

Uncovering the Neural Correlates of Anhedonia Subtypes in Major Depressive Disorder: Implications for Intervention Strategies

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
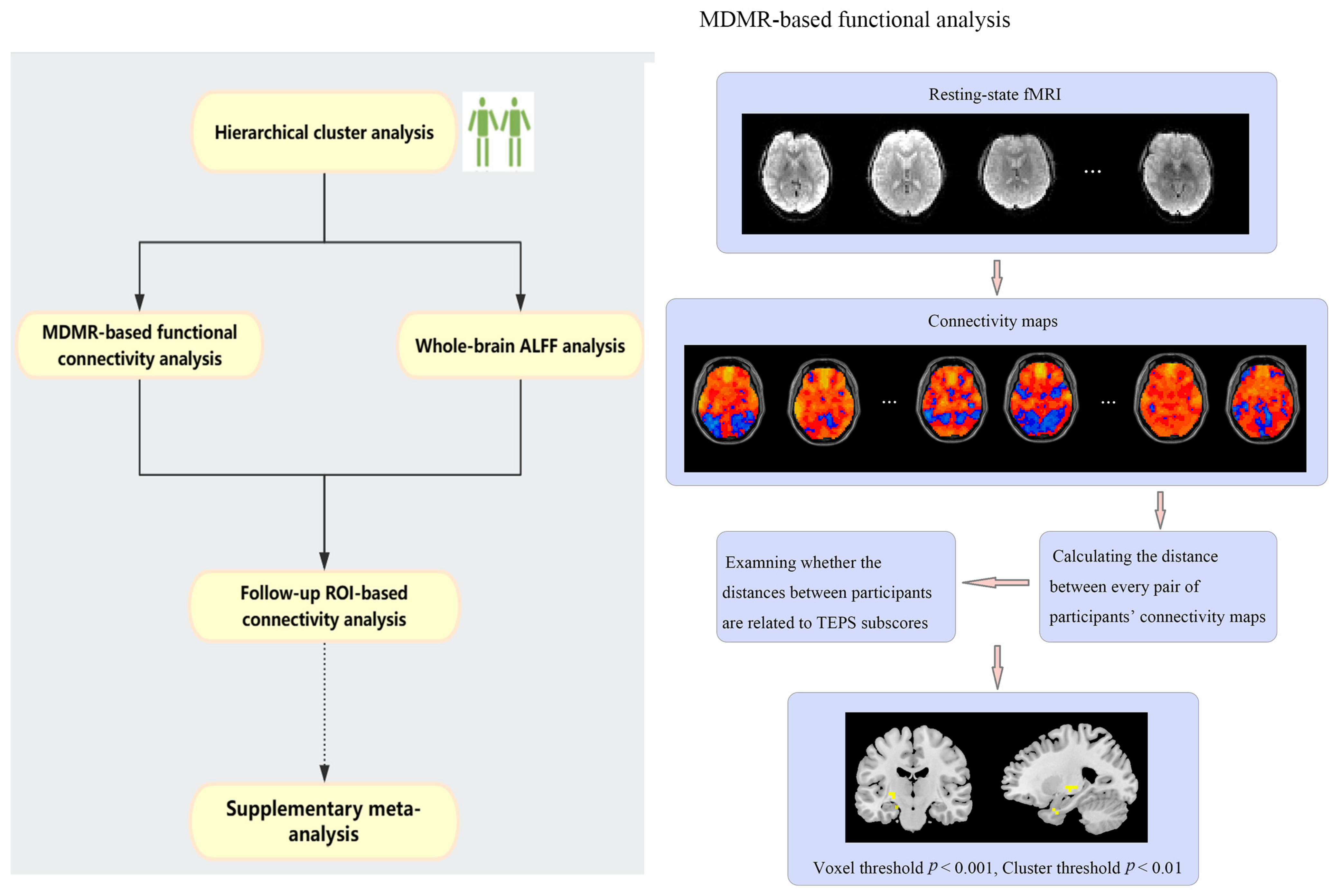
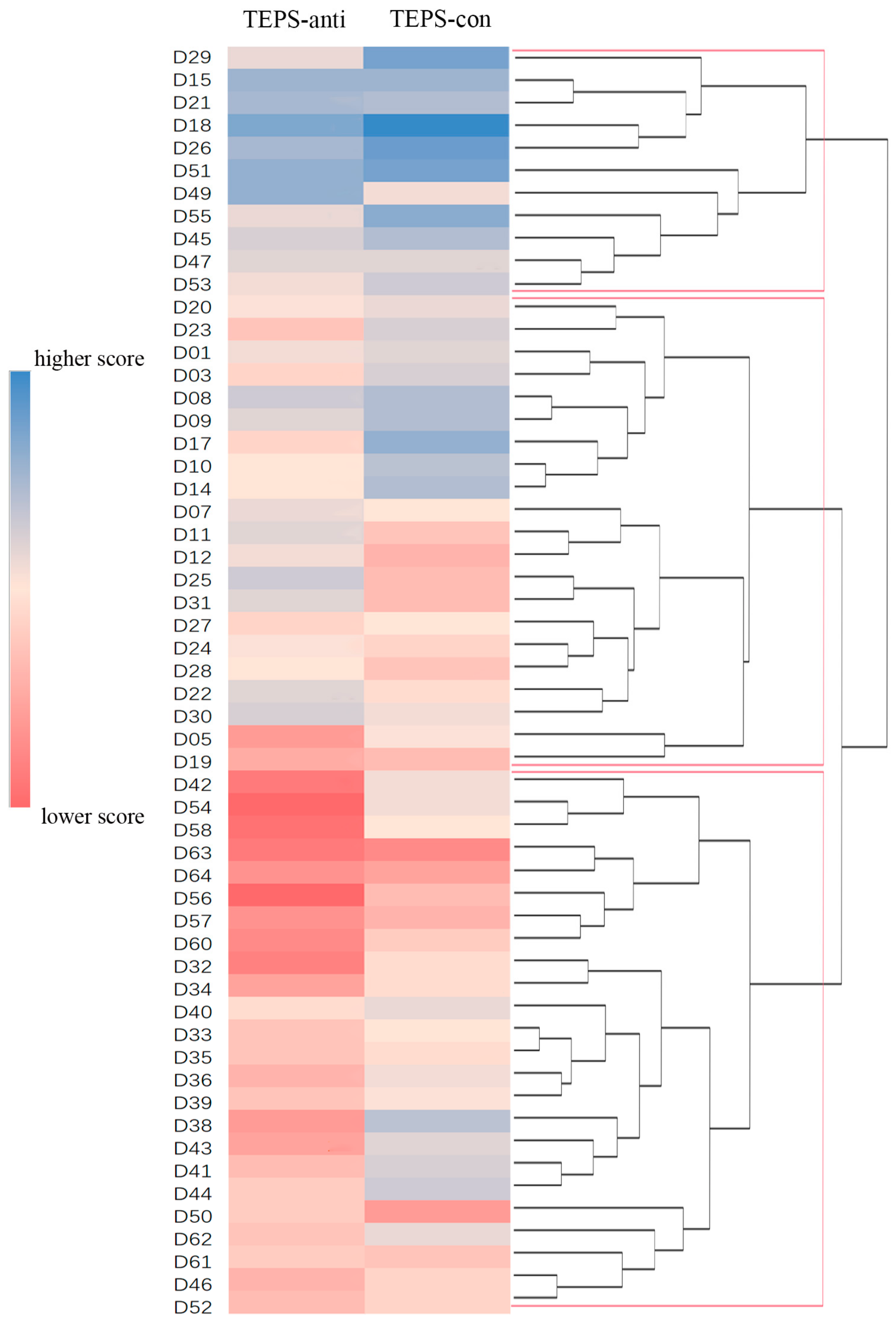
2.2. Hierarchical Cluster Analysis
2.3. MDMR-Based Functional Connectivity Analysis
2.4. Whole-Brain ALFF Analysis
2.5. Follow-Up ROI-Based Connectivity Analysis
2.6. Supplementary Meta-Analysis
3. Results
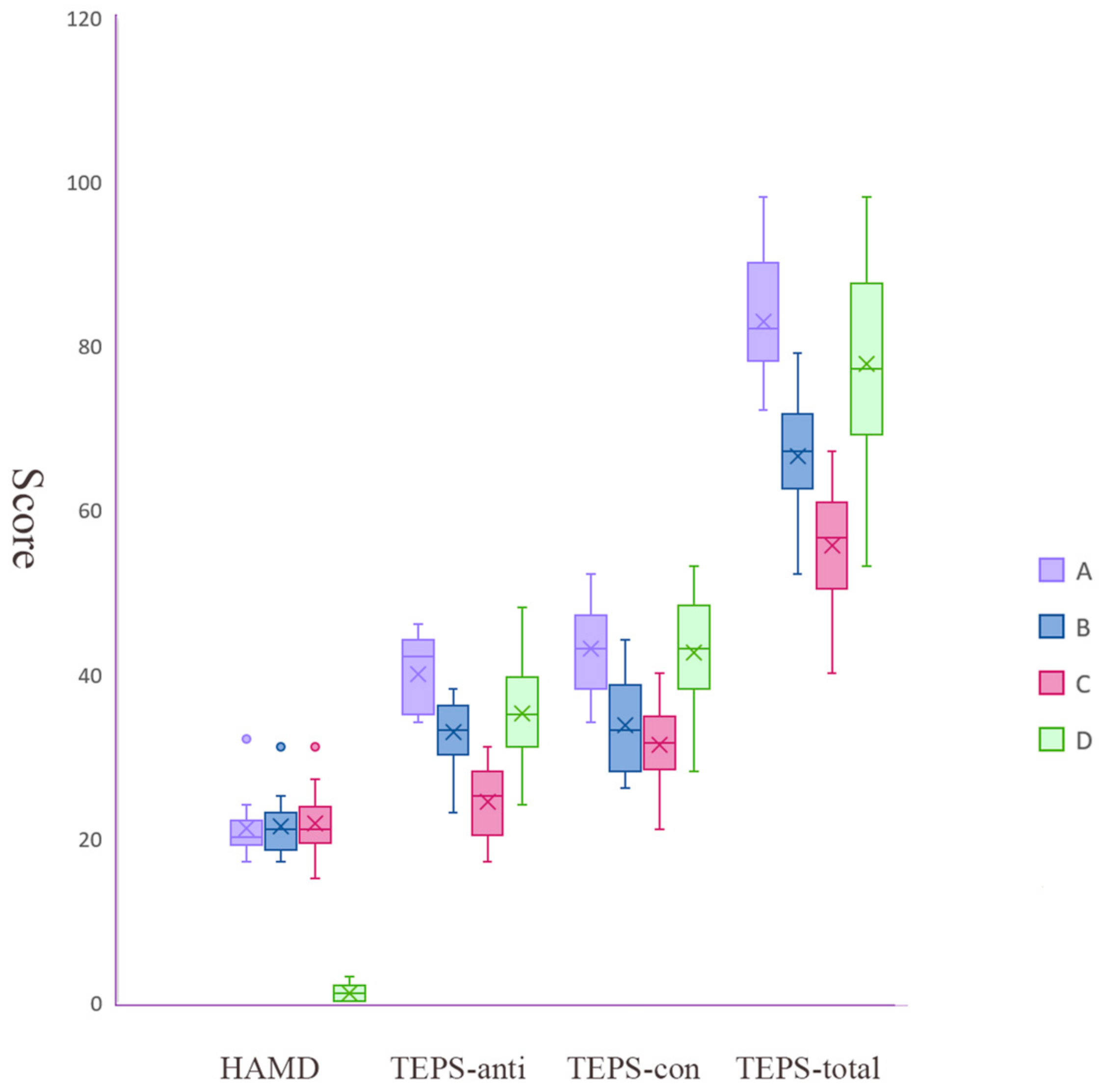
3.1. Three MDD Subgroups Identified by Hierarchical Clustering
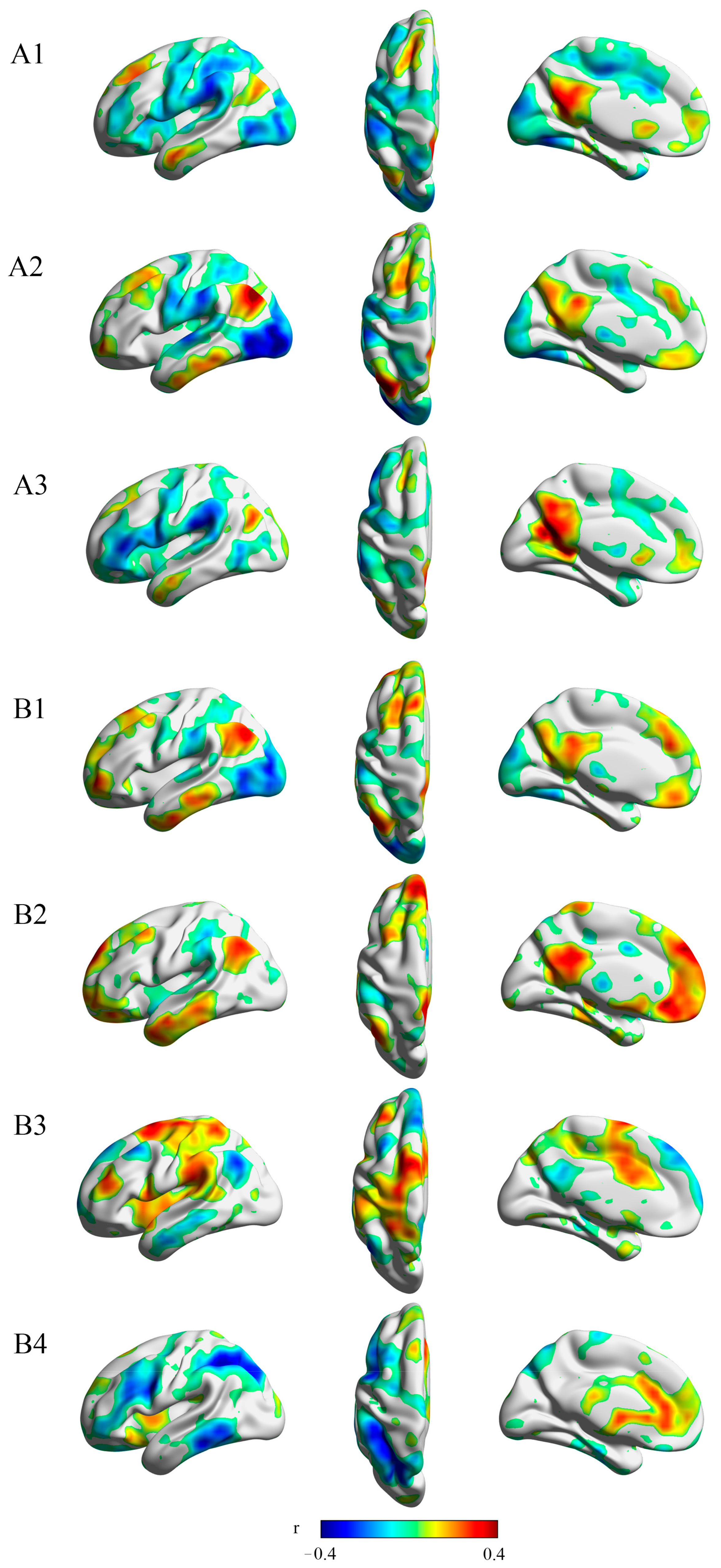
3.2. Resting-State Connectivity to Several Regions within the Reward Network Is Correlated to Anticipatory and Consummatory Pleasure
3.3. ALFF Values Differences across 4 Groups (3 MDD Subgroups and HC)
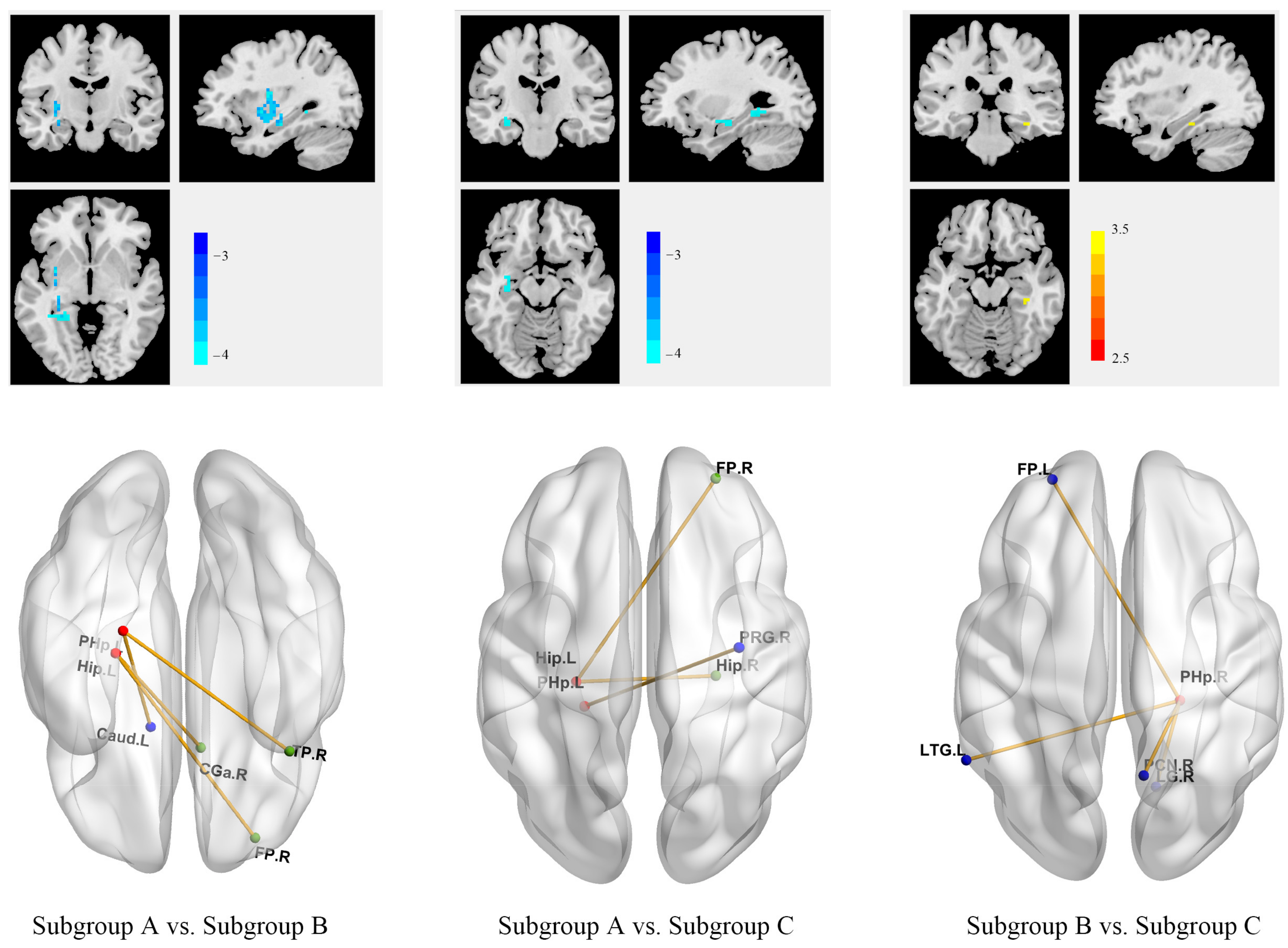
3.4. Different Anhedonia Profiles across 3 MDD Subgroups Were Associated with Different Patterns of Within- and Between-Network Connectivity
3.5. MDD Patients Revealed Reduced Activity in Dorsal Striatum during Reward Feedback
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.; Vos, T.; Lozano, R.; Naghavi, M.; Flaxman, A.D.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; et al. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2197–2223. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, B.N. The RDoC framework: Facilitating transition from ICD/DSM to dimensional approaches that integrate neuroscience and psychopathology. World Psychiatry 2014, 13, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, D.; Zhou, X.; Chen, L.; Liu, L.; Lin, R.; Li, X.; Liu, Y.; Qiu, H.; Cao, H.; et al. Neurological and metabolic related pathophysiologies and treatment of comorbid diabetes with depression. CNS Neurosci. Ther. 2023, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Chen, C. Editorial: Towards a mechanistic understanding of depression, anxiety, and their comorbidity: Perspectives from cognitive neuroscience. Front. Behav. Neurosci. 2023, 17, 1268156. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.J.; Pizzagalli, D.A.; Sproule, B.A.; Kennedy, S.H. Assessing anhedonia in depression: Potentials and pitfalls. Neurosci. Biobehav. Rev. 2016, 65, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Whitton, A.E.; Treadway, M.T.; Pizzagalli, D.A. Reward processing dysfunction in major depression, bipolar disorder and schizophrenia. Curr. Opin. Psychiatry 2015, 28, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Snaith, R.P.; Hamilton, M.; Morley, S.; Humayan, A.; Hargreaves, D.; Trigwell, P. A scale for the assessment of hedonic tone the Snaith-Hamilton Pleasure Scale. Br. J. Psychiatry 1995, 167, 99–103. [Google Scholar] [CrossRef]
- Sherdell, L.; Waugh, C.E.; Gotlib, I.H. Anticipatory pleasure predicts motivation for reward in major depression. J. Abnorm. Psychol. 2012, 121, 51–60. [Google Scholar] [CrossRef]
- Li, Y.; Mou, X.; Jiang, W.; Yang, Z.; Shen, X.; Jin, Z.; Dai, Z.; Liu, Y.; Mao, S.; Zhang, J.; et al. A comparative study of anhedonia components between major depression and schizophrenia in Chinese populations. Ann. Gen. Psychiatry 2015, 14, 24. [Google Scholar] [CrossRef]
- Nakonezny, P.A.; Carmody, T.J.; Morris, D.W.; Kurian, B.T.; Trivedi, M.H. Psychometric evaluation of the Snaith-Hamilton pleasure scale in adult outpatients with major depressive disorder. Int. Clin. Psychopharmacol. 2010, 25, 328–333. [Google Scholar] [CrossRef]
- Parker, G.; Bassett, D.; Outhred, T.; Morris, G.; Hamilton, A.; Das, P.; Baune, B.T.; Berk, M.; Boyce, P.; Lyndon, B.; et al. Defining melancholia: A core mood disorder. Bipolar Disord. 2017, 19, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Association, A.P. Diagnostic and Statistical Manual of Mental Disorders: DSM-5; American Psychiatric Press: Washington, DC, USA, 2013. [Google Scholar]
- Parker, G.H.-P.D.; Boyce, P. Issues in classification II: Classifying melancholia. In Melancholia: A Disorder of Movement and Mood: A Phenomenological and Neurobiological Review; Parker, G.H.-P.D., Ed.; Cambridge University Press: Cambridge, UK, 1996; pp. 20–37. [Google Scholar]
- Day, C.V.; Gatt, J.M.; Etkin, A.; DeBattista, C.; Schatzberg, A.F.; Williams, L.M. Cognitive and emotional biomarkers of melancholic depression: An iSPOT-D report. J. Affect. Disord. 2015, 176, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Foti, D.; Carlson, J.M.; Sauder, C.L.; Proudfit, G.H. Reward dysfunction in major depression: Multimodal neuroimaging evidence for refining the melancholic phenotype. Neuroimage 2014, 101, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, A.; Shankman, S.A. Blunted reward processing in remitted melancholic depression. Clin. Psychol. Sci. 2017, 5, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.; Blanch, B.; Paterson, A.; Hadzi-Pavlovic, D.; Sheppard, E.; Manicavasagar, V.; Synnott, H.; Graham, R.K.; Friend, P.; Gilfillan, D.; et al. The superiority of antidepressant medication to cognitive behavior therapy in melancholic depressed patients: A 12-week single-blind randomized study. Acta Psychiatr. Scand. 2013, 128, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Su, Y.; Rizvi, S.J.; Jagoda, J.; Li, J.; Wu, Y.; Dai, Y.; Zhang, Y.; Kennedy, S.H.; Si, T. Define and characterize the anhedonia in major depressive disorder: An explorative study. J. Affect. Disord. 2022, 313, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Heshmati, M.; Russo, S.J. Anhedonia and the brain reward circuitry in depression. Curr. Behav. Neurosci. Rep. 2015, 2, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Leri, F.; Rizvi, S.J. Anhedonia as a central factor in depression: Neural mechanisms revealed from preclinical to clinical evidence. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 110, 110289. [Google Scholar] [CrossRef]
- Oldham, S.; Murawski, C.; Fornito, A.; Youssef, G.; Yucel, M.; Lorenzetti, V. The anticipation and outcome phases of reward and loss processing: A neuroimaging meta-analysis of the monetary incentive delay task. Hum. Brain Mapp. 2018, 39, 3398–3418. [Google Scholar] [CrossRef]
- Keren, H.; O’Callaghan, G.; Vidal-Ribas, P.; Buzzell, G.A.; Brotman, M.A.; Leibenluft, E.; Pan, P.M.; Meffert, L.; Kaiser, A.; Wolke, S.; et al. Reward Processing in Depression: A Conceptual and Meta-Analytic Review Across fMRI and EEG Studies. Am. J. Psychiatry 2018, 175, 1111–1120. [Google Scholar] [CrossRef]
- Wang, L.; Li, F.; Mitchell, P.B.; Wang, C.Y.; Si, T.M. Striatal Resting-State Connectivity Abnormalities Associated with Different Clinical Stages of Major Depressive Disorder. J. Clin. Psychiatry 2020, 81, 4265. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; An, J.; Gao, H.M.; Zhang, P.; Chen, C.; Li, K.; Mitchell, P.B.; Si, T.M. Duloxetine effects on striatal resting-state functional connectivity in patients with major depressive disorder. Hum. Brain Mapp. 2019, 40, 3338–3346. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, Y.; Chen, X.; Zhang, Z.; Xiao, L.; Zhou, Y. Anhedonia correlates with functional connectivity of the nucleus accumbens subregions in patients with major depressive disorder. Neuroimage Clin. 2021, 30, 102599. [Google Scholar] [CrossRef] [PubMed]
- Pizzagalli, D.A. Toward a Better Understanding of the Mechanisms and Pathophysiology of Anhedonia: Are We Ready for Translation? Am. J. Psychiatry 2022, 179, 458–469. [Google Scholar] [CrossRef]
- Nagy, G.A.; Cernasov, P.; Pisoni, A.; Walsh, E.; Dichter, G.S.; Smoski, M.J. Reward Network Modulation as a Mechanism of Change in Behavioral Activation. Behav. Modif. 2020, 44, 186–213. [Google Scholar] [CrossRef] [PubMed]
- Pizzagalli, D.A.; Smoski, M.; Ang, Y.S.; Whitton, A.E.; Sanacora, G.; Mathew, S.J.; Nurnberger, J., Jr.; Lisanby, S.H.; Iosifescu, D.V.; Murrough, J.W.; et al. Selective kappa-opioid antagonism ameliorates anhedonic behavior: Evidence from the Fast-fail Trial in Mood and Anxiety Spectrum Disorders (FAST-MAS). Neuropsychopharmacology 2020, 45, 1656–1663. [Google Scholar] [CrossRef] [PubMed]
- Krystal, A.D.; Pizzagalli, D.A.; Smoski, M.; Mathew, S.J.; Nurnberger, J., Jr.; Lisanby, S.H.; Iosifescu, D.; Murrough, J.W.; Yang, H.; Weiner, R.D.; et al. A randomized proof-of-mechanism trial applying the ‘fast-fail’ approach to evaluating kappa-opioid antagonism as a treatment for anhedonia. Nat. Med. 2020, 26, 760–768. [Google Scholar] [CrossRef]
- Maisonneuve, I.M.; Archer, S.; Glick, S.D. U50,488, a kappa opioid receptor agonist, attenuates cocaine-induced increases in extracellular dopamine in the nucleus accumbens of rats. Neurosci. Lett. 1994, 181, 57–60. [Google Scholar] [CrossRef]
- Bell, C.C. DSM-IV: Diagnostic and Statistical Manual of Mental Disorders. JAMA 1994, 272, 828–829. [Google Scholar] [CrossRef]
- Hamilton, M. A rating scale for depression. J. Neurol. Neurosurg. Psychiatry 1960, 23, 56–62. [Google Scholar] [CrossRef]
- Gard, D.E.; Gard, M.G.; Kring, A.M.; John, O.P. Anticipatory and consummatory components of the experience of pleasure: A scale development study. J. Res. Personal. 2006, 40, 1086–1102. [Google Scholar] [CrossRef]
- Szekely, G.J.; Rizzo, M.L. Hierarchical Clustering via Joint Between-Within Distances: Extending Ward’s Minimum Variance Method. J. Classif. 2005, 22, 151–183. [Google Scholar] [CrossRef]
- Frid, M.G.; McKeon, B.A.; Thurman, J.M.; Maron, B.A.; Li, M.; Zhang, H.; Kumar, S.; Sullivan, T.; Laskowsky, J.; Fini, M.A.; et al. Immunoglobulin-driven Complement Activation Regulates Proinflammatory Remodeling in Pulmonary Hypertension. Am. J. Respir. Crit. Care Med. 2020, 201, 224–239. [Google Scholar] [CrossRef]
- Misaki, M.; Phillips, R.; Zotev, V.; Wong, C.K.; Wurfel, B.E.; Krueger, F.; Feldner, M.; Bodurka, J. Connectome-wide investigation of altered resting-state functional connectivity in war veterans with and without posttraumatic stress disorder. Neuroimage Clin. 2018, 17, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Wolf, D.H.; Ciric, R.; Kable, J.W.; Moore, T.M.; Vandekar, S.N.; Katchmar, N.; Daldal, A.; Ruparel, K.; Davatzikos, C.; et al. Common Dimensional Reward Deficits Across Mood and Psychotic Disorders: A Connectome-Wide Association Study. Am. J. Psychiatry 2017, 174, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Shehzad, Z.; Kelly, C.; Reiss, P.T.; Cameron Craddock, R.; Emerson, J.W.; McMahon, K.; Copland, D.A.; Castellanos, F.X.; Milham, M.P. A multivariate distance-based analytic framework for connectome-wide association studies. Neuroimage 2014, 93 Pt 1, 74–94. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, K.R.; Sabuncu, M.R.; Buckner, R.L. The influence of head motion on intrinsic functional connectivity MRI. Neuroimage 2012, 59, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Satterthwaite, T.D.; Wolf, D.H.; Loughead, J.; Ruparel, K.; Elliott, M.A.; Hakonarson, H.; Gur, R.C.; Gur, R.E. Impact of in-scanner head motion on multiple measures of functional connectivity: Relevance for studies of neurodevelopment in youth. Neuroimage 2012, 60, 623–632. [Google Scholar] [CrossRef]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar] [CrossRef]
- Zuo, X.N.; Di Martino, A.; Kelly, C.; Shehzad, Z.E.; Gee, D.G.; Klein, D.F.; Castellanos, F.X.; Biswal, B.B.; Milham, M.P. The oscillating brain: Complex and reliable. Neuroimage 2010, 49, 1432–1445. [Google Scholar] [CrossRef]
- Albajes-Eizagirre, A.; Solanes, A.; Vieta, E.; Radua, J. Voxel-based meta-analysis via permutation of subject images (PSI): Theory and implementation for SDM. Neuroimage 2019, 186, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Radua, J.; Grau, M.; van den Heuvel, O.A.; Thiebaut de Schotten, M.; Stein, D.J.; Canales-Rodriguez, E.J.; Catani, M.; Mataix-Cols, D. Multimodal voxel-based meta-analysis of white matter abnormalities in obsessive-compulsive disorder. Neuropsychopharmacology 2014, 39, 1547–1557. [Google Scholar] [CrossRef] [PubMed]
- Muller, V.I.; Cieslik, E.C.; Laird, A.R.; Fox, P.T.; Radua, J.; Mataix-Cols, D.; Tench, C.R.; Yarkoni, T.; Nichols, T.E.; Turkeltaub, P.E.; et al. Ten simple rules for neuroimaging meta-analysis. Neurosci. Biobehav. Rev. 2018, 84, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Rice, F.; Riglin, L.; Lomax, T.; Souter, E.; Potter, R.; Smith, D.J.; Thapar, A.K.; Thapar, A. Adolescent and adult differences in major depression symptom profiles. J. Affect. Disord. 2019, 243, 175–181. [Google Scholar] [CrossRef]
- Manfro, P.H.; Belem da Silva, C.T.; Anselmi, L.; Barros, F.; Eaton, W.W.; Goncalves, H.; Murray, J.; Oliveira, I.O.; Tovo-Rodrigues, L.; Wehrmeister, F.C.; et al. Depression in a youth population-based sample from Brazil: Prevalence and symptom structure. J. Affect. Disord. 2021, 292, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Mata, J.; Furman, D.J.; Whitmer, A.J.; Gotlib, I.H.; Thompson, R.J. Anticipatory and consummatory pleasure and displeasure in major depressive disorder: An experience sampling study. J. Abnorm. Psychol. 2017, 126, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, L.; Li, M.; Ren, Z.; Ma, P. Characterizing the subtype of anhedonia in major depressive disorder: A symptom-specific multimodal MRI study. Psychiatry Res. Neuroimaging 2021, 308, 111239. [Google Scholar] [CrossRef]
- Weller, B.E.; Bowen, N.K.; Faubert, S.J. Latent Class Analysis: A Guide to Best Practice. J. Black Psychol. 2020, 46, 287–311. [Google Scholar] [CrossRef]
- Zhang, B.; Lin, P.; Shi, H.; Öngür, D.; Auerbach, R.P.; Wang, X.; Yao, S.; Wang, X. Mapping anhedonia-specific dysfunction in a transdiagnostic approach: An ALE meta-analysis. Brain Imaging Behav. 2016, 10, 920–939. [Google Scholar] [CrossRef]
- Smith, K.S.; Berridge, K.C. Opioid limbic circuit for reward: Interaction between hedonic hotspots of nucleus accumbens and ventral pallidum. J. Neurosci. 2007, 27, 1594–1605. [Google Scholar] [CrossRef]
- Smith, K.S.; Tindell, A.J.; Aldridge, J.W.; Berridge, K.C. Ventral pallidum roles in reward and motivation. Behav. Brain Res. 2009, 196, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.S.; Berridge, K.C. The ventral pallidum and hedonic reward: Neurochemical maps of sucrose “liking” and food intake. J. Neurosci. 2005, 25, 8637–8649. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Vorel, S.R.; Tranguch, A.J.; Kenny, E.T.; Mazzoni, P.; van Gorp, W.G.; Kleber, H.D. Anhedonia after a selective bilateral lesion of the globus pallidus. Am. J. Psychiatry 2006, 163, 786–788. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraghavan, L.; Vaidya, J.G.; Humphreys, C.T.; Beglinger, L.J.; Paradiso, S. Emotional and motivational changes after bilateral lesions of the globus pallidus. Neuropsychology 2008, 22, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y.; Zhang, R.T.; Li, Y.; Wang, Y.; Wang, Y.M.; Wang, S.K.; Ongur, D.; Cheung, E.F.C.; Chan, R.C.K. Functional connectivity of the default mode network is associated with prospection in schizophrenia patients and individuals with social anhedonia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 92, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cao, C.; Fang, R.; Liu, P.; Luo, S.; Liberzon, I.; Wang, L. Neural correlates of posttraumatic anhedonia symptoms: Decreased functional connectivity between ventral pallidum and default mode network regions. J. Psychiatr. Res. 2021, 140, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Hoflich, A.; Michenthaler, P.; Kasper, S.; Lanzenberger, R. Circuit Mechanisms of Reward, Anhedonia, and Depression. Int. J. Neuropsychopharmacol. 2019, 22, 105–118. [Google Scholar] [CrossRef]
- Jonasson, L.S.; Axelsson, J.; Riklund, K.; Braver, T.S.; Ogren, M.; Backman, L.; Nyberg, L. Dopamine release in nucleus accumbens during rewarded task switching measured by [(1)(1)C]raclopride. Neuroimage 2014, 99, 357–364. [Google Scholar] [CrossRef]
- Spies, M.; Knudsen, G.M.; Lanzenberger, R.; Kasper, S. The serotonin transporter in psychiatric disorders: Insights from PET imaging. Lancet Psychiatry 2015, 2, 743–755. [Google Scholar] [CrossRef]
- Gryglewski, G.; Lanzenberger, R.; Kranz, G.S.; Cumming, P. Meta-analysis of molecular imaging of serotonin transporters in major depression. J. Cereb. Blood Flow. Metab. 2014, 34, 1096–1103. [Google Scholar] [CrossRef]
- Pringle, A.; McCabe, C.; Cowen, P.J.; Harmer, C.J. Antidepressant treatment and emotional processing: Can we dissociate the roles of serotonin and noradrenaline? J. Psychopharmacol. 2013, 27, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Meyniel, F.; Goodwin, G.M.; Deakin, J.W.; Klinge, C.; MacFadyen, C.; Milligan, H.; Mullings, E.; Pessiglione, M.; Gaillard, R. A specific role for serotonin in overcoming effort cost. eLife 2016, 5, e17282. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, J.; Li, Y.; Hu, F.; Lu, Y.; Ma, M.; Feng, Q.; Zhang, J.E.; Wang, D.; Zeng, J.; et al. Dorsal raphe neurons signal reward through 5-HT and glutamate. Neuron 2014, 81, 1360–1374. [Google Scholar] [CrossRef] [PubMed]
- Gleich, T.; Lorenz, R.C.; Pohland, L.; Raufelder, D.; Deserno, L.; Beck, A.; Heinz, A.; Kuhn, S.; Gallinat, J. Frontal glutamate and reward processing in adolescence and adulthood. Brain Struct. Funct. 2015, 220, 3087–3099. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A.; Waters, N.; Carlsson, M.L. Neurotransmitter interactions in schizophrenia—Therapeutic implications. Biol. Psychiatry 1999, 46, 1388–1395. [Google Scholar] [CrossRef] [PubMed]
- Duncan, N.W.; Wiebking, C.; Tiret, B.; Marjanska, M.; Hayes, D.J.; Lyttleton, O.; Doyon, J.; Northoff, G. Glutamate concentration in the medial prefrontal cortex predicts resting-state cortical-subcortical functional connectivity in humans. PLoS ONE 2013, 8, e60312. [Google Scholar] [CrossRef]
- Lally, N.; Nugent, A.C.; Luckenbaugh, D.A.; Ameli, R.; Roiser, J.P.; Zarate, C.A. Anti-anhedonic effect of ketamine and its neural correlates in treatment-resistant bipolar depression. Transl. Psychiatry 2014, 4, e469. [Google Scholar] [CrossRef]
- Downar, J.; Geraci, J.; Salomons, T.V.; Dunlop, K.; Wheeler, S.; McAndrews, M.P.; Bakker, N.; Blumberger, D.M.; Daskalakis, Z.J.; Kennedy, S.H.; et al. Anhedonia and reward-circuit connectivity distinguish nonresponders from responders to dorsomedial prefrontal repetitive transcranial magnetic stimulation in major depression. Biol. Psychiatry 2014, 76, 176–185. [Google Scholar] [CrossRef]
- Duprat, R.; Wu, G.R.; De Raedt, R.; Baeken, C. Accelerated iTBS treatment in depressed patients differentially modulates reward system activity based on anhedonia. World J. Biol. Psychiatry 2018, 19, 497–508. [Google Scholar] [CrossRef]
- Bewernick, B.H.; Hurlemann, R.; Matusch, A.; Kayser, S.; Grubert, C.; Hadrysiewicz, B.; Axmacher, N.; Lemke, M.; Cooper-Mahkorn, D.; Cohen, M.X.; et al. Nucleus accumbens deep brain stimulation decreases ratings of depression and anxiety in treatment-resistant depression. Biol. Psychiatry 2010, 67, 110–116. [Google Scholar] [CrossRef]
- Schlaepfer, T.E.; Cohen, M.X.; Frick, C.; Kosel, M.; Brodesser, D.; Axmacher, N.; Joe, A.Y.; Kreft, M.; Lenartz, D.; Sturm, V. Deep brain stimulation to reward circuitry alleviates anhedonia in refractory major depression. Neuropsychopharmacology 2008, 33, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, U.; Lee, I.; Beermann, A.; Eack, S.; Keshavan, M.; Brady, R. Individual Variation in Functional Brain Network Topography is Linked to Schizophrenia Symptomatology. Schizophr. Bull. 2021, 47, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Carrozzino, D.; Patierno, C.; Fava, G.A.; Guidi, J. The Hamilton Rating Scales for Depression: A Critical Review of Clinimetric Properties of Different Versions. Psychother. Psychosom. 2020, 89, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Sen, Z.D.; Danyeli, L.V.; Woelfer, M.; Lamers, F.; Wagner, G.; Sobanski, T.; Walter, M. Linking atypical depression and insulin resistance-related disorders via low-grade chronic inflammation: Integrating the phenotypic, molecular and neuroanatomical dimensions. Brain Behav. Immun. 2021, 93, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Wang, L.Z.; Zhao, S.H.; Ning, Y.P.; Chan, R.C. Anhedonia and emotional word memory in patients with depression. Psychiatry Res. 2012, 200, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, I.; Everaert, J.; Dainer-Best, J.; Loeys, T.; Beevers, C.G.; Koster, E.H.W. Specificity and overlap of attention and memory biases in depression. J. Affect. Disord. 2018, 225, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liao, W.; Long, Z.; Tao, B.; Zhao, Q.; Luo, C.; Tamminga, C.A.; Keshavan, M.S.; Pearlson, G.D.; Clementz, B.A.; et al. Subtyping Schizophrenia Patients Based on Patterns of Structural Brain Alterations. Schizophr. Bull. 2022, 48, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Pearlson, G.D.; Liu, J.; Sui, J.; Yu, Q.; He, H.; Castro, E.; Calhoun, V.D. A group ICA based framework for evaluating resting fMRI markers when disease categories are unclear: Application to schizophrenia, bipolar, and schizoaffective disorders. Neuroimage 2015, 122, 272–280. [Google Scholar] [CrossRef]
- Shan, X.; Uddin, L.Q.; Xiao, J.; He, C.; Ling, Z.; Li, L.; Huang, X.; Chen, H.; Duan, X. Mapping the Heterogeneous Brain Structural Phenotype of Autism Spectrum Disorder Using the Normative Model. Biol. Psychiatry 2022, 91, 967–976. [Google Scholar] [CrossRef]
- Admon, R.; Nickerson, L.D.; Dillon, D.G.; Holmes, A.J.; Bogdan, R.; Kumar, P.; Dougherty, D.D.; Iosifescu, D.V.; Mischoulon, D.; Fava, M.; et al. Dissociable cortico-striatal connectivity abnormalities in major depression in response to monetary gains and penalties. Psychol. Med. 2015, 45, 121–131. [Google Scholar] [CrossRef]
- Admon, R.; Kaiser, R.H.; Dillon, D.G.; Beltzer, M.; Goer, F.; Olson, D.P.; Vitaliano, G.; Pizzagalli, D.A. Dopaminergic Enhancement of Striatal Response to Reward in Major Depression. Am. J. Psychiatry 2017, 174, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Arrondo, G.; Segarra, N.; Metastasio, A.; Ziauddeen, H.; Spencer, J.; Reinders, N.R.; Dudas, R.B.; Robbins, T.W.; Fletcher, P.C.; Murray, G.K. Reduction in ventral striatal activity when anticipating a reward in depression and schizophrenia: A replicated cross-diagnostic finding. Front. Psychol. 2015, 6, 1280. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekhar Pammi, V.S.; Pillai Geethabhavan Rajesh, P.; Kesavadas, C.; Rappai Mary, P.; Seema, S.; Radhakrishnan, A.; Sitaram, R. Neural loss aversion differences between depression patients and healthy individuals: A functional MRI investigation. Neuroradiol. J. 2015, 28, 97–105. [Google Scholar] [CrossRef] [PubMed]
- DelDonno, S.R.; Mickey, B.J.; Pruitt, P.J.; Stange, J.P.; Hsu, D.T.; Weldon, A.L.; Zubieta, J.K.; Langenecker, S.A. Influence of childhood adversity, approach motivation traits, and depression on individual differences in brain activation during reward anticipation. Biol. Psychol. 2019, 146, 107709. [Google Scholar] [CrossRef] [PubMed]
- Dillon, D.G.; Dobbins, I.G.; Pizzagalli, D.A. Weak reward source memory in depression reflects blunted activation of VTA/SN and parahippocampus. Soc. Cogn. Affect. Neurosci. 2014, 9, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Dichter, G.S.; Kozink, R.V.; McClernon, F.J.; Smoski, M.J. Remitted major depression is characterized by reward network hyperactivation during reward anticipation and hypoactivation during reward outcomes. J. Affect. Disord. 2012, 136, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Forbes, E.E.; Hariri, A.R.; Martin, S.L.; Silk, J.S.; Moyles, D.L.; Fisher, P.M.; Brown, S.M.; Ryan, N.D.; Birmaher, B.; Axelson, D.A.; et al. Altered striatal activation predicting real-world positive affect in adolescent major depressive disorder. Am. J. Psychiatry 2009, 166, 64–73. [Google Scholar] [CrossRef]
- Gaillard, C.; Guillod, M.; Ernst, M.; Federspiel, A.; Schoebi, D.; Recabarren, R.E.; Ouyang, X.; Mueller-Pfeiffer, C.; Horsch, A.; Homan, P.; et al. Striatal reactivity to reward under threat-of-shock and working memory load in adults at increased familial risk for major depression: A preliminary study. Neuroimage Clin. 2020, 26, 102193. [Google Scholar] [CrossRef]
- Gorka, S.M.; Huggins, A.A.; Fitzgerald, D.A.; Nelson, B.D.; Phan, K.L.; Shankman, S.A. Neural response to reward anticipation in those with depression with and without panic disorder. J. Affect. Disord. 2014, 164, 50–56. [Google Scholar] [CrossRef]
- Gotlib, I.H.; Hamilton, J.P.; Cooney, R.E.; Singh, M.K.; Henry, M.L.; Joormann, J. Neural processing of reward and loss in girls at risk for major depression. Arch. Gen. Psychiatry 2010, 67, 380–387. [Google Scholar] [CrossRef]
- Gradin, V.B.; Kumar, P.; Waiter, G.; Ahearn, T.; Stickle, C.; Milders, M.; Reid, I.; Hall, J.; Steele, J.D. Expected value and prediction error abnormalities in depression and schizophrenia. Brain 2011, 134, 1751–1764. [Google Scholar] [CrossRef] [PubMed]
- Hägele, C.; Schlagenhauf, F.; Rapp, M.; Sterzer, P.; Beck, A.; Bermpohl, F.; Stoy, M.; Ströhle, A.; Wittchen, H.U.; Dolan, R.J.; et al. Dimensional psychiatry: Reward dysfunction and depressive mood across psychiatric disorders. Psychopharmacology 2015, 232, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Hall, G.B.; Milne, A.M.; Macqueen, G.M. An fMRI study of reward circuitry in patients with minimal or extensive history of major depression. Eur. Arch. Psychiatry Clin. Neurosci. 2014, 264, 187–198. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhang, D.; Muhlert, N.; Elliott, R. Neural substrates for anticipation and consumption of social and monetary incentives in depression. Soc. Cogn. Affect. Neurosci. 2019, 14, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Narayanan, A.; Perlman, G.; Luking, K.; DeLorenzo, C.; Hajcak, G.; Klein, D.N.; Kotov, R.; Mohanty, A. Orbitofrontal cortex activity and connectivity predict future depression symptoms in adolescence. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 610–618. [Google Scholar] [CrossRef]
- Johnston, B.A.; Tolomeo, S.; Gradin, V.; Christmas, D.; Matthews, K.; Steele, J.D. Failure of hippocampal deactivation during loss events in treatment-resistant depression. Brain 2015, 138 Pt 9, 2766–2776. [Google Scholar] [CrossRef] [PubMed]
- Knutson, B.; Bhanji, J.P.; Cooney, R.E.; Atlas, L.Y.; Gotlib, I.H. Neural responses to monetary incentives in major depression. Biol. Psychiatry 2008, 63, 686–692. [Google Scholar] [CrossRef]
- Kumar, P.; Goer, F.; Murray, L.; Dillon, D.G.; Beltzer, M.L.; Cohen, A.L.; Brooks, N.H.; Pizzagalli, D.A. Impaired reward prediction error encoding and striatal-midbrain connectivity in depression. Neuropsychopharmacology 2018, 43, 1581–1588. [Google Scholar] [CrossRef]
- Liu, W.H.; Valton, V.; Wang, L.Z.; Zhu, Y.H.; Roiser, J.P. Association between habenula dysfunction and motivational symptoms in unmedicated major depressive disorder. Soc. Cogn. Affect. Neurosci. 2017, 12, 1520–1533. [Google Scholar] [CrossRef]
- Luking, K.R.; Pagliaccio, D.; Luby, J.L.; Barch, D.M. Depression Risk Predicts Blunted Neural Responses to Gains and Enhanced Responses to Losses in Healthy Children. J. Am. Acad. Child Adolesc. Psychiatry 2016, 55, 328–337. [Google Scholar] [CrossRef]
- Mori, A.; Okamoto, Y.; Okada, G.; Takagaki, K.; Jinnin, R.; Takamura, M.; Kobayakawa, M.; Yamawaki, S. Behavioral activation can normalize neural hypoactivation in subthreshold depression during a monetary incentive delay task. J. Affect. Disord. 2016, 189, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Olino, T.M.; McMakin, D.L.; Dahl, R.E.; Ryan, N.D.; Silk, J.S.; Birmaher, B.; Axelson, D.A.; Forbes, E.E. “I won, but I’m not getting my hopes up”: Depression moderates the relationship of outcomes and reward anticipation. Psychiatry Res. 2011, 194, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Olino, T.M.; McMakin, D.L.; Morgan, J.K.; Silk, J.S.; Birmaher, B.; Axelson, D.A.; Williamson, D.E.; Dahl, R.E.; Ryan, N.D.; Forbes, E.E. Reduced reward anticipation in youth at high-risk for unipolar depression: A preliminary study. Dev. Cogn. Neurosci. 2014, 8, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Pizzagalli, D.A.; Holmes, A.J.; Dillon, D.G.; Goetz, E.L.; Birk, J.L.; Bogdan, R.; Dougherty, D.D.; Iosifescu, D.V.; Rauch, S.L.; Fava, M. Reduced caudate and nucleus accumbens response to rewards in unmedicated individuals with major depressive disorder. Am. J. Psychiatry 2009, 166, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Redlich, R.; Dohm, K.; Grotegerd, D.; Opel, N.; Zwitserlood, P.; Heindel, W.; Arolt, V.; Kugel, H.; Dannlowski, U. Reward Processing in Unipolar and Bipolar Depression: A Functional MRI Study. Neuropsychopharmacology 2015, 40, 2623–2631. [Google Scholar] [CrossRef] [PubMed]
- Remijnse, P.L.; Nielen, M.M.; van Balkom, A.J.; Hendriks, G.J.; Hoogendijk, W.J.; Uylings, H.B.; Veltman, D.J. Differential frontal-striatal and paralimbic activity during reversal learning in major depressive disorder and obsessive-compulsive disorder. Psychol. Med. 2009, 39, 1503–1518. [Google Scholar] [CrossRef] [PubMed]
- Robinson, O.J.; Cools, R.; Carlisi, C.O.; Sahakian, B.J.; Drevets, W.C. Ventral striatum response during reward and punishment reversal learning in unmedicated major depressive disorder. Am. J. Psychiatry 2012, 169, 152–159. [Google Scholar] [CrossRef]
- Sankar, A.; Yttredahl, A.A.; Fourcade, E.W.; Mickey, B.J.; Love, T.M.; Langenecker, S.A.; Hsu, D.T. Dissociable Neural Responses to Monetary and Social Gain and Loss in Women with Major Depressive Disorder. Front. Behav. Neurosci. 2019, 13, 149. [Google Scholar] [CrossRef]
- Satterthwaite, T.D.; Kable, J.W.; Vandekar, L.; Katchmar, N.; Bassett, D.S.; Baldassano, C.F.; Ruparel, K.; Elliott, M.A.; Sheline, Y.I.; Gur, R.C.; et al. Common and Dissociable Dysfunction of the Reward System in Bipolar and Unipolar Depression. Neuropsychopharmacology 2015, 40, 2258–2268. [Google Scholar] [CrossRef]
- Schneider, M.; Elbau, I.G.; Nantawisarakul, T.; Pöhlchen, D.; Brückl, T.; Be, C.W.G.; Czisch, M.; Saemann, P.G.; Lee, M.D.; Binder, E.B.; et al. Pupil Dilation during Reward Anticipation Is Correlated to Depressive Symptom Load in Patients with Major Depressive Disorder. Brain Sci. 2020, 10, 906. [Google Scholar] [CrossRef]
- Segarra, N.; Metastasio, A.; Ziauddeen, H.; Spencer, J.; Reinders, N.R.; Dudas, R.B.; Arrondo, G.; Robbins, T.W.; Clark, L.; Fletcher, P.C.; et al. Abnormal Frontostriatal Activity During Unexpected Reward Receipt in Depression and Schizophrenia: Relationship to Anhedonia. Neuropsychopharmacology 2016, 41, 2001–2010. [Google Scholar] [CrossRef]
- Sharp, C.; Kim, S.; Herman, L.; Pane, H.; Reuter, T.; Strathearn, L. Major depression in mothers predicts reduced ventral striatum activation in adolescent female offspring with and without depression. J. Abnorm. Psychol. 2014, 123, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Smoski, M.J.; Felder, J.; Bizzell, J.; Green, S.R.; Ernst, M.; Lynch, T.R.; Dichter, G.S. fMRI of alterations in reward selection, anticipation, and feedback in major depressive disorder. J. Affect. Disord. 2009, 118, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Smoski, M.J.; Rittenberg, A.; Dichter, G.S. Major depressive disorder is characterized by greater reward network activation to monetary than pleasant image rewards. Psychiatry Res. 2011, 194, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Steele, J.D.; Kumar, P.; Ebmeier, K.P. Blunted response to feedback information in depressive illness. Brain 2007, 130 Pt 9, 2367–2374. [Google Scholar] [CrossRef] [PubMed]
- Stoy, M.; Schlagenhauf, F.; Sterzer, P.; Bermpohl, F.; Hägele, C.; Suchotzki, K.; Schmack, K.; Wrase, J.; Ricken, R.; Knutson, B.; et al. Hyporeactivity of ventral striatum towards incentive stimuli in unmedicated depressed patients normalizes after treatment with escitalopram. J. Psychopharmacol. 2012, 26, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Stringaris, A.; Vidal-Ribas Belil, P.; Artiges, E.; Lemaitre, H.; Gollier-Briant, F.; Wolke, S.; Vulser, H.; Miranda, R.; Penttila, J.; Struve, M.; et al. The Brain’s Response to Reward Anticipation and Depression in Adolescence: Dimensionality, Specificity, and Longitudinal Predictions in a Community-Based Sample. Am. J. Psychiatry 2015, 172, 1215–1223. [Google Scholar] [CrossRef]
- Simmons, W.K.; Burrows, K.; Avery, J.A.; Kerr, K.L.; Bodurka, J.; Savage, C.R.; Drevets, W.C. Depression-Related Increases and Decreases in Appetite: Dissociable Patterns of Aberrant Activity in Reward and Interoceptive Neurocircuitry. Am. J. Psychiatry 2016, 173, 418–428. [Google Scholar] [CrossRef]
- Takamura, M.; Okamoto, Y.; Okada, G.; Toki, S.; Yamamoto, T.; Ichikawa, N.; Mori, A.; Minagawa, H.; Takaishi, Y.; Fujii, Y.; et al. Patients with major depressive disorder exhibit reduced reward size coding in the striatum. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79 Pt B, 317–323. [Google Scholar] [CrossRef]
- Ubl, B.; Kuehner, C.; Kirsch, P.; Ruttorf, M.; Diener, C.; Flor, H. Altered neural reward and loss processing and prediction error signalling in depression. Soc. Cogn. Affect. Neurosci. 2015, 10, 1102–1112. [Google Scholar] [CrossRef]
- Ubl, B.; Kuehner, C.; Kirsch, P.; Ruttorf, M.; Flor, H.; Diener, C. Neural reward processing in individuals remitted from major depression. Psychol. Med. 2015, 45, 3549–3558. [Google Scholar] [CrossRef]
- Wiggins, J.L.; Schwartz, K.T.; Kryza-Lacombe, M.; Spechler, P.A.; Blankenship, S.L.; Dougherty, L.R. Neural reactivity to reward in school-age offspring of depressed mothers. J. Affect. Disord. 2017, 214, 81–88. [Google Scholar] [CrossRef]
- Yan, C.; Zhang, Y. DPARSF: A MATLAB Toolbox for “Pipeline” Data Analysis of Resting-State fMRI. Front. Syst. Neurosci. 2010, 4, 13. [Google Scholar] [CrossRef]
- Hahamy, A.; Calhoun, V.; Pearlson, G.; Harel, M.; Stern, N.; Attar, F.; Malach, R.; Salomon, R. Save the global: Global signal connectivity as a tool for studying clinical populations with functional magnetic resonance imaging. Brain Connect. 2014, 4, 395–403. [Google Scholar] [CrossRef]
- Albajes-Eizagirre, A.; Solanes, A.; Radua, J. Meta-analysis of non-statistically significant unreported effects. Stat. Methods Med. Res. 2019, 28, 3741–3754. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subgroup A | Subgroup B | Subgroup C | HC | p | |
|---|---|---|---|---|---|
| N (female %) | 11 (54.5%) | 21 (66.7%) | 24 (79%) | 32 (53%) | 0.145 |
| Mean (SD) | Mean (SD) | Mean (SD) | Mean (SD) | ||
| Age | 30.27 (7.02) | 32.81 (8.04) | 27.92 (5.22) | 29.59 (5.00) | 0.071 |
| Education (years) | 14.45 (3.50) | 12.38 (3.04) | 14.71 (3.07) | 14.59 (2.82) | 0.039, B < A = C = HC |
| Duration of illness | 5.82 (4.14) | 6.67 (4.77) | 6.10 (4.27) | - | 0.854 |
| HAMD | 21.09 (4.06) | 21.29 (3.39) | 21.67 (3.58) | 0.94 (0.95) | <0.001, A = B = C > HC |
| TEPS-anti | 39.82 (4.42) | 32.76 (4.04) | 24.25 (4.33) | 35.13 (6.10) | <0.001, A > B = HC > C |
| TEPS-con | 42.91 (5.52) | 33.67 (5.57) | 31.29 (4.77) | 42.53 (6.63) | <0.001, HC = A > B = C |
| TEPS-total | 82.73 (8.05) | 66.43 (6.68) | 55.54 (7.14) | 77.66 (11.60) | <0.001, HC = A > B > C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Y.; Ou, Y.; Yan, H.; Liu, F.; Li, H.; Li, P.; Xie, G.; Cui, X.; Guo, W. Uncovering the Neural Correlates of Anhedonia Subtypes in Major Depressive Disorder: Implications for Intervention Strategies. Biomedicines 2023, 11, 3138. https://doi.org/10.3390/biomedicines11123138
Ding Y, Ou Y, Yan H, Liu F, Li H, Li P, Xie G, Cui X, Guo W. Uncovering the Neural Correlates of Anhedonia Subtypes in Major Depressive Disorder: Implications for Intervention Strategies. Biomedicines. 2023; 11(12):3138. https://doi.org/10.3390/biomedicines11123138
Chicago/Turabian StyleDing, Yudan, Yangpan Ou, Haohao Yan, Feng Liu, Huabing Li, Ping Li, Guangrong Xie, Xilong Cui, and Wenbin Guo. 2023. "Uncovering the Neural Correlates of Anhedonia Subtypes in Major Depressive Disorder: Implications for Intervention Strategies" Biomedicines 11, no. 12: 3138. https://doi.org/10.3390/biomedicines11123138