
High-Altitude Hypoxia Induces Excessive Erythrocytosis in Mice via Upregulation of the Intestinal HIF2a/Iron-Metabolism Pathway
1
Research Center for High Altitude Medicine, Qinghai University, Xining 810001, China
2
Key Laboratory of High-Altitude Medicine, Ministry of Education, Xining 810001, China
3
Key Laboratory of Application and Foundation for High Altitude Medicine Research in Qinghai Province, Xining 810001, China
*
Author to whom correspondence should be addressed.
Biomedicines 2023, 11(11), 2992; https://doi.org/10.3390/biomedicines11112992
Submission received: 5 October 2023
/
Revised: 2 November 2023
/
Accepted: 3 November 2023
/
Published: 7 November 2023
(This article belongs to the Special Issue Molecular Mechanisms of Normal and Malignant Hematopoiesis)
Abstract
:Excessive erythrocytosis (EE) is a preclinical form of chronic mountain sickness (CMS). The dysregulation of iron metabolism in high-altitude hypoxia may induce EE. The intestinal hypoxia-inducible factor 2 alpha (HIF2a) regulates the genes involved in iron metabolism. Considering these findings, we aimed to investigate the function and mechanism of intestinal HIF2α and the iron metabolism pathway in high-altitude EE mice. C57BL/6J mice were randomized into four groups: the low-altitude group, the high-altitude group, the high-altitude + HIF2α inhibitor group, and the high-altitude + vehicle group. In-vitro experiments were performed using the human intestinal cell line HCT116 cultured under hypoxic conditions for 24 h. Results showed that high-altitude hypoxia significantly increased the expression of intestinal HIF2α and iron metabolism-related genes, including Dmt1, Dcytb, Fpn, Tfrc, and Fth in EE mice. Genetic blockade of the intestinal HIF2α-iron metabolism pathway decreased iron availability in HCT116 cells during hypoxia. The HIF2α inhibitor PT2385 suppressed intestinal HIF2α expression, decreased iron hypermetabolism, and reduced excessive erythrocytosis in mice. These data support the hypothesis that exposure to high-altitude hypoxia can lead to iron hypermetabolism by activating intestinal HIF2α transcriptional regulation, and reduced iron availability improves EE by inhibiting intestinal HIF2α signaling.
1. Introduction
High-altitude hypoxia can induce excessive erythrocytosis (EE) [1,2]. EE is thought to be a sign of inadequate high-altitude acclimatization [3,4,5], and is one of the critical traits of chronic mountain sickness (CMS) or Monge’s disease in high-altitude populations worldwide [6,7,8]. This excessive pathobiological response has deleterious effects; it results in high hemoglobin (Hb) levels, causing severe hypoxemia and clinical symptoms such as headaches, breathlessness, fatigue, and sleep disorders [9,10,11]. Moreover, the exacerbated erythropoietic response has been suggested to be caused by the high-altitude hypoxia-induced systemic physiological changes in respiratory, cardiovascular, and hormonal responses [12,13,14]. However, the pathophysiological mechanism that results in EE remains under debate.
EE is an uncommon disorder that is marked by an abnormally increased red cell mass and abnormal changes in the body’s physiology. These changes affect the balance between iron availability and the physiology of hypoxic responses, which are important for maintaining oxygen homeostasis [15]. Hypoxia and iron are functionally related; hypoxia stimulates erythropoiesis, which increases the body’s ability to transfer oxygen, and the boost in red blood cells requires a large amount of extra iron, which must be obtained through dietary iron absorption as well as mobilization from iron stores [16]. Over the past decade, research has revealed that the liver-derived hormone hepcidin regulates the major organs of iron metabolism, and intestinal iron absorption is required to sustain systemic iron homeostasis [17]. However, a definite physiological connection between EE and iron metabolism remains unclear.
Hypoxia-inducible factors (HIFs) are transcription factors that bind to specific DNA regions known as hypoxia responsive elements (HREs) to initiate homeostatic gene transcription involved in iron metabolism and oxygen availability [18]. HIFs regulate many genes that are involved in erythropoiesis and iron metabolism, which are required for tissue oxygen delivery [19]. HIF2α activity increases with low oxygen levels in small intestinal enterocytes, activating dietary iron absorption and iron release into the circulation [20]. HIF2α plays a key role in regulating cellular iron and oxygen levels, which activates iron transporters at the apical side of the enterocyte and iron exporters at the basolateral side, releasing iron into the circulation and maintaining systemic iron levels [21,22]. HIF2α also regulates dietary iron hyperabsorption, which leads to disorders of systemic iron overload, such as -thalassemia illness [23,24]. However, the precise molecular cues by which intestinal HIF2α responds to both erythropoietic requirements and systemic iron metabolism during hypoxia are unknown. Furthermore, it is unknown whether there is a coordinated molecular integration of the intestinal HIF2α signaling pathway for EE via the dysregulation of iron metabolism caused by high-altitude hypoxia.
In this study, we hypothesized that high-altitude hypoxia activates intestinal HIF2α signaling, thereby increasing iron availability, and inducing EE in high-altitude mice. We examined whether pharmacological or genetic blockade of the intestinal HIF2α–iron gene pathway may alleviate iron hypermetabolism in vivo and in vitro and ameliorate EE.
2. Materials and Methods
2.1. Animals and Treatments
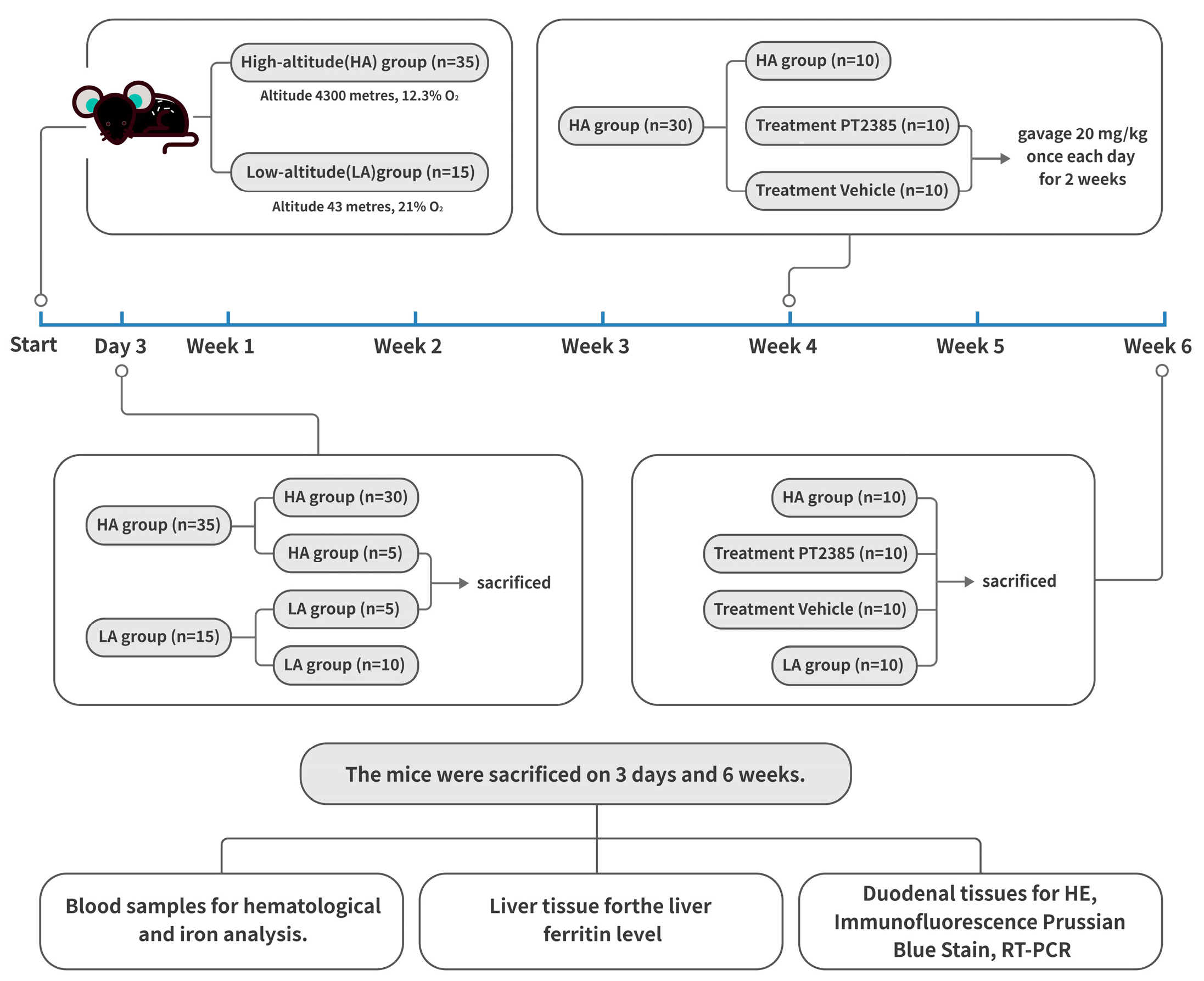
All mice-related operations described in this study were authorized by the experimental committee of Qinghai University and followed the Chinese Ministry of Health’s animal management policies. The mice used in our experiments were all 6- to 8-week-old male C57BL/6J mice obtained from the Vital River Laboratory Animal Technology Company (Beijing, China). The 50 mice were randomized into four groups. For the low-altitude (LA) group (n = 15), mice were housed at an elevation of 43 m and 21% O2 (Beijing, China) for six weeks. The high-altitude mice were housed at an elevation of 4300 m, 12.3% O2 (Maduo, China) for six weeks. The high-altitude hypoxia mice were divided into three groups: the high-altitude (HA) group (n = 15), the PT2385 treatment group (n = 10), and the blank drug control (vehicle) group (n = 10). For the HIF2α inhibitor studies, mice were administered and oral gavage at 20 mg/kg of body weight of either PT2385 (HY12867 MedChem Express, South Brunswick Township, NJ, USA) suspended in saline with 10% DMSO, 40% PEG300, 5% Tween 80, and 45% saline, or a vehicle (10% DMSO, 40% PEG300, 5% Tween 80, and 45% saline) as previously described [25]. Before the treatments, there were no differences in body weight between the mice. All mice were given standard chow and were exposed to a 12-h light-dark cycle. The mouse experimental design is shown in Figure 1.
2.2. Cell Culture and Transfection
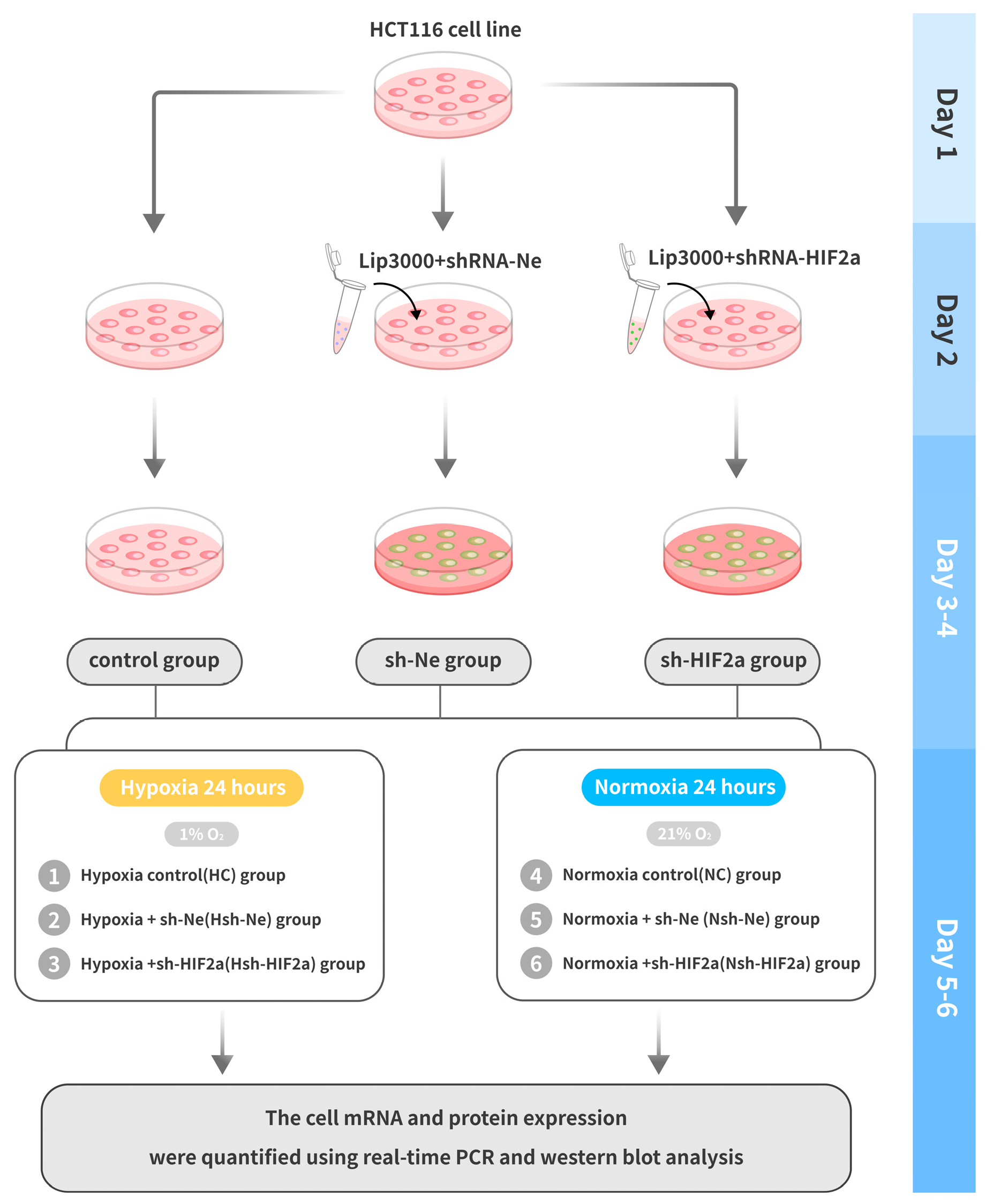
The human intestinal cell line (HCT116) was obtained from the National Infrastructure of Cell Lines (Beijing, China). All cells were maintained in IMDM (Thermo Fisher Scientific, Waltham, MA, USA) with 10% fetal bovine serum (GIBCO, Thermo Fisher Scientific) and 1% penicillin/streptomycin (Solarbio, Beijing, China) in 5% CO2 and 21% O2 at 37 °C. The short hairpin RNAs targeting human HIF2α (sh-HIF2a) were designed and produced by Genechem (Shanghai, China). The HIF2a-RNAi targeting sequences were cgACCTGAAGATTGAAGTGAT (only the sense strand is shown). The LipofectamineTM 3000 transfection reagent (L3000001 Thermo Fisher Scientific) was used to transfect sh-HIF2a into HCT116 cells. The shRNA negative control (sh-Ne) was used and delivered into cells using the same conditions. After successful cell transfection, the cells were inoculated at a density of 1 × 106/mL (2 mL/well) and cultured in 6-well plates. A total of five passages were performed, and hypoxic and normoxic control studies were performed for each passage. To perform hypoxic studies, cells were grown for 24 h at 37 °C in 1% O2 and 5% CO2 with balanced N2 [26].The experiment cells were divided into six groups: 1. Hypoxia control (HC) group, cells were grown with 1% O2; 2. Hypoxia + sh-Ne (Hsh-Ne) group, cells were transfected with negative control shRNA; 3. Hypoxia +sh-HIF2a (Hsh-HIF2a) group, cells were transfected with shRNA HIF2α; 4. Normoxia control (NC) group, normal oxygen partial pressure (21% O2) treatment; 5. Normoxia + sh-Ne (Nsh-Ne) group; 6. Normoxia + sh-HIF2a (Nsh-HIF2a) group. The successfully generated HCT118 cell knockdown clone’s mRNA and protein expression were quantified using real-time PCR and western blot analysis, respectively. The cell experimental design is shown in Figure 2.
2.3. Hematological and Iron Analysis
Mice were anaesthetized by injecting urethane (100 mg/kg) into their abdomens. Blood samples were collected for hematological and iron analysis by the retroorbital venous plexus exsanguination. Hemoglobin (Hb), hematocrit (Hct), and red blood counts (RBC) were analyzed using a Mindray Biomedical Electronics BC-5000Vet blood cell analyzer (Shenzhen, China). A Total Iron-Binding Capacity (TIBC) and a Serum Iron Assay Kit (ab239715; Abcam, Cambridge, UK) were used to measure serum iron and serum transferrin saturation (TS). The liver ferritin level was determined via a mouse ferritin enzyme-linked immunosorbent assay kit (E-EL-M0491c; Elabscience, Wuhan, China).
2.4. Histology, Immunofluorescence Staining, and Tissue Iron Staining
Fresh duodenal tissues were preserved in 4% paraformaldehyde and encapsulated in paraffin. Hematoxylin-Eosin (HE) Stain Kit was used on duodenal section tissues (G1120; Solarbio). After xylene deparaffinization (3 times, 5min for each), ethanol was used to clean the sections of tissues (wash for 3 min each with 100% ethanol, 95% ethanol, 80% ethanol, and 75% ethanol). Sections (5 µm) were stained with hematoxylin solution for 5 min, rinsed with water, stained for 5 min with eosin solution, and then rinsed with water again. For immunofluorescence staining, duodenal frozen tissues were sectioned (5 µm) and fixed for 15 min in 4% paraformaldehyde. Sections were blocked in a 10% goat serum for 60 min at room temperature before being probed with the polyclonal rabbit anti-HIF2α (1:100 ab199; Abcam) overnight at 4 °C and subsequently with rhodamine-labeled goat anti-rabbit IgG (1:100, ZF-0316; ZSGB-BIO, Beijing, China) for 120 min. After that, a counterstain with DAPI for 5 min. Tissue iron staining was performed with a Prussian Blue Stain (ab150674; Abcam). After deparaffinization in xylene, slides were incubated in a working Iron Stain solution for 3 min, rinsed in water, stained in a nuclear fast red solution for 5 min, rinsed 4 times in water, dehydrated in 95% alcohol, and mounted as directed by the manufacturer.
2.5. Real-Time Quantitative PCR
Mouse duodenal tissues and human intestinal cell lines were used to extract total RNA using an RNA Easy Fast Tissue/Cell Kit (DP451; TIANGEN Biotech Co., Beijing, China). The cDNA was constructed utilizing the FastKing gDNA Dispelling RT SuperMix (KR118; TIANGEN) and the QuantStudio5 PCR System was used to amplify genes. The cycling conditions used a 20 μL total reaction volume, and the qPCR process was as follows: pre-amplification at 95 °C for 15 min, followed by 40 cycles of amplification at 95 °C for 10 s, and elongation at 60 °C for 32 s. The fold-change in the genes was measured using the ∆∆Ct method with β-actin as the housekeeping gene. The primers are listed in Table 1.
2.6. Western Blot Analysis
RIPA buffer was applied to create whole-cell lysates from human intestinal cell lines (R0010; Solarbio). Total protein content was measured by Pierce™ BCA Protein Assay Kit (23227, Thermo Fisher Scientific). The protein lysates containing 30–40 µg of per well were loaded onto 10% SDS-PAGE, and the separated protein bands were transferred onto a 0.22-µm PVDF membrane and blocked with 5% BSA Blocker solution for 2 h at room temperature. The primary antibodies targeted HIF2α (1:500, ab199; Abcam), FTH (1:1000, ab183781; Abcam), FPN (1:1000, A14884; ABclonal, Wuhan, China), DMT1 (1:1000, ab262715; Abcam), DCYTB (1:1000, ab66048; Abcam), and ACTIN (1:1000, ab8226; Abcam). The secondary antibodies were anti-Rabbit/Mouse from ABclonal. Proteins on the blots were detected using chemiluminescence. ImageJ (version 1.49, Bethesda, MD, USA) was used to calculate protein levels.
2.7. Statistical Analysis
Statistical tests and graphs were performed using SPSS 27.0 (IBM Corp., Armonk, NY, USA) and GraphPad Prism 8.0 (GraphPad Software LLC, San Diego, CA, USA). Data are expressed as mean ± standard deviation. The two-tailed unpaired t-test was used for comparisons between two groups, whereas a one-way analysis of variance (ANOVA) was used to conduct multi-group analyses. Statistical significance is indicated as * p < 0.05.
3. Results
3.1. EE in High-Altitude Hypoxia Mice Is Accompanied by Enhanced Iron Availability
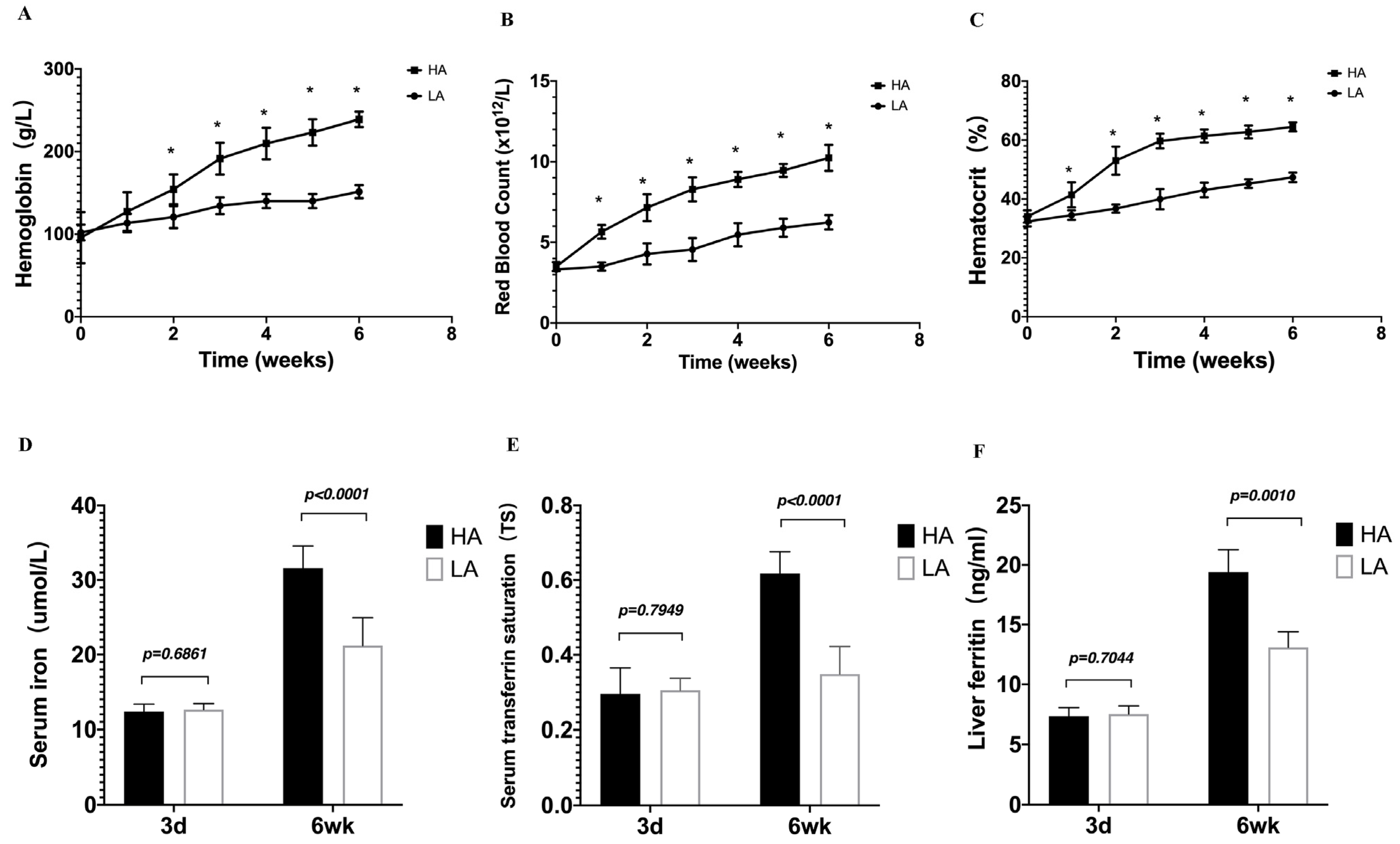
To understand the physiological connection between EE and iron availability during the high-altitude hypoxia, the mice were observed for 6 weeks, and no deaths occurred during the high-altitude hypoxia exposure period. Hb levels, RBC counts, and Hct in HA mice were notably higher than those in LA mice in a time-dependent manner (Figure 3A–C). Serum iron, serum TS, and liver ferritin reflect the capacity for iron absorption, transportation, and storage. We detected serum iron, TS, and liver ferritin increases at 6 weeks after exposure to high-altitude hypoxia. Compared to the LA group, the iron status of the HA group was significantly increased (Figure 3D–F). These findings suggest that high-altitude hypoxia mice with EE have increased iron availability.
3.2. Intestinal HIF2α Signaling Is Associated with Iron Regulation in High-Altitude EE Mice
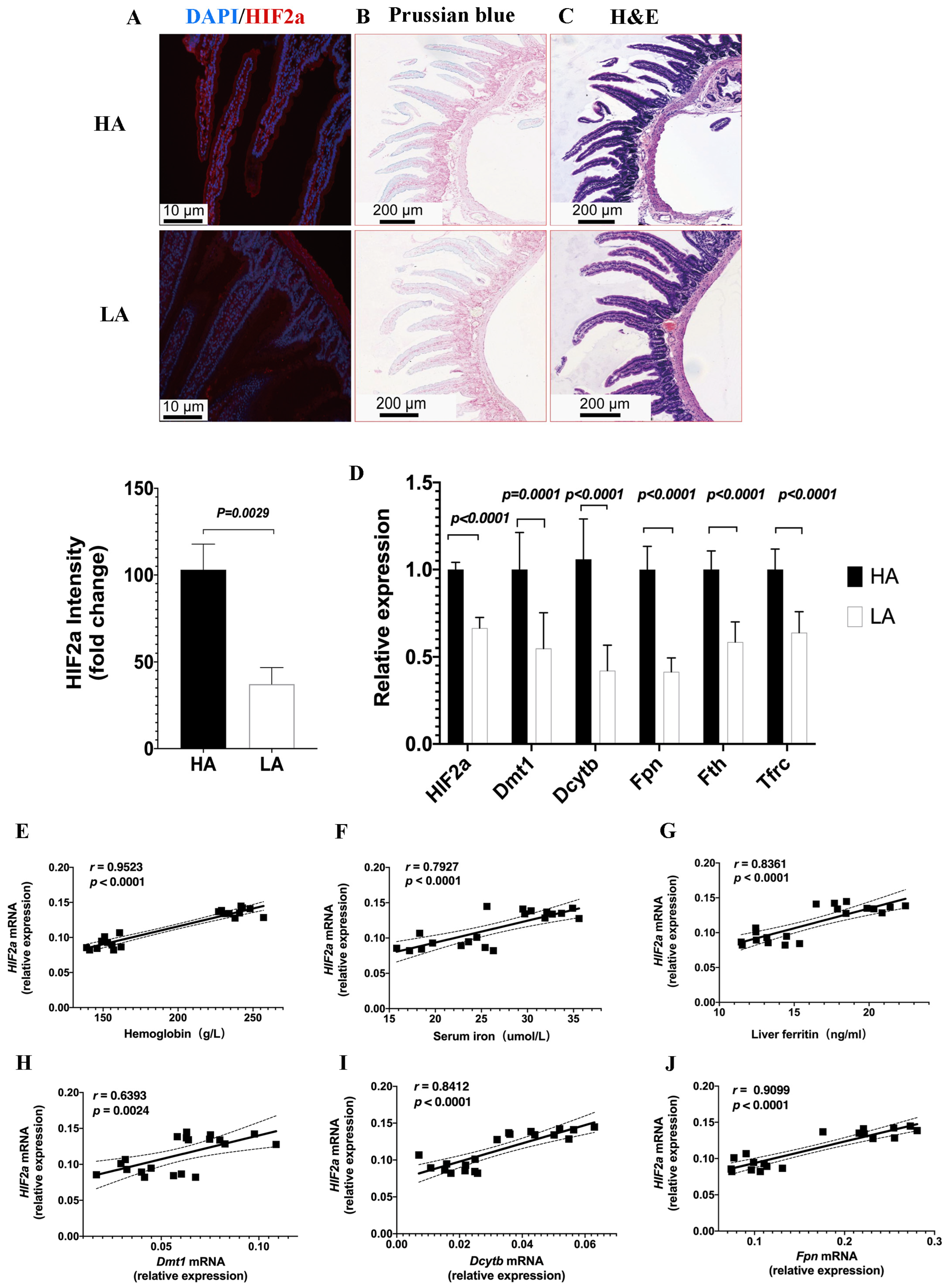
To investigate the relationship between intestinal HIF2α transcriptional expression and iron availability in high-altitude hypoxia, we assessed HIF2α expression on duodenal sections from the HA group using immunofluorescence staining (Figure 4A), which revealed significantly higher HIF2α expression in the HA group compared to the LA group, suggesting that high-altitude hypoxia responses may trigger intestinal HIF2α expression. To confirm whether iron and hypoxia are functionally linked, Prussian blue staining was performed which revealed enhanced progressive iron acquisition in duodenal sections of the high-altitude EE mice compared to the low-altitude mice (Figure 4B). The Hematoxylin and Eosin staining revealed no structural variations between the HA and LA groups (Figure 4C). Moreover, the iron-absorptive and iron-mobilization genes, divalent metal transporter (Dmt1), duodenal cytochrome b (Dcytb), ferroportin (Fpn), and ferritin (Fth), as well as transferrin receptor (Tfrc), were also markedly upregulated in high-altitude EE mice (Figure 4D). The expression of the intestinal HIF2α mRNA was positively associated with Hb, serum iron, and liver ferritin levels, as well as the Dmt1 mRNA, Dcytb mRNA, and Fpn mRNA expression levels (Figure 4E–J). These data indicated that activated HIF2α transcriptional signaling was present in the intestines of high-altitude EE mice, along with increased expression of iron metabolism genes.
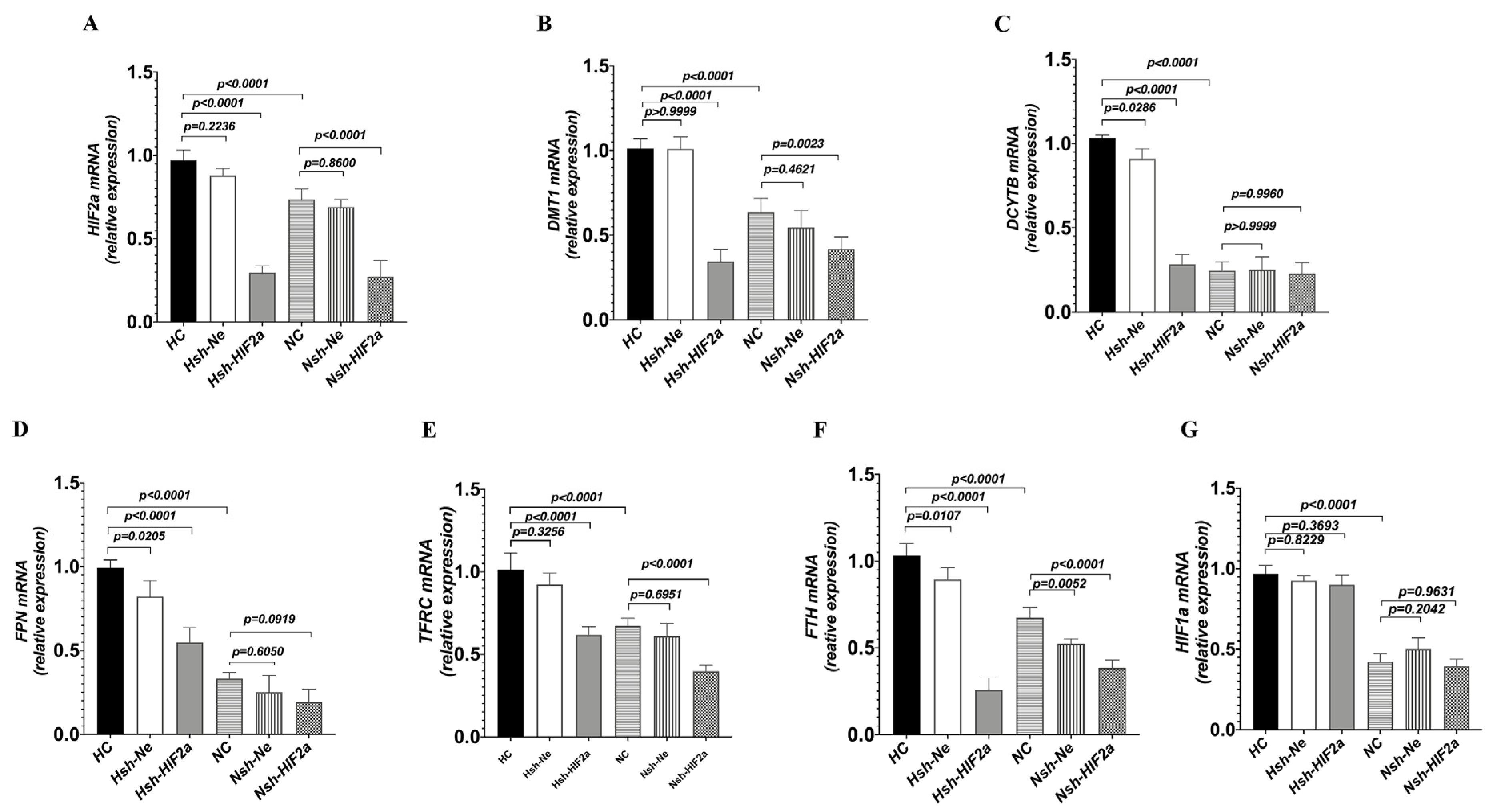
3.3. Knockdown of HIF2a Downregulates Genes Related to Iron Metabolism in HCT116 Cells under Hypoxic Conditions
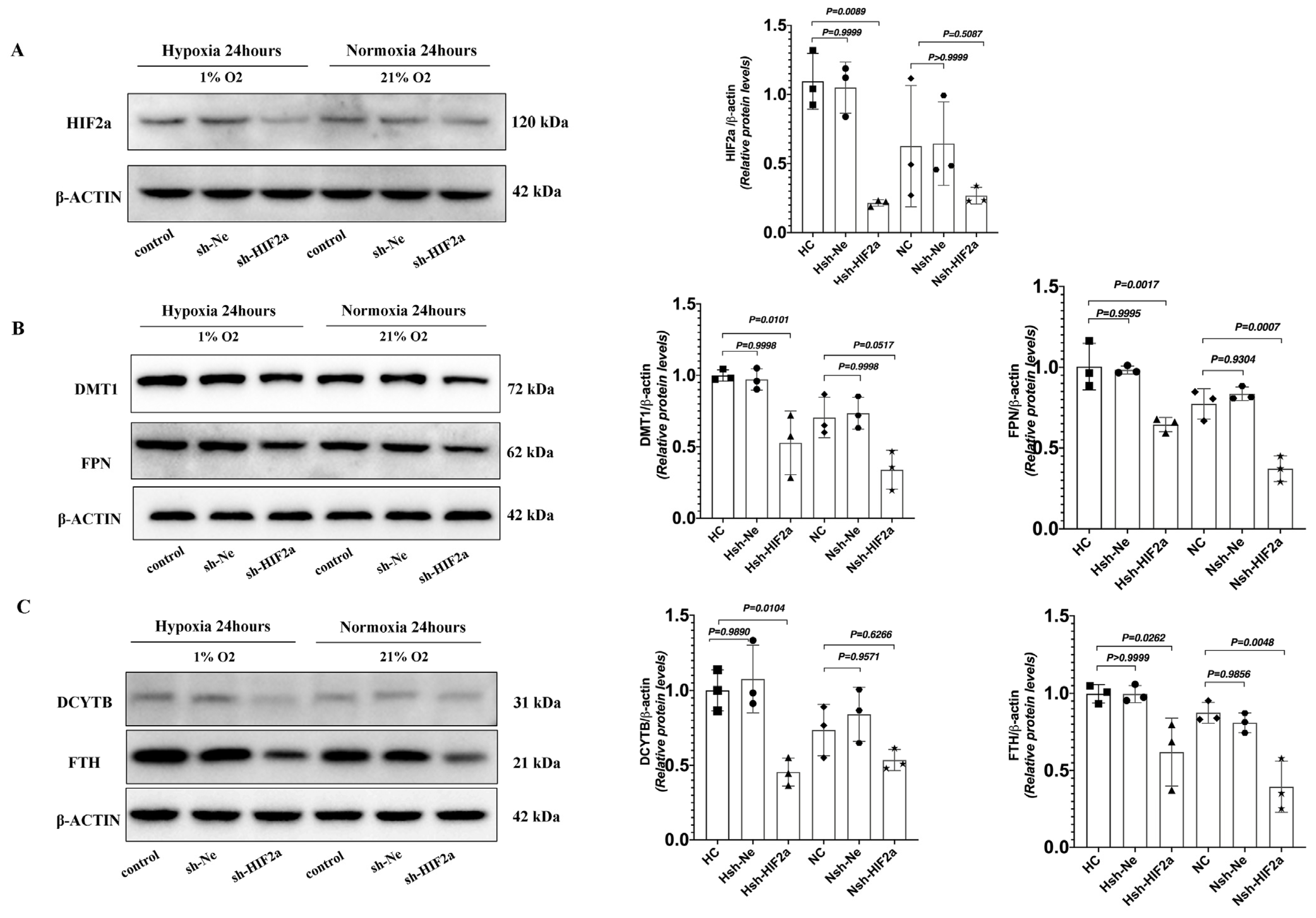
To definitively establish the connection between intestinal HIF2α transcriptional activity and iron metabolism in hypoxia, we used short hairpin RNAs (shRNAs) to generate stable knockdowns of HIF2α mRNA expression in HCT116 cells (sh-HIF2a) (Figure 5A). Compared to shRNA negative control cells (sh-Ne), sh-HIF2a significantly decreased the expression of the iron metabolism genes DMT1, DCYTB, FPN, TFRC, and FTH mRNA (Figure 5B–F) under hypoxia; however, it did not affect the expression of HIF-1a mRNA (Figure 5G). Furthermore, sh-HIF2a significantly decreased hypoxia-mediated induction of HIF2a and iron metabolism genes DMT1, FPN, DCYTB, and FTH protein expression levels in the intestinal cell line (Figure 6A–C). These results further support that intestinal HIF2α signals can promote iron metabolism and increase iron availability during hypoxia.
3.4. Inhibition of Intestinal HIF2α Transcription Decreases Iron Metabolism Gene Expression and Improves Excessive Erythrocytosis in High-Altitude Mice
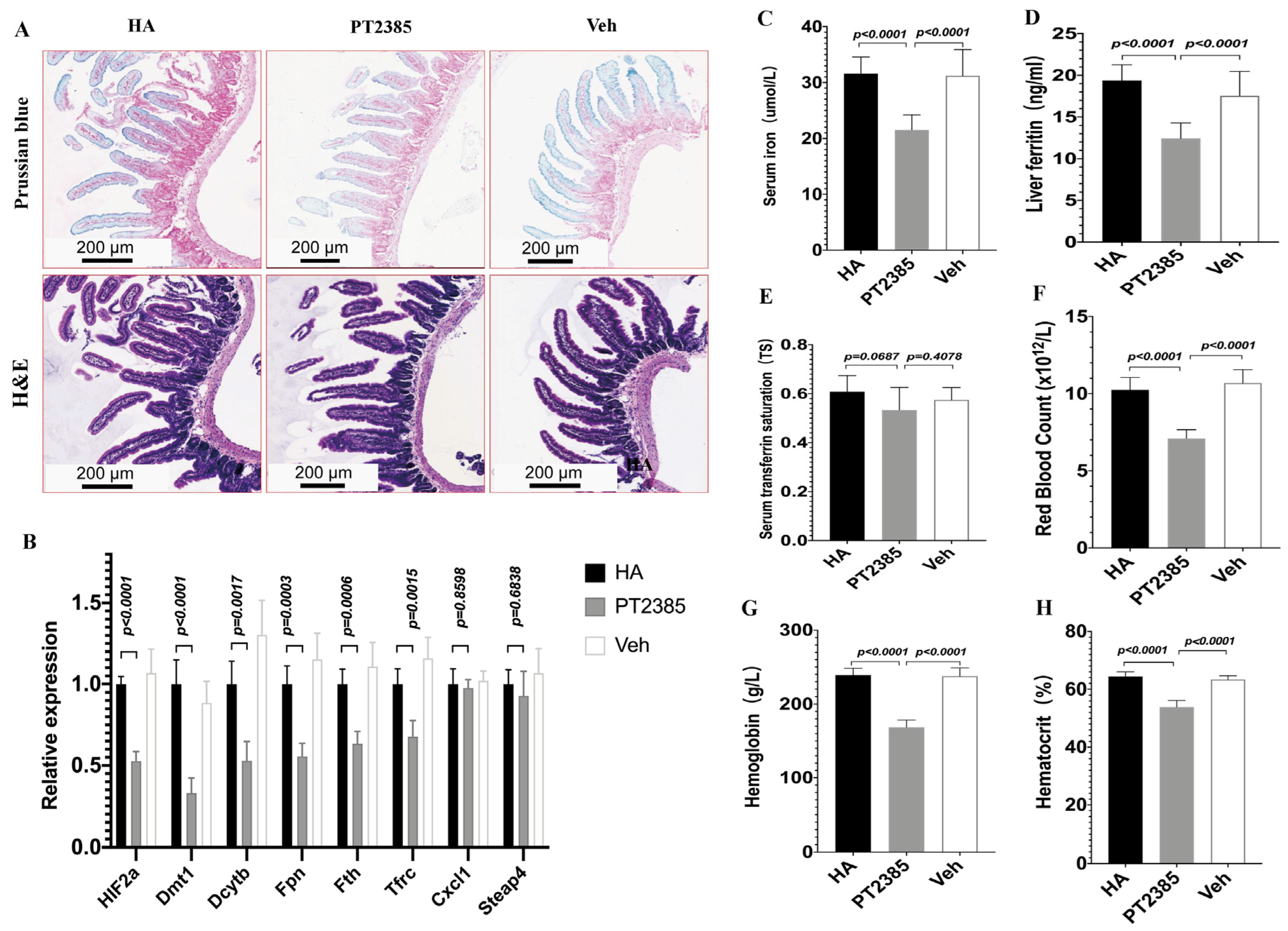
To explore whether decreased iron availability in high-altitude mice ameliorated EE by inhibiting intestinal HIF2α transcription, we designed a controlled experiment in HA mice. Since LA mice did not have a high-altitude erythrocytosis phenotype, we treated HA mice with either the PT2385 drug or a blank drug (vehicle) for two weeks. Notably, Prussian blue staining revealed that in progressive duodenal sections, the iron acquisition rate was reduced in PT2385 mice compared to the untreated HA and vehicle mice. Histological examination revealed no major morphological changes between PT2385 and vehicle mice (Figure 7A). Moreover, the PT2385 treatment significantly decreased the levels of mRNA for HIF2α, Dmt1, Dcytb, Fpn, Tfrc, and Fth compared to those of the untreated HA and Veh groups, but not the HIF2α-specific inflammatory transcripts Cxcl1\Steap4 (Figure 7B). Additionally, our qualitative analysis showed that PT2385 mice exhibited lower serum iron and liver ferritin compared to the untreated HA and vehicle mice. However, TS was not different between the HIF-2α inhibitor PT2385 mice and vehicle mice (Figure 7C–E); Furthermore, RBC numbers significantly increased in the untreated HA and vehicle mice, as did Hb and Hct levels, and in PT2385 mice, these increases were reduced (Figure 7F–H). These data show that decreased iron availability in high-altitude mice ameliorated EE by inhibiting intestinal HIF2α transcription.
4. Discussion
Transcription factors play essential roles in human physiology and disease, highlighting the importance of the continued efforts to understand and regulate the expression of the genome in physiological and pathological processes [27]. In this study, we demonstrated that intestinal HIF2α is critical for activating a transcriptional program that may increase iron availability through the upregulation of iron metabolism genes expression, and promote EE, providing a potential strategy for the treatment of EE diseases.
Dietary iron intake and body iron stores increase with long-term exposure to high-altitude hypoxia [28]. In the present research, we revealed that excessive erythropoiesis is related to enhanced iron intake and utilization in high-altitude mice. A similar pattern of results was reported previously. Populations living at 2210 m altitude had notably higher levels of hemoglobin, hematocrit, and serum ferritin than their peers living at sea level [29]. Moreover, according to a recent study, Hb and ferritin levels rise significantly every 300 m of residential altitude, starting at 300 m above sea level [20]. In another study, high-altitude migrants had greater serum TS levels than sea-level residents, indicating that more iron was prepared due to increased erythropoietic needs [30]. Consistent with these results, in the present study, we observed increased serum iron levels, TS levels, and liver ferritin levels in high-altitude EE mice, owing to enhanced iron availability from external absorption and intrinsic reuse [31].
High-altitude hypoxia may cause EE. In native Tibetans, a strongly selected HIF2α mutation was found to be related to normal hemoglobin at high altitude [32,33]. Similar to this research, we found a significantly higher intestinal HIF2α expression in high-altitude EE mice, which suggests that high-altitude hypoxia responses might trigger intestinal HIF2α transcriptional expression. HIF2α plays an important role in iron hyperabsorption in primary and secondary hemochromatosis [23,24,34]. The activation of HIF2α in the intestine promotes dietary iron intake and iron release from storage organs [15]. Based on the findings, the present study raises the hypothesis that intestinal HIF2α plays an important role in the regulation of systemic iron homeostasis in high-altitude EE mice, which is supported by the close relationship between HIF2α mRNA and protein expression patterns and iron metabolism genes. We also found that intestinal HIF2α transcriptional signaling was positively correlated with Hb and iron availability in high-altitude EE mice, and that activated intestinal HIF2α resulted in upregulated iron metabolism genes, which can cause an excess of iron supply. Furthermore, the easy access to iron provided by active intestinal HIF2α is a risk factor for stimulating EE [28]. Our study indicated that intestinal HIF2α transcription is important in the regulation of systemic iron homeostasis and that activating HIF2α transcription in the intestines of the high-altitude EE mice resulted in increased iron acquisition, utilization, and storage.
Hypoxic-stimulated erythropoiesis requires the availability of extra iron for heme production [19]. HIF2α is essential and sufficient to drive the adaptive increase in iron uptake under hypoxia and erythropoiesis demand by directly transcriptionally activating iron absorption pathways [26,35,36]. In the present study, we evaluated the degree to which intestinal HIF2α contributed to the hypoxic modulation of iron metabolism genes. These included the genes implicated in iron-absorptive and iron-transport genes, Dmt1, Dcytb, Fpn and Tfrc, which were also markedly upregulated in high-altitude EE mice [37]. We validated this finding in vitro by treating HCT116 cells with HIF2αshRNA, confirming that HIF-2α, but not HIF-1α, was a critical regulator of the iron metabolism pathway by measuring the gene and protein expression of DMT1, DCYTB, FPN, and FTH in HCT116 cells during hypoxia. We revealed that the intestinal HIF2α signal centrally influences the activity of the iron metabolism genes in vivo and in vitro.
High-altitude polycythemia (HAPC) patients currently rely on phlebotomy to reduce erythrocyte levels. However, chronic phlebotomy has side effects such as fatigue that can make patients less likely to stick with their treatment plan [11]. According to recent reports, PT2385 selectively suppresses HIF2α transcriptional activity by allosteric regulation, inhibiting HIF2α heterodimerization and its transcriptional dimerization partner ARNT/HIF1β [38,39,40]. In this study, we demonstrated that inhibiting intestinal HIF2α transcriptional activity with PT2385 prevents and improves abnormal iron absorption and metabolism in high-altitude EE mice, followed by a reduction in serum iron and ferritin levels. However, TS did not differ between the HIF2α inhibitor mice and vehicle mice, suggesting a contribution of recycled and stored iron by macrophages of the reticuloendothelial system [41,42]. Reticuloendothelial macrophages act as iron storage and recycle components that are not affected by the HIF2α inhibitor [43]. Recent studies have demonstrated associations between HIF2α downregulation and blood-related phenotypes [44]. Notably, our findings show that hematologic parameters (Hb, RBC, and Hct) greatly increased in an HIF2a-dependent manner during high-altitude hypoxia, which was reversed by treatment with PT2385. The oxygen sensing pathway in duodenal cells was demonstrated to be independent of an individual’s Hb content [20]. Accordingly, with low oxygenation, HIF2α activity increased in duodenal enterocytes, activating dietary iron intake and utilization in circulation. Our findings provide compelling support for the use of HIF2α inhibitors in the treatment of high-altitude hypoxia with EE diseases, many of which are characterized by intestinal HIF2α dysfunction and iron hypermetabolism.
In conclusion, we used a high-altitude EE mouse model to show that intestinal HIF2α contributes to iron hyperabsorption and may cause EE. Moreover, we showed that in high-altitude EE mice, decreasing iron availability ameliorates EE by inhibiting intestinal HIF2α transcription. We discussed that therapeutic intervention in intestinal HIF2α activity could help reduce iron hypermetabolism and improve EE in high-altitude hypoxia. However, animal models do not fully replicate the extent of the EE phenotype seen in humans. Therefore, more preclinical research is needed to determine whether molecules targeting HIF2α treatment are beneficial in patients.
Author Contributions
S.Z.: study design and data collection, original drafting, and statistical analysis. R.-L.G.: project design, study supervision, manuscript revision, and acquisition of financial support. J.Y.: technical assistance and data analysis. K.S.: manuscript revision and technical assistance. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Qinghai Plateau Medicine Clinical Research Center Project (2017-SF-11), the National Natural Science Foundation of China (82072107), and the Qinghai Fundamental Scientific and Technological Research Plan (2018-ZJ-730 and 2019-SF-134).
Institutional Review Board Statement
All animal experiments were performed in compliance with the regulations of the Ministry of Science and Technology of China and approved by the Experimental Committee of Qinghai University (QHDX-2019-09). The HCT116 cell line was obtained from the National Infrastructure of Cell Lines (Beijing, China).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We would like to thank the Research Center for High Altitude Medicine for providing the experimental platform.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
| CMS | Chronic Mountain sickness |
| EE | Excessive erythrocytosis |
| HA | High-altitude |
| HC | Hypoxia control |
| HIF | Hypoxia-inducible factors |
| DMT1 | Divalent metal transporter |
| DCYTB | Duodenal cytochrome b |
| FPN | Ferroportin |
| TFRC | Transferrin receptor |
| FTH | Ferritin |
| CXCL1 | C-X-C motif chemokine ligand 1 |
| STEAP4 | STEAP family member 4 |
| LA | Low-altitude |
| NC | Normoxia control |
| RBC | Red blood cells |
| Hb | Hemoglobin |
| Hct | Hematocrit |
| TIBC | Total Iron Binding Capacity |
| TS | Transferrin saturation |
References
- Hancco, I.; Bailly, S.; Baillieul, S.; Doutreleau, S.; Germain, M.; Pépin, J.-L.; Verges, S. Excessive Erythrocytosis and Chronic Mountain Sickness in Dwellers of the Highest City in the World. Front. Physiol. 2020, 11, 773. [Google Scholar] [CrossRef] [PubMed]
- Vargas, E.; Spielvogel, H. Chronic mountain sickness, optimal hemoglobin, and heart disease. High Alt. Med. Biol. 2006, 7, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Beall, C.M.; Cavalleri, G.L.; Deng, L.; Elston, R.C.; Gao, Y.; Knight, J.; Li, C.; Li, J.C.; Liang, Y.; McCormack, M.; et al. Natural selection on EPAS1 (HIF2α) associated with low hemoglobin concentration in Tibetan highlanders. Proc. Natl. Acad. Sci. USA 2010, 107, 11459–11464. [Google Scholar] [CrossRef] [PubMed]
- Monge, C.; Leon-Velarde, F.; Laguë, S.L.; Coppel, J.; Court, J.; van der Kaaij, J.; Vercueil, A.; Feelisch, M.; Levett, D.; Mythen, M.; et al. Physiological adaptation to high altitude: Oxygen transport in mammals and birds. Physiol. Rev. 1991, 71, 1135–1172. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.G. Measuring high-altitude adaptation. J. Appl. Physiol. 2017, 123, 1371–1385. [Google Scholar] [CrossRef] [PubMed]
- De Ferrari, A.; Miranda, J.J.; Gilman, R.H.; Dávila-Román, V.G.; León-Velarde, F.; Rivera-Ch, M.; Huicho, L.; Bernabé-Ortiz, A.; Wise, R.A.; Checkley, W. Prevalence, clinical profile, iron status, and subject-specific traits for excessive erythrocytosis in andean adults living permanently at 3825 meters above sea level. Chest 2014, 146, 1327–1336. [Google Scholar] [CrossRef]
- Sahota, I.; Panwar, N. Prevalence of Chronic Mountain Sickness in high altitude districts of Himachal Pradesh. Indian J. Occup. Environ. Med. 2013, 17, 94–100. [Google Scholar] [CrossRef]
- Monge, C.; León-Velarde, F.; Arregui, A. Increasing prevalence of excessive erythrocytosis with age among healthy high-altitude miners. N. Engl. J. Med. 1989, 321, 1271. [Google Scholar] [CrossRef]
- Arregui, A.; León-Velarde, F.; Cabrera, J.; Paredes, S.; Vizcarra, D.; Umeres, H. Migraine, polycythemia and chronic mountain sickness. Cephalalgia 1994, 14, 339–341. [Google Scholar] [CrossRef]
- León-Velarde, F.; Maggiorini, M.; Reeves, J.T.; Aldashev, A.; Asmus, I.; Bernardi, L.; Ge, R.L.; Hackett, P.; Kobayashi, T.; Moore, L.G.; et al. Consensus statement on chronic and subacute high altitude diseases. High Alt. Med. Biol. 2005, 6, 147–157. [Google Scholar] [CrossRef]
- Villafuerte, F.C.; Corante, N.; Ucrós, S.; Granados, C.M.; Castro-Rodríguez, J.A.; Hill, C.M.; Parr, N.; Wilkes, M.; Hawkes, L.A.; Grissom, C.K.; et al. Chronic Mountain Sickness: Clinical Aspects, Etiology, Management, and Treatment. High Alt. Med. Biol. 2016, 17, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Azad, P.; Stobdan, T.; Zhou, D.; Hartley, I.; Akbari, A.; Bafna, V.; Haddad, G.G. High-altitude adaptation in humans: From genomics to integrative physiology. J. Mol. Med. 2017, 95, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- León-Velarde, F.; Richalet, J.P. Respiratory control in residents at high altitude: Physiology and pathophysiology. High Alt. Med. Biol. 2006, 7, 125–137. [Google Scholar] [CrossRef]
- Stobdan, T.; Akbari, A.; Azad, P.; Zhou, D.; Poulsen, O.; Appenzeller, O.; Gonzales, G.F.; Telenti, A.; Wong, E.H.M.; Saini, S.; et al. New Insights into the Genetic Basis of Monge’s Disease and Adaptation to High-Altitude. Mol. Biol. Evol. 2017, 34, 3154–3168. [Google Scholar] [CrossRef]
- Gassmann, M.; Muckenthaler, M.U. Adaptation of iron requirement to hypoxic conditions at high altitude. J. Appl. Physiol. 2015, 119, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Muckenthaler, M.U.; Mairbäurl, H.; Gassmann, M. Iron metabolism in high-altitude residents. J. Appl. Physiol. 2020, 129, 920–925. [Google Scholar] [CrossRef]
- Atanasiu, V.; Manolescu, B.; Stoian, I. Hepcidin–central regulator of iron metabolism. Eur. J. Haematol. 2007, 78, 1–10. [Google Scholar] [CrossRef]
- Hirota, K. An intimate crosstalk between iron homeostasis and oxygen metabolism regulated by the hypoxia-inducible factors (HIFs). Free Radic. Biol. Med. 2019, 133, 118–129. [Google Scholar] [CrossRef]
- Yoon, D.; Ponka, P.; Prchal, J.T. Hypoxia. 5. Hypoxia and hematopoiesis. Am. J. Physiol. Physiol. 2011, 300, C1215–C1222. [Google Scholar] [CrossRef]
- Staub, K.; Haeusler, M.; Bender, N.; Morozova, I.; Eppenberger, P.E.; Panczak, R.; Zwahlen, M.; Schaer, D.J.; Maggiorini, M.V.; Ulrich, S.; et al. Hemoglobin concentration of young men at residential altitudes between 200 and 2000m mirrors Switzerland’s topography. Blood 2020, 135, 1066–1069. [Google Scholar] [CrossRef]
- Shah, Y.M.; Matsubara, T.; Ito, S.; Yim, S.-H.; Gonzalez, F.J. Intestinal hypoxia-inducible transcription factors are essential for iron absorption following iron deficiency. Cell Metab. 2009, 9, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Mastrogiannaki, M.; Matak, P.; Keith, B.; Simon, M.C.; Vaulont, S.; Peyssonnaux, C. HIF-2α, but not HIF-1α, promotes iron absorption in mice. J. Clin. Investig. 2009, 119, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Xie, L.; Ramakrishnan, S.K.; Campbell, A.; Rivella, S.; Shah, Y.M. Intestine-specific Disruption of Hypoxia-inducible Factor (HIF)-2α Improves Anemia in Sickle Cell Disease. J. Biol. Chem. 2015, 290, 23523–23527. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.R.; Taylor, M.; Xue, X.; Ramakrishnan, S.K.; Martin, A.; Xie, L.; Bredell, B.X.; Gardenghi, S.; Rivella, S.; Shah, Y.M. Intestinal HIF2α promotes tissue-iron accumulation in disorders of iron overload with anemia. Proc. Natl. Acad. Sci. USA 2013, 110, E4922–E4930. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Yagai, T.; Luo, Y.; Liang, X.; Chen, T.; Wang, Q.; Sun, D.; Zhao, J.; Ramakrishnan, S.K.; Sun, L.; et al. Activation of intestinal hypoxia-inducible factor 2α during obesity contributes to hepatic steatosis. Nat. Med. 2017, 23, 1298–1308. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.R.; Xue, X.; Shah, Y.M. Intestinal hypoxia-inducible factor-2α (HIF-2α) is critical for efficient erythropoiesis. J. Biol. Chem. 2011, 286, 19533–19540. [Google Scholar] [CrossRef]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef]
- Liu, Y.-S.; Huang, H.; Zhou, S.-M.; Tian, H.-J.; Li, P. Excessive Iron Availability Caused by Disorders of Interleukin-10 and Interleukin-22 Contributes to High Altitude Polycythemia. Front. Physiol. 2018, 9, 548. [Google Scholar] [CrossRef]
- Brothers, M.D.; Doan, B.K.; Zupan, M.F.; Wile, A.L.; Wilber, R.L.; Byrnes, W.C.; Barnes, K.R.; Kilding, A.E.; Hamid, N.A.; Azidin, R.M.F.R.; et al. Hematological and physiological adaptations following 46 weeks of moderate altitude residence. High Alt. Med. Biol. 2010, 11, 199–208. [Google Scholar] [CrossRef]
- Northrop-Clewes, C.A.; Thurnham, D.I. Biomarkers for the differentiation of anemia and their clinical usefulness. J. Blood Med. 2013, 4, 11–22. [Google Scholar] [CrossRef]
- Salvin, H.E.; Pasricha, S.-R.; Marks, D.C.; Speedy, J. Iron deficiency in blood donors: A national cross-sectional study. Transfusion 2014, 54, 2434–2444. [Google Scholar] [CrossRef] [PubMed]
- Painschab, M.S.; Malpartida, G.E.; Dávila-Roman, V.G.; Gilman, R.H.; Kolb, T.M.; León-Velarde, F.; Miranda, J.J.; Checkley, W. Association between serum concentrations of hypoxia inducible factor responsive proteins and excessive erythrocytosis in high altitude Peru. High Alt. Med. Biol. 2015, 16, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.; Simonson, T.S.; Gordeuk, V.; Prchal, J.T.; McClain, D.A. Metabolic aspects of high-altitude adaptation in Tibetans. Exp. Physiol. 2015, 100, 1247–1255. [Google Scholar] [CrossRef]
- Mastrogiannaki, M.; Matak, P.; Delga, S.; Deschemin, J.-C.; Vaulont, S.; Peyssonnaux, C. Deletion of HIF-2α in the enterocytes decreases the severity of tissue iron loading in hepcidin knockout mice. Blood 2012, 119, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.; Qu, A.; Anderson, E.R.; Matsubara, T.; Martin, A.; Gonzalez, F.J.; Shah, Y.M. Hypoxia-inducible factor-2α mediates the adaptive increase of intestinal ferroportin during iron deficiency in mice. Gastroenterology 2011, 140, 2044–2055. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, S.K.; Anderson, E.R.; Martin, A.; Centofanti, B.; Shah, Y.M. Maternal intestinal HIF-2α is necessary for sensing iron demands of lactation in mice. Proc. Natl. Acad. Sci. USA 2015, 112, E3738–E3747. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.S.; Percy, M.J. The HIF pathway and erythrocytosis. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 165–192. [Google Scholar] [CrossRef]
- Wallace, E.M.; Rizzi, J.P.; Han, G.; Wehn, P.M.; Cao, Z.; Du, X.; Cheng, T.; Czerwinski, R.M.; Dixon, D.D.; Goggin, B.S.; et al. A Small-Molecule Antagonist of HIF2α Is Efficacious in Preclinical Models of Renal Cell Carcinoma. Cancer Res. 2016, 76, 5491–5500. [Google Scholar] [CrossRef]
- Cho, H.; Du, X.; Rizzi, J.P.; Liberzon, E.; Chakraborty, A.A.; Gao, W.; Carvo, I.; Signoretti, S.; Bruick, R.K.; Josey, J.A.; et al. On-target efficacy of a HIF-2α antagonist in preclinical kidney cancer models. Nature 2016, 539, 107–111. [Google Scholar] [CrossRef]
- Chen, W.; Hill, H.; Christie, A.; Kim, M.S.; Holloman, E.; Pavia-Jimenez, A.; Homayoun, F.; Ma, Y.; Patel, N.; Yell, P.; et al. Targeting renal cell carcinoma with a HIF-2 antagonist. Nature 2016, 539, 112–117. [Google Scholar] [CrossRef]
- Camaschella, C. Iron-Deficiency Anemia. N. Engl. J. Med. 2015, 372, 1832–1843. [Google Scholar] [CrossRef] [PubMed]
- de Back, D.Z.; Kostova, E.B.; van Kraaij, M.; van den Berg, T.K.; van Bruggen, R. Of macrophages and red blood cells; a complex love story. Front. Physiol. 2014, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, J.R.; Heinis, M.; Zumerle, S.; Delga, S.; Le Bon, A.; Peyssonnaux, C. Investigating the real role of HIF-1 and HIF-2 in iron recycling by macrophages. Haematologica 2014, 99, e112–e114. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Cui, C.; He, Y.; Ouzhuluobu; Zhang, H.; Yang, D.; Zhang, Q.; Bianbazhuoma; Yang, L.; He, Y.; et al. Down-Regulation of EPAS1 Transcription and Genetic Adaptation of Tibetans to High-Altitude Hypoxia. Mol. Biol. Evol. 2017, 34, 818–830. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flow chart of the mouse experimental design.

Figure 2.
Flow chart of the cell experimental design.

Figure 3.
High-altitude hypoxia affects hematological parameters and iron status indicators in mice. (A) Hemoglobin levels, (B) red blood cell counts, (C) hematocrit levels, (D) serum iron, (E) serum transferrin saturation, and (F) liver ferritin. (A–C) n = 10 per group; (D–F) n = 5–10 per group. Data are presented as the mean ± SD. * p < 0.05. Significance was calculated by a two-tailed, unpaired t-test.
Figure 3.
High-altitude hypoxia affects hematological parameters and iron status indicators in mice. (A) Hemoglobin levels, (B) red blood cell counts, (C) hematocrit levels, (D) serum iron, (E) serum transferrin saturation, and (F) liver ferritin. (A–C) n = 10 per group; (D–F) n = 5–10 per group. Data are presented as the mean ± SD. * p < 0.05. Significance was calculated by a two-tailed, unpaired t-test.

Figure 4.
High-altitude hypoxia increases intestinal HIF2α levels and is associated with Hb and iron metabolism. (A) Immunofluorescence staining of HIF2α expression on duodenal sections from the HA and LA groups. Original magnification was ×200. Quantifications of the fluorescence intensity signals are shown in the bar graphs. (B) Prussian blue iron staining of duodenal tissues. (C) Hematoxylin and eosin staining of duodenal tissues. (D) Quantitative PCR for HIF2α and iron metabolism genes: HIF2α\Dmt1\Dcytb\Fpn\Fth\Tfrc. (E–J) Correlative analyses of intestinal HIF2α mRNA levels with Hb (E), serum iron (F), liver ferritin (G), Dmt1 mRNA (H), Dcytb mRNA (I), and Fpn mRNA (J). (A–C) n = 3 per group, (D) n = 10 per group, (E–J) n = 10 per group. Correlations were assessed via Pearson’s test. Data are presented as the mean ± SD. Significance was calculated by a two-tailed, unpaired t-test.
Figure 4.
High-altitude hypoxia increases intestinal HIF2α levels and is associated with Hb and iron metabolism. (A) Immunofluorescence staining of HIF2α expression on duodenal sections from the HA and LA groups. Original magnification was ×200. Quantifications of the fluorescence intensity signals are shown in the bar graphs. (B) Prussian blue iron staining of duodenal tissues. (C) Hematoxylin and eosin staining of duodenal tissues. (D) Quantitative PCR for HIF2α and iron metabolism genes: HIF2α\Dmt1\Dcytb\Fpn\Fth\Tfrc. (E–J) Correlative analyses of intestinal HIF2α mRNA levels with Hb (E), serum iron (F), liver ferritin (G), Dmt1 mRNA (H), Dcytb mRNA (I), and Fpn mRNA (J). (A–C) n = 3 per group, (D) n = 10 per group, (E–J) n = 10 per group. Correlations were assessed via Pearson’s test. Data are presented as the mean ± SD. Significance was calculated by a two-tailed, unpaired t-test.

Figure 5.
Knockdown of HIF2α expression in HCT116 cells decreased iron metabolism gene expression under hypoxic conditions. The cells were transduced with sh-HIF2a, as a control sh-Ne to silence the HIF2α gene, and then incubated under hypoxic or normoxic conditions for 24 h. (A) Quantitative PCR showed the efficiency of HIF-2α silencing. (B–F) Quantitative PCR showed the efficacy of HIF2α silencing on DMT1, DCYTB, FPN, TFRC, and FTH, as well as (G) HIF1a expression. (A) n = 5; (B–G) n = 5. Statistical significance was calculated by one-way analysis of variance with Tukey’s post hoc test.
Figure 5.
Knockdown of HIF2α expression in HCT116 cells decreased iron metabolism gene expression under hypoxic conditions. The cells were transduced with sh-HIF2a, as a control sh-Ne to silence the HIF2α gene, and then incubated under hypoxic or normoxic conditions for 24 h. (A) Quantitative PCR showed the efficiency of HIF-2α silencing. (B–F) Quantitative PCR showed the efficacy of HIF2α silencing on DMT1, DCYTB, FPN, TFRC, and FTH, as well as (G) HIF1a expression. (A) n = 5; (B–G) n = 5. Statistical significance was calculated by one-way analysis of variance with Tukey’s post hoc test.

Figure 6.
Western Blot analysis of sh-HIF2a in HCT116 cells decreased iron metabolism genes protein expression under hypoxic conditions. (A) The protein expression of HIF2a was determined by Western blot in HCT116 cells. (B) The protein expression of DMT1 and FPN was determined by Western blot in HCT116cells. (C) The protein expression of DCYTB and FTH was determined by Western blot in HCT116cells. (A–C) n = 3 per group. Statistical significance was calculated by one-way analysis of variance with Tukey’s post hoc test.
Figure 6.
Western Blot analysis of sh-HIF2a in HCT116 cells decreased iron metabolism genes protein expression under hypoxic conditions. (A) The protein expression of HIF2a was determined by Western blot in HCT116 cells. (B) The protein expression of DMT1 and FPN was determined by Western blot in HCT116cells. (C) The protein expression of DCYTB and FTH was determined by Western blot in HCT116cells. (A–C) n = 3 per group. Statistical significance was calculated by one-way analysis of variance with Tukey’s post hoc test.

Figure 7.
Inhibition of intestinal HIF2α using PT2385 decreases iron availability and ameliorates excessive erythrocytosis in high-altitude mice. (A) Prussian blue iron staining, Hematoxylin and eosin staining of the double tissue. (B) Quantitative PCR analysis of HIF2α mRNA and related genes Dmt1\Dcytb\Fpn\Fth\Tfrc\Cxcl1\Steap4. (C) Serum iron, (D) liver ferritin, and (E) serum transferrin saturation. (F–H) Hematological analyses (RBC, Hb, and Hct). (A) n = 3 per group; (B) n = 5 per group. (C–H) n = 10 per group. Data are presented as the mean ± SD. Statistical significance was calculated by one-way analysis of variance with Tukey’s post hoc test.
Figure 7.
Inhibition of intestinal HIF2α using PT2385 decreases iron availability and ameliorates excessive erythrocytosis in high-altitude mice. (A) Prussian blue iron staining, Hematoxylin and eosin staining of the double tissue. (B) Quantitative PCR analysis of HIF2α mRNA and related genes Dmt1\Dcytb\Fpn\Fth\Tfrc\Cxcl1\Steap4. (C) Serum iron, (D) liver ferritin, and (E) serum transferrin saturation. (F–H) Hematological analyses (RBC, Hb, and Hct). (A) n = 3 per group; (B) n = 5 per group. (C–H) n = 10 per group. Data are presented as the mean ± SD. Statistical significance was calculated by one-way analysis of variance with Tukey’s post hoc test.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers and base sequences for RT-PCR detection.
| Species | Gene | Forward Primer | Reverse Primer |
|---|---|---|---|
| Mouse | -actin | CTACCTCATGAAGATCCTGACC | CACAGCTTCTCTTTGATGTCAC |
| HIF-2a | GAGAACCTGACTCTCAAAAACG | GTTGTTGTAGACTCTCACTTGC | |
| Dmt1 | TTTTGGACAAATATGGCTTGCG | TACTCATATCCAAACGTGAGGG | |
| Dcytb | GTGTTTGAGTACCACAATGTCC | TGGAAGCAGAAAGACGAAAAAG | |
| Fpn | TTGTGTGTGATCTCCGTATTCA | GTTGTAAAGACGGTCTCAGGTA | |
| Tfrc | TCACACTCTCTCAGCTTTAGTG | TGGTTTCTGAAGAGGGTTTCAT | |
| Fth | TAAAGAAACCAGACCGTGATGA | ATTCACACTCTTTTCCAAGTGC | |
| Cxcl1 | GGCTGGGATTCACCTCAAGAACATC | TGAGTGTGGCTATGACTTCGGTTTG | |
| Steap4 | CCTCTGTGCTGTGCGTCTTCTTC | ACACGATTCGGGATGGAAATGGC | |
| Human | -ACTIN | GGCACCACACCTTCTACAATGAGC | GATAGCACAGCCTGGATAGCAACG |
| HIF-1a | CCATTAGAAAGCAGTTCCGCAAGC | GTGGTAGTGGTGGCATTAGCAGTAG | |
| HIF-2a | ATCAGCAAGTTCATGGGACTTA | AAACCAGAGCCATTTTTGAGAC | |
| DMT1 | CATCCTCACATTTACGAGCTTG | CCAACCCAAGTAGAACACAAAG | |
| DCYTB | GCCAGAAGGTGTTTTCGTAAAT | TGGTAGAATTTGGCTCCTTAGG | |
| FPN | ACAATACGAAGGATTGACCAGT | ATACCAAGTTCCATCCCGAAAT | |
| TFRC | TGAACCAATACAGAGCAGACAT | GTTTTCTCAGCATTCCCGAAAT | |
| FTH | CTCCTACGTTTACCTGTCCATG | CAAGTCATCAGGCACATACAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, S.; Yan, J.; Song, K.; Ge, R.-L. High-Altitude Hypoxia Induces Excessive Erythrocytosis in Mice via Upregulation of the Intestinal HIF2a/Iron-Metabolism Pathway. Biomedicines 2023, 11, 2992. https://doi.org/10.3390/biomedicines11112992
AMA Style
Zhou S, Yan J, Song K, Ge R-L. High-Altitude Hypoxia Induces Excessive Erythrocytosis in Mice via Upregulation of the Intestinal HIF2a/Iron-Metabolism Pathway. Biomedicines. 2023; 11(11):2992. https://doi.org/10.3390/biomedicines11112992
Chicago/Turabian StyleZhou, Sisi, Jun Yan, Kang Song, and Ri-Li Ge. 2023. "High-Altitude Hypoxia Induces Excessive Erythrocytosis in Mice via Upregulation of the Intestinal HIF2a/Iron-Metabolism Pathway" Biomedicines 11, no. 11: 2992. https://doi.org/10.3390/biomedicines11112992
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.