Decitabine Inhibits Bone Resorption in Periodontitis by Upregulating Anti-Inflammatory Cytokines and Suppressing Osteoclastogenesis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Reagents
2.2. Ligature-Induced Periodontitis Model
2.3. Analysis of Periodontal Bone Loss
2.4. Staining for Tartrate-Resistant Acid Phosphatase (TRAP)
2.5. Immunohistochemistry
2.6. Osteoclast Differentiation
2.7. Osteoblastic Cell Line MC3T3-E1 Culture and Osteogenic Differentiation
2.8. Real-Time PCR
2.9. Luciferase Assay
2.10. Statistical Analysis
3. Results
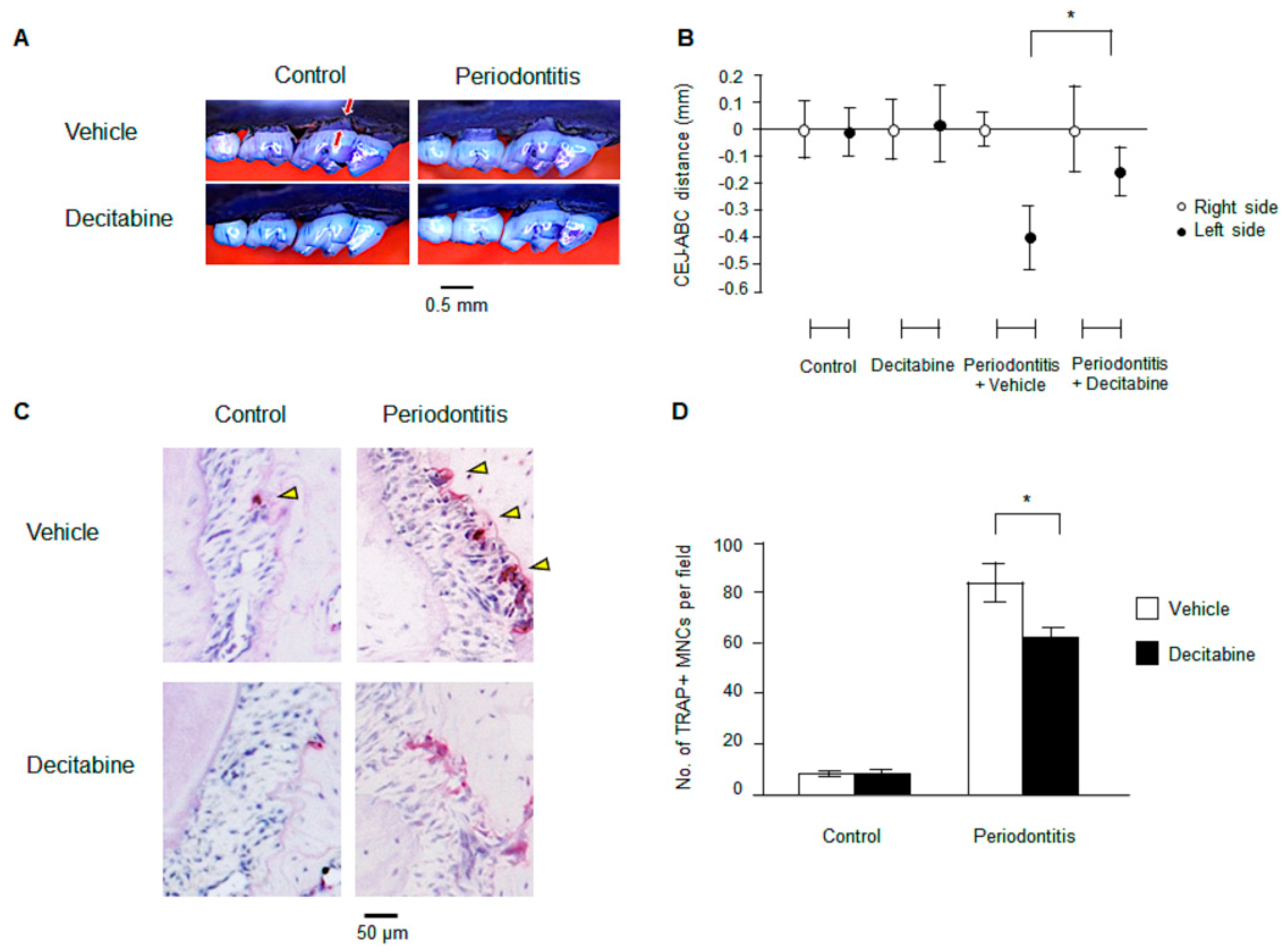
3.1. Decitabine Inhibits Bone Resorption in Mice with Ligature-Induced Periodontitis
3.2. Decitabine Suppresses Osteoclastogenesis in Mice with Ligature-Induced Periodontitis
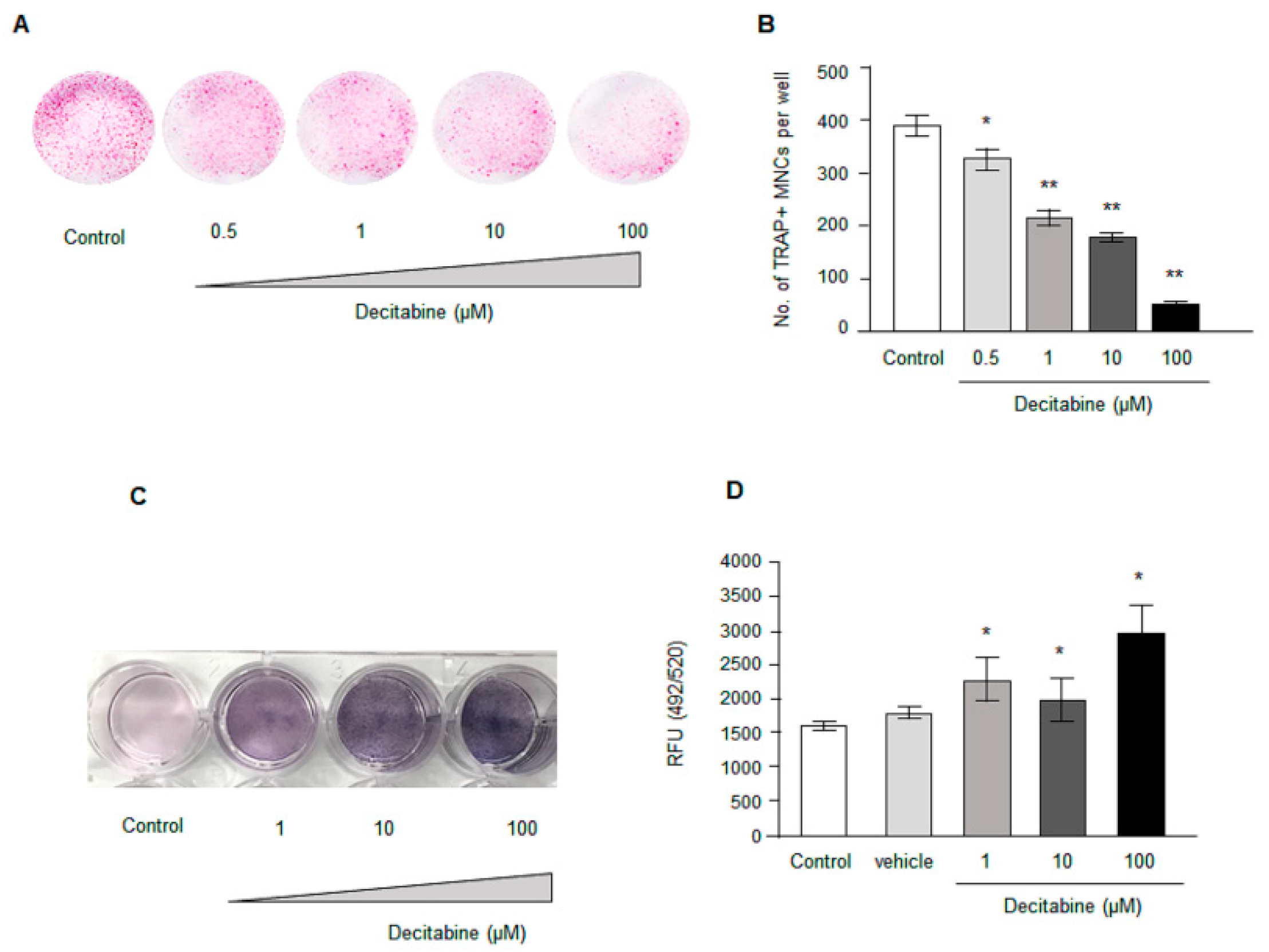
3.3. Decitabine Decreases Osteoclastogenesis in Human CD14+ Monocytes In Vitro in a Dose-Dependent Manner
3.4. Decitabine Increases Osteogenic Differentiation in Osteoblast Precursor Cell Line MC3CT3-E1 in a Dose Dependent Manner
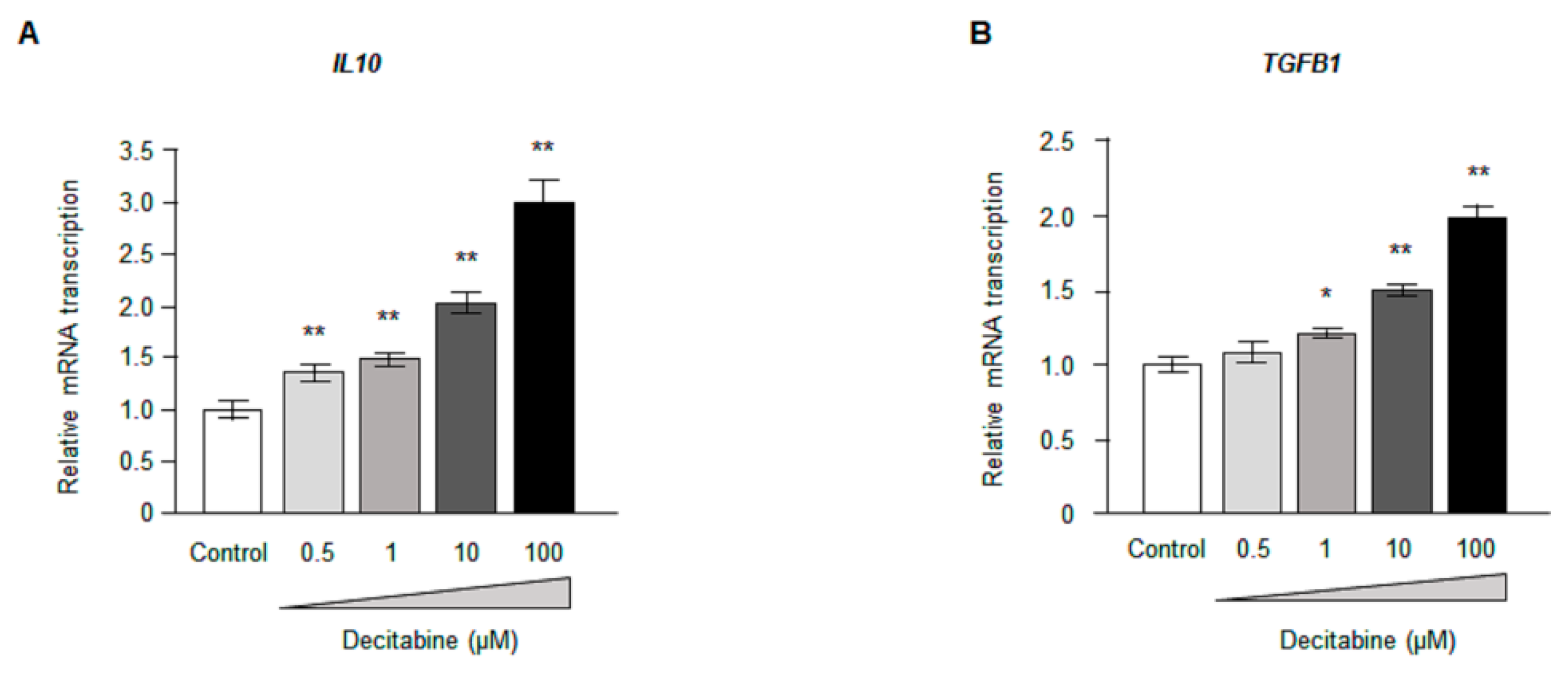
3.5. Decitabine Increases the Expression of Anti-Inflammatory Cytokines in CD14+ Monocytes
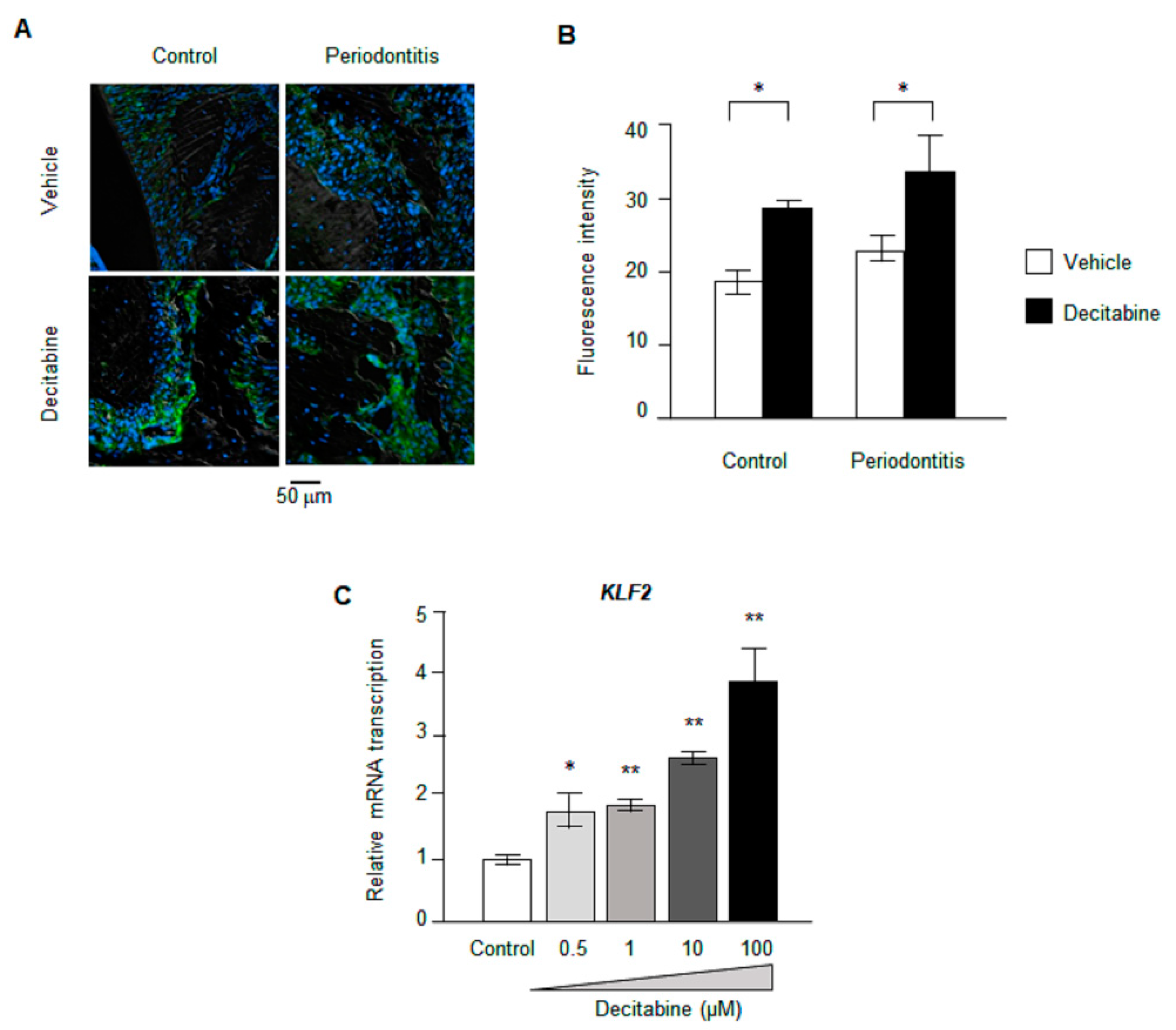
3.6. Decitabine Increases the Expression of KLF2
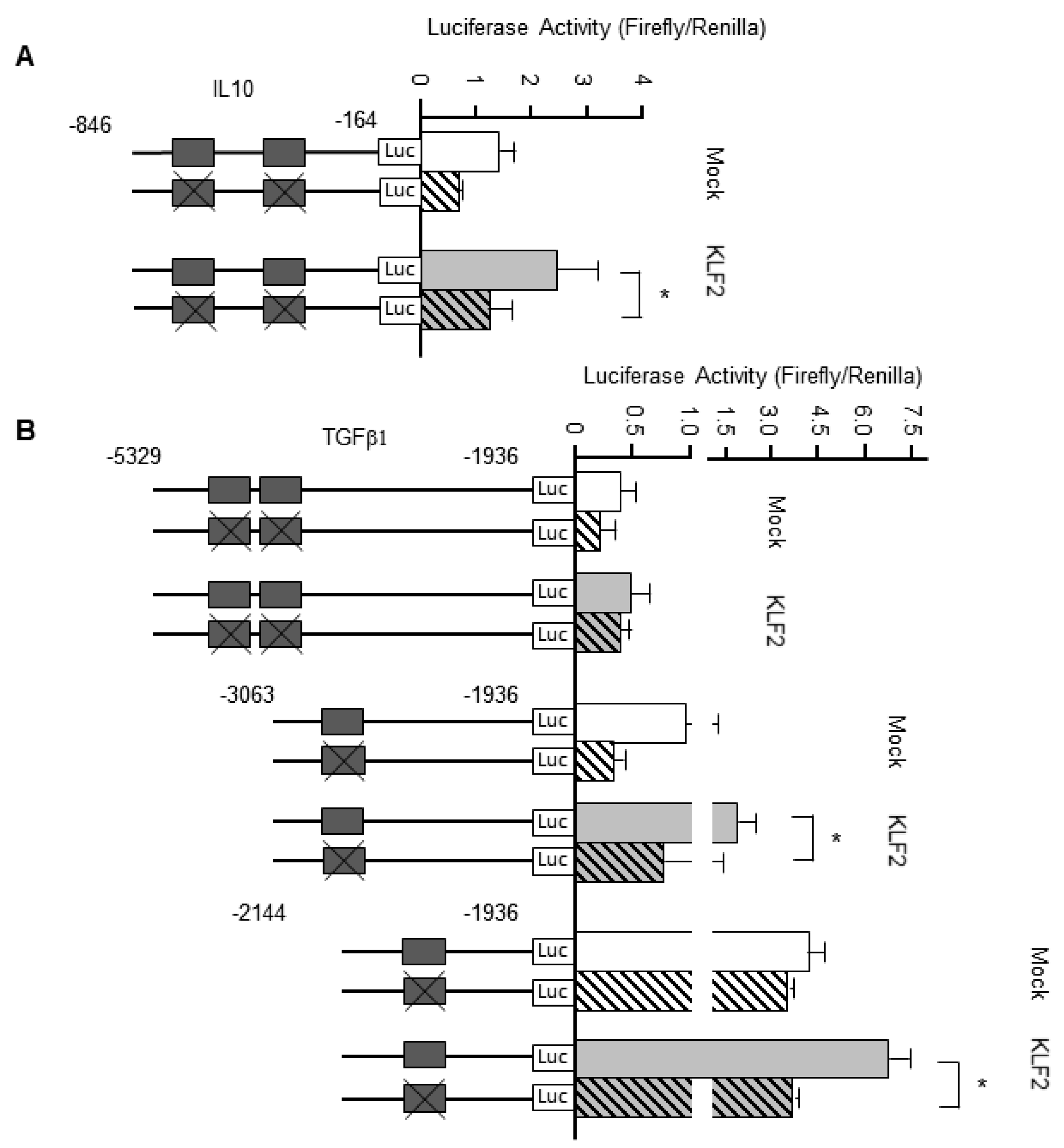
3.7. KLF2 Upregulates the Transcription of Genes Encoding Anti-Inflammatory Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tonetti, M.S.; Claffey, N. European Workshop in Periodontology group, Advances in the progression of periodontitis and proposal of definitions of a periodontitis case and disease progression for use in risk factor re-search. Group C consensus report of the 5th European Workshop in Periodontology. J. Clin. Periodontol. 2005, 32, 210–213. [Google Scholar]
- Stabholz, A.; Soskolne, W.A.; Shapira, L. Genetic and environmental risk factors for chronic periodontitis and aggressive periodontitis. Periodontol. 2000 2010, 53, 138–153. [Google Scholar] [CrossRef] [PubMed]
- Graziani, F.; Karapetsa, D.; Alonso, B.; Herrera, D. Nonsurgical and surgical treatment of periodontitis: How many options for one disease? J. Periodontol. 2017, 75, 152–188. [Google Scholar] [CrossRef]
- Kitamura, M.; Akamatsu, M.; Kawanami, M.; Furuichi, Y.; Fujii, T.; Mori, M.; Kunimatsu, K.; Shimauchi, H.; Ogata, Y.; Yamamoto, M.; et al. Randomized Placebo-Controlled and Controlled Non-Inferiority Phase III Trials Comparing Trafermin, a Recombinant Human Fibroblast Growth Factor 2, and Enamel Matrix Derivative in Periodontal Regeneration in Intrabony Defects. J. Bone Miner. Res. 2016, 31, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Ramseier, C.A.; Rasperini, G.; Batia, S.; Giannobile, W.V. Advanced reconstructive technologies for periodontal tissue repair. Periodontol. 2000 2012, 59, 185–202. [Google Scholar] [CrossRef] [Green Version]
- Tonetti, M.S.; Lang, N.P.; Cortellini, P.; Suvan, J.E.; Adriaens, P.; Dubravec, D.; Fonzar, A.; Fourmousis, I.; Mayfield, L.; Rossi, R.; et al. Enamel matrix proteins in the regenerative therapy of deep intrabony defects. J. Clin. Periodontol. 2002, 29, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Benakanakere, M.; Abdolhosseini, M.; Hosur, K.; Finoti, L.; Kinane, D. TLR2 Promoter Hypermethylation Creates Innate Immune Dysbiosis. J. Dent. Res. 2015, 94, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Palioto, D.B.; Finoti, L.S.; Kinane, D.F.; Benakanakere, M. Epigenetic and inflammatory events in experimental periodontitis following systemic microbial challenge. J. Clin. Periodontol. 2019, 46, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Almiñana-Pastor, P.J.; Boronat-Catalá, M.; Micó-Martinez, P.; Bellot-Arcís, C.; Lopez-Roldan, A.; Alpiste-Illueca, F.M.; Lopez-Rolda, A. Epigenetics and periodontics: A systematic review. Medicina Oral Patología Oral y Cirugia Bucal 2019, 24, e659–e672. [Google Scholar] [CrossRef] [PubMed]
- Andia, D.C.; De Oliveira, N.F.; Casarin, R.C.; Casati, M.Z.; Line, S.R.; De Souza, A.P. DNA Methylation Status of theIL8Gene Promoter in Aggressive Periodontitis. J. Periodontol. 2010, 81, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, A.M.; Rocha, L.P.C.; Amormino, S.A.D.F.; Gomes, C.C.; Dutra, W.O.; Gomez, R.S.; Da Costa, J.E.; Moreira, P.R. DNA methylation profile of genes related to immune response in generalized periodontitis. J. Periodontal Res. 2020, 55, 426–431. [Google Scholar] [CrossRef]
- Amormino, S.A.D.F.; Arão, T.C.; Saraiva, A.M.; Gomez, R.S.; Dutra, W.O.; Da Costa, J.E.; Silva, J.D.F.C.; Moreira, P.R. Hypermethylation and low transcription of TLR2 gene in chronic periodontitis. Hum. Immunol. 2013, 74, 1231–1236. [Google Scholar] [CrossRef]
- Zhang, S.; Barros, S.P.; Moretti, A.J.; Yu, N.; Zhou, J.; Preisser, J.S.; Niculescu, M.D.; Offenbacher, S. Epigenetic regulation of TNFA expression in periodontal disease. J. Periodontol. 2013, 84, 1606–1616. [Google Scholar] [PubMed] [Green Version]
- Gore, S.D.; Jones, C.; Kirkpatrick, P. Decitabine. Nat. Rev. Drug Discov. 2006, 5, 891–892. [Google Scholar] [CrossRef] [PubMed]
- Kantarjian, H.; Issa, J.P.; Rosenfeld, C.S.; Bennett, J.M.; Albitar, M.; DiPersio, J.; Klimek, V.; Slack, J.; de Castro, C.; Ravandi, F.; et al. Decitabine improves patient outcomes in myelodysplastic syndromes: Results of a phase III randomized study. Cancer 2006, 106, 1794–1803. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, M. Myelodysplastic Syndromes. N. Engl. J. Med. 2020, 383, 1358–1374. [Google Scholar] [CrossRef] [PubMed]
- Makishima, H.; Yoshizato, T.; Yoshida, K.; A Sekeres, M.; Radivoyevitch, T.; Suzuki, H.; Przychodzen, B.; Nagata, Y.; Meggendorfer, M.; Sanada, M.; et al. Dynamics of clonal evolution in myelodysplastic syndromes. Nat. Genet. 2017, 49, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Fenaux, P.; Mufti, G.J.; Hellstrom-Lindberg, E.; Santini, V.; Finelli, C.; Giagounidis, A.; Schoch, R.; Gattermann, N.; Sanz, G.; List, A.; et al. Efficacy of azacitidine compared with that of conventional care regimens in the treatment of higher-risk myelodysplastic syndromes: A randomised, open-label, phase III study. Lancet Oncol. 2009, 10, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Christman, J.K. 5-Azacytidine and 5-aza-2′-deoxycytidine as inhibitors of DNA methylation: Mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibney, E.R.; Nolan, C.M. Epigenetics and gene expression. Heredity 2010, 105, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A Landscape Takes Shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steensma, D.P.; Baer, M.R.; Slack, J.L.; Buckstein, R.; Godley, L.A.; Garcia-Manero, G.; Albitar, M.; Larsen, J.S.; Arora, S.; Cullen, M.T.; et al. Multicenter Study of Decitabine Administered Daily for 5 Days Every 4 Weeks to Adults With Myelodysplastic Syndromes: The Alternative Dosing for Outpatient Treatment (ADOPT) Trial. J. Clin. Oncol. 2009, 27, 3842–3848. [Google Scholar] [CrossRef]
- Zhang, Z.; He, Q.; Tao, Y.; Guo, J.; Xu, F.; Wu, L.-Y.; Zhao, Y.-S.; Wu, N.; Zhou, L.-Y.; Su, J.-Y.; et al. Decitabine treatment sensitizes tumor cells to T-cell-mediated cytotoxicity in patients with myelodysplastic syndromes. Am. J. Transl. Res. 2017, 9, 454–465. [Google Scholar]
- Abe, T.; Hajishengallis, G. Optimization of the ligature-induced periodontitis model in mice. J. Immunol. Methods 2013, 394, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Guo, J.; Ou, H.; Deng, Y.; Jiao, F. Lipopolysaccharide Downregulates Kruppel-Like Factor 2 (KLF2) via Inducing DNMT1-Mediated Hypermethylation in Endothelial Cells. Inflammation 2017, 40, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Barros, S.P.; Offenbacher, S. Modifiable risk factors in periodontal disease: Epigenetic regulation of gene ex-pression in the inflammatory response. J. Periodontol. 2014, 64, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Bobetsis, Y.; Barros, S.; Lin, D.; Weidman, J.; Dolinoy, D.; Jirtle, R.; Boggess, K.; Beck, J.; Offenbacher, S. Bacterial Infection Promotes DNA Hypermethylation. J. Dent. Res. 2007, 86, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Lindroth, A.M.; Park, Y.J. Epigenetic biomarkers: A step forward for understanding periodontitis. J. Periodontal Implant. Sci. 2013, 43, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, H.; Mi, B.; Li, Y.; Wu, W.; Tan, P.; Fang, Z.; Li, J.; Zhang, Y.; Li, F. Decitabine represses osteoclastogenesis through inhibition of RANK and NF-kappaB. Cell Signal. 2015, 27, 969–977. [Google Scholar] [CrossRef]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.-I.; et al. Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Teitelbaum, S.L. Bone Resorption by Osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Al-Rasheed, A.; Scheerens, H.; Rennick, D.M.; Fletcher, H.M.; Tatakis, D.N. Accelerated Alveolar Bone Loss in Mice Lacking Interleukin-10. J. Dent. Res. 2003, 82, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Al-Rasheed, A.; Scheerens, H.; Srivastava, A.K.; Rennick, D.M.; Tatakis, D.N. Accelerated alveolar bone loss in mice lacking interleukin-10: Late onset. J. Periodontal Res. 2004, 39, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Li, Z.; Jiang, H.; Cao, Z.; Liu, J.; Zhang, X. Gene Modification of Transforming Growth Factor beta (TGF-beta) and Interleukin 10 (IL-10) in Sup-pressing Mt Sonicate Induced Osteoclast Formation and Bone Absorption. Med. Sci. Monit. 2018, 24, 5200–5207. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Oh, Y.; Jo, S.; Kim, T.H.; Ji, J.D. A dual role of TGF-beta in human osteoclast differentiation mediated by Smad1 versus Smad3 sig-naling. Immunol. Lett. 2019, 206, 33–40. [Google Scholar] [CrossRef]
- Contaldo, M.; Itro, A.; Lajolo, C.; Gioco, G.; Inchingolo, F.; Serpico, R. Overview on Osteoporosis, Periodontitis and Oral Dysbiosis: The Emerging Role of Oral Mi-crobiota. Appl. Sci. 2020, 10, 6000. [Google Scholar] [CrossRef]
- Potempa, J.; Mydel, P.; Koziel, J. The case for periodontitis in the pathogenesis of rheumatoid arthritis. Nat. Rev. Rheumatol. 2017, 13, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Das, H.; Kumar, A.; Lin, Z.; Patino, W.D.; Hwang, P.M.; Feinberg, M.W.; Majumder, P.K.; Jain, M.K. Kruppel-like factor 2 (KLF2) regulates proinflammatory activation of monocytes. Proc. Natl. Acad. Sci. USA 2006, 103, 6653–6658. [Google Scholar] [CrossRef] [Green Version]
- Nayak, L.; Goduni, L.; Takami, Y.; Sharma, N.; Kapil, P.; Jain, M.K.; Mahabeleshwar, G.H. Kruppel-Like Factor 2 Is a Transcriptional Regulator of Chronic and Acute Inflammation. Am. J. Pathol. 2013, 182, 1696–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.; Wang, Z.; Tao, Y.; Bai, J.; Yu, B.; Shen, J.; Sun, H.; Xiao, L.; Xu, Y.; Zhou, J.; et al. KLF2 regulates osteoblast differentiation by targeting of Runx2. Lab. Investig. 2018, 99, 271–280. [Google Scholar] [CrossRef]
- Welch, J.S.; Petti, A.A.; Miller, C.A.; Fronick, C.C.; O’Laughlin, M.; Fulton, R.S.; Wilson, R.K.; Baty, J.D.; Duncavage, E.J.; Tandon, B.; et al. TP53 and Decitabine in Acute Myeloid Leukemia and Myelodysplastic Syndromes. N. Engl. J. Med. 2016, 375, 2023–2036. [Google Scholar] [CrossRef] [PubMed]
- Dinardo, C.D.; Pratz, K.; Pullarkat, V.; Jonas, B.A.; Arellano, M.; Becker, P.S.; Frankfurt, O.; Konopleva, M.; Wei, A.H.; Kantarjian, H.M.; et al. Venetoclax combined with decitabine or azacitidine in treatment-naive, elderly patients with acute myeloid leukemia. Blood 2019, 133, 7–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, U.; Kajioka, S.; Finoti, L.S.; Palioto, D.B.; Kinane, D.F.; Benakanakere, M.R. Decitabine Inhibits Bone Resorption in Periodontitis by Upregulating Anti-Inflammatory Cytokines and Suppressing Osteoclastogenesis. Biomedicines 2021, 9, 199. https://doi.org/10.3390/biomedicines9020199
Tanaka U, Kajioka S, Finoti LS, Palioto DB, Kinane DF, Benakanakere MR. Decitabine Inhibits Bone Resorption in Periodontitis by Upregulating Anti-Inflammatory Cytokines and Suppressing Osteoclastogenesis. Biomedicines. 2021; 9(2):199. https://doi.org/10.3390/biomedicines9020199
Chicago/Turabian StyleTanaka, Urara, Shunichi Kajioka, Livia S. Finoti, Daniela B. Palioto, Denis F. Kinane, and Manjunatha R. Benakanakere. 2021. "Decitabine Inhibits Bone Resorption in Periodontitis by Upregulating Anti-Inflammatory Cytokines and Suppressing Osteoclastogenesis" Biomedicines 9, no. 2: 199. https://doi.org/10.3390/biomedicines9020199