
Nerve Growth Factor, Antimicrobial Peptides and Chemotherapy: Glioblastoma Combination Therapy to Improve Their Efficacy
 ,
,
Abstract
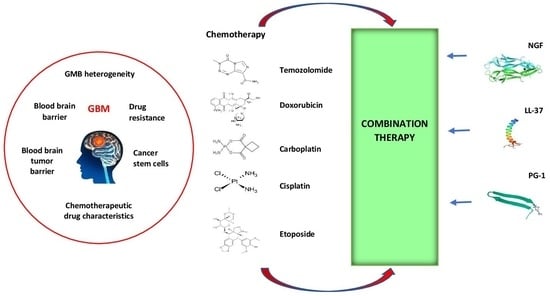
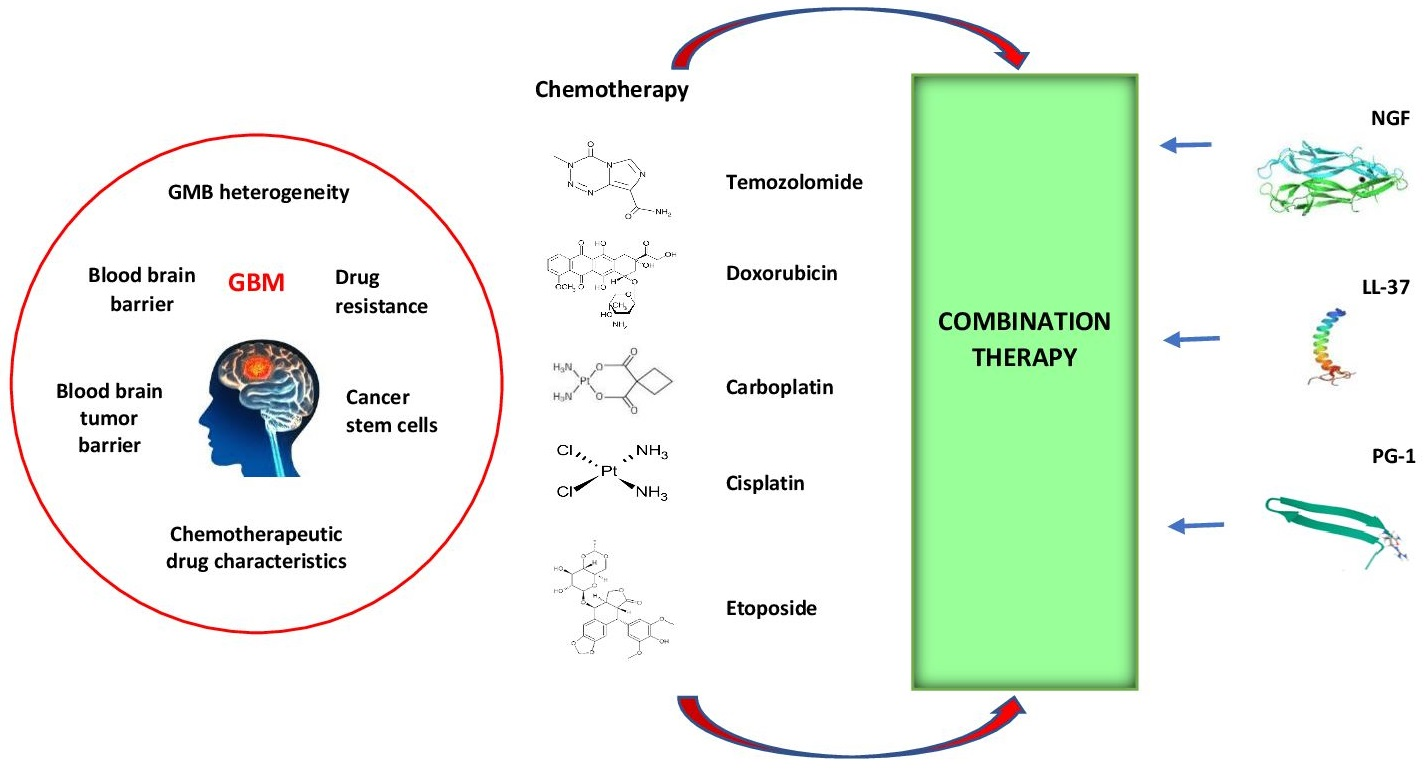
:
1. Introduction
2. Materials and Methods
2.1. Pharmacological Agents, Chemicals, Peptides, and Reagents
2.2. Cell Culture
2.3. Concentrations of Compounds
2.4. MTT Assay
2.5. Assessment of Drug-Concentration Effect and Calculation of the Combination Index
2.6. Assessment of Cell Viability with Flow Cytometry Using YO-PRO-1 and PI
2.7. Caspase Activation Analysis
2.8. Statistical Analysis
3. Results
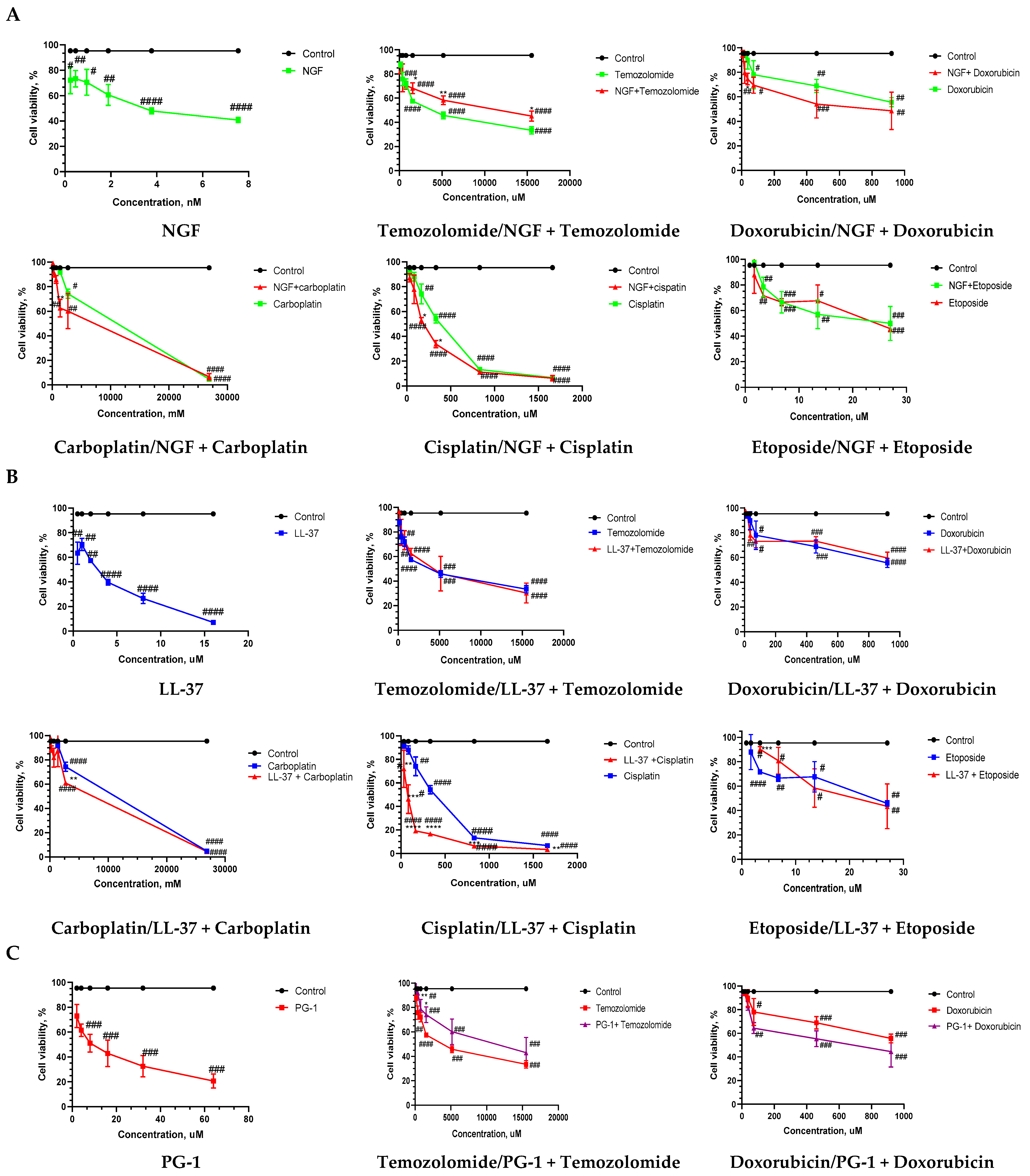
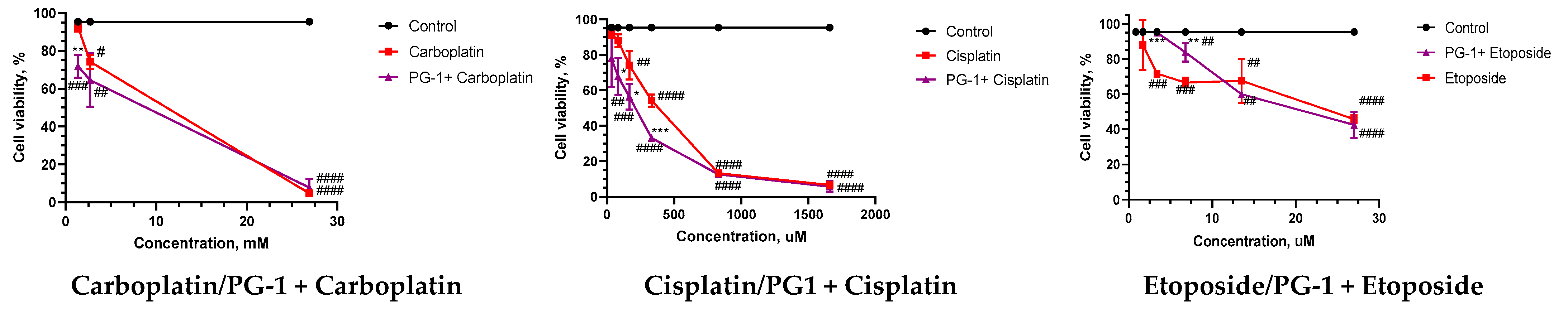
3.1. Sensitivity of U251GBM Cells to NGF, LL-37, PG-1, and Chemotherapy and IC50 Calculation
3.2. The Combination Index (CI) and Anti-Tumor Activity of Combined Treatment of NGF, LL-37, and PG-1 with Chemotherapy
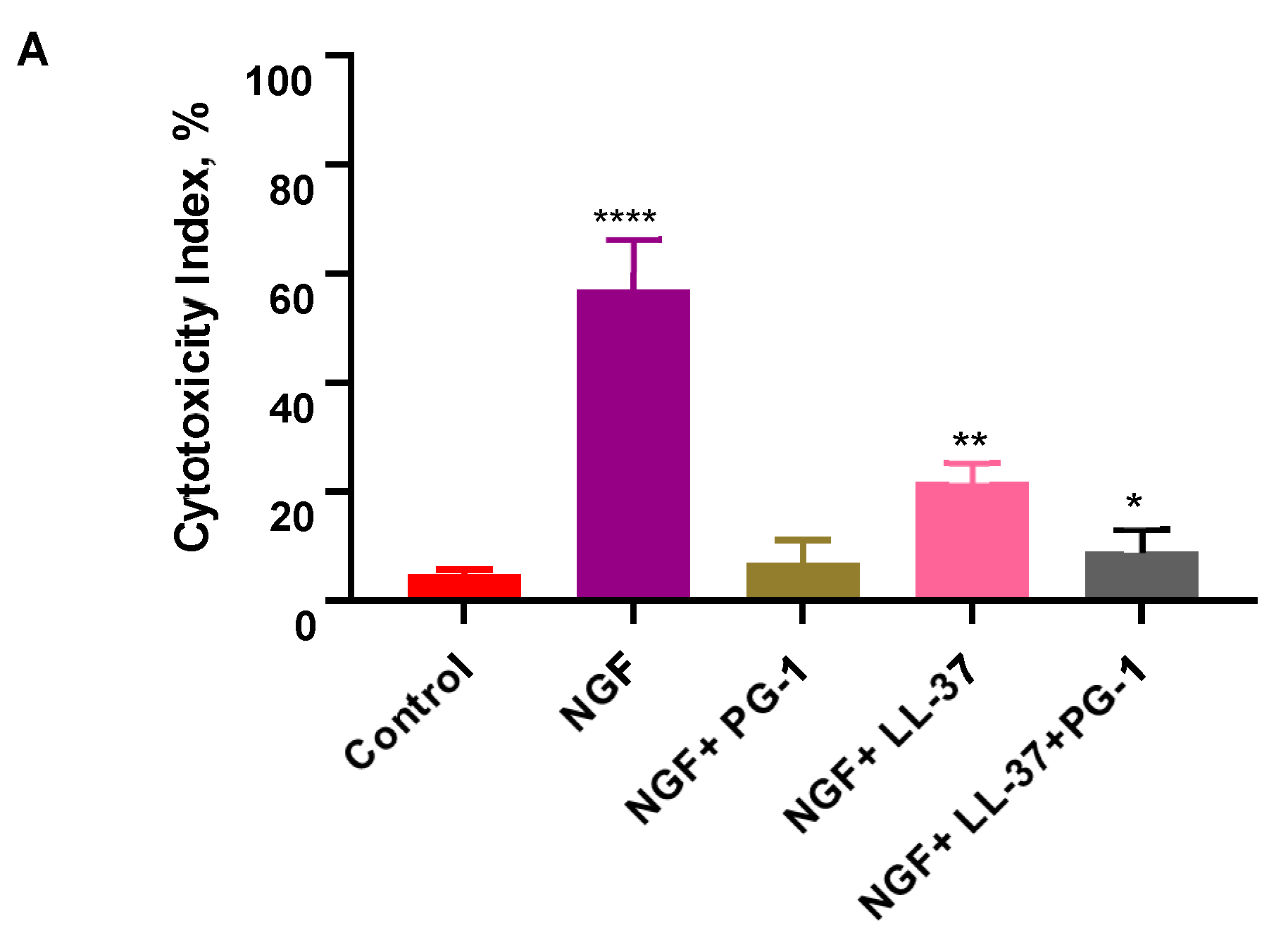
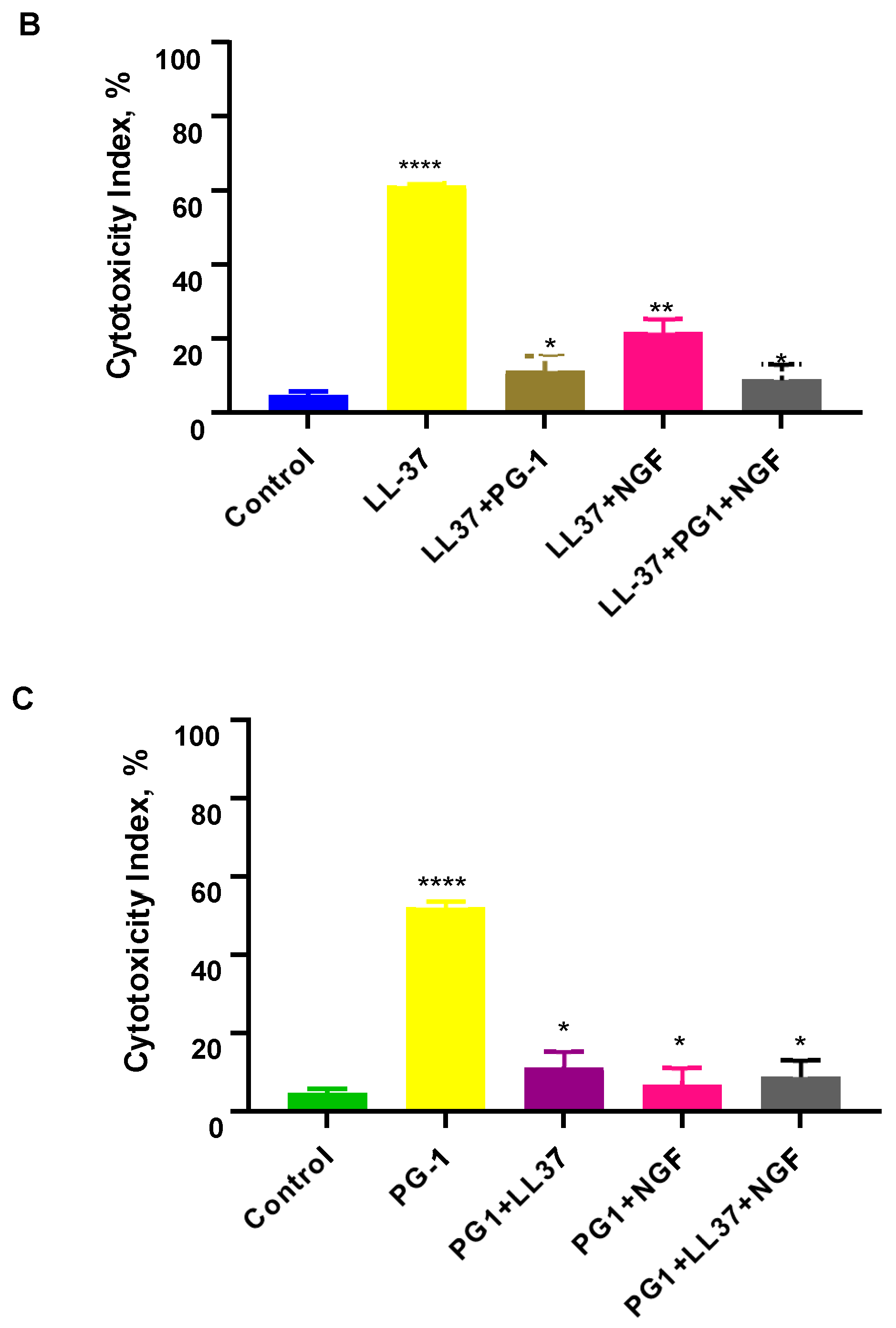
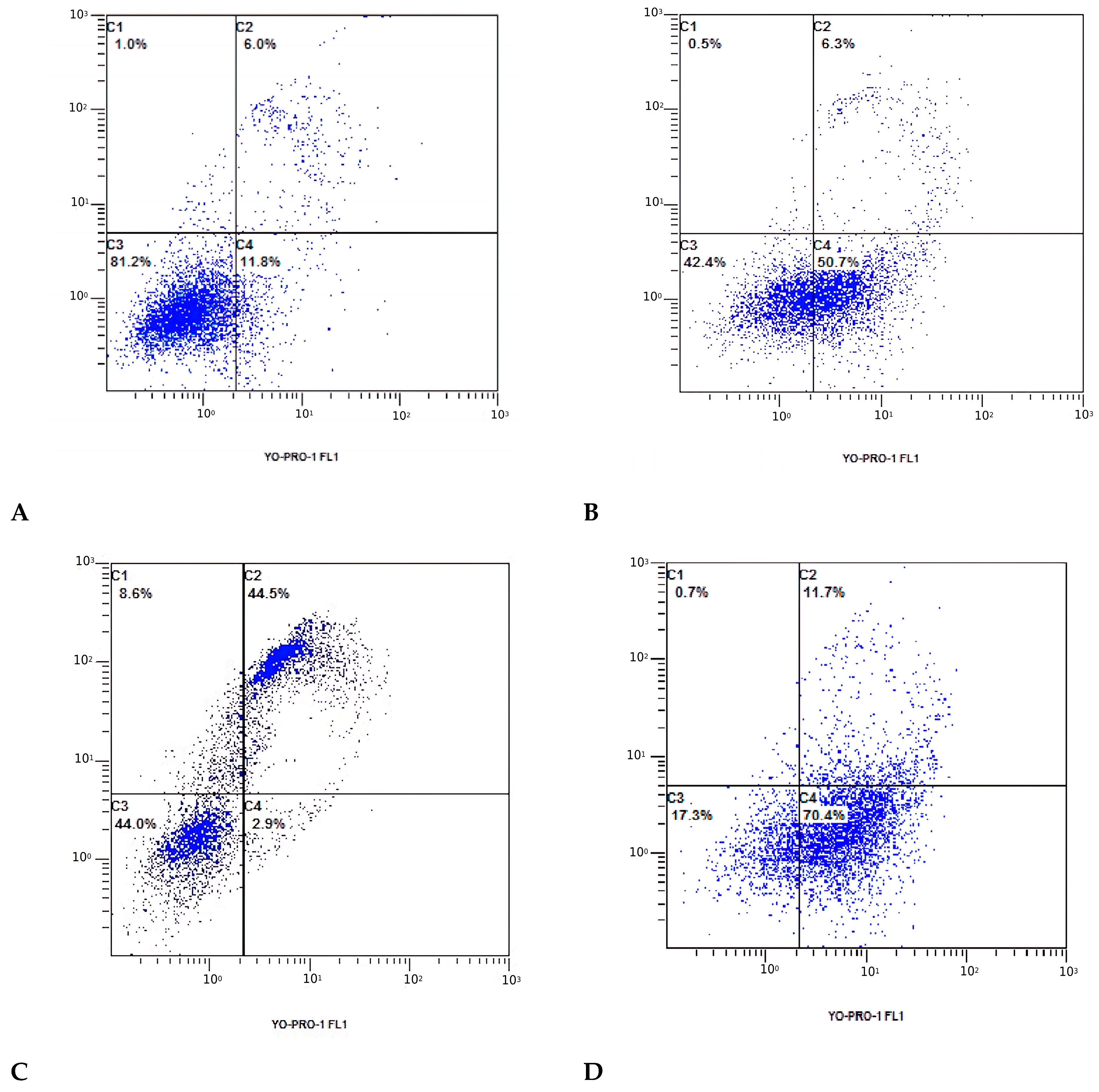
3.3. Cell Apoptosis Detected with Flow Cytometry
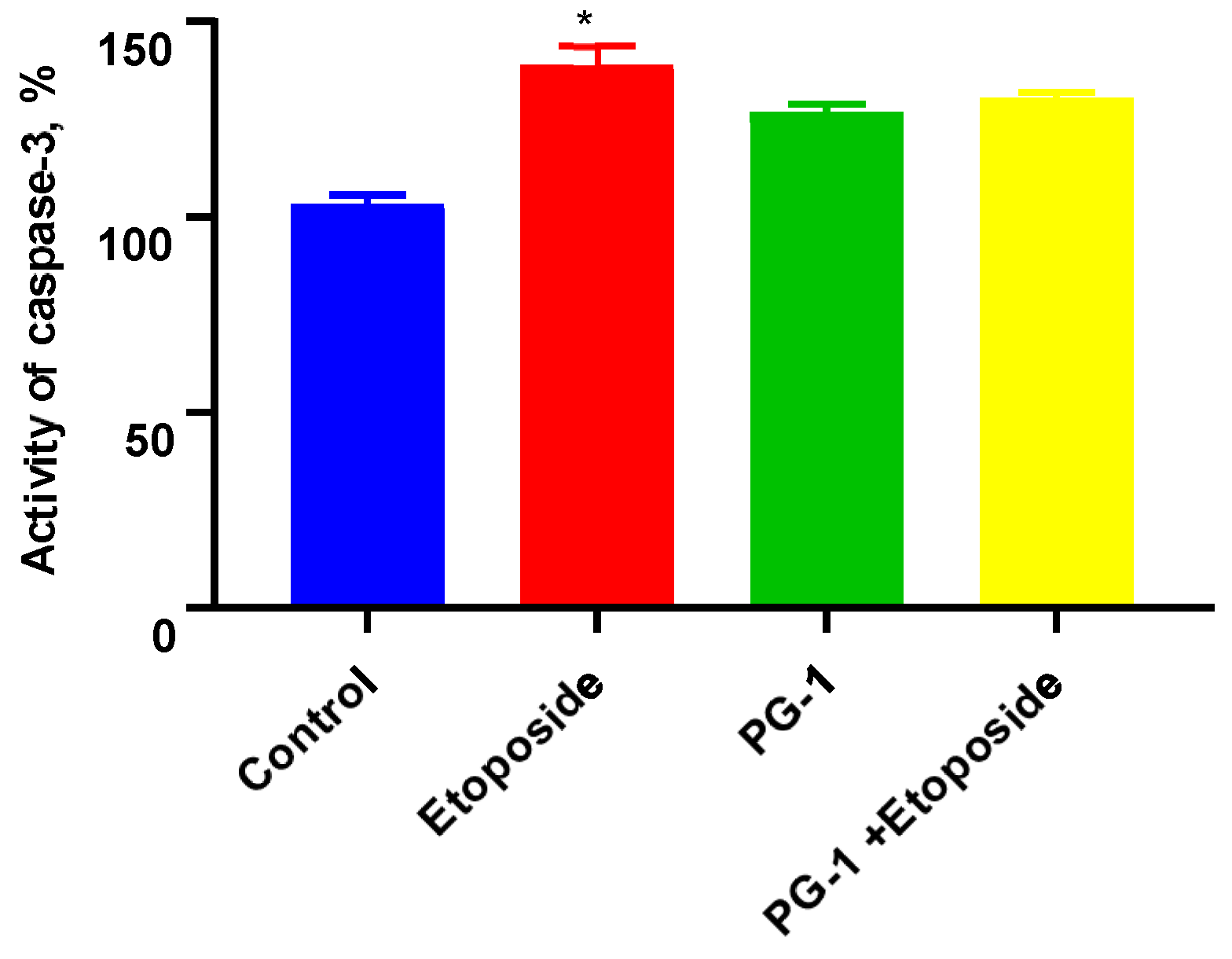
3.4. Activation of Caspase-3 on U251 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2016 Brain and Other CNS Cancer Collaborators. Global, regional, and national burden of brain and other CNS cancer, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 376–393. [Google Scholar] [CrossRef]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global cancer observatory: Cancer today. Lyon Fr. Int. Agency Res. Cancer 2018, 3, 2019. [Google Scholar]
- Nabors, L.B.; Portnow, J.; Ahluwalia, M.; Baehring, J.; Brem, H.; Brem, S.; Butowski, N.; Campian, J.L.; Clark, S.W.; Fabiano, A.J.; et al. Central Nervous System Cancers, Version 3.2020, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. JNCCN 2020, 18, 1537–1570. [Google Scholar] [CrossRef]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of Tumor-Treating Fields Plus Maintenance Temozolomide vs Maintenance Temozolomide Alone on Survival in Patients With Glioblastoma: A Randomized Clinical Trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, M.; Gan, H.; Wang, H.; Lee, J.H.; Fang, D.; Kitange, G.J.; He, L.; Hu, Z.; Parney, I.F.; et al. A novel enhancer regulates MGMT expression and promotes temozolomide resistance in glioblastoma. Nat. Commun. 2018, 9, 2949. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, J.S.; Alexander, E., 3rd; Hochberg, F.H.; Wen, P.Y.; Morris, J.H.; Schoene, W.C.; Siddon, R.L.; Morse, R.H.; Black, P.M. Clinical patterns of failure following stereotactic interstitial irradiation for malignant gliomas. Int. J. Radiat. Oncol. Biol. Phys. 1990, 19, 1455–1462. [Google Scholar] [CrossRef]
- Saunders, N.A.; Simpson, F.; Thompson, E.W.; Hill, M.M.; Endo-Munoz, L.; Leggatt, G.; Minchin, R.F.; Guminski, A. Role of intratumoural heterogeneity in cancer drug resistance: Molecular and clinical perspectives. EMBO Mol. Med. 2012, 4, 675–684. [Google Scholar] [CrossRef]
- Furnari, F.B.; Cloughesy, T.F.; Cavenee, W.K.; Mischel, P.S. Heterogeneity of epidermal growth factor receptor signalling networks in glioblastoma. Nat. Rev. Cancer 2015, 15, 302–310. [Google Scholar] [CrossRef]
- Qazi, M.A.; Vora, P.; Venugopal, C.; Sidhu, S.S.; Moffat, J.; Swanton, C.; Singh, S.K. Intratumoral heterogeneity: Pathways to treatment resistance and relapse in human glioblastoma. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2017, 28, 1448–1456. [Google Scholar] [CrossRef]
- Gupta, S.K.; Kizilbash, S.H.; Daniels, D.J.; Sarkaria, J.N. Editorial: Targeted Therapies for Glioblastoma: A Critical Appraisal. Front. Oncol. 2019, 9, 1216. [Google Scholar] [CrossRef]
- Di Tacchio, M.; Macas, J.; Weissenberger, J.; Sommer, K.; Bähr, O.; Steinbach, J.P.; Senft, C.; Seifert, V.; Glas, M.; Herrlinger, U.; et al. Tumor Vessel Normalization, Immunostimulatory Reprogramming, and Improved Survival in Glioblastoma with Combined Inhibition of PD-1, Angiopoietin-2, and VEGF. Cancer Immunol. Res. 2019, 7, 1910–1927. [Google Scholar] [CrossRef] [PubMed]
- Polivka, J.; Polivka, J.; Holubec, L.; Kubikova, T.; Priban, V.; Hes, O.; Pivovarcikova, K.; Treskova, I. Advances in Experimental Targeted Therapy and Immunotherapy for Patients with Glioblastoma Multiforme. Anticancer Res. 2017, 37, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Balandin, S.V.; Emelianova, A.A.; Kalashnikova, M.B.; Kokryakov, V.N.; Shamova, O.V.; Ovchinnikova, T.V. Molecular Mechanisms of Anticancer Action of Natural Antimicrobial Peptides. Bioorganic Chem. 2016, 42, 633–648. [Google Scholar]
- CIBA Foundation Symposium. Growth Factors in Biology and Medicine; Evered, D., Nugent, J., Whelan, J., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 0470718676/9780470718674. [Google Scholar]
- Apfel, S.C.; Arezzo, J.C.; Brownlee, M.; Federoff, H.; Kessler, J.A. Nerve growth factor administration protects against experimental diabetic sensory neuropathy. Brain Res. 1994, 634, 7–12. [Google Scholar] [CrossRef]
- Aloe, L.; Rocco, M.L.; Balzamino, B.O.; Micera, A. Nerve growth factor: Role in growth, differentiation and controlling cancer cell development. J. Exp. Clin. Cancer Res. CR 2016, 35, 116. [Google Scholar] [CrossRef]
- Verge, V.M.; Merlio, J.P.; Grondin, J.; Ernfors, P.; Persson, H.; Riopelle, R.J.; Hökfelt, T.; Richardson, P.M. Colocalization of NGF binding sites, trk mRNA, and low-affinity NGF receptor mRNA in primary sensory neurons: Responses to injury and infusion of NGF. J. Neurosci. Off. J. Soc. Neurosci. 1992, 12, 4011–4022. [Google Scholar] [CrossRef]
- Chiaretti, A.; Antonelli, A.; Genovese, O.; Fernandez, E.; Di Giuda, D.; Mariotti, P.; Riccardi, R. Intraventricular nerve growth factor infusion improves cerebral blood flow and stimulates doublecortin expression in two infants with hypoxic-ischemic brain injury. Neurol. Res. 2008, 30, 223–228. [Google Scholar] [CrossRef]
- Goretzki, P.E.; Wahl, R.A.; Becker, R.; Koller, C.; Branscheid, D.; Grussendorf, M.; Roeher, H.D. Nerve growth factor (NGF) sensitizes human medullary thyroid carcinoma (hMTC) cells for cytostatic therapy in vitro. Surgery 1987, 102, 1035–1042. [Google Scholar]
- Rakowicz-Szulczyńska, E.M.; Herlyn, M.; Koprowski, H. Nerve growth factor receptors in chromatin of melanoma cells, proliferating melanocytes, and colorectal carcinoma cells in vitro. Cancer Res. 1988, 48 Pt 1, 7200–7206. [Google Scholar]
- Rakowicz-Szulczynska, E.M.; Reddy, U.; Vorbrodt, A.; Herlyn, D.; Koprowski, H. Chromatin and cell surface receptors mediate melanoma cell growth response to nerve growth factor. Mol. Carcinog. 1991, 4, 388–396. [Google Scholar] [CrossRef]
- Revoltella, R.P.; Butler, R.H. Nerve growth factor may stimulate either division or differentiation of cloned C1300 neuroblastoma cells in serum-free cultures. J. Cell. Physiol. 1980, 104, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, Y.; Gong, A.; Wang, C.; Liang, Y.; Chenet, Y. Localization of NGF and TrkA at mitotic apparatus in human glioma cell line U251. Biochem. Biophys. Res. Commun. 2005, 337, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.W.; Friess, H.; Wang, L.; Di Mola, F.F.; Zimmermann, A.; Büchler, M.W. Down-regulation of nerve growth factor in poorly differentiated and advanced human esophageal cancer. Anticancer Res. 2000, 20, 125–132. [Google Scholar] [PubMed]
- Chiaretti, A.; Falsini, B.; Servidei, S.; Marangoni, D.; Pierri, F.; Riccardi, R. Nerve growth factor eye drop administration improves visual function in a patient with optic glioma. Neurorehabilit. Neural Repair 2011, 25, 386–390. [Google Scholar] [CrossRef]
- Falsini, B.; Chiaretti, A.; Barone, G.; Piccardi, M.; Pierri, F.; Colosimo, C.; Lazzareschi, I.; Ruggiero, A.; Parisi, V.; Fadda, A.; et al. Topical nerve growth factor as a visual rescue strategy in pediatric optic gliomas: A pilot study including electrophysiology. Neurorehabilit. Neural Repair 2011, 25, 512–520. [Google Scholar] [CrossRef]
- Falsini, B.; Chiaretti, A.; Rizzo, D.; Piccardi, M.; Ruggiero, A.; Manni, L.; Soligo, M.; Dickmann, A.; Federici, M.; Salerni, A.; et al. Nerve growth factor improves visual loss in childhood optic gliomas: A randomized, double-blind, phase II clinical trial. Brain A J. Neurol. 2016, 139 Pt 2, 404–414. [Google Scholar] [CrossRef]
- Kimura, S.; Yoshino, A.; Katayama, Y.; Watanabe, T.; Fukushima, T. Growth control of C6 glioma in vivo by nerve growth factor. J. Neuro-Oncol. 2002, 59, 199–205. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, H.; Lu, H.; Li, G.; Huang, Q. LAMP: A Database Linking Antimicrobial Peptides. PLoS ONE 2013, 8, e66557. [Google Scholar] [CrossRef]
- Do, N.; Weindl, G.; Grohmann, L.; Salwiczek, M.; Koksch, B.; Korting, H.C.; Schäfer-Korting, M. Cationic membrane-active peptides-anticancer and antifungal activity as well as penetration into human skin. Exp. Dermatol. 2014, 23, 326–331. [Google Scholar] [CrossRef]
- Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Antimicrobial peptides: Versatile biological properties. Int. J. Pept. 2013, 2013, 675391. [Google Scholar] [CrossRef]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Et Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef]
- Mader, J.S.; Hoskin, D.W. Cationic antimicrobial peptides as novel cytotoxic agents for cancer treatment. Expert Opin. Investig. Drugs 2006, 15, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Burns, J.L.; Montelaro, R.C. Engineered cationic antimicrobial peptides to overcome multidrug resistance by ESKAPE pathogens. Antimicrob. Agents Chemother. 2015, 59, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Steckbeck, J.D.; Deslouches, B.; Montelaro, R.C. Antimicrobial peptides: New drugs for bad bugs? Expert Opin. Biol. Ther. 2014, 14, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Mietzner, T.A.; Montelaro, R.C. Rational design of engineered cationic antimicrobial peptides consisting exclusively of arginine and tryptophan, and their activity against multidrug-resistant pathogens. Antimicrob. Agents Chemother. 2013, 57, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Scocchi, M.; Mardirossian, M.; Runti, G.; Benincasa, M. Non-Membrane Permeabilizing Modes of Action of Antimicrobial Peptides on Bacteria. Curr. Top. Med. Chem. 2016, 16, 76–88. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Gkeka, P.; Sarkisov, L. Interactions of phospholipid bilayers with several classes of amphiphilic alpha-helical peptides: Insights from coarse-grained molecular dynamics simulations. J. Phys. Chem. B 2010, 114, 826–839. [Google Scholar] [CrossRef]
- Wang, L.; Dong, C.; Li, X.; Han, W.; Su, X. Anticancer potential of bioactive peptides from animal sources (Review). Oncol. Rep. 2017, 38, 637–651. [Google Scholar] [CrossRef]
- Imamovic, L.; Sommer, M.O. Use of collateral sensitivity networks to design drug cycling protocols that avoid resistance development. Sci. Transl. Med. 2013, 5, 204ra132. [Google Scholar] [CrossRef]
- Cokol, M.; Chua, H.N.; Tasan, M.; Mutlu, B.; Weinstein, Z.B.; Suzuki, Y.; Nergiz, M.E.; Costanzo, M.; Baryshnikova, A.; Giaever, G.; et al. Systematic exploration of synergistic drug pairs. Mol. Syst. Biol. 2011, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- Freshney, R.I. Animal Cell Culture: A Practical Approach, 3rd ed.; Oxford University Press: London, UK, 2000. [Google Scholar]
- Zhang, S.; Xie, R.; Wan, F.; Ye, F.; Guo, D.; Lei, T. Identification of U251 glioma stem cells and their heterogeneous stem-like phenotypes. Oncol. Lett. 2013, 6, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Florento, L.; Matias, R.; Tuaño, E.; Santiago, K.; Dela Cruz, F.; Tuazon, A. Comparison of Cytotoxic Activity of Anticancer Drugs against Various Human Tumor Cell Lines Using In Vitro Cell-Based Approach. Int. J. Biomed. Sci. IJBS 2012, 8, 76–80. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Mohamadi, N.; Kazemi, S.M.; Mohammadian, M.; Milani, A.T.; Moradi, Y.; Yasemi, M.; Ebrahimifar, M.; Tabrizi, M.M.; Shahmabadi, H.E.; Khiyavi, A.A. Toxicity of Cisplatin-Loaded Poly Butyl Cyanoacrylate Nanoparticles in a Brain Cancer Cell Line: Anionic Polymerization Results. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 629–632. [Google Scholar]
- Svirnovsky, A.I. Methodological studies of drug sensitivity of leukemic cells. Probl. Health Ecol. 2011, 3, 89–91. [Google Scholar]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Mindukshev, I.; Kudryavtsev, I.; Serebriakova, M.; Trulioff, A.; Gambaryan, S.; Sudnitsyna, J.; Khmelevskoy, D.; Voitenko, N.; Avdonin, P.; Jenkins, R.; et al. Flow Cytometry and Light Scattering Technique in Evaluation of Nutraceuticals. In Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2016; pp. 319–332. [Google Scholar]
- Dubashynskaya, N.V.; Golovkin, A.S.; Kudryavtsev, I.V.; Prikhodko, S.S.; Trulioff, A.S.; Bokatyi, A.N.; Poshina, D.N.; Raik, S.V.; Skorik, Y.A. Mucoadhesive cholesterol-chitosan self-assembled particles for topical ocular delivery of dexamethasone. Int. J. Biol. Macromol. 2020, 158, 811–818. [Google Scholar] [CrossRef]
- Caspase 3 Assay Kit, Colorimetric: Technical Bulletin (2004) Sigma-Aldrich, Inc 4 p. Available online: https://www.abcam.com/products/assay-kits/caspase-3-assay-kit-colorimetric-ab39401.html (accessed on 29 October 2023).
- van Belle, G.; Fisher, L.D.; Heagerty, P.J.; Lumley, T. Biostatistics: A Methodology for the Health Sciences; Fisher, L.D., van Belle, G., Eds.; Jonh Wiley and Sons Inc.: Hoboken, NJ, USA, 2004. [Google Scholar]
- Voronina, T.A.; Guzeevatykh, L.S.; Mironov, A.N. Guidelines for Conducting Preclinical Studies of Drugs; Mulyar, A.G., Chichenkov, O.N., Eds.; Grif and K: Moscow, Russia, 2012. [Google Scholar]
- Chernov, A.N.; Tsapieva, A.; Alaverdian, D.A.; Filatenkova, T.A.; Galimova, E.S.; Suvorova, M.; Shamova, O.V.; Suvorov, A.N. In vitro evaluation of the cytotoxic effect of Streptococcus pyogenes strains, protegrin PG-1, cathelicidin LL-37, nerve growth factor and chemotherapy on the C6 glioma cell line. Molecules 2022, 27, 569. [Google Scholar] [CrossRef]
- Chernov, A.N.; Filatenkova, T.A.; Glushakov, R.I.; Buntovskaya, A.S.; Alaverdian, D.A.; Tsapieva, A.N.; Kim, A.V.; Fedorov, E.V.; Skliar, S.S.; Matsko, M.V.; et al. Anticancer effect of cathelicidin LL-37, protegrin PG-1, nerve growth factor NGF, and temozolomide: Impact on the mitochondrial metabolism, clonogenic potential, and migration of human U251 glioma cells. Molecules 2022, 27, 4988. [Google Scholar] [CrossRef]
- Giraud, S.; Loum, E.; Bessette, B.; Mathonnet, M.; Lalloué, F. P75 neurotrophin receptor is sequestered in the Golgi apparatus of the U-87 MG human glioblastoma cell line. Int. J. Oncol. 2011, 38, 391–399. [Google Scholar] [PubMed]
- Oelmann, E.; Sreter, L.; Schuller, I.; Serve, H.; Koenigsmann, M.; Wiedenmann, B.; Oberberg, D.; Reufi, B.; Thiel, E.; Berdel, W.E. Nerve growth factor stimulates clonal growth of human lung cancer cell lines and a human glioblastoma cell line expressing high-affinity nerve growth factor binding sites involving tyrosine kinase signaling. Cancer Res. 1995, 55, 2212–2219. [Google Scholar] [PubMed]
- Singer, H.S.; Hansen, B.; Martinie, D.; Karp, C.L. Mitogenesis in glioblastoma multiforme cell lines: A role for NGF and its TrkA receptors. J. Neuro-Oncol. 1999, 45, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Wang, H.; Wang, F.J.; Xi, Y.F.; Chen, L.H. Expression of nerve growth factor and brain-derived neurotrophic factor in astrocytomas. Oncol. Lett. 2018, 15, 533–537. [Google Scholar] [CrossRef]
- Yang, Y.; Peng, H.; Chen, C.-B.; Yu, H. Expression and biological significance of nerve growth factor and its receptor in human glioma cell line U251. Chin. J. Clin. Rehabil. 2006, 10, 94–96. [Google Scholar]
- Johnston, A.L.; Lun, X.; Rahn, J.J.; Liacini, A.; Wang, L.; Hamilton, M.G.; Parney, I.F.; Hempstead, B.L.; Robbins, S.M.; Forsyth, P.A.; et al. The p75 neurotrophin receptor is a central regulator of glioma invasion. PLoS Biol. 2007, 5, e212. [Google Scholar] [CrossRef]
- Hempstead, B.L.; Martin-Zanca, D.; Kaplan, D.R.; Parada, L.F.; Chao, M.V. High-affinity NGF binding requires coexpression of the trk proto-oncogene and the low-affinity NGF receptor. Nature 1991, 350, 678–683. [Google Scholar] [CrossRef]
- Piktel, E.; Niemirowicz, K.; Wnorowska, U.; Wątek, M.; Wollny, T.; Głuszek, K.; Góźdź, S.; Levental, I.; Bucki, R. The Role of Cathelicidin LL-37 in Cancer Development. Arch. Immunol. Et Ther. Exp. 2016, 64, 33–46. [Google Scholar] [CrossRef]
- Chen, X.; Zou, X.; Qi, G.; Tang, Y.; Guo, Y.; Si, J.; Liang, L. Roles and Mechanisms of Human Cathelicidin LL-37 in Cancer. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 47, 1060–1073. [Google Scholar] [CrossRef]
- An, L.L.; Ma, X.-T.; Yang, Y.-H.; Lin, Y.-M.; Song, Y.-H.; Wu, K.-F. Marked reduction of LL-37/hCAP-18, an antimicrobial peptide, in patients with acute myeloid leukemia. Int. J. Hematol. 2005, 81, 45–47. [Google Scholar] [CrossRef]
- Yang, Y.H.; Zheng, G.-G.; Li, G.; Zhang, B.; Song, Y.-H.; Wu, K.-F. Expression of LL-37/hCAP-18 gene in human leukemia cells. Leuk. Res. 2003, 27, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.K.; Yiu Sung, J.J.; To, K.F.; Yu, L.; Li, H.T.; Li, Z.J.; Chu, K.M.; Yu, J.; Cho, C.H. The host defense peptide LL-37 activates the tumor-suppressing bone morphogenetic protein signaling via inhibition of proteasome in gastric cancer cells. J. Cell. Physiol. 2010, 223, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Heilborn, J.D.; Nilsson, M.F.; Chamorro Jimenez, C.I.; Sandstedt, B.; Borregaard, N.; Tham, E.; Sørensen, O.E.; Weber, G.; Ståhle, M. Antimicrobial protein hCAP18/LL-37 is highly expressed in breast cancer and is a putative growth factor for epithelial cells. Int. J. Cancer 2005, 114, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Coffelt, S.B.; Waterman, R.S.; Florez, L.; Höner zu Bentrup, K.; Zwezdaryk, K.J.; Tomchuck, S.L.; LaMarca, H.L.; Danka, E.S.; Morris, C.A.; Scandurro, A.B. Ovarian cancers overexpress the antimicrobial protein hCAP-18 and its derivative LL-37 increases ovarian cancer cell proliferation and invasion. Int. J. Cancer 2008, 122, 1030–1039. [Google Scholar] [CrossRef]
- von Haussen, J.; Koczulla, R.; Shaykhiev, R.; Herr, C.; Pinkenburg, O.; Reimer, D.; Wiewrodt, R.; Biesterfeld, S.; Aigner, A.; Czubayko, F.; et al. The host defence peptide LL-37/hCAP-18 is a growth factor for lung cancer cells. Lung Cancer 2008, 59, 12–23. [Google Scholar] [CrossRef]
- Ren, S.X.; Cheng, A.S.; To, K.F.; Tong, J.H.; Li, M.S.; Shen, J.; Wong, C.C.; Zhang, L.; Chan, R.L.; Wang, X.J.; et al. Host immune defense peptide LL-37 activates caspase-independent apoptosis and suppresses colon cancer. Cancer Res. 2012, 72, 6512–6523. [Google Scholar] [CrossRef]
- Colle, J.H.; Périchon, B.; Garcia, A. Antitumor and antibacterial properties of virally encoded cationic sequences. Biol. Targets Ther. 2019, 13, 117–126. [Google Scholar] [CrossRef]
- Coelho Neto, G.T.; de Lima, T.M.; Barbeiro, H.V.; Chammas, R.; César Machado, M.C.; da Silva, F.P. Cathelicidin LL-37 Promotes or Inhibits Cancer Cell Stemness Depending on the Tumor Origin. Oncomedicine 2016, 1, 14–17. [Google Scholar] [CrossRef]
- Bolintineanu, D.; Hazrati, E.; Davis, H.T.; Lehrer, R.I.; Kaznessis, Y.N. Antimicrobial mechanism of pore-forming protegrin peptides: 100 pores to kill E. coli. Peptides 2010, 31, 1–8. [Google Scholar] [CrossRef]
- Edwards, I.A.; Elliott, A.G.; Kavanagh, A.M.; Zuegg, J.; Blaskovich, M.A.T.; Cooper, M.A. Contribution of Amphipathicity and Hydrophobicity to the Antimicrobial Activity and Cytotoxicity of β-Hairpin Peptides. ACS Infect. Dis. 2016, 2, 442–450. [Google Scholar] [CrossRef]
- Drin, G.; Temsamani, J. Translocation of protegrin I through phospholipid membranes: Role of peptide folding. Biochim. Et Biophys. Acta 2002, 1559, 160–170. [Google Scholar] [CrossRef]
- Lipkin, R.B.; Lazaridis, T. Implicit Membrane Investigation of the Stability of Antimicrobial Peptide β-Barrels and Arcs. J. Membr. Biol. 2015, 248, 469–486. [Google Scholar] [CrossRef]
- Lam, K.L.; Wang, H.; Siaw, T.A.; Chapman, M.R.; Waring, A.J.; Kindt, J.T.; Lee, K.Y. Mechanism of structural transformations induced by antimicrobial peptides in lipid membranes. Biochim. Et Biophys. Acta 2012, 1818, 194–204. [Google Scholar] [CrossRef]
- Paredes-Gamero, E.J.; Martins, M.N.; Cappabianco, F.A.; Ide, J.S.; Miranda, A. Characterization of dual effects induced by antimicrobial peptides: Regulated cell death or membrane disruption. Biochim. Et Biophys. Acta 2012, 1820, 1062–1072. [Google Scholar] [CrossRef]
- Soletti, R.C.; del Barrio, L.; Daffre, S.; Miranda, A.; Borges, H.L.; Moura-Neto, V.; Lopez, M.G.; Gabilan, N.H. Peptide gomesin triggers cell death through L-type channel calcium influx, MAPK/ERK, PKC and PI3K signaling and generation of reactive oxygen species. Chem.-Biol. Interact. 2010, 186, 135–143. [Google Scholar] [CrossRef]
- Cruz-Chamorro, L.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. In vitro biological activities of magainin alone or in combination with nisin. Peptides 2006, 27, 1201–1209. [Google Scholar] [CrossRef]
- Soundrarajan, N.; Park, S.; Van Chanh, Q.L.; Cho, H.-S.; Raghunathan, G.; Ahn, B.; Song, H.; Kim, J.-H.; Park, C. Protegrin-1 cytotoxicity towards mammalian cells positively correlates with the magnitude of conformational changes of the unfolded form upon cell interaction. Sci. Rep. 2019, 9, 11569. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | N | Monotherapy, IC50 µM | NGF + Chemotherapy, IC50 µM | LL37 + Chemotherapy, IC50 µM | PG1 + Chemotherapy, IC50 µM |
|---|---|---|---|---|---|
| MTT test | |||||
| Doxorubicin | 3 | 1554.98 ± 207.5 | 600.5 ± 55.75 | 5265.5 ± 1031.0 | 730.9 ± 12.43 |
| Carboplatin | 3 | 3652.9 ± 670.8 | 2880.6 ± 275.5 | 3513.3 ± 493.0 | 2938.9 ± 529.7 |
| Temozolomide | 3 | 1725.7 ± 494.0 | 16,804.0 ± 937.4 | 4007.0 ± 365.5 | 9761.7 ± 997.0 |
| Cisplatin | 3 | 371.5 ± 23.50 | 207.4 ± 8.477 | 869 ± 107.5 | 237.9 ± 29.83 |
| Etoposide | 3 | 25.90 ± 0.61 | 13.7 ± 0.1129 | 17.9 ± 3.03 | 17.0 ± 1.80 |
| NGF (nM) | 3 | 2.14 ± 5.0 | |||
| LL-37 | 3 | 3.1 ± 0.4063 | |||
| PG-1 | 3 | 26.1 ± 7.6 |
| Compounds | PG1 + Chemotherapy | LL37 + Chemotherapy | NGF + Chemotherapy |
|---|---|---|---|
| MTT test | |||
| Doxorubicin | 0.94 additivity | 188.6 very strong antagonism | 2.42 antagonism |
| Carboplatin | 1.46 antagonism | 4.31 strong antagonism | 2.69 antagonism |
| Temozolomide | 6.04 strong antagonism | 8.95 strong antagonism | 9.73 strong antagonism |
| Cisplatin | 1.38 antagonism | 2.67 antagonism | 2.78 antagonism |
| Etoposide | 0.65 synergy | 1.85 antagonism | 2.34 antagonism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernov, A.; Kudryavtsev, I.; Komlev, A.; Alaverdian, D.; Tsapieva, A.; Galimova, E.; Shamova, O. Nerve Growth Factor, Antimicrobial Peptides and Chemotherapy: Glioblastoma Combination Therapy to Improve Their Efficacy. Biomedicines 2023, 11, 3009. https://doi.org/10.3390/biomedicines11113009
Chernov A, Kudryavtsev I, Komlev A, Alaverdian D, Tsapieva A, Galimova E, Shamova O. Nerve Growth Factor, Antimicrobial Peptides and Chemotherapy: Glioblastoma Combination Therapy to Improve Their Efficacy. Biomedicines. 2023; 11(11):3009. https://doi.org/10.3390/biomedicines11113009
Chicago/Turabian StyleChernov, Alexandr, Igor Kudryavtsev, Aleksei Komlev, Diana Alaverdian, Anna Tsapieva, Elvira Galimova, and Olga Shamova. 2023. "Nerve Growth Factor, Antimicrobial Peptides and Chemotherapy: Glioblastoma Combination Therapy to Improve Their Efficacy" Biomedicines 11, no. 11: 3009. https://doi.org/10.3390/biomedicines11113009