

Exploring the Therapeutic Potential of Gamma-Aminobutyric Acid in Stress and Depressive Disorders through the Gut–Brain Axis

{kind=link}
Abstract
:1. Introduction
2. GABAergic Hypothesis in Depression
3. GABA and Cognitive Function in Depression
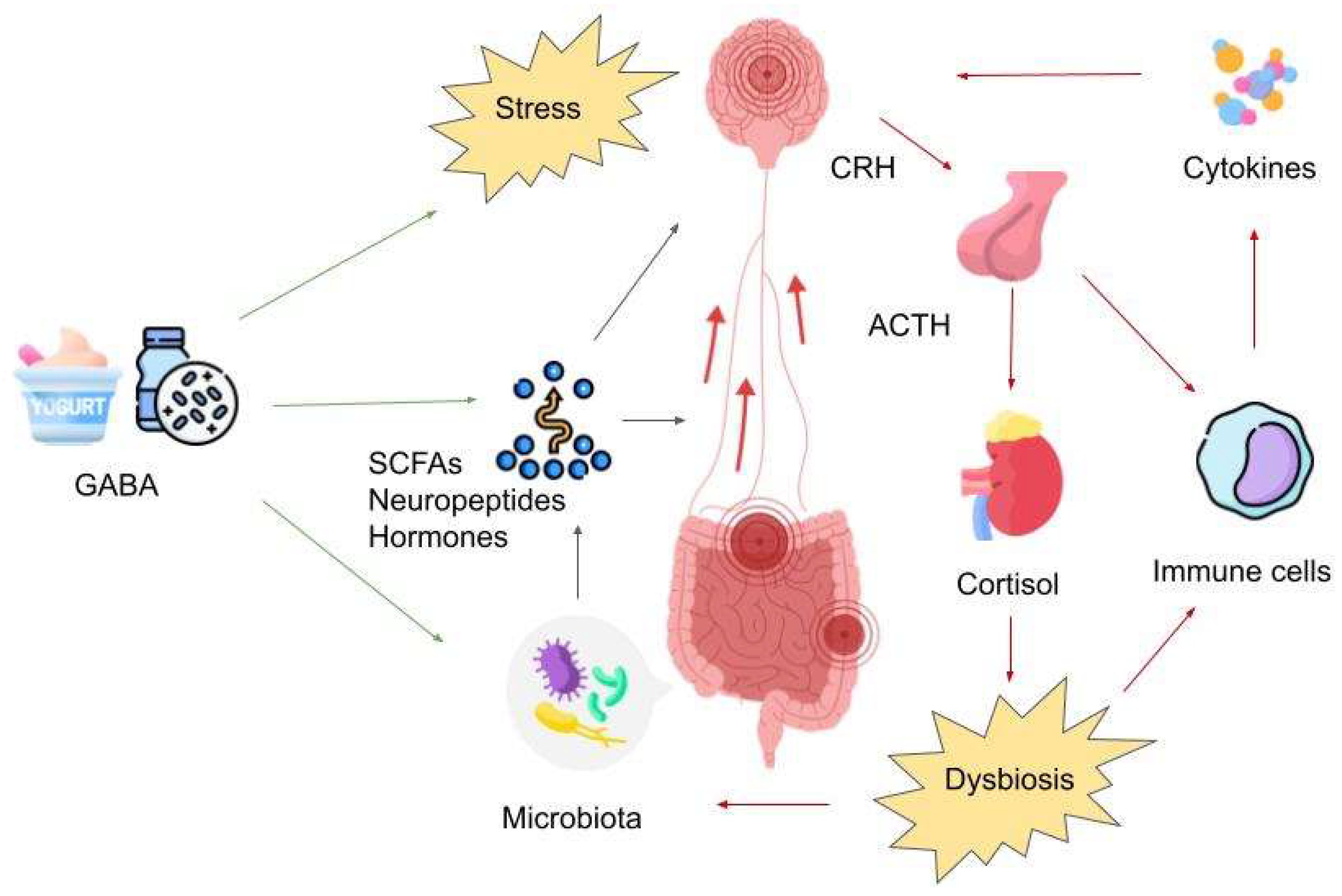
4. GABA and the Microbiota–Gut–Brain Axis
5. GABA’s Impact on the Enteric Nervous System
6. Traditional Diets and Their Impact on Mood
7. Fermented Foods Enriched with GABA
8. GABA-Enriched Fermented Foods as Neuro-Therapeutics
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Institute of Health Metrics and Evaluation Global Health Data Exchange (GHDx). Available online: https://ghdx.healthdata.org/keyword/unipolar-depressive-disorders (accessed on 20 November 2023).
- World Health Organization Depression. Available online: https://www.who.int/news-room/fact-sheets/detail/depression (accessed on 20 November 2023).
- Bierman, E.J.M.; Comijs, H.C.; Jonker, C.; Beekman, A.T.F. Effects of Anxiety Versus Depression on Cognition in Later Life. Am. J. Geriatr. Psychiatry 2005, 13, 686–693. [Google Scholar] [CrossRef]
- Dere, E.; Pause, B.M.; Pietrowsky, R. Emotion and Episodic Memory in Neuropsychiatric Disorders. Behav. Brain Res. 2010, 215, 162–171. [Google Scholar] [CrossRef]
- Majer, M.; Ising, M.; Künzel, H.; Binder, E.B.; Holsboer, F.; Modell, S.; Zihl, J. Impaired Divided Attention Predicts Delayed Response and Risk to Relapse in Subjects with Depressive Disorders. Psychol. Med. 2004, 34, 1453–1463. [Google Scholar] [CrossRef]
- McIntyre, R.S.; Cha, D.S.; Soczynska, J.K.; Woldeyohannes, H.O.; Gallaugher, L.A.; Kudlow, P.; Alsuwaidan, M.; Baskaran, A. Cognitive Deficits and Functional Outcomes in Major Depressive Disorder: Determinants, Substrates, and Treatment Interventions. Depress. Anxiety 2013, 30, 515–527. [Google Scholar] [CrossRef]
- Semkovska, M.; Quinlivan, L.; O’Grady, T.; Johnson, R.; Collins, A.; O’Connor, J.; Knittle, H.; Ahern, E.; Gload, T. Cognitive Function Following a Major Depressive Episode: A Systematic Review and Meta-Analysis. Lancet Psychiatry 2019, 6, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, P.E.; Fournier, A.-A.; Sisitsky, T.; Simes, M.; Berman, R.; Koenigsberg, S.H.; Kessler, R.C. The Economic Burden of Adults with Major Depressive Disorder in the United States (2010 and 2018). Pharmacoeconomics 2021, 39, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Bertolote, J.M.; Fleischmann, A.; De Leo, D.; Wasserman, D. Psychiatric Diagnoses and Suicide: Revisiting the Evidence. Crisis 2004, 25, 147–155. [Google Scholar] [CrossRef]
- Evans-Lacko, S.; Aguilar-Gaxiola, S.; Al-Hamzawi, A.; Alonso, J.; Benjet, C.; Bruffaerts, R.; Chiu, W.T.; Florescu, S.; de Girolamo, G.; Gureje, O.; et al. Socio-Economic Variations in the Mental Health Treatment Gap for People with Anxiety, Mood, and Substance Use Disorders: Results from the WHO World Mental Health (WMH) Surveys. Psychol. Med. 2018, 48, 1560–1571. [Google Scholar] [CrossRef]
- Malhi, G.S.; Bassett, D.; Boyce, P.; Bryant, R.; Fitzgerald, P.B.; Fritz, K.; Hopwood, M.; Lyndon, B.; Mulder, R.; Murray, G.; et al. Royal Australian and New Zealand College of Psychiatrists Clinical Practice Guidelines for Mood Disorders. Aust. N. Z. J. Psychiatry 2015, 49, 1087–1206. [Google Scholar] [CrossRef]
- Middleton, H.; Shaw, I.; Hull, S.; Feder, G. NICE Guidelines for the Management of Depression. BMJ 2005, 330, 267–268. [Google Scholar] [CrossRef] [PubMed]
- Härter, M.; Prien, P. The Diagnosis and Treatment of Unipolar Depression—National Disease Management Guideline. Dtsch. Arztebl. Int. 2023, 120, 355–361. [Google Scholar] [CrossRef]
- Cuijpers, P.; Karyotaki, E.; Eckshtain, D.; Ng, M.Y.; Corteselli, K.A.; Noma, H.; Quero, S.; Weisz, J.R. Psychotherapy for Depression Across Different Age Groups: A Systematic Review and Meta-Analysis. JAMA Psychiatry 2020, 77, 694–702. [Google Scholar] [CrossRef]
- Malhi, G.S.; Bell, E. Make News: Treatment-Resistant Depression—An Irreversible Problem in Need of a Reversible Solution? Aust. N. Z. J. Psychiatry 2020, 54, 111–113. [Google Scholar] [CrossRef]
- Malhi, G.S.; Das, P.; Mannie, Z.; Irwin, L. Treatment-Resistant Depression: Problematic Illness or a Problem in Our Approach? Br. J. Psychiatry 2019, 214, 1–3. [Google Scholar] [CrossRef]
- Thase, M.E.; Friedman, E.S.; Biggs, M.M.; Wisniewski, S.R.; Trivedi, M.H.; Luther, J.F.; Fava, M.; Nierenberg, A.A.; McGrath, P.J.; Warden, D.; et al. Cognitive Therapy versus Medication in Augmentation and Switch Strategies as Second-Step Treatments: A STAR*D Report. Am. J. Psychiatry 2007, 164, 739–752. [Google Scholar] [CrossRef]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Stewart, J.W.; Warden, D.; Niederehe, G.; Thase, M.E.; Lavori, P.W.; Lebowitz, B.D.; et al. Acute and Longer-Term Outcomes in Depressed Outpatients Requiring One or Several Treatment Steps: A STAR*D Report. Am. J. Psychiatry 2006, 163, 1905–1917. [Google Scholar] [CrossRef]
- Overall, J.E. Methodologic Issues in the Epidemiology of Treatment Resistant Depression. Contribution to Epidemiology. Pharmacopsychiatry 1974, 7, 129–138. [Google Scholar] [CrossRef]
- Kupfer, D.J.; Charney, D.S. Difficult-to-Treat Depression. Biol. Psychiatry 2003, 53, 633–634. [Google Scholar] [CrossRef]
- Bayes, A.J.; Parker, G.B. Comparison of Guidelines for the Treatment of Unipolar Depression: A Focus on Pharmacotherapy and Neurostimulation. Acta Psychiatr. Scand. 2018, 137, 459–471. [Google Scholar] [CrossRef]
- Bwalya, G.M.; Srinivasan, V.; Wang, M. Electroconvulsive Therapy Anesthesia Practice Patterns: Results of a UK Postal Survey. J. ECT 2011, 27, 81–85. [Google Scholar] [CrossRef]
- González-Pando, D.; González-Menéndez, A.; Aparicio-Basauri, V.; Sanz de la Garza, C.L.; Torracchi-Carrasco, J.E.; Pérez-Álvarez, M. Ethical Implications of Electroconvulsive Therapy: A Review. Ethical Hum. Psychol. Psychiatry 2021, 23, 25–38. [Google Scholar] [CrossRef]
- Gergel, T.; Howard, R.; Lawrence, R.; Seneviratne, T. Time to Acknowledge Good Electroconvulsive Therapy Research. Lancet Psychiatry 2021, 8, 1032–1033. [Google Scholar] [CrossRef] [PubMed]
- Luscher, B.; Shen, Q.; Sahir, N. The GABAergic Deficit Hypothesis of Major Depressive Disorder. Mol. Psychiatry 2011, 16, 383–406. [Google Scholar] [CrossRef] [PubMed]
- Spiering, M.J. The Discovery of GABA in the Brain. J. Biol. Chem. 2018, 293, 19159–19160. [Google Scholar] [CrossRef]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and Immunity Enhancement Effects of γ-Aminobutyric Acid (GABA) Administration in Humans. BioFactors 2006, 26, 201–208. [Google Scholar] [CrossRef]
- Lee, B.-J.; Kim, J.-S.; Kang, Y.M.; Lim, J.-H.; Kim, Y.-M.; Lee, M.-S.; Jeong, M.-H.; Ahn, C.-B.; Je, J.-Y. Antioxidant Activity and γ-Aminobutyric Acid (GABA) Content in Sea Tangle Fermented by Lactobacillus brevis BJ20 Isolated from Traditional Fermented Foods. Food Chem. 2010, 122, 271–276. [Google Scholar] [CrossRef]
- Li, W.; Wei, M.; Wu, J.; Rui, X.; Dong, M. Novel Fermented Chickpea Milk with Enhanced Level of γ -Aminobutyric Acid and Neuroprotective Effect on PC12 Cells. PeerJ 2016, 4, e2292. [Google Scholar] [CrossRef]
- Deng, Y.; Xu, L.; Zeng, X.; Li, Z.; Qin, B.; He, N. New Perspective of GABA as an Inhibitor of Formation of Advanced Lipoxidation End-Products: It’s Interaction with Malondiadehyde. J. Biomed. Nanotechnol. 2010, 6, 318–324. [Google Scholar] [CrossRef]
- Aoki, H.; Furuya, Y.; Endo, Y.; Fujimoto, K. Effect of Gamma-Aminobutyric Acid-Enriched Tempeh-like Fermented Soybean (GABA-Tempeh) on the Blood Pressure of Spontaneously Hypertensive Rats. Biosci. Biotechnol. Biochem. 2003, 67, 1806–1808. [Google Scholar] [CrossRef]
- Jang, E.K.; Kim, N.Y.; Ahn, H.J.; Ji, G.E. γ-Aminobutyric Acid (GABA) Production and Angiotensin-I Converting Enzyme (ACE) Inhibitory Activity of Fermented Soybean Containing Sea Tangle by the Co-Culture of Lactobacillus brevis with Aspergillus oryzae. J. Microbiol. Biotechnol. 2015, 25, 1315–1320. [Google Scholar] [CrossRef]
- Huang, Q.; Liu, C.; Wang, C.; Hu, Y.; Qiu, L.; Xu, P. Neurotransmitter γ-Aminobutyric Acid-Mediated Inhibition of the Invasive Ability of Cholangiocarcinoma Cells. Oncol. Lett. 2011, 2, 519–523. [Google Scholar] [CrossRef]
- Foster, A.C.; Kemp, J.A. Glutamate- and GABA-Based CNS Therapeutics. Curr. Opin. Pharmacol. 2006, 6, 7–17. [Google Scholar] [CrossRef]
- Patel, A.B.; Rothman, D.L.; Cline, G.W.; Behar, K.L. Glutamine Is the Major Precursor for GABA Synthesis in Rat Neocortex in Vivo Following Acute GABA-Transaminase Inhibition. Brain Res. 2001, 919, 207–220. [Google Scholar] [CrossRef]
- Möhler, H. Molecular Regulation of Cognitive Functions and Developmental Plasticity: Impact of GABAA Receptors. J. Neurochem. 2007, 102, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mody, I.; Pearce, R.A. Diversity of Inhibitory Neurotransmission through GABA(A) Receptors. Trends Neurosci. 2004, 27, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Mombereau, C.; Kaupmann, K.; Froestl, W.; Sansig, G.; van der Putten, H.; Cryan, J.F. Genetic and Pharmacological Evidence of a Role for GABA(B) Receptors in the Modulation of Anxiety- and Antidepressant-like Behavior. Neuropsychopharmacology 2004, 29, 1050–1062. [Google Scholar] [CrossRef]
- Mombereau, C.; Kaupmann, K.; Gassmann, M.; Bettler, B.; van der Putten, H.; Cryan, J.F. Altered Anxiety and Depression-Related Behaviour in Mice Lacking GABAB(2) Receptor Subunits. NeuroReport 2005, 16, 307–310. [Google Scholar] [CrossRef]
- Gonzalez-Burgos, G.; Fish, K.N.; Lewis, D.A. GABA Neuron Alterations, Cortical Circuit Dysfunction and Cognitive Deficits in Schizophrenia. Neural Plast. 2011, 2011, 723184. [Google Scholar] [CrossRef]
- Brambilla, P.; Perez, J.; Barale, F.; Schettini, G.; Soares, J.C. GABAergic Dysfunction in Mood Disorders. Mol. Psychiatry 2003, 8, 721–737. [Google Scholar] [CrossRef]
- Horder, J.; Petrinovic, M.M.; Mendez, M.A.; Bruns, A.; Takumi, T.; Spooren, W.; Barker, G.J.; Künnecke, B.; Murphy, D.G. Glutamate and GABA in Autism Spectrum Disorder-a Translational Magnetic Resonance Spectroscopy Study in Man and Rodent Models. Transl. Psychiatry 2018, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Lener, M.S.; Niciu, M.J.; Ballard, E.D.; Park, M.; Park, L.T.; Nugent, A.C.; Zarate, C.A. Glutamate and Gamma-Aminobutyric Acid Systems in the Pathophysiology of Major Depression and Antidepressant Response to Ketamine. Biol. Psychiatry 2017, 81, 886–897. [Google Scholar] [CrossRef]
- Knudsen, M.K.; Near, J.; Blicher, A.B.; Videbech, P.; Blicher, J.U. Magnetic Resonance (MR) Spectroscopic Measurement of γ-Aminobutyric Acid (GABA) in Major Depression before and after Electroconvulsive Therapy. Acta Neuropsychiatr. 2019, 31, 17–26. [Google Scholar] [CrossRef]
- Sanacora, G.; Treccani, G.; Popoli, M. Towards a Glutamate Hypothesis of Depression: An Emerging Frontier of Neuropsychopharmacology for Mood Disorders. Neuropharmacology 2012, 62, 63–77. [Google Scholar] [CrossRef]
- Lener, M.S.; Iosifescu, D. V In Pursuit of Neuroimaging Biomarkers to Guide Treatment Selection in Major Depressive Disorder: A Review of the Literature. Ann. N. Y. Acad. Sci. 2015, 1344, 50–65. [Google Scholar] [CrossRef]
- Yüksel, C.; Öngür, D. Magnetic Resonance Spectroscopy Studies of Glutamate-Related Abnormalities in Mood Disorders. Biol. Psychiatry 2010, 68, 785–794. [Google Scholar] [CrossRef]
- Arnone, D.; Mumuni, A.N.; Jauhar, S.; Condon, B.; Cavanagh, J. Indirect Evidence of Selective Glial Involvement in Glutamate-Based Mechanisms of Mood Regulation in Depression: Meta-Analysis of Absolute Prefrontal Neuro-Metabolic Concentrations. Eur. Neuropsychopharmacol. 2015, 25, 1109–1117. [Google Scholar] [CrossRef]
- Yildiz-Yesiloglu, A.; Ankerst, D.P. Review of 1H Magnetic Resonance Spectroscopy Findings in Major Depressive Disorder: A Meta-Analysis. Psychiatry Res. 2006, 147, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Rajkowska, G.; O’Dwyer, G.; Teleki, Z.; Stockmeier, C.A.; Miguel-Hidalgo, J.J. GABAergic Neurons Immunoreactive for Calcium Binding Proteins Are Reduced in the Prefrontal Cortex in Major Depression. Neuropsychopharmacology 2007, 32, 471–482. [Google Scholar] [CrossRef]
- Petty, F.; Schlesser, M.A. Plasma GABA in Affective Illness. J. Affect. Disord. 1981, 3, 339–343. [Google Scholar] [CrossRef]
- Petty, F.; Sherman, A.D. Plasma GABA Levels in Psychiatric Illness. J. Affect. Disord. 1984, 6, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Gerner, R.H.; Hare, T.A. CSF GABA in Normal Subjects and Patients with Depression, Schizophrenia, Mania, and Anorexia Nervosa. Am. J. Psychiatry 1981, 138, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Honig, A.; Bartlett, J.R.; Bouras, N.; Bridges, P.K. Amino Acid Levels in Depression: A Preliminary Investigation. J. Psychiatr. Res. 1988, 22, 159–164. [Google Scholar] [CrossRef]
- Price, R.B.; Shungu, D.C.; Mao, X.; Nestadt, P.; Kelly, C.; Collins, K.A.; Murrough, J.W.; Charney, D.S.; Mathew, S.J. Amino Acid Neurotransmitters Assessed by Proton Magnetic Resonance Spectroscopy: Relationship to Treatment Resistance in Major Depressive Disorder. Biol. Psychiatry 2009, 65, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Schür, R.R.; Draisma, L.W.R.; Wijnen, J.P.; Boks, M.P.; Koevoets, M.G.J.C.; Joëls, M.; Klomp, D.W.; Kahn, R.S.; Vinkers, C.H. Brain GABA Levels across Psychiatric Disorders: A Systematic Literature Review and Meta-Analysis of 1H-MRS Studies. Hum. Brain Mapp. 2016, 37, 3337–3352. [Google Scholar] [CrossRef]
- Kohrs, R.; Durieux, M.E. Ketamine: Teaching an old drug new tricks. Anesth. Analg. 1998, 87, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Fond, G.; Loundou, A.; Rabu, C.; Macgregor, A.; Lançon, C.; Brittner, M.; Micoulaud-Franchi, J.-A.; Richieri, R.; Courtet, P.; Abbar, M.; et al. Ketamine Administration in Depressive Disorders: A Systematic Review and Meta-Analysis. Psychopharmacology 2014, 231, 3663–3676. [Google Scholar] [CrossRef] [PubMed]
- Newport, D.J.; Carpenter, L.L.; McDonald, W.M.; Potash, J.B.; Tohen, M.; Nemeroff, C.B. Ketamine and Other NMDA Antagonists: Early Clinical Trials and Possible Mechanisms in Depression. Am. J. Psychiatry 2015, 172, 950–966. [Google Scholar] [CrossRef] [PubMed]
- Esel, E.; Kose, K.; Hacimusalar, Y.; Ozsoy, S.; Kula, M.; Candan, Z.; Turan, T. The Effects of Electroconvulsive Therapy on GABAergic Function in Major Depressive Patients. J. ECT 2008, 24, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Vitali, M.; Tedeschini, E.; Mistretta, M.; Fehling, K.; Aceti, F.; Ceccanti, M.; Fava, M. Adjunctive Pregabalin in Partial Responders with Major Depressive Disorder and Residual Anxiety. J. Clin. Psychopharmacol. 2013, 33, 95–98. [Google Scholar] [CrossRef]
- Stein, D.J.; Baldwin, D.S.; Baldinetti, F.; Mandel, F. Efficacy of Pregabalin in Depressive Symptoms Associated with Generalized Anxiety Disorder: A Pooled Analysis of 6 Studies. Eur. Neuropsychopharmacol. 2008, 18, 422–430. [Google Scholar] [CrossRef]
- Sanacora, G.; Fenton, L.R.; Fasula, M.K.; Rothman, D.L.; Levin, Y.; Krystal, J.H.; Mason, G.F. Cortical Gamma-Aminobutyric Acid Concentrations in Depressed Patients Receiving Cognitive Behavioral Therapy. Biol. Psychiatry 2006, 59, 284–286. [Google Scholar] [CrossRef]
- Sanacora, G.; Mason, G.F.; Rothman, D.L.; Hyder, F.; Ciarcia, J.J.; Ostroff, R.B.; Berman, R.M.; Krystal, J.H. Increased Cortical GABA Concentrations in Depressed Patients Receiving ECT. Am. J. Psychiatry 2003, 160, 577–579. [Google Scholar] [CrossRef]
- Jiang, Y.; Peng, T.; Gaur, U.; Silva, M.; Little, P.; Chen, Z.; Qiu, W.; Zhang, Y.; Zheng, W. Role of Corticotropin Releasing Factor in the Neuroimmune Mechanisms of Depression: Examination of Current Pharmaceutical and Herbal Therapies. Front. Cell. Neurosci. 2019, 13, 290. [Google Scholar] [CrossRef] [PubMed]
- Cobb, S.R.; Buhl, E.H.; Halasy, K.; Paulsen, O.; Somogyi, P. Synchronization of Neuronal Activity in Hippocampus by Individual GABAergic Interneurons. Nature 1995, 378, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Q.; Barberis, A.; Higley, M.J. Preserving the Balance: Diverse Forms of Long-Term GABAergic Synaptic Plasticity. Nat. Rev. Neurosci. 2019, 20, 272–281. [Google Scholar] [CrossRef]
- Schmidt-Wilcke, T.; Fuchs, E.; Funke, K.; Vlachos, A.; Müller-Dahlhaus, F.; Puts, N.A.J.; Harris, R.E.; Edden, R.A.E. GABA—From Inhibition to Cognition: Emerging Concepts. Neuroscientist 2018, 24, 501–515. [Google Scholar] [CrossRef]
- Möhler, H.; Rudolph, U. Disinhibition, an Emerging Pharmacology of Learning and Memory. F1000Research 2017, 6, 101. [Google Scholar] [CrossRef]
- Collinson, N.; Kuenzi, F.M.; Jarolimek, W.; Maubach, K.A.; Cothliff, R.; Sur, C.; Smith, A.; Otu, F.M.; Howell, O.; Atack, J.R.; et al. Enhanced Learning and Memory and Altered GABAergic Synaptic Transmission in Mice Lacking the Alpha 5 Subunit of the GABAA Receptor. J. Neurosci. 2002, 22, 5572–5580. [Google Scholar] [CrossRef]
- Martel, G.; Dutar, P.; Epelbaum, J.; Viollet, C. Somatostatinergic Systems: An Update on Brain Functions in Normal and Pathological Aging. Front. Endocrinol. 2012, 3, 154. [Google Scholar] [CrossRef]
- Kumar, U. Expression of Somatostatin Receptor Subtypes (SSTR1-5) in Alzheimer’s Disease Brain: An Immunohistochemical Analysis. Neuroscience 2005, 134, 525–538. [Google Scholar] [CrossRef]
- Fee, C.; Banasr, M.; Sibille, E. Somatostatin-Positive Gamma-Aminobutyric Acid Interneuron Deficits in Depression: Cortical Microcircuit and Therapeutic Perspectives. Biol. Psychiatry 2017, 82, 549–559. [Google Scholar] [CrossRef]
- Gentet, L.J.; Kremer, Y.; Taniguchi, H.; Huang, Z.J.; Staiger, J.F.; Petersen, C.C.H. Unique Functional Properties of Somatostatin-Expressing GABAergic Neurons in Mouse Barrel Cortex. Nat. Neurosci. 2012, 15, 607–612. [Google Scholar] [CrossRef]
- Piantadosi, S.C.; French, B.J.; Poe, M.M.; Timić, T.; Marković, B.D.; Pabba, M.; Seney, M.L.; Oh, H.; Orser, B.A.; Savić, M.M.; et al. Sex-Dependent Anti-Stress Effect of an A5 Subunit Containing GABAA Receptor Positive Allosteric Modulator. Front. Pharmacol. 2016, 7, 446. [Google Scholar] [CrossRef]
- Koh, M.T.; Rosenzweig-Lipson, S.; Gallagher, M. Selective GABA(A) A5 Positive Allosteric Modulators Improve Cognitive Function in Aged Rats with Memory Impairment. Neuropharmacology 2013, 64, 145–152. [Google Scholar] [CrossRef]
- Prévot, T.; Sibille, E. Altered GABA-Mediated Information Processing and Cognitive Dysfunctions in Depression and Other Brain Disorders. Mol. Psychiatry 2021, 26, 151–167. [Google Scholar] [CrossRef]
- Hasler, G.; Northoff, G. Discovering Imaging Endophenotypes for Major Depression. Mol. Psychiatry 2011, 16, 604–619. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Cruz-Pereira, J.S.; Rea, K.; Nolan, Y.M.; O’Leary, O.F.; Dinan, T.G.; Cryan, J.F. Depression’s Unholy Trinity: Dysregulated Stress, Immunity, and the Microbiome. Annu. Rev. Psychol. 2020, 71, 49–78. [Google Scholar] [CrossRef]
- Winter, G.; Hart, R.A.; Charlesworth, R.P.G.; Sharpley, C.F. Gut Microbiome and Depression: What We Know and What We Need to Know. Rev. Neurosci. 2018, 29, 629–643. [Google Scholar] [CrossRef]
- Kelly, J.R.; Borre, Y.; O’ Brien, C.; Patterson, E.; El Aidy, S.; Deane, J.; Kennedy, P.J.; Beers, S.; Scott, K.; Moloney, G.; et al. Transferring the Blues: Depression-Associated Gut Microbiota Induces Neurobehavioural Changes in the Rat. J. Psychiatr. Res. 2016, 82, 109–118. [Google Scholar] [CrossRef]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut Microbiome Remodeling Induces Depressive-like Behaviors through a Pathway Mediated by the Host’s Metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef]
- Li, Y.; Ning, L.; Yin, Y.; Wang, R.; Zhang, Z.; Hao, L.; Wang, B.; Zhao, X.; Yang, X.; Yin, L.; et al. Age-Related Shifts in Gut Microbiota Contribute to Cognitive Decline in Aged Rats. Aging 2020, 12, 7801–7817. [Google Scholar] [CrossRef]
- Boehme, M.; Guzzetta, K.E.; Bastiaanssen, T.F.S.; van de Wouw, M.; Moloney, G.M.; Gual-Grau, A.; Spichak, S.; Olavarría-Ramírez, L.; Fitzgerald, P.; Morillas, E.; et al. Microbiota from Young Mice Counteracts Selective Age-Associated Behavioral Deficits. Nat. Aging 2021, 1, 666–676. [Google Scholar] [CrossRef]
- Liu, L.; Wang, H.; Zhang, H.; Chen, X.; Zhang, Y.; Wu, J.; Zhao, L.; Wang, D.; Pu, J.; Ji, P.; et al. Toward a Deeper Understanding of Gut Microbiome in Depression: The Promise of Clinical Applicability. Adv. Sci. 2022, 9, e2203707. [Google Scholar] [CrossRef]
- Firth, J.; Gangwisch, J.E.; Borsini, A.; Wootton, R.E.; Mayer, E.A. Food and Mood: How Do Diet and Nutrition Affect Mental Wellbeing? BMJ 2020, 369, m2382. [Google Scholar] [CrossRef]
- Liwinski, T.; Elinav, E. Harnessing the Microbiota for Therapeutic Purposes. Am. J. Transplant. 2020, 20, 1482–1488. [Google Scholar] [CrossRef]
- Schaub, A.-C.; Schneider, E.; Vazquez-Castellanos, J.F.; Schweinfurth, N.; Kettelhack, C.; Doll, J.P.K.; Yamanbaeva, G.; Mählmann, L.; Brand, S.; Beglinger, C.; et al. Clinical, Gut Microbial and Neural Effects of a Probiotic Add-on Therapy in Depressed Patients: A Randomized Controlled Trial. Transl. Psychiatry 2022, 12, 227. [Google Scholar] [CrossRef]
- Ng, Q.X.; Peters, C.; Ho, C.Y.X.; Lim, D.Y.; Yeo, W.-S. A Meta-Analysis of the Use of Probiotics to Alleviate Depressive Symptoms. J. Affect. Disord. 2018, 228, 13–19. [Google Scholar] [CrossRef]
- Nikolova, V.L.; Cleare, A.J.; Young, A.H.; Stone, J.M. Acceptability, Tolerability, and Estimates of Putative Treatment Effects of Probiotics as Adjunctive Treatment in Patients with Depression. JAMA Psychiatry 2023, 80, 842–847. [Google Scholar] [CrossRef]
- Diez-Gutiérrez, L.; San Vicente, L.; Barrón, L.J.R.; Villarán, M.d.C.; Chávarri, M. Gamma-Aminobutyric Acid and Probiotics: Multiple Health Benefits and Their Future in the Global Functional Food and Nutraceuticals Market. J. Funct. Foods 2020, 64, 103669. [Google Scholar] [CrossRef]
- Vodnar, D.; Paucean, A.; Dulf, F.; Socaciu, C. HPLC Characterization of Lactic Acid Formation and FTIR Fingerprint of Probiotic Bacteria during Fermentation Processes. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 109–113. [Google Scholar]
- Aslam, H.; Green, J.; Jacka, F.N.; Collier, F.; Berk, M.; Pasco, J.; Dawson, S.L. Fermented Foods, the Gut and Mental Health: A Mechanistic Overview with Implications for Depression and Anxiety. Nutr. Neurosci. 2020, 23, 659–671. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of Bacteria–Gut–Brain Signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef]
- Shaibe, E.; Metzer, E.; Halpern, Y.S. Metabolic Pathway for the Utilization of L-Arginine, L-Ornithine, Agmatine, and Putrescine as Nitrogen Sources in Escherichia Coli K-12. J. Bacteriol. 1985, 163, 933–937. [Google Scholar] [CrossRef]
- Smith, D.K.; Kassam, T.; Singh, B.; Elliott, J.F. Escherichia Coli Has Two Homologous Glutamate Decarboxylase Genes That Map to Distinct Loci. J. Bacteriol. 1992, 174, 5820–5826. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Johnson, C.; Luk, B.; Uribe, G.; Fu, Y.; Oezguen, N.; Matsunami, R.K.; Lugo, M.; Major, A.; Mori-Akiyama, Y.; et al. GABA-producing Bifidobacterium dentium Modulates Visceral Sensitivity in the Intestine. Neurogastroenterol. Motil. 2017, 29, e12904. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus Strain Regulates Emotional Behavior and Central GABA Receptor Expression in a Mouse via the Vagus Nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-Altering Microorganisms: The Impact of the Gut Microbiota on Brain and Behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric Acid Production by Culturable Bacteria from the Human Intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Auteri, M.; Zizzo, M.G.; Serio, R. GABA and GABA Receptors in the Gastrointestinal Tract: From Motility to Inflammation. Pharmacol. Res. 2015, 93, 11–21. [Google Scholar] [CrossRef]
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef]
- Bravo, J.A.; Julio-Pieper, M.; Forsythe, P.; Kunze, W.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Communication between Gastrointestinal Bacteria and the Nervous System. Curr. Opin. Pharmacol. 2012, 12, 667–672. [Google Scholar] [CrossRef]
- Carron, R.; Roncon, P.; Lagarde, S.; Dibué, M.; Zanello, M.; Bartolomei, F. Latest Views on the Mechanisms of Action of Surgically Implanted Cervical Vagal Nerve Stimulation in Epilepsy. Neuromodul. Technol. Neural Interface 2023, 26, 498–506. [Google Scholar] [CrossRef]
- Austelle, C.W.; O’Leary, G.H.; Thompson, S.; Gruber, E.; Kahn, A.; Manett, A.J.; Short, B.; Badran, B.W. A Comprehensive Review of Vagus Nerve Stimulation for Depression. Neuromodul. Technol. Neural Interface 2022, 25, 309–315. [Google Scholar] [CrossRef]
- Marrosu, F.; Serra, A.; Maleci, A.; Puligheddu, M.; Biggio, G.; Piga, M. Correlation between GABAA Receptor Density and Vagus Nerve Stimulation in Individuals with Drug-Resistant Partial Epilepsy. Epilepsy Res. 2003, 55, 59–70. [Google Scholar] [CrossRef]
- Ben-Menachem, E.; Hamberger, A.; Hedner, T.; Hammond, E.J.; Uthman, B.M.; Slater, J.; Treig, T.; Stefan, H.; Ramsay, R.E.; Wernicke, J.F.; et al. Effects of Vagus Nerve Stimulation on Amino Acids and Other Metabolites in the CSF of Patients with Partial Seizures. Epilepsy Res. 1995, 20, 221–227. [Google Scholar] [CrossRef]
- Junker, B. Fermentation. In Kirk-Othmer Encyclopedia of Chemical Technology; Wiley: Hoboken, NJ, USA, 2004. [Google Scholar]
- Steinkraus, K.H. Fermentations in World Food Processing. Compr. Rev. Food Sci. Food Saf. 2002, 1, 23–32. [Google Scholar] [CrossRef]
- Henderson, J.S.; Joyce, R.A.; Hall, G.R.; Hurst, W.J.; McGovern, P.E. Chemical and Archaeological Evidence for the Earliest Cacao Beverages. Proc. Natl. Acad. Sci. USA 2007, 104, 18937–18940. [Google Scholar] [CrossRef] [PubMed]
- McGovern Anticancer Activity of Botanical Compounds in Ancient Fermented Beverages (Review). Int. J. Oncol. 2010, 37, 5–14. [CrossRef]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.; et al. Fermented Beverages of Pre- and Proto-Historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [PubMed]
- Caplice, E. Food Fermentations: Role of Microorganisms in Food Production and Preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Borresen, E.C.; Henderson, A.J.; Kumar, A.; Weir, T.L.; Ryan, E.P. Fermented Foods: Patented Approaches and Formulations for Nutritional Supplementation and Health Promotion. Recent Pat. Food Nutr. Agric. 2012, 4, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Selhub, E.M.; Logan, A.C.; Bested, A.C. Fermented Foods, Microbiota, and Mental Health: Ancient Practice Meets Nutritional Psychiatry. J. Physiol. Anthropol. 2014, 33, 2. [Google Scholar] [CrossRef]
- Hidaka, B.H. Depression as a Disease of Modernity: Explanations for Increasing Prevalence. J. Affect. Disord. 2012, 140, 205–214. [Google Scholar] [CrossRef]
- Logan, A.C.; Selhub, E.M. Vis Medicatrix Naturae: Does Nature “Minister to the Mind”? Biopsychosoc. Med. 2012, 6, 11. [Google Scholar] [CrossRef]
- Sanchez-Villegas, A.; Martínez-González, M.A. Diet, a New Target to Prevent Depression? BMC Med. 2013, 11, 3. [Google Scholar] [CrossRef]
- Li, N.P.; van Vugt, M.; Colarelli, S.M. The Evolutionary Mismatch Hypothesis: Implications for Psychological Science. Curr. Dir. Psychol. Sci. 2018, 27, 38–44. [Google Scholar] [CrossRef]
- Chang, Y.; Durante, K.M. Why Consumers Have Everything but Happiness: An Evolutionary Mismatch Perspective. Curr. Opin. Psychol. 2022, 46, 101347. [Google Scholar] [CrossRef]
- Hesseltine, C.W.; Wang, H.L. Traditional Fermented Foods. Biotechnol. Bioeng. 1967, 9, 275–288. [Google Scholar] [CrossRef]
- Takashima, N.; Katayama, A.; Dokai Mochimasu, K.; Hishii, S.; Suzuki, H.; Miyatake, N. A Pilot Study of the Relationship between Diet and Mental Health in Community Dwelling Japanese Women. Medicina 2019, 55, 513. [Google Scholar] [CrossRef]
- Ventriglio, A.; Sancassiani, F.; Contu, M.P.; Latorre, M.; Di Slavatore, M.; Fornaro, M.; Bhugra, D. Mediterranean Diet and Its Benefits on Health and Mental Health: A Literature Review. Clin. Pract. Epidemiol. Ment. Health 2020, 16, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Toyomaki, A.; Miyazaki, A.; Nakai, Y.; Yamaguchi, A.; Kubo, C.; Suzuki, J.; Ohkubo, I.; Shimizu, M.; Musashi, M.; et al. Mediators of the Effects of Rice Intake on Health in Individuals Consuming a Traditional Japanese Diet Centered on Rice. PLoS ONE 2017, 12, e0185816. [Google Scholar] [CrossRef]
- Nanri, A.; Kimura, Y.; Matsushita, Y.; Ohta, M.; Sato, M.; Mishima, N.; Sasaki, S.; Mizoue, T. Dietary Patterns and Depressive Symptoms among Japanese Men and Women. Eur. J. Clin. Nutr. 2010, 64, 832–839. [Google Scholar] [CrossRef]
- Nanri, A.; Mizoue, T.; Poudel-Tandukar, K.; Noda, M.; Kato, M.; Kurotani, K.; Goto, A.; Oba, S.; Inoue, M.; Tsugane, S. Dietary Patterns and Suicide in Japanese Adults: The Japan Public Health Center-Based Prospective Study. Br. J. Psychiatry 2013, 203, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miyaki, K.; Tsutsumi, A.; Hashimoto, H.; Kawakami, N.; Takahashi, M.; Shimazu, A.; Inoue, A.; Kurioka, S.; Kakehashi, M.; et al. Japanese Dietary Pattern Consistently Relates to Low Depressive Symptoms and It Is Modified by Job Strain and Worksite Supports. J. Affect. Disord. 2013, 150, 490–498. [Google Scholar] [CrossRef]
- Sanada, M.; Imai, T.; Sezaki, A.; Miyamoto, K.; Kawase, F.; Shirai, Y.; Abe, C.; Suzuki, N.; Inden, A.; Kato, T.; et al. Changes in the Association between the Traditional Japanese Diet Score and Suicide Rates over 26 Years: A Global Comparative Study. J. Affect. Disord. 2021, 294, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Murooka, Y.; Yamshita, M. Traditional Healthful Fermented Products of Japan. J. Ind. Microbiol. Biotechnol. 2008, 35, 791–798. [Google Scholar] [CrossRef]
- Jacka, F.N.; Mykletun, A.; Berk, M.; Bjelland, I.; Tell, G.S. The Association Between Habitual Diet Quality and the Common Mental Disorders in Community-Dwelling Adults. Psychosom. Med. 2011, 73, 483–490. [Google Scholar] [CrossRef]
- Jacka, F.N.; Pasco, J.A.; Mykletun, A.; Williams, L.J.; Hodge, A.M.; O’Reilly, S.L.; Nicholson, G.C.; Kotowicz, M.A.; Berk, M. Association of Western and Traditional Diets with Depression and Anxiety in Women. Am. J. Psychiatry 2010, 167, 305–311. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Delgado-Rodríguez, M.; Alonso, A.; Schlatter, J.; Lahortiga, F.; Majem, L.S.; Martínez-González, M.A. Association of the Mediterranean Dietary Pattern with the Incidence of Depression. Arch. Gen. Psychiatry 2009, 66, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Akbaraly, T.N.; Brunner, E.J.; Ferrie, J.E.; Marmot, M.G.; Kivimaki, M.; Singh-Manoux, A. Dietary Pattern and Depressive Symptoms in Middle Age. Br. J. Psychiatry 2009, 195, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Skarupski, K.A.; Tangney, C.C.; Li, H.; Evans, D.A.; Morris, M.C. Mediterranean Diet and Depressive Symptoms among Older Adults over Time. J. Nutr. Health Aging 2013, 17, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Rienks, J.; Dobson, A.J.; Mishra, G.D. Mediterranean Dietary Pattern and Prevalence and Incidence of Depressive Symptoms in Mid-Aged Women: Results from a Large Community-Based Prospective Study. Eur. J. Clin. Nutr. 2013, 67, 75–82. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Henríquez, P.; Bes-Rastrollo, M.; Doreste, J. Mediterranean Diet and Depression. Public Health Nutr. 2006, 9, 1104–1109. [Google Scholar] [CrossRef]
- Lang, U.E.; Beglinger, C.; Schweinfurth, N.; Walter, M.; Borgwardt, S. Nutritional Aspects of Depression. Cell. Physiol. Biochem. 2015, 37, 1029–1043. [Google Scholar] [CrossRef]
- Yin, W.; Löf, M.; Chen, R.; Hultman, C.M.; Fang, F.; Sandin, S. Mediterranean Diet and Depression: A Population-Based Cohort Study. Int. J. Behav. Nutr. Phys. Act. 2021, 18, 153. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Martínez-González, M.A.; Estruch, R.; Salas-Salvadó, J.; Corella, D.; Covas, M.I.; Arós, F.; Romaguera, D.; Gómez-Gracia, E.; Lapetra, J.; et al. Mediterranean Dietary Pattern and Depression: The PREDIMED Randomized Trial. BMC Med. 2013, 11, 208. [Google Scholar] [CrossRef]
- Bayes, J.; Schloss, J.; Sibbritt, D. A Randomised Controlled Trial Assessing the Effect of a Mediterranean Diet on the Symptoms of Depression in Young Men (the ‘AMMEND’ Study): A Study Protocol. Br. J. Nutr. 2021, 126, 730–737. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Cabrera-Suárez, B.; Molero, P.; González-Pinto, A.; Chiclana-Actis, C.; Cabrera, C.; Lahortiga-Ramos, F.; Florido-Rodríguez, M.; Vega-Pérez, P.; Vega-Pérez, R.; et al. Preventing the Recurrence of Depression with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil. The PREDI-DEP Trial: Study Protocol. BMC Psychiatry 2019, 19, 63. [Google Scholar] [CrossRef] [PubMed]
- Parletta, N.; Zarnowiecki, D.; Cho, J.; Wilson, A.; Bogomolova, S.; Villani, A.; Itsiopoulos, C.; Niyonsenga, T.; Blunden, S.; Meyer, B.; et al. A Mediterranean-Style Dietary Intervention Supplemented with Fish Oil Improves Diet Quality and Mental Health in People with Depression: A Randomized Controlled Trial (HELFIMED). Nutr. Neurosci. 2019, 22, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Opie, R.S.; O’Neil, A.; Jacka, F.N.; Pizzinga, J.; Itsiopoulos, C. A Modified Mediterranean Dietary Intervention for Adults with Major Depression: Dietary Protocol and Feasibility Data from the SMILES Trial. Nutr. Neurosci. 2018, 21, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Wade, A.T.; Davis, C.R.; Dyer, K.A.; Hodgson, J.M.; Woodman, R.J.; Keage, H.A.D.; Murphy, K.J. A Mediterranean Diet Supplemented with Dairy Foods Improves Mood and Processing Speed in an Australian Sample: Results from the MedDairy Randomized Controlled Trial. Nutr. Neurosci. 2020, 23, 646–658. [Google Scholar] [CrossRef]
- Morris, L.; Bhatnagar, D. The Mediterranean Diet. Curr. Opin. Lipidol. 2016, 27, 89–91. [Google Scholar] [CrossRef]
- Davis, C.; Bryan, J.; Hodgson, J.; Murphy, K. Definition of the Mediterranean Diet; a Literature Review. Nutrients 2015, 7, 9139–9153. [Google Scholar] [CrossRef]
- Naureen, Z.; Bonetti, G.; Medori, M.C.; Aquilanti, B.; Velluti, V.; Matera, G.; Iaconelli, A.; Bertelli, M. Foods of the Mediterranean Diet: Lacto-Fermented Food, the Food Pyramid and Food Combinations. J. Prev. Med. Hyg. 2022, 63, E28–E35. [Google Scholar] [CrossRef]
- Briguglio, M.; Dell’Osso, B.; Panzica, G.; Malgaroli, A.; Banfi, G.; Zanaboni Dina, C.; Galentino, R.; Porta, M. Dietary Neurotransmitters: A Narrative Review on Current Knowledge. Nutrients 2018, 10, 591. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, P.; Pan, D.; Zeng, X.; Guo, Y.; Zhao, G. Effect of Adzuki Bean Sprout Fermented Milk Enriched in γ-Aminobutyric Acid on Mild Depression in a Mouse Model. J. Dairy Sci. 2021, 104, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Wan-Mohtar, W.A.A.Q.I.; Sohedein, M.N.A.; Ibrahim, M.F.; Ab Kadir, S.; Suan, O.P.; Weng Loen, A.W.; Sassi, S.; Ilham, Z. Isolation, Identification, and Optimization of γ-Aminobutyric Acid (GABA)-Producing Bacillus Cereus Strain KBC from a Commercial Soy Sauce Moromi in Submerged-Liquid Fermentation. Processes 2020, 8, 652. [Google Scholar] [CrossRef]
- Linares, D.M.; O’Callaghan, T.F.; O’Connor, P.M.; Ross, R.P.; Stanton, C. Streptococcus Thermophilus APC151 Strain Is Suitable for the Manufacture of Naturally GABA-Enriched Bioactive Yogurt. Front. Microbiol. 2016, 7, 1876. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Chamignon, C.; Mhedbi-Hajri, N.; Chain, F.; Derrien, M.; Escribano-Vázquez, U.; Garault, P.; Cotillard, A.; Pham, H.P.; Chervaux, C.; et al. The Potential Probiotic Lactobacillus rhamnosus CNCM I-3690 Strain Protects the Intestinal Barrier by Stimulating Both Mucus Production and Cytoprotective Response. Sci. Rep. 2019, 9, 5398. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.-W.; Cheng, M.-Y.; Yang, X.; Lu, Y.-Y.; Yin, H.-D.; Zeng, Y.; Wang, R.-Y.; Jiang, Y.-L.; Yang, W.-T.; Wang, J.-Z.; et al. Probiotic Lactobacillus rhamnosus GG Promotes Mouse Gut Microbiota Diversity and T Cell Differentiation. Front. Microbiol. 2020, 11, 607735. [Google Scholar] [CrossRef] [PubMed]
- Janik, R.; Thomason, L.A.M.; Stanisz, A.M.; Forsythe, P.; Bienenstock, J.; Stanisz, G.J. Magnetic Resonance Spectroscopy Reveals Oral Lactobacillus Promotion of Increases in Brain GABA, N-Acetyl Aspartate and Glutamate. Neuroimage 2016, 125, 988–995. [Google Scholar] [CrossRef]
- Slykerman, R.F.; Hood, F.; Wickens, K.; Thompson, J.M.D.; Barthow, C.; Murphy, R.; Kang, J.; Rowden, J.; Stone, P.; Crane, J.; et al. Effect of Lactobacillus rhamnosus HN001 in Pregnancy on Postpartum Symptoms of Depression and Anxiety: A Randomised Double-Blind Placebo-Controlled Trial. eBioMedicine 2017, 24, 159–165. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA Production and Structure of GadB/GadC Genes in Lactobacillus and Bifidobacterium Strains from Human Microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef]
- Zhou, B.; Jin, G.; Pang, X.; Mo, Q.; Bao, J.; Liu, T.; Wu, J.; Xie, R.; Liu, X.; Liu, J.; et al. Lactobacillus rhamnosus GG Colonization in Early Life Regulates Gut-Brain Axis and Relieves Anxiety-like Behavior in Adulthood. Pharmacol. Res. 2022, 177, 106090. [Google Scholar] [CrossRef]
- Lorusso, A.; Coda, R.; Montemurro, M.; Rizzello, C.G. Use of Selected Lactic Acid Bacteria and Quinoa Flour for Manufacturing Novel Yogurt-Like Beverages. Foods 2018, 7, 51. [Google Scholar] [CrossRef]
- Hashemi Gahruie, H.; Eskandari, M.H.; Mesbahi, G.; Hanifpour, M.A. Scientific and Technical Aspects of Yogurt Fortification: A Review. Food Sci. Hum. Wellness 2015, 4, 1–8. [Google Scholar] [CrossRef]
- McKinley, M.C. The Nutrition and Health Benefits of Yoghurt. Int. J. Dairy Technol. 2005, 58, 1–12. [Google Scholar] [CrossRef]
- Lee, X.Y.; Tan, J.S.; Cheng, L.H. Gamma Aminobutyric Acid (GABA) Enrichment in Plant-Based Food—A Mini Review. Food Rev. Int. 2022, 39, 5864–5885. [Google Scholar] [CrossRef]
- Nikmaram, N.; Dar, B.; Roohinejad, S.; Koubaa, M.; Barba, F.J.; Greiner, R.; Johnson, S.K. Recent Advances in γ -Aminobutyric Acid (GABA) Properties in Pulses: An Overview. J. Sci. Food Agric. 2017, 97, 2681–2689. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Shirai, T.; Ochiai, H.; Kasao, M.; Hayakawa, K.; Kimura, M.; Sansawa, H. Blood-Pressure-Lowering Effect of a Novel Fermented Milk Containing γ-Aminobutyric Acid (GABA) in Mild Hypertensives. Eur. J. Clin. Nutr. 2003, 57, 490–495. [Google Scholar] [CrossRef]
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as Food Supplements: The Effects of GABA on Brain and Behavior. Front. Psychol. 2015, 6, 1520. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, Y.; Nagata, Y.; Hirano, S. Active Transport of Gamma-Aminobutyric Acid in Brain Cortex Slices, with Special Reference to Phosphorus-32 Turnover of Phospholipids in Cytoplasmic Particulates. Nature 1960, 186, 474–475. [Google Scholar] [CrossRef] [PubMed]
- Gelder, N.M.; Elliott, K.A.C. Disposition of γ-Aminobutyric Acid Administered to Mammals. J. Neurochem. 1958, 3, 139–143. [Google Scholar] [CrossRef]
- Takanaga, H.; Ohtsuki, S.; Hosoya, K.; Terasaki, T. GAT2/BGT-1 as a System Responsible for the Transport of Gamma-Aminobutyric Acid at the Mouse Blood-Brain Barrier. J. Cereb. Blood Flow Metab. 2001, 21, 1232–1239. [Google Scholar] [CrossRef]
- Hossain, S.J.; Hamamoto, K.; Aoshima, H.; Hara, Y. Effects of Tea Components on the Response of GABA A Receptors Expressed in Xenopus Oocytes. J. Agric. Food Chem. 2002, 50, 3954–3960. [Google Scholar] [CrossRef]
- Thanapreedawat, P.; Kobayashi, H.; Inui, N.; Sakamoto, K.; Kim, M.; Yoto, A.; Yokogoshi, H. GABA Affects Novel Object Recognition Memory and Working Memory in Rats. J. Nutr. Sci. Vitaminol. 2013, 59, 152–157. [Google Scholar] [CrossRef]
- Yoto, A.; Murao, S.; Motoki, M.; Yokoyama, Y.; Horie, N.; Takeshima, K.; Masuda, K.; Kim, M.; Yokogoshi, H. Oral Intake of γ-Aminobutyric Acid Affects Mood and Activities of Central Nervous System during Stressed Condition Induced by Mental Tasks. Amino Acids 2012, 43, 1331–1337. [Google Scholar] [CrossRef]
- Smith, A.K.; Wade, A.R.; Penkman, K.E.; Baker, D.H. Dietary Modulation of Cortical Excitation and Inhibition. J. Psychopharmacol. 2017, 31, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.J.; Cooper, M.A.; Simmons, R.D.J.; Weir, C.J.; Belelli, D. Neurosteroids: Endogenous Allosteric Modulators of GABAA Receptors. Psychoneuroendocrinology 2009, 34, S48–S58. [Google Scholar] [CrossRef] [PubMed]
- Hinton, T.; Jelinek, H.F.; Viengkhou, V.; Johnston, G.A.; Matthews, S. Effect of GABA-Fortified Oolong Tea on Reducing Stress in a University Student Cohort. Front. Nutr. 2019, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Waagepetersen, H.S.; Sonnewald, U.; Schousboe, A. The GABA Paradox. J. Neurochem. 2002, 73, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Aoshima, H.; Tenpaku, Y. Modulation of GABA Receptors Expressed in Xenopus Oocytes by 13-L-Hydoxylinoleic Acid and Food Additives. Biosci. Biotechnol. Biochem. 1997, 61, 2051–2057. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-H.; Moon, Y.-J.; Oh, C.-H. γ-Aminobutyric Acid (GABA) Content of Selected Uncooked Foods. Prev. Nutr. Food Sci. 2003, 8, 75–78. [Google Scholar] [CrossRef]
- Yamatsu, A.; Yamashita, Y.; Pandharipande, T.; Maru, I.; Kim, M. Effect of Oral γ-Aminobutyric Acid (GABA) Administration on Sleep and Its Absorption in Humans. Food Sci. Biotechnol. 2016, 25, 547–551. [Google Scholar] [CrossRef]
- Byun, J.-I.; Shin, Y.Y.; Chung, S.-E.; Shin, W.C. Safety and Efficacy of Gamma-Aminobutyric Acid from Fermented Rice Germ in Patients with Insomnia Symptoms: A Randomized, Double-Blind Trial. J. Clin. Neurol. 2018, 14, 291–295. [Google Scholar] [CrossRef]
- Kanehira, T.; Nakamura, Y.; Nakamura, K.; Horie, K.; Horie, N.; Furugori, K.; Sauchi, Y.; Yokogoshi, H. Relieving Occupational Fatigue by Consumption of a Beverage Containing γ-Amino Butyric Acid. J. Nutr. Sci. Vitaminol. 2011, 57, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Yamatsu, A.; Yamashita, Y.; Horie, K.; Takeshima, K.; Horie, N.; Masuda, K. Beneficial Action of GABA on Sleep and Frequent Night Urination in the Elderly. Jpn. Pharmacol. Ther. 2013, 41, 985–988. [Google Scholar]
- Yamastsu, A.; Yoneyama, M.; Kim, M.; Yamashita, Y.; Horie, K.; Yokogoshi, H. The Beneficial Effects of Coffee on Stress and Fatigue Can Be Enhanced by the Addition of GABA—A Randomized, Double-Blind, Placebo Controlled, Crossover-Designed Study. Jpn. Pharmacol. Ther 2015, 43, 515–519. [Google Scholar]
- Nakamura, H.; Takishima, T.; Kometani, T.; Yokogoshi, H. Psychological Stress-Reducing Effect of Chocolate Enriched with γ-Aminobutyric Acid (GABA) in Humans: Assessment of Stress Using Heart Rate Variability and Salivary Chromogranin A. Int. J. Food Sci. Nutr. 2009, 60, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Sugishita, T.; Murakami, T.; Murai, H.; Saikusa, T.; Horino, T.; Onoda, A.; Kajimoto, O.; Takahashi, R.; Takahashi, T. Effect of the Defatted Rice Germ Enriched with GABA for Sleeplessness, Depression, Autonomic Disorder by Oral Administration. Nippon. Shokuhin Kagaku Kogaku Kaishi 2000, 47, 596–603. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of Gaba (γ—Aminobutyric Acid) by Microorganisms: A Review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef]
- Reid, S.N.S.; Ryu, J.; Kim, Y.; Jeon, B.H. GABA-Enriched Fermented Laminaria Japonica Improves Cognitive Impairment and Neuroplasticity in Scopolamine- and Ethanol-Induced Dementia Model Mice. Nutr. Res. Pract. 2018, 12, 199–207. [Google Scholar] [CrossRef]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-Derived Neurotrophic Factor (BDNF) Is Required for the Enhancement of Hippocampal Neurogenesis Following Environmental Enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF Val66met Polymorphism Affects Activity-Dependent Secretion of BDNF and Human Memory and Hippocampal Function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Park, H.-J.; Lee, M.-S.; Shim, H.S.; Lee, G.-R.; Chung, S.Y.; Kang, Y.M.; Lee, B.-J.; Seo, Y.B.; Kim, K.S.; Shim, I. Fermented Saccharina Japonica (Phaeophyta) Improves Neuritogenic Activity and TMT-Induced Cognitive Deficits in Rats. ALGAE 2016, 31, 73–84. [Google Scholar] [CrossRef]
- van de Wouw, M.; Walsh, A.M.; Crispie, F.; van Leuven, L.; Lyte, J.M.; Boehme, M.; Clarke, G.; Dinan, T.G.; Cotter, P.D.; Cryan, J.F. Distinct Actions of the Fermented Beverage Kefir on Host Behaviour, Immunity and Microbiome Gut-Brain Modules in the Mouse. Microbiome 2020, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Tınok, A.A.; Karabay, A.; de Jong, J.; Balta, G.; Akyürek, E.G. Effects of Gamma-Aminobutyric Acid on Working Memory and Attention: A Randomized, Double-Blinded, Placebo-Controlled, Crossover Trial. J. Psychopharmacol. 2023, 37, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Leonte, A.; Colzato, L.S.; Steenbergen, L.; Hommel, B.; Akyürek, E.G. Supplementation of Gamma-Aminobutyric Acid (GABA) Affects Temporal, but Not Spatial Visual Attention. Brain Cogn. 2018, 120, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Rudzki, L.; Ostrowska, L.; Pawlak, D.; Małus, A.; Pawlak, K.; Waszkiewicz, N.; Szulc, A. Probiotic Lactobacillus plantarum 299v Decreases Kynurenine Concentration and Improves Cognitive Functions in Patients with Major Depression: A Double-Blind, Randomized, Placebo Controlled Study. Psychoneuroendocrinology 2019, 100, 213–222. [Google Scholar] [CrossRef]
- Schneider, E.; Doll, J.P.K.; Schweinfurth, N.; Kettelhack, C.; Schaub, A.-C.; Yamanbaeva, G.; Varghese, N.; Mählmann, L.; Brand, S.; Eckert, A.; et al. Effect of Short-Term, High-Dose Probiotic Supplementation on Cognition, Related Brain Functions and BDNF in Patients with Depression: A Secondary Analysis of a Randomized Controlled Trial. J. Psychiatry Neurosci. 2023, 48, E23–E33. [Google Scholar] [CrossRef]
- Shyamaladevi, N.; Jayakumar, A.R.; Sujatha, R.; Paul, V.; Subramanian, E.H. Evidence That Nitric Oxide Production Increases γ-Amino Butyric Acid Permeability of Blood-Brain Barrier. Brain Res. Bull. 2002, 57, 231–236. [Google Scholar] [CrossRef]
- Thorne Research, Inc. Gamma-Aminobutyric Acid (GABA), Monograph. Altern. Med. Rev. 2007, 12, 274–279. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liwinski, T.; Lang, U.E.; Brühl, A.B.; Schneider, E. Exploring the Therapeutic Potential of Gamma-Aminobutyric Acid in Stress and Depressive Disorders through the Gut–Brain Axis. Biomedicines 2023, 11, 3128. https://doi.org/10.3390/biomedicines11123128
Liwinski T, Lang UE, Brühl AB, Schneider E. Exploring the Therapeutic Potential of Gamma-Aminobutyric Acid in Stress and Depressive Disorders through the Gut–Brain Axis. Biomedicines. 2023; 11(12):3128. https://doi.org/10.3390/biomedicines11123128
Chicago/Turabian StyleLiwinski, Timur, Undine E. Lang, Annette B. Brühl, and Else Schneider. 2023. "Exploring the Therapeutic Potential of Gamma-Aminobutyric Acid in Stress and Depressive Disorders through the Gut–Brain Axis" Biomedicines 11, no. 12: 3128. https://doi.org/10.3390/biomedicines11123128