
Syndecan-1 Expression Is Increased in the Aortic Wall of Patients with Type 2 Diabetes but Is Unrelated to Elevated Fasting Plasma Glucagon-Like Peptide-1

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
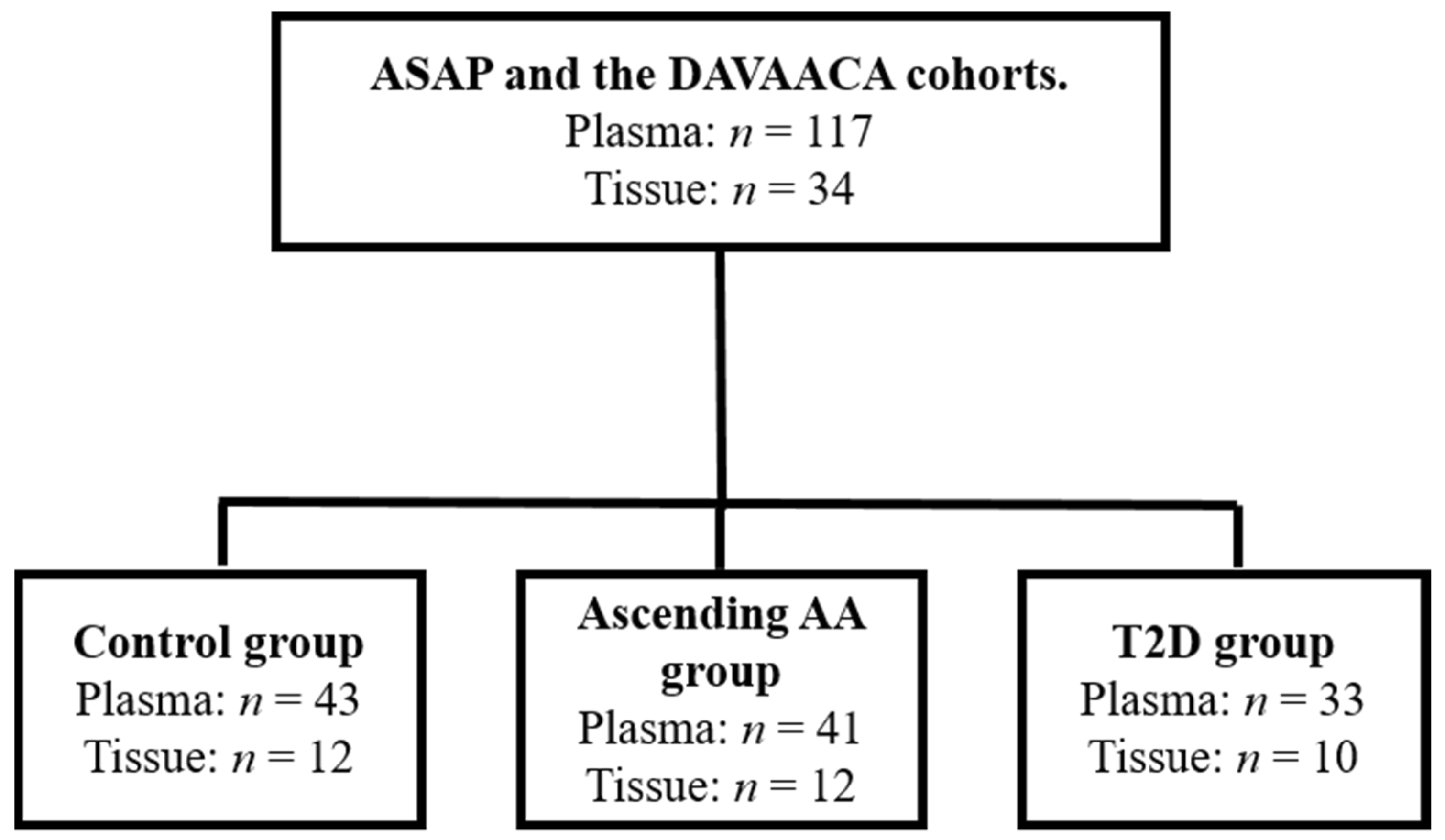
2.1. Patient Information
2.2. Measurement of the fpGLP-1
2.3. Matrix Metalloproteinase-2 Activity Assay
2.4. Syndecan-1 Shedding
2.5. Cytokines
2.6. Western Blot
2.7. Statistical Analysis
3. Results
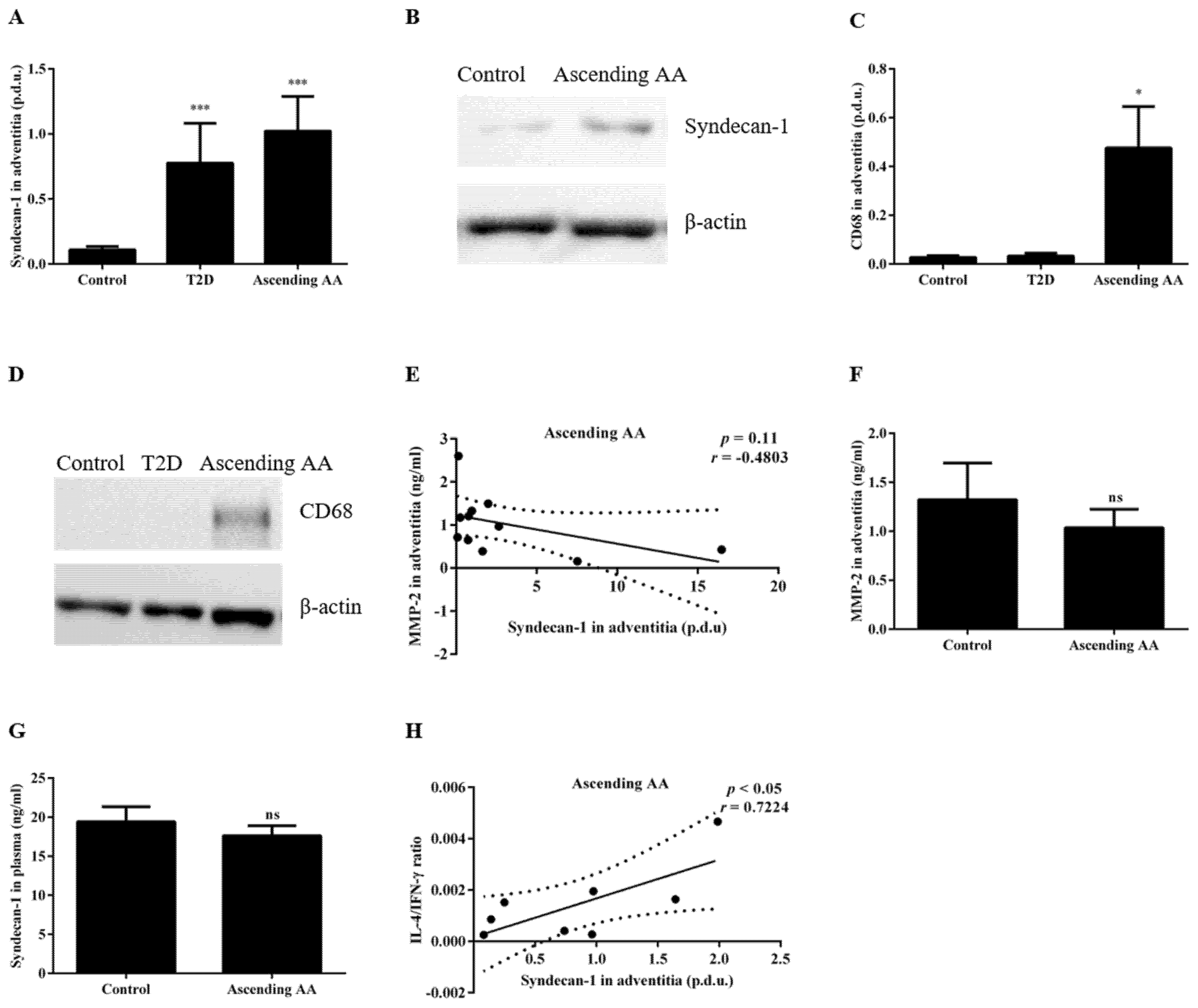
3.1. Type 2 Diabetes Is Associated with Decreased Plasma Sdc-1 and Increased Expression of Sdc-1 in Aortic Tissue
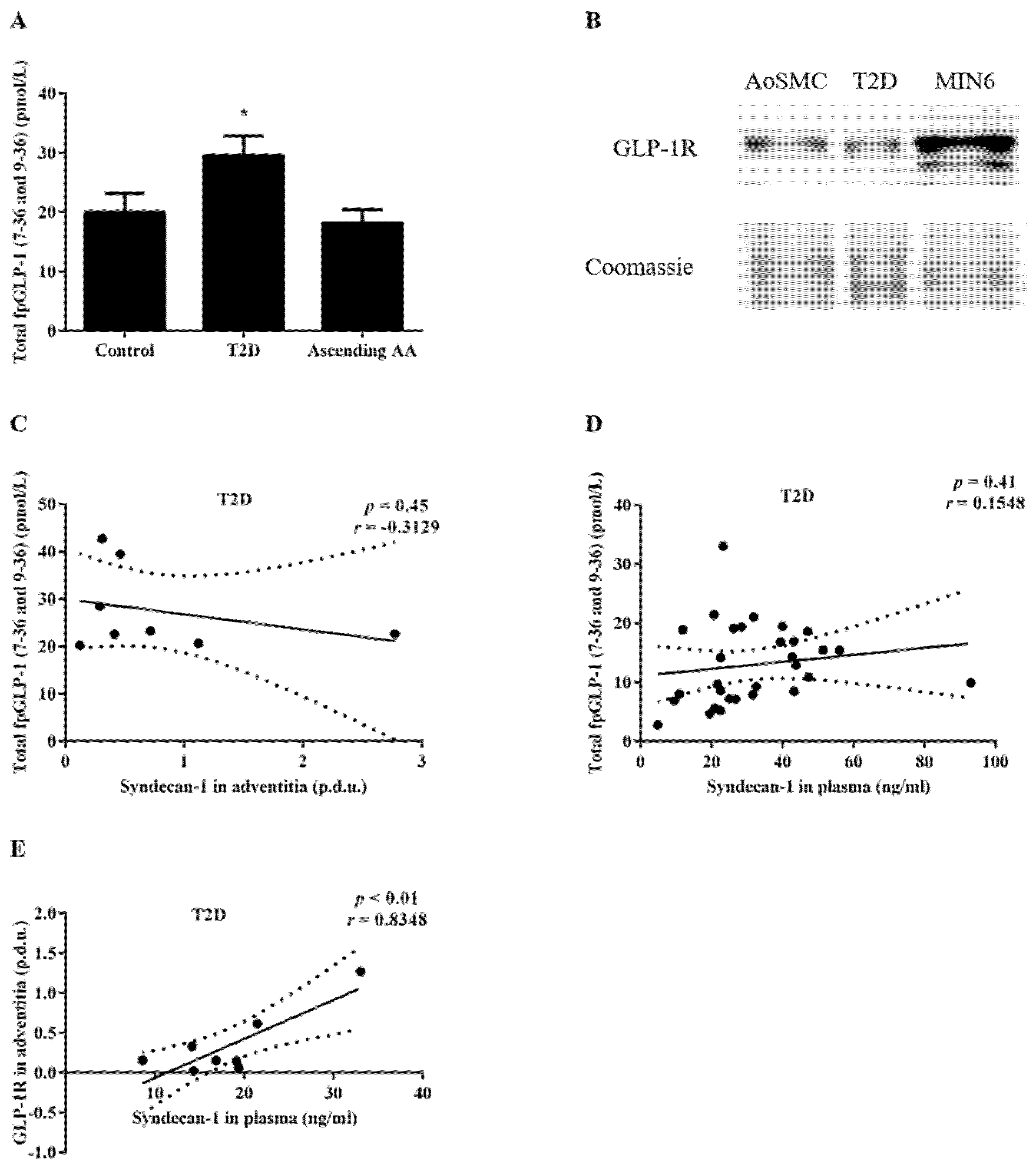
3.2. Increased FpGLP-1 in T2D Is Not Significantly Associated with an Increased Sdc-1 Expression in the Adventitia of Patients with T2D
3.3. The Sdc-1 Is Increased in the Aortic Tissue of Patients with an Ascending AA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuzmik, G.A.; Sang, A.X.; Elefteriades, J.A. Natural history of thoracic aortic aneurysms. J. Vasc. Surg. 2012, 56, 565–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federle, M.P.; Rosado-de-Christenson, M.L.; Raman, S.P.; Carter, B.W.; Woodward, P.J.; Shaaban, A.M. Heart. In Imaging Anatomy: Chest, Abdomen, Pelvis, 2nd ed.; Federle, M.P., Rosado-de-Christenson, M.L., Raman, S.P., Carter, B.W., Woodward, P.J., Shaaban, A.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 336–379. [Google Scholar]
- Hiratzka, L.F.; Bakris, G.L.; Beckman, J.A.; Bersin, R.; Carr, V.; Casey, D., Jr.; Eagle, K.; Hermann, L.; Isselbacher, E.; Kazerooni, E. ACCF. Circulation 2010, 121, 1544. [Google Scholar]
- Saliba, E.; Sia, Y.; Dore, A.; El Hamamsy, I. The ascending aortic aneurysm: When to intervene? Int. J. Cardiol. Heart Vasc. 2015, 6, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.L.; Lin, C.L.; Wu, Y.Y.; Shieh, D.C.; Sung, F.C.; Kao, C.H. Advanced complicated diabetes mellitus is associated with a reduced risk of thoracic and abdominal aortic aneurysm rupture: A population-based cohort study. Diabetes/Metab. Res. Rev. 2015, 31, 190–197. [Google Scholar] [CrossRef]
- Prakash, S.K.; Pedroza, C.; Khalil, Y.A.; Milewicz, D.M. Diabetes and reduced risk for thoracic aortic aneurysms and dissections: A nationwide case-control study. J. Am. Heart Assoc. 2012, 1, e000323. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.Y.; Huang, C.Y.; Shih, C.M.; Chang, W.H.; Tsai, C.S.; Lin, F.Y.; Shih, C.C. Dipeptidyl peptidase-4 inhibitor decreases abdominal aortic aneurysm formation through GLP-1-dependent monocytic activity in mice. PLoS ONE 2015, 10, e0121077. [Google Scholar] [CrossRef]
- Yu, J.; Morimoto, K.; Bao, W.; Yu, Z.; Okita, Y.; Okada, K. Glucagon-like peptide-1 prevented abdominal aortic aneurysm development in rats. Surg. Today 2016, 46, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Takahara, Y.; Tokunou, T.; Ichiki, T. Suppression of Abdominal Aortic Aneurysm Formation in Mice by Teneligliptin, a Dipeptidyl Peptidase-4 Inhibitor. J. Atheroscler. Thromb. 2018, 25, 698–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikedo, T.; Minami, M.; Kataoka, H.; Hayashi, K.; Nagata, M.; Fujikawa, R.; Higuchi, S.; Yasui, M.; Aoki, T.; Fukuda, M.; et al. Dipeptidyl Peptidase-4 Inhibitor Anagliptin Prevents Intracranial Aneurysm Growth by Suppressing Macrophage Infiltration and Activation. J. Am. Heart Assoc. 2017, 6, e004777. [Google Scholar] [CrossRef]
- Goto, H.; Nomiyama, T.; Mita, T.; Yasunari, E.; Azuma, K.; Komiya, K.; Arakawa, M.; Jin, W.L.; Kanazawa, A.; Kawamori, R.; et al. Exendin-4, a glucagon-like peptide-1 receptor agonist, reduces intimal thickening after vascular injury. Biochem. Biophys. Res. Commun. 2011, 405, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Morimoto, K.; Hasegawa, T.; Sasaki, N.; Yamashita, T.; Hirata, K.; Okita, Y.; Okada, K. Orally administered dipeptidyl peptidase-4 inhibitor (alogliptin) prevents abdominal aortic aneurysm formation through an antioxidant effect in rats. J. Vasc. Surg. 2014, 59, 1098–1108. [Google Scholar] [CrossRef] [Green Version]
- El-Hamamsy, I.; Yacoub, M.H. Cellular and molecular mechanisms of thoracic aortic aneurysms. Nat. Rev. Cardiol. 2009, 6, 771–786. [Google Scholar] [CrossRef]
- Petit, C.; Mousavi, S.J.; Avril, S. Chapter 6—Review of the Essential Roles of SMCs in ATAA Biomechanics. In Advances in Biomechanics and Tissue Regeneration; Doweidar, M.H., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 95–114. [Google Scholar]
- Agaimy, A.; Weyand, M.; Strecker, T. Inflammatory thoracic aortic aneurysm (lymphoplasmacytic thoracic aortitis): A 13-year-experience at a German Heart Center with emphasis on possible role of IgG4. Int. J. Clin. Exp. Pathol. 2013, 6, 1713–1722. [Google Scholar] [PubMed]
- Wu, D.; Choi, J.C.; Sameri, A.; Minard, C.G.; Coselli, J.S.; Shen, Y.H.; LeMaire, S.A. Inflammatory Cell Infiltrates in Acute and Chronic Thoracic Aortic Dissection. Aorta 2013, 1, 259–267. [Google Scholar] [CrossRef] [Green Version]
- del Porto, F.; Proietta, M.; Tritapepe, L.; Miraldi, F.; Koverech, A.; Cardelli, P.; Tabacco, F.; de Santis, V.; Vecchione, A.; Mitterhofer, A.P.; et al. Inflammation and immune response in acute aortic dissection. Ann. Med. 2010, 42, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Dinesh, N.E.H.; Reinhardt, D.P. Inflammation in thoracic aortic aneurysms. Herz 2019, 44, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, M.E.; Dietz, H.C. Lessons on the pathogenesis of aneurysm from heritable conditions. Nature 2011, 473, 308–316. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Guo, D.-C.; Estrera, A.L.; Safi, H.J.; Huynh, T.T.; Yin, Z.; Cao, S.-N.; Lin, J.; Kurian, T.; Buja, L.M.; et al. Characterization of the inflammatory and apoptotic cells in the aortas of patients with ascending thoracic aortic aneurysms and dissections. J. Thorac. Cardiovasc. Surg. 2006, 131, 671–678.e672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, K.; Takino, T.; Miyamori, H.; Kinsen, H.; Yoshizaki, T.; Furukawa, M.; Sato, H. Cleavage of syndecan-1 by membrane type matrix metalloproteinase-1 stimulates cell migration. J. Biol. Chem. 2003, 278, 40764–40770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brule, S.; Charnaux, N.; Sutton, A.; Ledoux, D.; Chaigneau, T.; Saffar, L.; Gattegno, L. The shedding of syndecan-4 and syndecan-1 from HeLa cells and human primary macrophages is accelerated by SDF-1/CXCL12 and mediated by the matrix metalloproteinase-9. Glycobiology 2006, 16, 488–501. [Google Scholar] [CrossRef] [Green Version]
- Bernfield, M.; Götte, M.; Park, P.W.; Reizes, O.; Fitzgerald, M.L.; Lincecum, J.; Zako, M. Functions of cell surface heparan sulfate proteoglycans. Annu. Rev. Biochem. 1999, 68, 729–777. [Google Scholar] [CrossRef]
- Xiao, J.; Angsana, J.; Wen, J.; Smith, S.V.; Park, P.W.; Ford, M.L.; Haller, C.A.; Chaikof, E.L. Syndecan-1 displays a protective role in aortic aneurysm formation by modulating T cell-mediated responses. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 386–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaterji, S.; Lam, C.H.; Ho, D.S.; Proske, D.C.; Baker, A.B. Syndecan-1 regulates vascular smooth muscle cell phenotype. PLoS ONE 2014, 9, e89824. [Google Scholar] [CrossRef] [Green Version]
- Yeaman, C.; Rapraeger, A.C. Post-transcriptional regulation of syndecan-1 expression by cAMP in peritoneal macrophages. J. Cell Biol. 1993, 122, 941–950. [Google Scholar] [CrossRef]
- Fehmann, H.C.; Goke, R.; Goke, B. Cell and molecular biology of the incretin hormones glucagon-like peptide-I and glucose-dependent insulin releasing polypeptide. Endocr. Rev. 1995, 16, 390–410. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Philippe, J.; Mojsov, S.; Chick, W.L.; Habener, J.F. Glucagon-like peptide I stimulates insulin gene expression and increases cyclic AMP levels in a rat islet cell line. Proc. Natl. Acad. Sci. USA 1987, 84, 3434–3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelone, D.F.; Wessels, M.R.; Coughlin, M.; Suter, E.E.; Valentini, P.; Kalish, L.A.; Levy, O. Innate Immunity of the Human Newborn Is Polarized Toward a High Ratio of IL-6/TNF-α Production In Vitro and In Vivo. Pediatr. Res. 2006, 60, 205–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krizhanovskii, C.; Ntika, S.; Olsson, C.; Eriksson, P.; Franco-Cereceda, A. Elevated circulating fasting glucagon-like peptide-1 in surgical patients with aortic valve disease and diabetes. Diabetol. Metab. Syndr. 2017, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Treska, V.; Topolcan, O.; Pecen, L. Cytokines as plasma markers of abdominal aortic aneurysm. Clin. Chem. Lab. Med. 2000, 38, 1161–1164. [Google Scholar] [CrossRef]
- Guo, D.C.; Papke, C.L.; He, R.; Milewicz, D.M. Pathogenesis of thoracic and abdominal aortic aneurysms. Ann. N. Y. Acad. Sci. 2006, 1085, 339–352. [Google Scholar] [CrossRef]
- Juvonen, J.; Surcel, H.M.; Satta, J.; Teppo, A.M.; Bloigu, A.; Syrjala, H.; Airaksinen, J.; Leinonen, M.; Saikku, P.; Juvonen, T. Elevated circulating levels of inflammatory cytokines in patients with abdominal aortic aneurysm. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 2843–2847. [Google Scholar] [CrossRef]
- Batra, R.; Suh, M.K.; Carson, J.S.; Dale, M.A.; Meisinger, T.M.; Fitzgerald, M.; Opperman, P.J.; Luo, J.; Pipinos, I.I.; Xiong, W.; et al. IL-1beta (Interleukin-1beta) and TNF-alpha (Tumor Necrosis Factor-alpha) Impact Abdominal Aortic Aneurysm Formation by Differential Effects on Macrophage Polarization. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 457–463. [Google Scholar] [CrossRef]
- Gabrielsson, S.; Soderlund, A.; Nilsson, C.; Lilja, G.; Nordlund, M.; Troye-Blomberg, M. Influence of atopic heredity on IL-4-, IL-12- and IFN-gamma-producing cells in in vitro activated cord blood mononuclear cells. Clin. Exp. Immunol. 2001, 126, 390–396. [Google Scholar] [CrossRef]
- Angsana, J.; Chen, J.; Smith, S.; Xiao, J.; Wen, J.; Liu, L.; Haller, C.A.; Chaikof, E.L. Syndecan-1 Modulates the Motility and Resolution Responses of Macrophages. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 332–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Kan, H.; Boye, A.; Jiang, Y.; Wu, C.; Yang, Y. Mitogen-activated protein kinase inhibitors reduce the nuclear accumulation of phosphorylated Smads by inhibiting Imp 7 or Imp 8 in HepG2 cells. Oncol. Lett. 2018, 15, 4867–4872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Jin, S.; Jin, C.; Jin, Z. Syndecan-1 regulates extracellular matrix expression in keloid fibroblasts via TGF-β1/Smad and MAPK signaling pathways. Life Sci. 2020, 254, 117326. [Google Scholar] [CrossRef] [PubMed]
- Van Gucht, I.; Meester, J.A.N.; Bento, J.R.; Bastiaansen, M.; Bastianen, J.; Luyckx, I.; Van Den Heuvel, L.; Neutel, C.H.G.; Guns, P.J.; Vermont, M.; et al. A human importin-β-related disorder: Syndromic thoracic aortic aneurysm caused by bi-allelic loss-of-function variants in IPO8. Am. J. Hum. Genet. 2021, 108, 1115–1125. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Hasegawa, Y.; Ishigaki, Y.; Yamada, T.; Gao, J.; Imai, J.; Uno, K.; Kaneko, K.; Ogihara, T.; Shimosawa, T.; et al. Importance of endothelial NF-κB signalling in vascular remodelling and aortic aneurysm formation. Cardiovasc. Res. 2012, 97, 106–114. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Liu, J.; Zhang, Z.; Chen, Y. Suppressing Syndecan-1 Shedding Ameliorates Intestinal Epithelial Inflammation through Inhibiting NF-κB Pathway and TNF-α. Gastroenterol. Res. Pract. 2016, 2016, 6421351. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, M.; Mita, T.; Azuma, K.; Ebato, C.; Goto, H.; Nomiyama, T.; Fujitani, Y.; Hirose, T.; Kawamori, R.; Watada, H. Inhibition of monocyte adhesion to endothelial cells and attenuation of atherosclerotic lesion by a glucagon-like peptide-1 receptor agonist, exendin-4. Diabetes 2010, 59, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Manon-Jensen, T.; Itoh, Y.; Couchman, J.R. Proteoglycans in health and disease: The multiple roles of syndecan shedding. FEBS J. 2010, 277, 3876–3889. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.L.; Wang, Z.; Park, P.W.; Murphy, G.; Bernfield, M. Shedding of syndecan-1 and -4 ectodomains is regulated by multiple signaling pathways and mediated by a TIMP-3-sensitive metalloproteinase. J. Cell Biol. 2000, 148, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Foley, E.M.; Gonzales, J.C.; Gordts, P.L.; Li, Y.; Esko, J.D. Shedding of syndecan-1 from human hepatocytes alters very low density lipoprotein clearance. Hepatology 2012, 55, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kind, S.; Merenkow, C.; Büscheck, F.; Möller, K.; Dum, D.; Chirico, V.; Luebke, A.M.; Höflmayer, D.; Hinsch, A.; Jacobsen, F.; et al. Prevalence of Syndecan-1 (CD138) Expression in Different Kinds of Human Tumors and Normal Tissues. Dis. Markers 2019, 2019, 4928315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledin, J.; Staatz, W.; Li, J.P.; Götte, M.; Selleck, S.; Kjellén, L.; Spillmann, D. Heparan sulfate structure in mice with genetically modified heparan sulfate production. J. Biol. Chem. 2004, 279, 42732–42741. [Google Scholar] [CrossRef] [Green Version]
- Henry-Stanley, M.J.; Zhang, B.; Erlandsen, S.L.; Wells, C.L. Synergistic effect of tumor necrosis factor-alpha and interferon-gamma on enterocyte shedding of syndecan-1 and associated decreases in internalization of Listeria monocytogenes and Staphylococcus aureus. Cytokine 2006, 34, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.H.-F.; Aquino, R.S.; Park, P.W. Molecular functions of syndecan-1 in disease. Matrix Biol. 2012, 31, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Fritchley, S.J.; Kirby, J.A.; Ali, S. The antagonism of interferon-gamma (IFN-gamma) by heparin: Examination of the blockade of class II MHC antigen and heat shock protein-70 expression. Clin. Exp. Immunol. 2000, 120, 247–252. [Google Scholar] [CrossRef]
- Araújo, F.; Fonte, P.; Santos, H.A.; Sarmento, B. Oral delivery of glucagon-like peptide-1 and analogs: Alternatives for diabetes control? J. Diabetes Sci. Technol. 2012, 6, 1486–1497. [Google Scholar] [CrossRef] [Green Version]
- Holst, J.J.; Deacon, C.F. Glucagon-like peptide-1 mediates the therapeutic actions of DPP-IV inhibitors. Diabetologia 2005, 48, 612–615. [Google Scholar] [CrossRef]
- Schmitt, R.; Tscheuschler, A.; Laschinski, P.; Uffelmann, X.; Discher, P.; Fuchs, J.; Kreibich, M.; Peyronnet, R.; Kari, F.A. A potential key mechanism in ascending aortic aneurysm development: Detection of a linear relationship between MMP-14/TIMP-2 ratio and active MMP-2. PLoS ONE 2019, 14, e0212859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taketani, T.; Imai, Y.; Morota, T.; Maemura, K.; Morita, H.; Hayashi, D.; Yamazaki, T.; Nagai, R.; Takamoto, S. Altered patterns of gene expression specific to thoracic aortic aneurysms. Int. Heart J. 2005, 46, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamin, M.M.; Khalil, R.A. Matrix metalloproteinase inhibitors as investigative tools in the pathogenesis and management of vascular disease. Exp. Suppl. 2012, 103, 209–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, A.H.; Hayashida, K.; Park, P.W. Molecular and cellular mechanisms of syndecans in tissue injury and inflammation. Mol. Cells 2007, 24, 153–166. [Google Scholar]
- Day, R.M.; Mitchell, T.J.; Knight, S.C.; Forbes, A. Regulation of epithelial syndecan-1 expression by inflammatory cytokines. Cytokine 2003, 21, 224–233. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Group | Number of Patients | Mean Age (± Standard error of Mean, SEM) | Gender (Male/Female) | Valve Pathology (Aortic Stenosis/Aortic Insufficiency) 1 |
|---|---|---|---|---|
| Control | 43 | 67.6 ± 2.1 | 26/17 | 24/15 |
| Ascending AA | 41 | 65.3 ± 1.7 | 26/15 | 3/29 |
| T2D | 33 | 71.7 ± 0.94 | 27/6 | 30/1 |
| Total Patients | 117 | 62.1 ± 1.1 | 79/38 | 57/45 |
| Correlation With Sdc-1 in the Adventitia | IFN-γ (pg/mL) | IL-1β (pg/mL) | IL-4 (pg/mL) | IL-5 (pg/mL) | IL-6 (pg/mL) | IL-12p70 (pg/mL) | TNF-α (pg/mL) |
|---|---|---|---|---|---|---|---|
| r (T2D patients) | 0.7136 | −0.0811 | 0.0879 | 0.1359 | 0.0843 | 0.9201 | 0.1865 |
| p value (T2D patients) | 0.07 | 0.88 | 0.85 | 0.77 | 0.86 | < 0.01 | 0.66 |
| r (control and T2D patients) | 0.1270 | −0.09170 | 0.1870 | 0.04293 | 0.03023 | 0.7263 | −0.1046 |
| p value (control and T2D patients) | 0.6394 | 0.7657 | 0.5046 | 0.8700 | 0.9083 | < 0.01 | 0.6796 |
| Correlation with Sdc-1 in the Adventitia | IFN-γ (pg/mL) | IL-1β (pg/mL) | IL-4 (pg/mL) | IL-5 (pg/mL) | IL-6 (pg/mL) | IL-12p70 (pg/mL) | TNF-α (pg/mL) | Sdc-1 in plasma (ng/mL) | FpGLP-1 (pmol/L) |
|---|---|---|---|---|---|---|---|---|---|
| r (ascending AA patients) | −0.2109 | −0.1219 | 0.2103 | −0.5401 | −0.5252 | −0.3461 | 0.3337 | 0.0983 | −0.0590 |
| p value (ascending AA patients) | 0.65 | 0.77 | 0.62 | 0.13 | 0.15 | 0.45 | 0.38 | 0.82 | 0.87 |
| r (control and ascending AA patients) | −0.2746 | −0.1686 | −0.0929 | −0.3476 | −0.1768 | −0.1496 | −0.0191 | −0.0777 | −0.2037 |
| p value (control and ascending AA patients) | 0.30 | 0.55 | 0.73 | 0.14 | 0.47 | 0.61 | 0.94 | 0.74 | 0.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntika, S.; Tracy, L.M.; Franco-Cereceda, A.; Björck, H.M.; Krizhanovskii, C. Syndecan-1 Expression Is Increased in the Aortic Wall of Patients with Type 2 Diabetes but Is Unrelated to Elevated Fasting Plasma Glucagon-Like Peptide-1. Biomedicines 2021, 9, 697. https://doi.org/10.3390/biomedicines9060697
Ntika S, Tracy LM, Franco-Cereceda A, Björck HM, Krizhanovskii C. Syndecan-1 Expression Is Increased in the Aortic Wall of Patients with Type 2 Diabetes but Is Unrelated to Elevated Fasting Plasma Glucagon-Like Peptide-1. Biomedicines. 2021; 9(6):697. https://doi.org/10.3390/biomedicines9060697
Chicago/Turabian StyleNtika, Stelia, Linda M. Tracy, Anders Franco-Cereceda, Hanna M. Björck, and Camilla Krizhanovskii. 2021. "Syndecan-1 Expression Is Increased in the Aortic Wall of Patients with Type 2 Diabetes but Is Unrelated to Elevated Fasting Plasma Glucagon-Like Peptide-1" Biomedicines 9, no. 6: 697. https://doi.org/10.3390/biomedicines9060697