

Involvement of Protease-Activated Receptor2 Pleckstrin Homology Binding Domain in Ovarian Cancer: Expression in Fallopian Tubes and Drug Design

Abstract
:1. Introduction
 -catenin signaling). In this context, we recently described RNF43 as a novel negative regulator of PAR2, capable of inhibiting cell surface PAR2 expression and the PAR-induced β-catenin stabilization pathway [17]. As a result, PAR2 joins FZDs as a target for RNF43 negative regulation.
-catenin signaling). In this context, we recently described RNF43 as a novel negative regulator of PAR2, capable of inhibiting cell surface PAR2 expression and the PAR-induced β-catenin stabilization pathway [17]. As a result, PAR2 joins FZDs as a target for RNF43 negative regulation.2. Materials and Methods
2.1. Animal Models
2.2. Human Tissues
2.3. Patient Data Analysis
2.4. Cell Culture
2.5. Cell transfections, Plasmids, and PAR Activation
2.6. Small Hairpin (sh) RNA Preparation and Generation of Viral Particles
2.7. Real-Time PCR (qRT PCR) and RT PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LGR5 | F: 5′ CCAACCTCAGCGTCTTCACC 3′ R: 5′ GGAGACTGGGCAGGGGATT 3′ |
| CD44 | F: 5′ CTGGGGACTCTGCCTCGT 3′ R: 5′ ACGTGGAATACACCTGCAAAGC 3′ |
| ALDH1 | F: 5′ CACGCCAGACTTACCTGTCC 3′ R: 5′ TGCCACTCACTGAATCATGCC 3′ |
| Oct-4 | F: 5′-GAG AATTT GTT CCT GCA GTG C-3′ R: 5′-GTT CCC AAT TCC TTC CTT AGT G-3′ |
| CD166 | F: 5′ GATCTCCGCCACCGTCTTC 3′ R: 5′ CGTCAAGTCGGCAAGGTATGG 3′ |
| Par2 | F: 5′-GGC CAA TCT GGC CTT GGC TGA C-3′ R: 5′-GGC AGG AAT GAA GAT GGT CTG-3′ |
| Par4 | F: 5′ CCCAGCGTCTACGACGAGA 3′ R: 5′ CACAGACTTGGCCTGGGTAG 3′ |
| GAPD | F: 5′-CCA CCC ATG GCA AAT TCC ATG GCA-3 R: 5′-TCT AGA CGG CAG GTC AGG TCC ACC-3′ |
2.8. Preparation of Cell Lysates, Immunoprecipitation, and Western Blot
2.9. Matrigel Invasion Assay
2.10. Peptide Synthesis
2.11. Pc(4-4) Cyclic Peptide
2.12. Tumor Xenograft Mouse Model
2.13. Immunohistochemistry
2.14. Statistical Analysis
3. Results
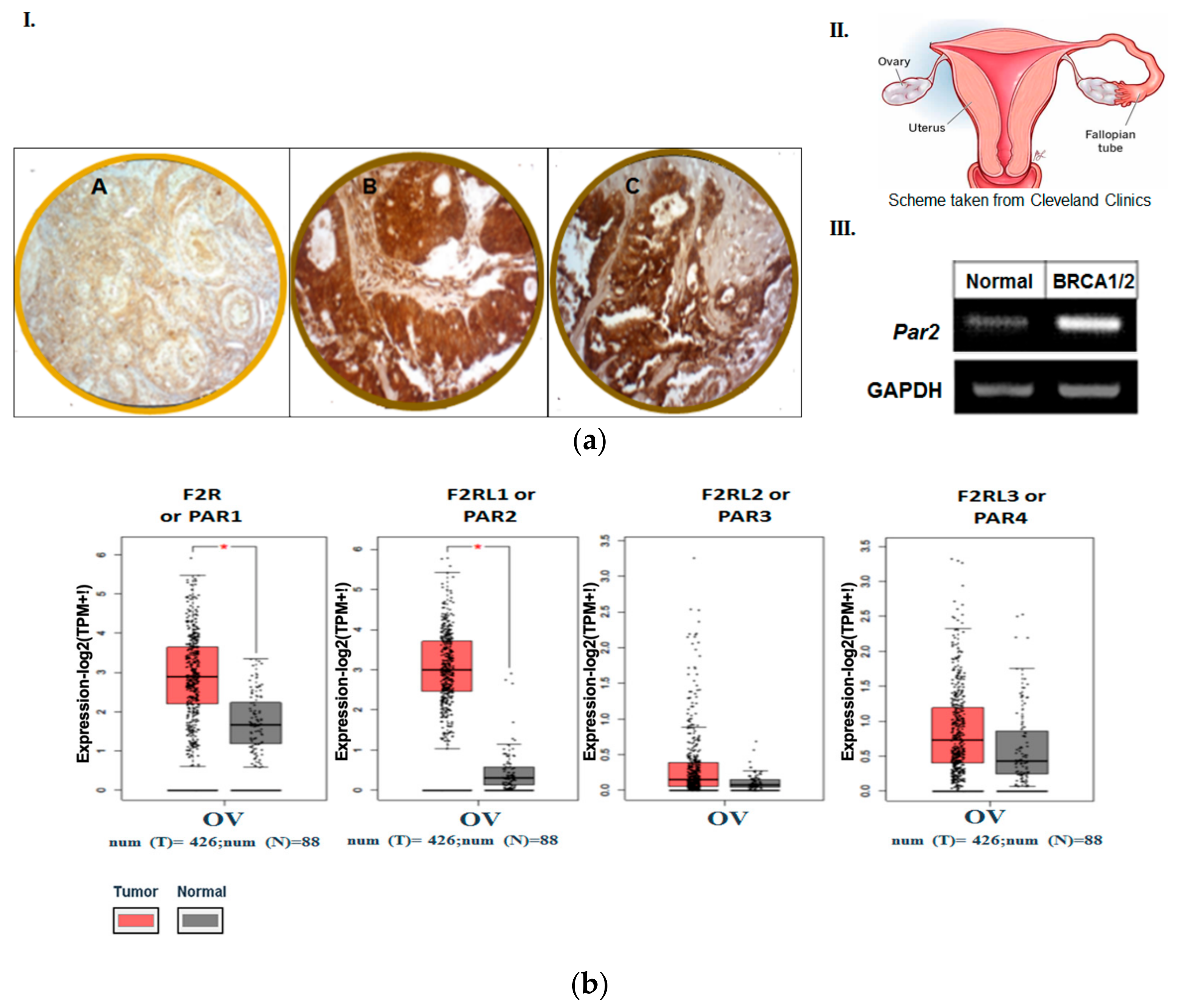
3.1. PAR2 Is Highly Expressed in FTs of BRCA Carriers
3.2. The Pattern of Members of PAR Family in Human Ovarian Tumors
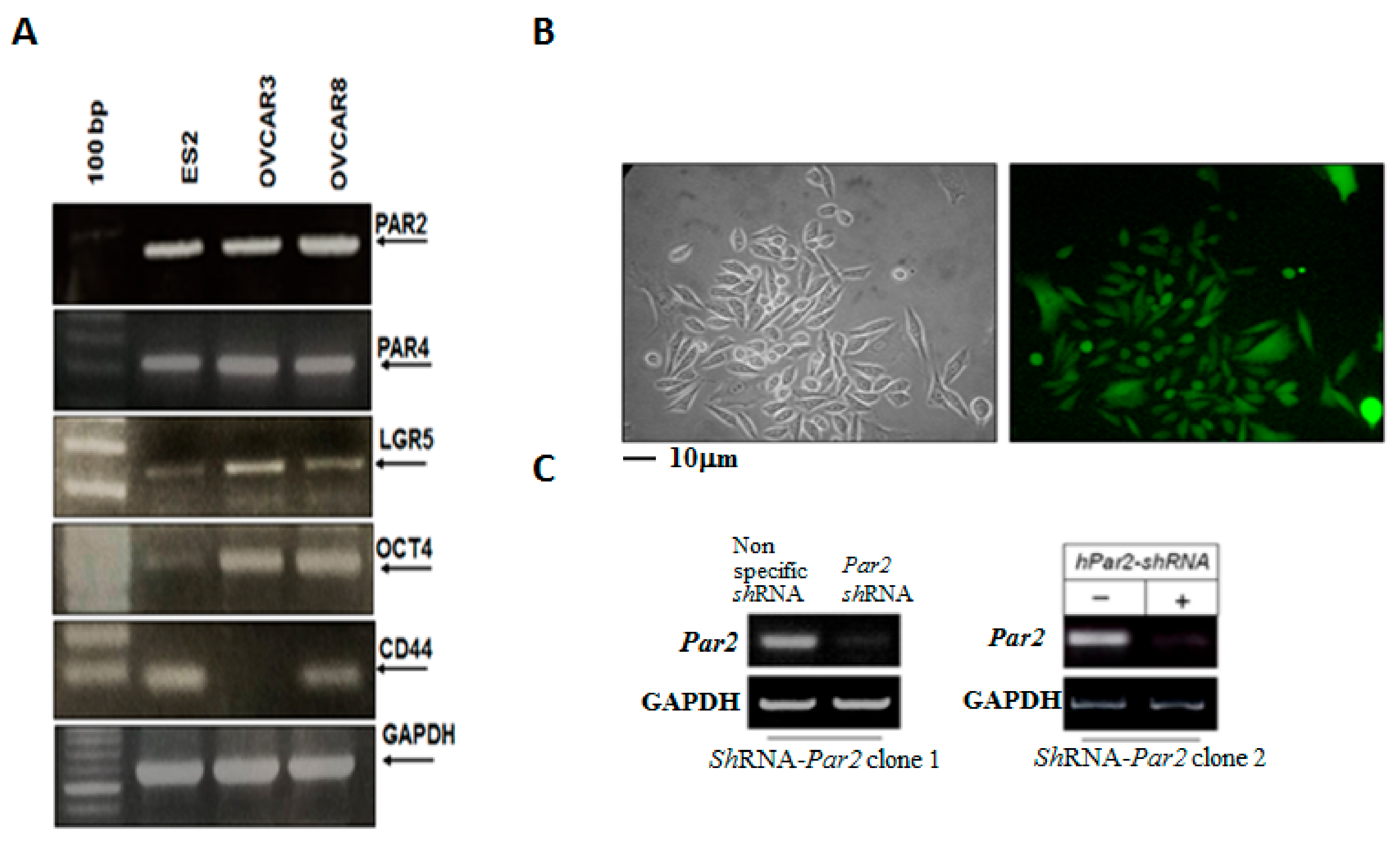
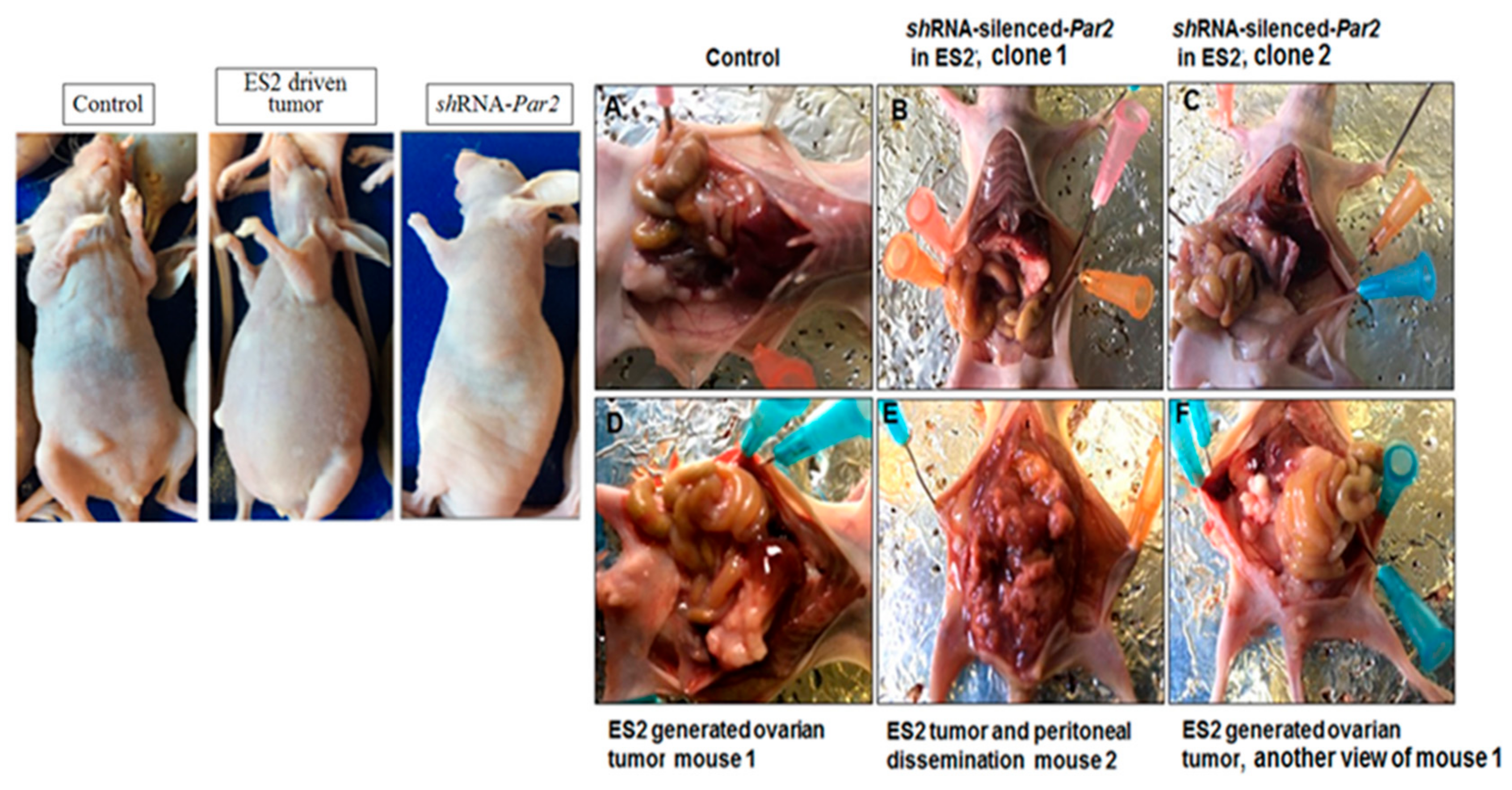
3.3. Knockdown of Par2/f2rl1 Markedly Inhibits Stem Cell Markers In Vitro and Ovarian Cancer Metastasis In Vivo
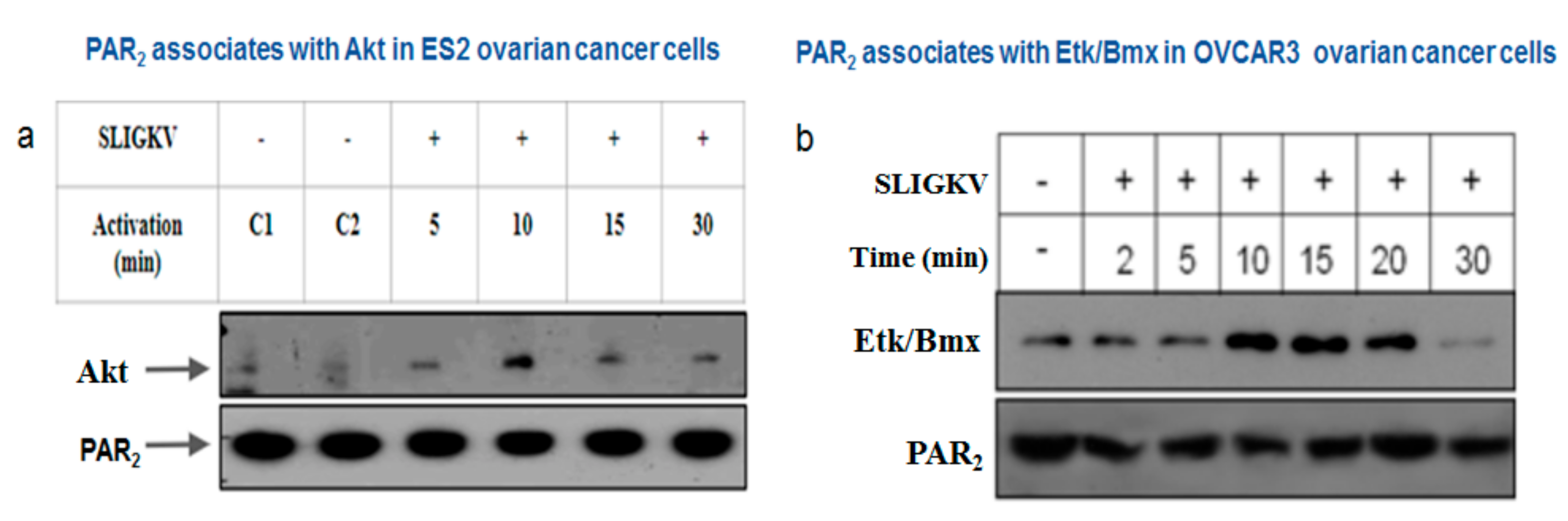
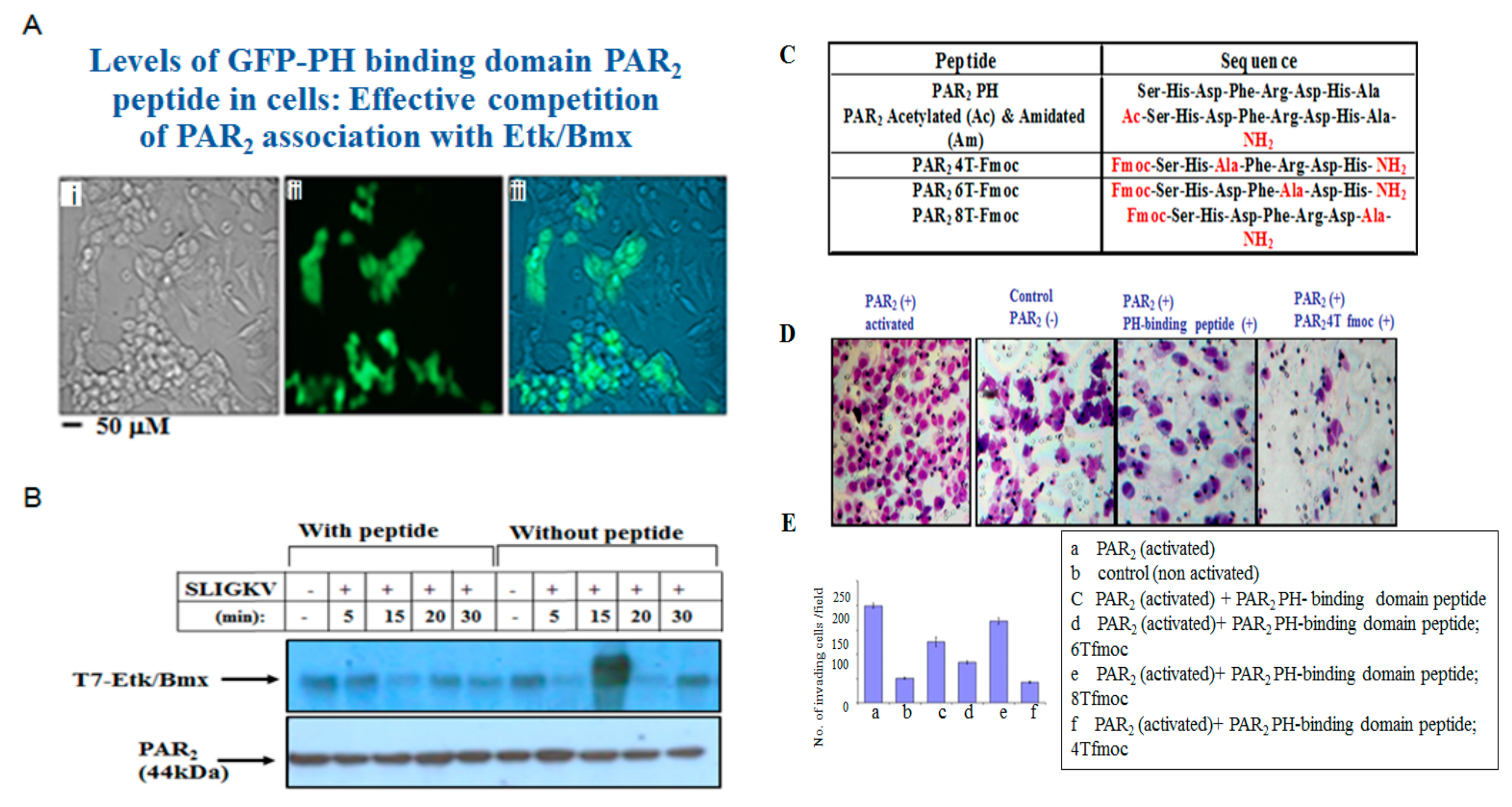
3.4. PAR2-PH Binding Motif and Signaling Partners in Ovarian Cancer Cell Lines
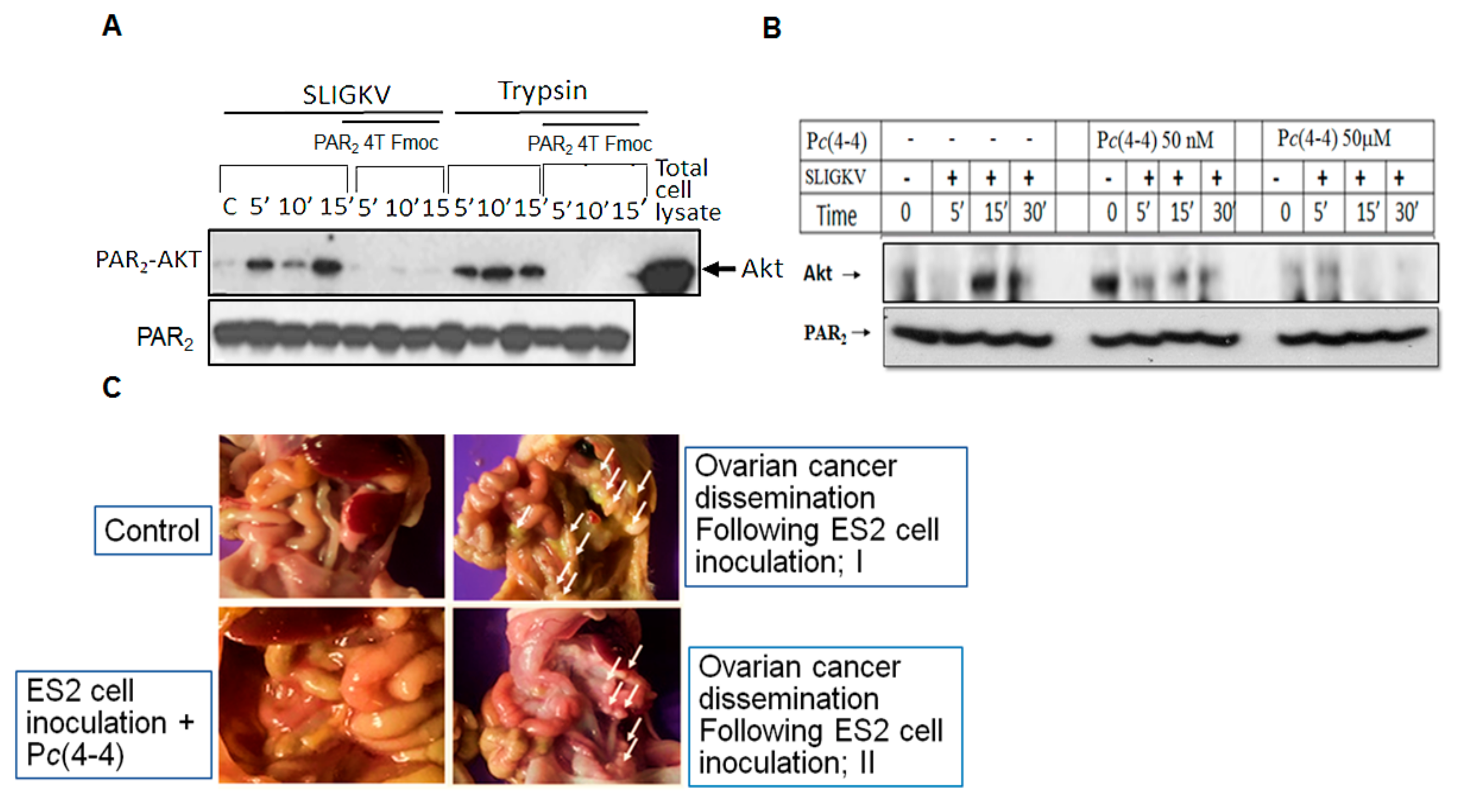
3.5. Characterization of PAR2-PH Binding Domain Peptides
3.6. Pc(4-4) Inhibits PAR2-Akt Association and Ovarian Cancer Peritoneal Dissemination
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samuel, D.; Diaz-Barbe, A.; Pinto, A.; Schlumbrecht, M.; George, S. Hereditary Ovarian Carcinoma: Cancer Pathogenesis Looking beyond BRCA1 and BRCA2. Cells 2022, 11, 539. [Google Scholar] [CrossRef] [PubMed]
- Huber, D.; Seitz, S.; Kast, K.; Emons, G.; Ortmann, O. Use of oral contraceptives in BRCA mutation carriers and risk for ovarian and breast cancer: A systematic review. Arch. Gynecol. Obstet. 2020, 301, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Lengyel, E. Ovarian cancer development and metastasis. Am. J. Pathol. 2010, 177, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Lei, N.; Zhou, J.; Chen, M.; Guo, R.; Qin, B.; Li, Y.; Chang, L. Extracellular vesicles in ovarian cancer chemoresistance, metastasis, and immune evasion. Cell Death Dis. 2022, 13, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Bast, R.C.; Hennessy, B., Jr.; Mills, G.B. The biology of ovarian cancer: New opportunities for translation. Nat. Rev. Cancer 2009, 9, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Feuer, D.J.; Broadley, K.E.; Shepherd, J.H.; Barton, D.P. Surgery for the resolution of symptoms in malignant bowel obstruction in advanced gynaecological and gastrointestinal cancer. Cochrane Database Syst. Rev. 2016, 1, CD002764. [Google Scholar]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28–45. [Google Scholar] [CrossRef]
- Gutkind, J.S.; Kostenis, E. Arrestins as rheostats of GPCR signalling. Nat. Rev. Mol. Cell Biol. 2018, 19, 615–616. [Google Scholar] [CrossRef]
- Chaudhary, P.K.; Kim, S. An Insight into GPCR and G-Proteins as Cancer Drivers. Cells 2021, 10, 3288. [Google Scholar] [CrossRef]
- Kancharla, A.; Maoz, M.; Jaber, M.; Agranovich, D.; Peretz, T.; Grisaru-Granovsky, S.; Uziely, B.; Bar-Shavit, R. PH motifs in PAR1&2 endow breast cancer growth. Nat. Commun. 2015, 6, 8853–8863. [Google Scholar]
- Nag, J.K.; Malka, H.; Sedley, S.; Appasamy, P.; Rudina, T.; Levi, T.; Hoffman, A.; Gilon, C.; Uziely, B.; Bar-Shavit, R. PH-Binding Motif in PAR4 Oncogene: From Molecular Mechanism to Drug Design. Molecular Cancer Therapeutics. Mol. Cancer Ther. 2022, 21, 1415–1429. [Google Scholar] [CrossRef] [PubMed]
- Rebecchi, M.J.; Scarlata, S. Pleckstrin homology domains: A common fold with diverse functions. Ann. Rev. Biophys. Biomol. Struct. 1998, 27, 503–528. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.X.; Xie, Y.; Zhang, Y.; Charlat, O.; Oster, E.; Avello, M.; Lei, H.; Mickanin, C.; Liu, D.; Ruffner, H.; et al. ZNRF3 promotes Wnt receptor turnover in an R-spondin-sensitive manner. Nature 2012, 485, 195–200. [Google Scholar] [CrossRef]
- Koo, B.K.; Spit, M.; Jordens, I.; Low, T.Y.; Stange, D.E.; van de Wetering, M.; Van Es, J.H.; Mohammed, S.; Heck, A.J.R.; Maurice, M.M.; et al. Tumour suppressor RNF43 is a stem-cell E3 ligase that induces endocytosis of Wnt receptors. Nature 2012, 488, 665–669. [Google Scholar] [CrossRef]
- Ryland, G.L.; Hunter, S.M.; Doyle, M.A.; Rowley, S.M.; Christie, M.; Allan, P.E.; Bowtell, D.D.; Australian Ovarian Cancer Study Group; Gorringe, K.L.; Campbell, I.G. RNF43 is a tumor suppressor gene mutated in mucinous tumours of the ovary. J. Pathol. 2013, 229, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hao, H.X.; Growney, J.D.; Woolfenden, S.; Bottiglio, C.; Ng, N.; Lu, B.; Hsieh, M.H.; Bagdasarian, L.; Meyer, R.; et al. Inactivating mutations of RNF43 confer Wnt dependency in pancreatic ductal adenocarcinoma. Proc. Natl. Acad. Sci. USA 2013, 110, 12649–12654. [Google Scholar] [CrossRef] [PubMed]
- Nag, J.K.; Appasamy, P.; Sedley, S.; Malka, H.; Rudina, T.; Bar-Shavit, R. RNF43 induces the turnover of protease-activated receptor 2 in colon cancer. FASEB J. 2023, 37, e22675. [Google Scholar] [CrossRef]
- Shih, I.M.; Wang, Y.; Wang, T.L. The Origin of Ovarian Cancer Species and Precancerous Landscape. Am. J. Pathol. 2022, 191, 26–39. [Google Scholar] [CrossRef]
- Kindelberger, D.W.; Lee, Y.; Miron, A.; Hirsch, M.S.; Feltmate, C.; Medeiros, F.; Callahan, M.J.; Garner, E.O.; Gordon, R.W.; Birch, C.; et al. Intraepithelial carcinoma of the fimbria and pelvic serous carcinoma: Evidence for a causal relationship. Am. J. Surg. Pathol. 2007, 31, 161–169. [Google Scholar] [CrossRef]
- Przybycin, C.G.; Kurman, R.J.; Ronnett, B.M.; Shih, I.M.; Vang, R. Are all pelvic (nonuterine) serous carcinomas of tubal origin? Am. J. Surg. Pathol. 2010, 34, 1407–1416. [Google Scholar] [CrossRef]
- Folkins, A.K.; Jarboe, E.A.; Saleemuddin, A.; Lee, Y.; Callahan, M.J.; Drapkin, R.; Garber, J.E.; Muto, M.G.; Tworoger, S.; Crum, C.P. A candidate precursor to pelvic serous cancer (p53 signature) and its prevalence in ovaries and fallopian tubes from women with BRCA mutations. Gynecol. Oncol. 2008, 109, 168–173. [Google Scholar] [CrossRef]
- Labidi-Galy, S.I.; Papp, E.; Hallberg, D.; Niknafs, N.; Adleff, V.; Noe, M.; Bhattacharya, R.; Novak, M.; Jones, S.; Phallen, J.; et al. High grade serous ovarian carcinomas originate in the fallopian tube. Nat. Commun. 2017, 8, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.C.; Wang, P.; Lin, S.F.; Zhang, M.; Song, Q.; Chu, T.; Wang, B.G.; Kurman, R.J.; Vang, R.; Kinzler, K.; et al. Genomic landscape and evolutionary trajectories of ovarian cancer precursor lesions. J. Pathol. 2019, 248, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.Y.; Fang, C.; Huang, H.S.; Wang, J.; Chu, T.Y. Natural history of ovarian high-grade serous carcinoma from time effects of ovulation inhibition and progesterone clearance of p53-defective lesions. Mod. Pathol. 2020, 33, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, K.; Shaw, P.; May, B.J.; Bahadirli-Talbott, A.; Kaushiva, A.; Risch, H.; Narod, S.; Wang, T.-L.; Parkash, V.; Vang, R.; et al. Fallopian Tube Lesions in Women at High Risk for Ovarian Cancer: A Multicenter Study. Cancer Prev. Res. 2018, 11, 697–706. [Google Scholar] [CrossRef]
- Perets, R.; Wyant, G.A.; Muto, K.W.; Bijron, J.G.; Poole, B.B.; Chin, K.T.; Chen, J.Y.; Ohman, A.W.; Stepule, C.D.; Kwak, S.; et al. Transformation of the fallopian tube secretory epithelium leads to high-grade serous ovarian cancer in Brca; Tp53; Pten models. Cancer Cell 2013, 24, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Turner, R.; Khazan, N.; Kodza, A.; Jones, A.; Singh, R.K.; Moore, R.G. Role of trypsin and protease-activated receptor-2 in ovarian cancer. PLoS ONE 2020, 15, e0232253. [Google Scholar] [CrossRef]
- Jiang, Y.; Lim, J.; Wu, K.C.; Xu, W.; Suen, J.Y.; Fairlie, D.P. PAR2 induces ovarian cancer cell motility by merging three signalling pathways to transactivate EGFR. Br. J. Pharmacol. 2021, 178, 913–932. [Google Scholar] [CrossRef]
- Pawar, N.R.; Buzza, M.S.; Duru, N.; Strong, A.A.; Antalis, T.M. Matriptase drives dissemination of ovarian cancer spheroids by a PAR-2/PI3K/Akt/MMP9 signaling axis. J. Cell Biol. 2023, 222, e202209114. [Google Scholar] [CrossRef]
- Devor, E.J.; Lapierre, J.R.; David P Bender, D.P. ES-2 Ovarian Cancer Cells Present a Genomic Profile Inconsistent with their Reported History. Obstet. Gynecol. Res. 2021, 4, 233–238. [Google Scholar] [CrossRef]
- Even-Ram, S.; Uziely, B.; Cohen, P.; Grisaru-Granovsky, S.; Maoz, M.; Ginzburg, Y.; Reich, R.; Vlodavsky, I.; Bar-Shavit, R. Thrombin receptor overexpression in malignant and physiological invasion processes. Nat. Med. 1998, 4, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell Mol. Med. 2005, 9, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Hanada, M.; Feng, J.; Hemmings, B.A. Structure, regulation and function of PKB/AKT-a major therapeutic target. Biochim. Biophys. Acta 2004, 1697, 3–16. [Google Scholar] [CrossRef]
- Lu, J.; Steeg, P.S.; Price, J.E.; Krishnamurthy, S.; Mani, S.A.; Reuben, J.; Cristofanilli, M.; Dontu, G.; Bidaut, L.; Valero, V.; et al. Breast cancer metastasis: Challenges and opportunities. Cancer Res. 2009, 69, 4951–4953. [Google Scholar] [CrossRef] [PubMed]
- Chu, N.; Salguero, A.L.; Liu, A.Z.; Chen, Z.; Dempsey, D.R.; Ficarro, S.B.; Alexander, W.M.; Marto, J.A.; Li, Y.; Amzel, L.M.; et al. Akt kinase Activation Mechanisms Revealed Using Protein Semisynthesis. Cell 2018, 174, 897–907. [Google Scholar] [CrossRef]
- Qiu, Y.; Kung, H.J. Signaling network of the Btk family kinases. Oncogene 2000, 19, 5651–5661. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Robinson, D.; Pretlow, T.G.; Kung, H.J. Etk/Bmx, a tyrosine kinase with a pleckstrin-homology domain, is an effector of phosphatidylinositol 3′- kinase and is involved in interleukin 6-induced neuroendocrine differentiation of prostate cancer cells. Proc. Natl. Acad. Sci. USA 1998, 95, 3644–3649. [Google Scholar] [CrossRef]
- Cohen, I.; Maoz, M.; Turm, H.; Grisaru-Granovsky, S.; Maly, B.; Uziely, B.; Weiss, E.; Abramovitch, R.; Gross, E.; Barzilay, O.; et al. Etk/Bmx regulates proteinase-activated-receptor1 (PAR1) in breast cancer invasion: Signaling partners, hierarchy and physiological significance. PLoS ONE 2010, 5, e11135. [Google Scholar] [CrossRef]
- Banerjee, S.; Tian, T.; Wei, Z.; Shih, N.; Feldman, M.D.; Alwine, J.C.; Coukos, G.; Robertson, E.S. The ovarian cancer oncobiome. Oncotarget 2017, 8, 36225–36245. [Google Scholar] [CrossRef]
- Chanakira, A.; Westmark, P.R.; Ong, I.M.; Sheehan, J.P. Tissue Factor-Factor VIIa Complex Triggers Protease Activated Receptor 2-Dependent Growth Factor Release and Migration in Ovarian Cancer. Gynecol. Oncol. 2017, 145, 167–175. [Google Scholar] [CrossRef]
- Tanimoto, H.; Shigemasa, K.; Tian, X.; Gu, L.; Beard, J.B.; Sawasaki, T.T.; J O’Brien, T.J. Transmembrane serine protease TADG-15 (ST14/Matriptase/MT-SP1): Expression and prognostic value in ovarian cancer. Br. J. Cancer 2005, 92, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Loessner, D.; Irving-Rodgers, H.; Obermair, A.; Nicklin, J.L.; Clements, J.A. Metastasis of ovarian cancer is mediated by kallikrein related peptidases. Clin. Exp. Metastasis 2014, 31, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, P.; Wang, B.G.; Murdock, T.; Cope, L.; Hsu, F.-C.; Wang, T.-L.; Shih, I.-M. Spatial Transcriptomic Analysis of Ovarian Cancer Precursors Reveals Reactivation of IGFBP2 During Pathogenesis. Cancer Res. 2022, 82, 4528–4541. [Google Scholar] [CrossRef] [PubMed]
- Salah, Z.; Uziely, B.; Jaber, M.; Maoz, M.; Cohen, I.; Hamburger, T.; Maly, B.; Peretz, T.; Bar-Shavit, R. Regulation of human protease-activated receptor 1 (hPar1) gene expression in breast cancer by estrogen. FASEB J. 2012, 26, 2031–2042. [Google Scholar] [CrossRef]
- Kuhn, E.; Kurman, R.J.; Soslow, R.; Han, G.; Sehdev, A.S.; Morin, P.J.; Tian-Li, W.; Shih, I.-M. The diagnostic and biological implications of laminin expression in serous tubal intraepithelial carcinoma. Am. J. Surg. Pathol. 2012, 36, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Sehdev, A.S.; Kurman, R.J.; Kuhn, E.; Shih, I.-M. Serous tubal intraepithelial carcinoma upregulates markers associated with high-grade serous carcinomas including Rsf-1 (HBXAP), cyclin E and fatty acid synthase. Mod. Pathol. 2010, 23, 844–855. [Google Scholar] [CrossRef]
- Karsta, A.M.; Levanona, K.; Duraisamya, S.; Liua, J.F.; Hirschb, M.S.; Hechtc, J.L.; Drapkin, R. Stathmin 1, a marker of PI3K pathway activation and regulator of microtubule dynamics, is expressed in early pelvic serous carcinomas. Gynecol. Oncol. 2011, 123, 5–12. [Google Scholar] [CrossRef]
- Aman, M.; Ohishi, Y.; Imamura, H.; Shinozaki, T.; Yasutake, N.; Kato, K.; Oda, Y. Expression of protease-activated receptor-2 (PAR-2) is related to advanced clinical stage and adverse prognosis in ovarian clear cell carcinoma. Hum. Pathol. 2017, 64, 156–163. [Google Scholar] [CrossRef]
- Arora, P.; Cuevas, B.D.; Russo, A.; Johnson, G.L.; Trejo, J. Persistent transactivation of EGFR and ErbB2/HER2 by protease-activated receptor-1 promotes breast carcinoma cell invasion. Oncogene 2008, 27, 4434–4445. [Google Scholar] [CrossRef]
- Palanisamy, S.; Xue, C.; Ishiyama, S.; Naga Prasad, S.V.; Gabrielson, K. GPCR-ErbB transactivation pathways and clinical implications. Cell Signal. 2021, 86, 1100–1119. [Google Scholar] [CrossRef]
- Andreev, J.; Galisteo, M.L.; Kranenburg, O.; Logan, S.K.; Chiu, E.S.; Okigaki, M.; Cary, L.A.; Moolenaar, W.H.; Schlessinger, J. Src and Pyk2 mediate G-protein-coupled receptor activation of epidermal growth factor receptor (EGFR) but are not required for coupling to the mitogen-activated protein (MAP) kinase signaling cascade. J. Biol. Chem. 2001, 276, 20130–20135. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nag, J.K.; Grisaru-Granovsky, S.; Armon, S.; Rudina, T.; Appasamy, P.; Bar-Shavit, R. Involvement of Protease-Activated Receptor2 Pleckstrin Homology Binding Domain in Ovarian Cancer: Expression in Fallopian Tubes and Drug Design. Biomedicines 2024, 12, 246. https://doi.org/10.3390/biomedicines12010246
Nag JK, Grisaru-Granovsky S, Armon S, Rudina T, Appasamy P, Bar-Shavit R. Involvement of Protease-Activated Receptor2 Pleckstrin Homology Binding Domain in Ovarian Cancer: Expression in Fallopian Tubes and Drug Design. Biomedicines. 2024; 12(1):246. https://doi.org/10.3390/biomedicines12010246
Chicago/Turabian StyleNag, Jeetendra Kumar, Sorina Grisaru-Granovsky, Shunit Armon, Tatyana Rudina, Priyanga Appasamy, and Rachel Bar-Shavit. 2024. "Involvement of Protease-Activated Receptor2 Pleckstrin Homology Binding Domain in Ovarian Cancer: Expression in Fallopian Tubes and Drug Design" Biomedicines 12, no. 1: 246. https://doi.org/10.3390/biomedicines12010246