue, F.; Chan, P.; Cai, X. A silicon based implantable microelectrode array for electrophysiological and dopamine recording from cortex to striatum in the non-human primate brain. Biosens. Bioelectron. 2016, 85, 53–61. [Google Scholar] [CrossRef]
- Shin, H.; Jeong, S.; Lee, J.H.; Sun, W.; Choi, N.; Cho, I.J. 3D high-density microelectrode array with optical stimulation and drug delivery for investigating neural circuit dynamics. Nat. Commun. 2021, 12, 492. [Google Scholar] [CrossRef] [PubMed]
- Song, E.; Li, J.; Won, S.M.; Bai, W.; Rogers, J.A. Materials for flexible bioelectronic systems as chronic neural interfaces. Nat. Mater. 2020, 19, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Won, S.M.; Song, E.; Zhao, J.; Li, J.; Rivnay, J.; Rogers, J.A. Recent Advances in Materials, Devices, and Systems for Neural Interfaces. Adv. Mater. 2018, 30, e1800534. [Google Scholar] [CrossRef] [PubMed]
- Nolta, N.F.; Christensen, M.B.; Crane, P.D.; Skousen, J.L.; Tresco, P.A. BBB leakage, astrogliosis, and tissue loss correlate with silicon microelectrode array recording performance. Biomaterials 2015, 53, 753–762. [Google Scholar] [CrossRef]
- Dai, Y.; Song, Y.; Xie, J.; Xiao, G.; Li, X.; Li, Z.; Gao, F.; Zhang, Y.; He, E.; Xu, S.; et al. CB1-Antibody Modified Liposomes for Targeted Modulation of Epileptiform Activities Synchronously Detected by Microelectrode Arrays. ACS Appl. Mater. Interfaces 2020, 12, 41148–41156. [Google Scholar] [CrossRef]
- Xie, J.; Song, Y.; Dai, Y.; Li, Z.; Gao, F.; Li, X.; Xiao, G.; Zhang, Y.; Wang, H.; Lu, Z.; et al. Nanoliposome-encapsulated caged-GABA for modulating neural electrophysiological activity with simultaneous detection by microelectrode arrays. Nano Res. 2020, 13, 1756–1763. [Google Scholar] [CrossRef]
- Lu, Z.; Xu, S.; Wang, H.; Liu, J.; Dai, Y.; Xie, J.; Song, Y.; Wang, Y.; Wang, Y.; Qu, L.; et al. Total sleep deprivation-induced electrophysiological activities changes in rat hippocampal CA1 detected by microelectrode arrays. Sens. Actuators A Phys. 2021, 331, 112983. [Google Scholar] [CrossRef]
- Vieira-Rasteli, E.B.; de Paula, B.B.; de Paiva, Y.B.; Coimbra, N.C.; Leite-Panissi, C.R.A. Restricted lesions of the ventrolateral or dorsal columns of the periaqueductal gray promotes distinct effects on tonic immobility and defensive analgesia in guinea pigs. Physiol. Behav. 2018, 194, 538–544. [Google Scholar] [CrossRef]
- Dielenberg, R.A.; Hunt, G.E.; McGregor, I.S. ‘When a rat smells a cat’: The distribution of Fos immunoreactivity in rat brain following exposure to a predatory odor. Neuroscience 2001, 104, 1085–1097. [Google Scholar] [CrossRef]
- Watson, T.C.; Cerminara, N.L.; Lumb, B.M.; Apps, R. Neural Correlates of Fear in the Periaqueductal Gray. J. Neurosci. 2016, 36, 12707–12719. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, B.; Fan, P.; Wang, Y.; Dai, Y.; Xie, J.; Yang, G.; Mo, F.; Xu, Z.; Song, Y.; Liu, J.; et al. Neuronal Electrophysiological Activities Detection of Defense Behaviors Using an Implantable Microelectrode Array in the Dorsal Periaqueductal Gray. Biosensors 2022, 12, 193. https://doi.org/10.3390/bios12040193
Lu B, Fan P, Wang Y, Dai Y, Xie J, Yang G, Mo F, Xu Z, Song Y, Liu J, et al. Neuronal Electrophysiological Activities Detection of Defense Behaviors Using an Implantable Microelectrode Array in the Dorsal Periaqueductal Gray. Biosensors. 2022; 12(4):193. https://doi.org/10.3390/bios12040193
Chicago/Turabian StyleLu, Botao, Penghui Fan, Yiding Wang, Yuchuan Dai, Jingyu Xie, Gucheng Yang, Fan Mo, Zhaojie Xu, Yilin Song, Juntao Liu, and et al. 2022. "Neuronal Electrophysiological Activities Detection of Defense Behaviors Using an Implantable Microelectrode Array in the Dorsal Periaqueductal Gray" Biosensors 12, no. 4: 193. https://doi.org/10.3390/bios12040193