
Circulating CD8+ T Cell Subsets in Primary Sjögren’s Syndrome
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Laboratory and Pathological Determination
2.2. Sample Collection
2.3. Immunophenotyping of Circulating CD8+ T Cell Subset Maturation Stages and ‘Polarized’ CD8+ T Cell Subsets
2.4. Cytokine and Chemokine Measurement
2.5. Statistical Analysis
3. Results
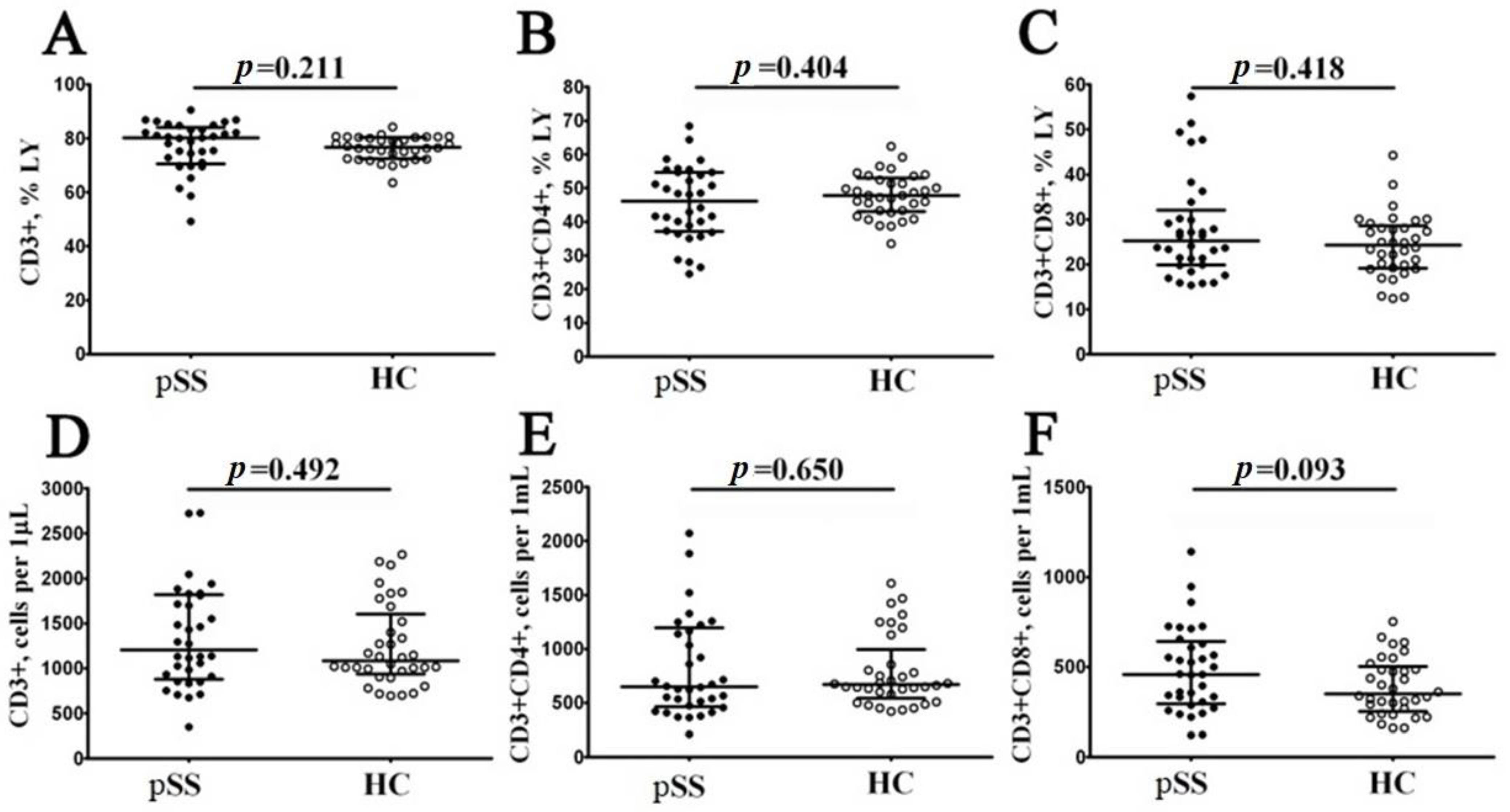
3.1. Main Peripheral Blood CD3+ T Cell Subsets in Patients with Sjögren’s Syndrome
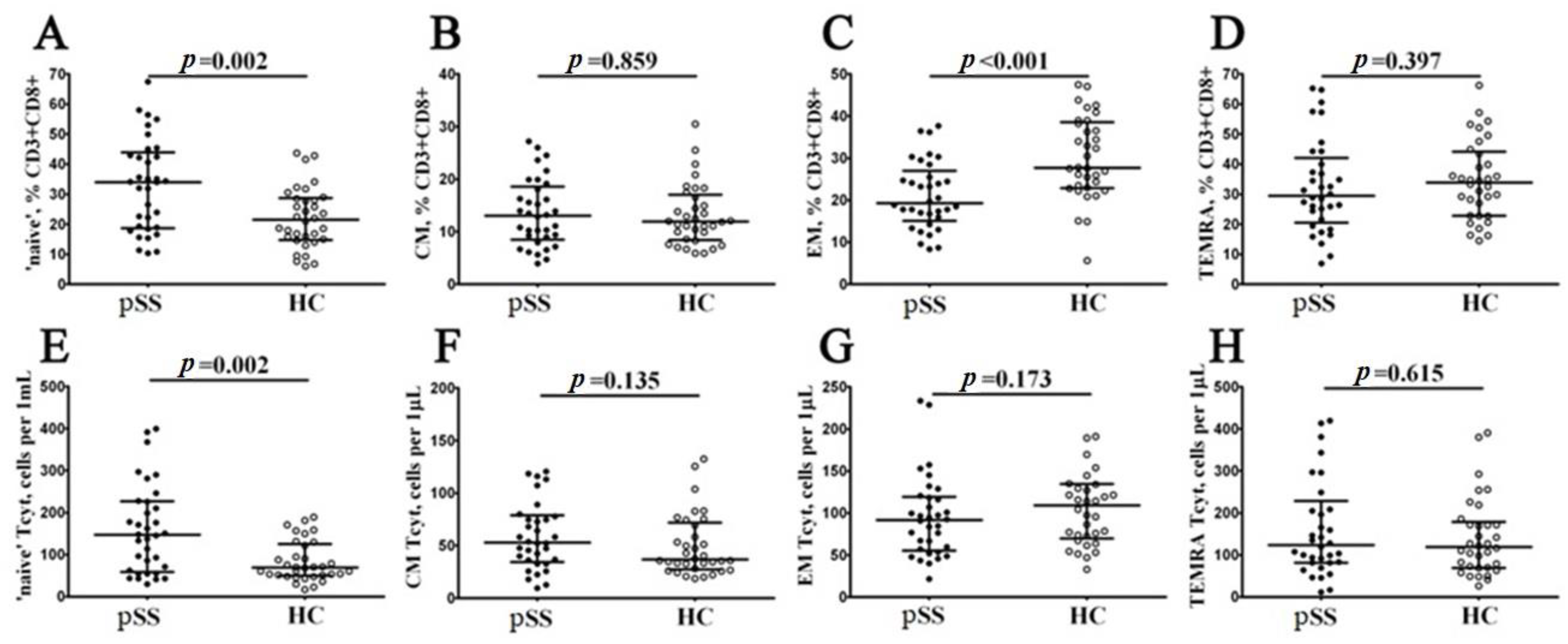
3.2. Alterations in CD8+ T Cell Maturation Subsets in Patients with Sjögren’s Syndrome
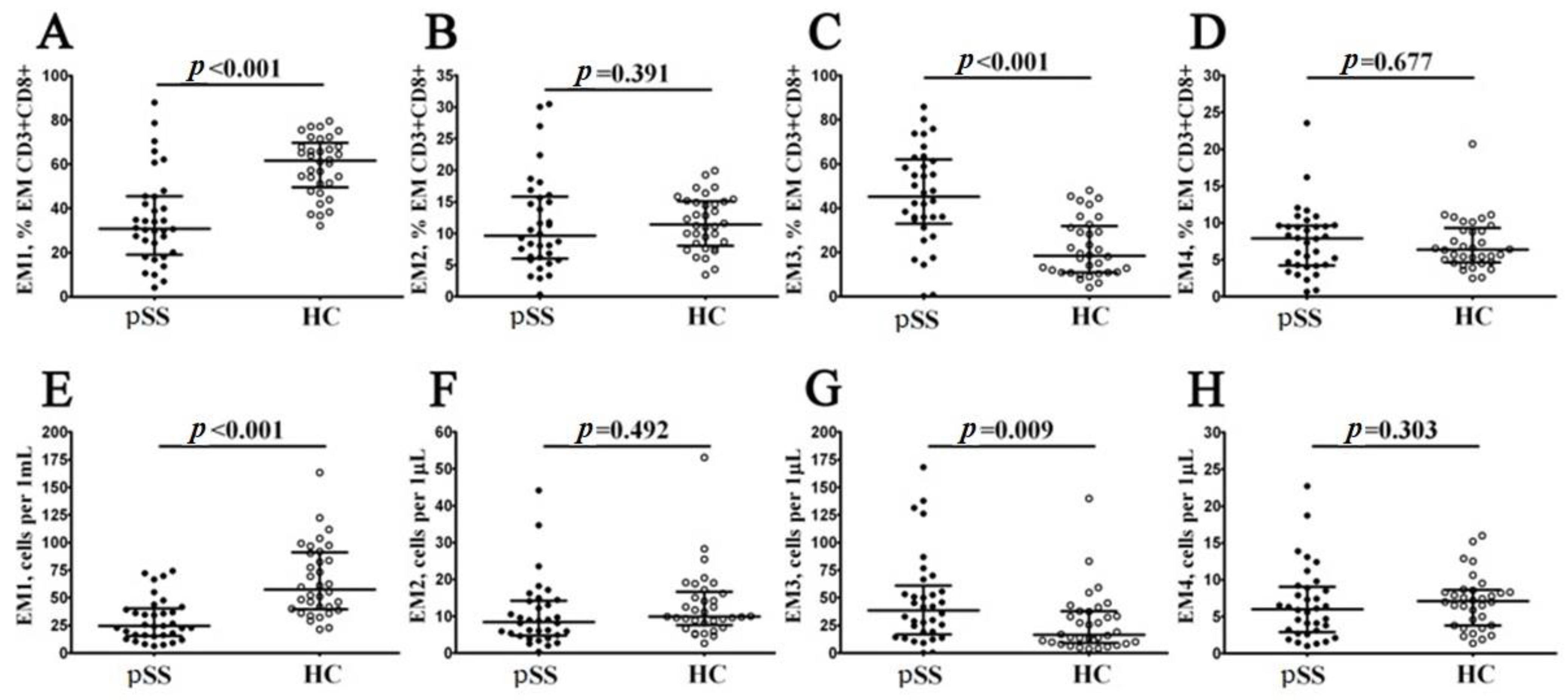
3.3. Imbalance in EM and TEMRA CD8+ T Cell Subsets in Patients with Sjögren’s Syndrome
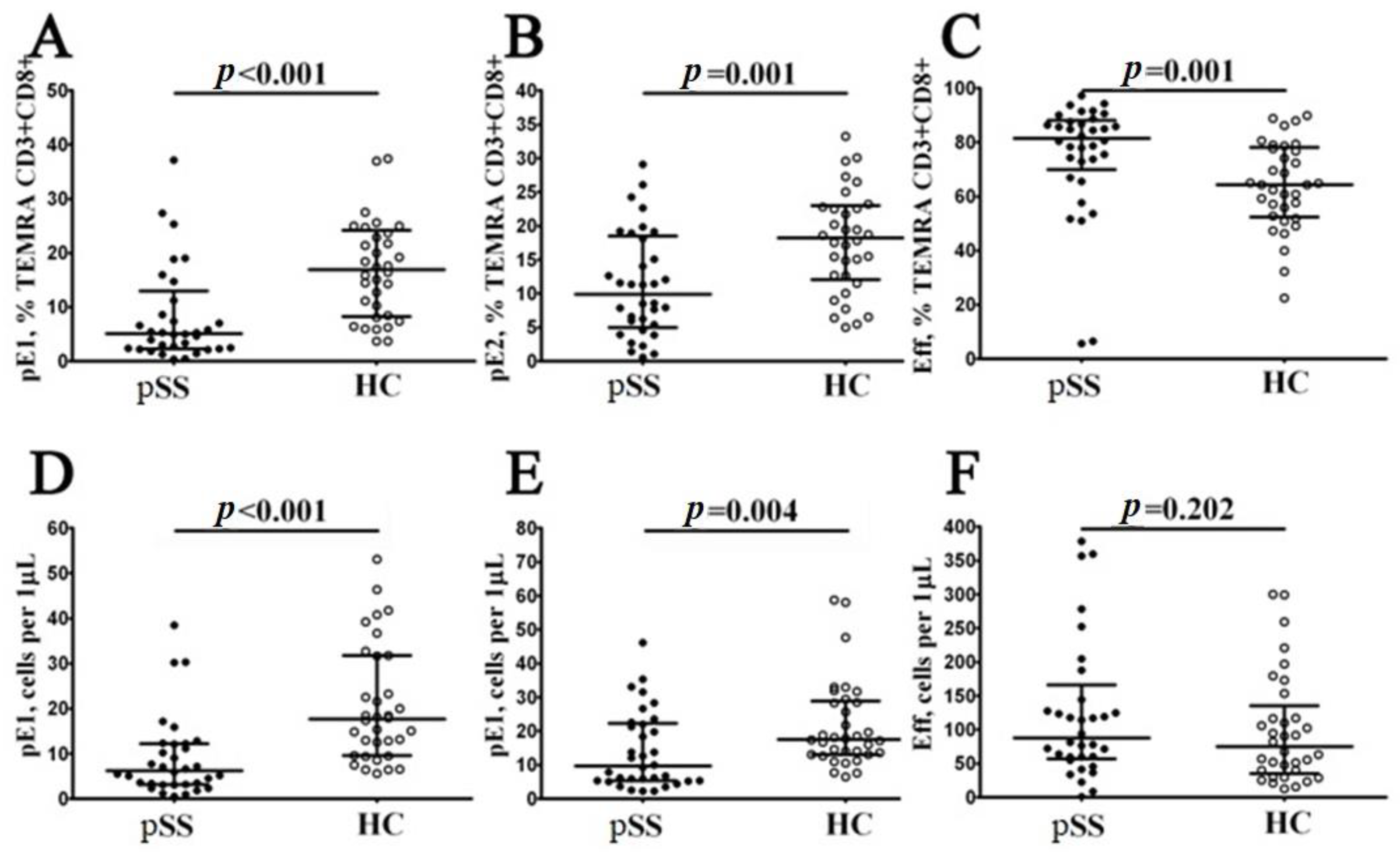
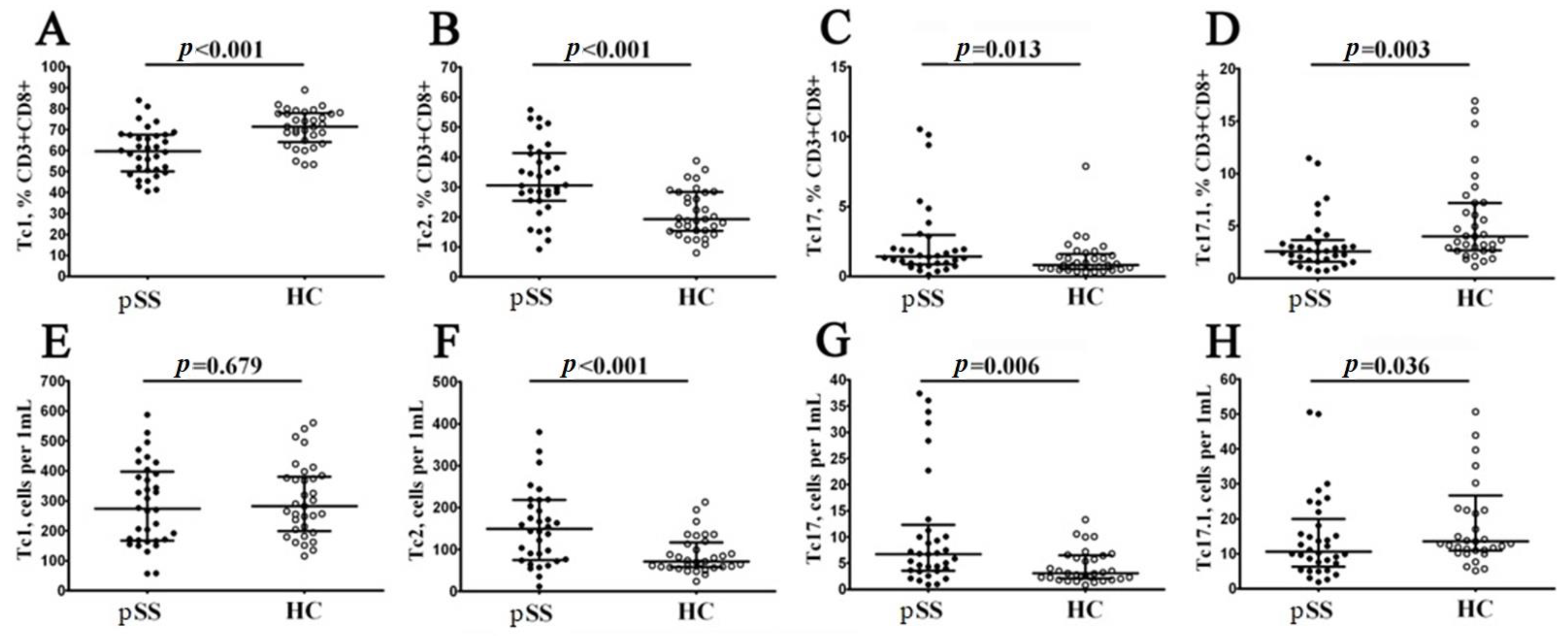
3.4. Imbalance in Peripheral Blood CD8+ T Cells ‘Polarization’ in Patients with Sjögren’s Syndrome
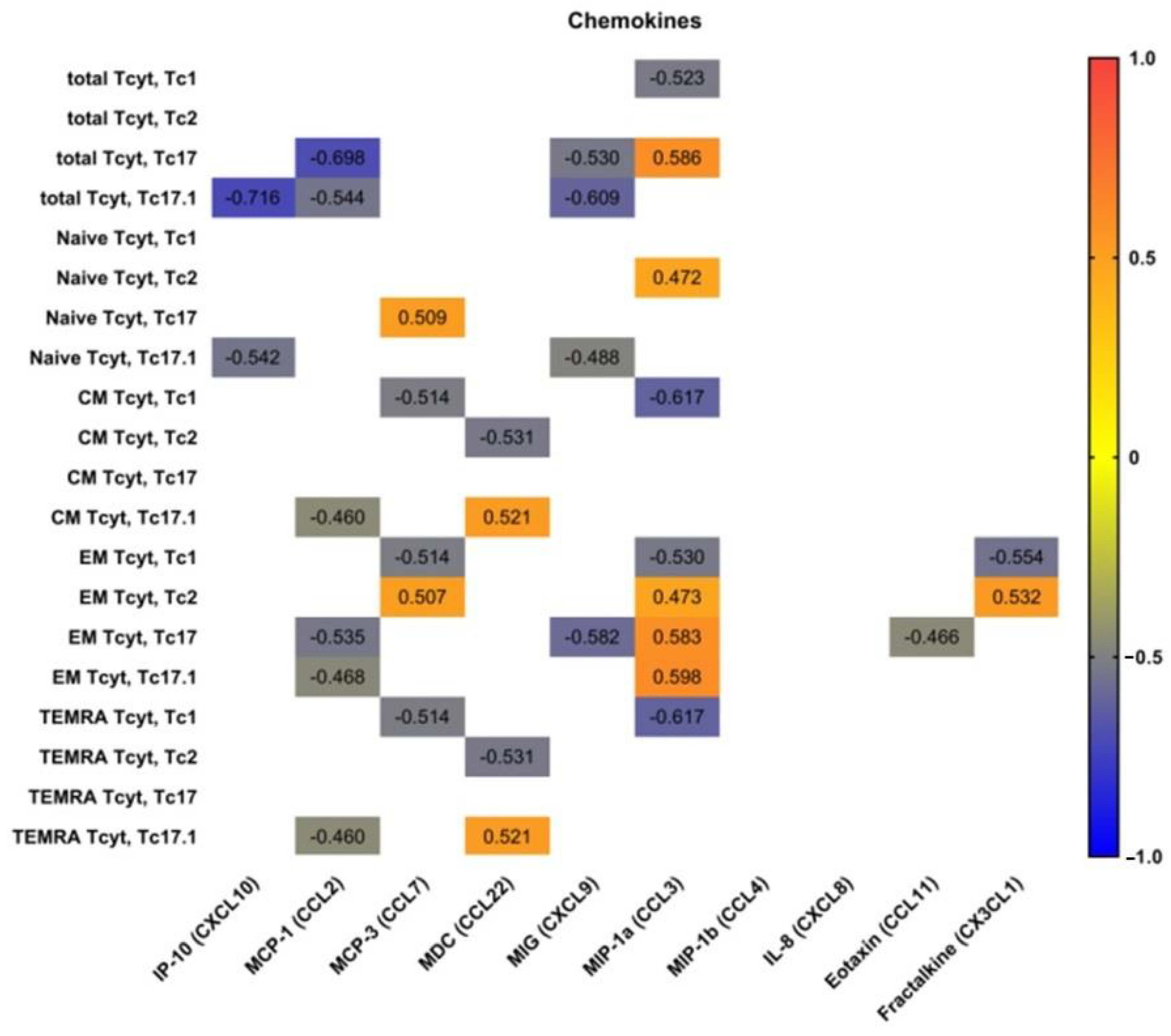
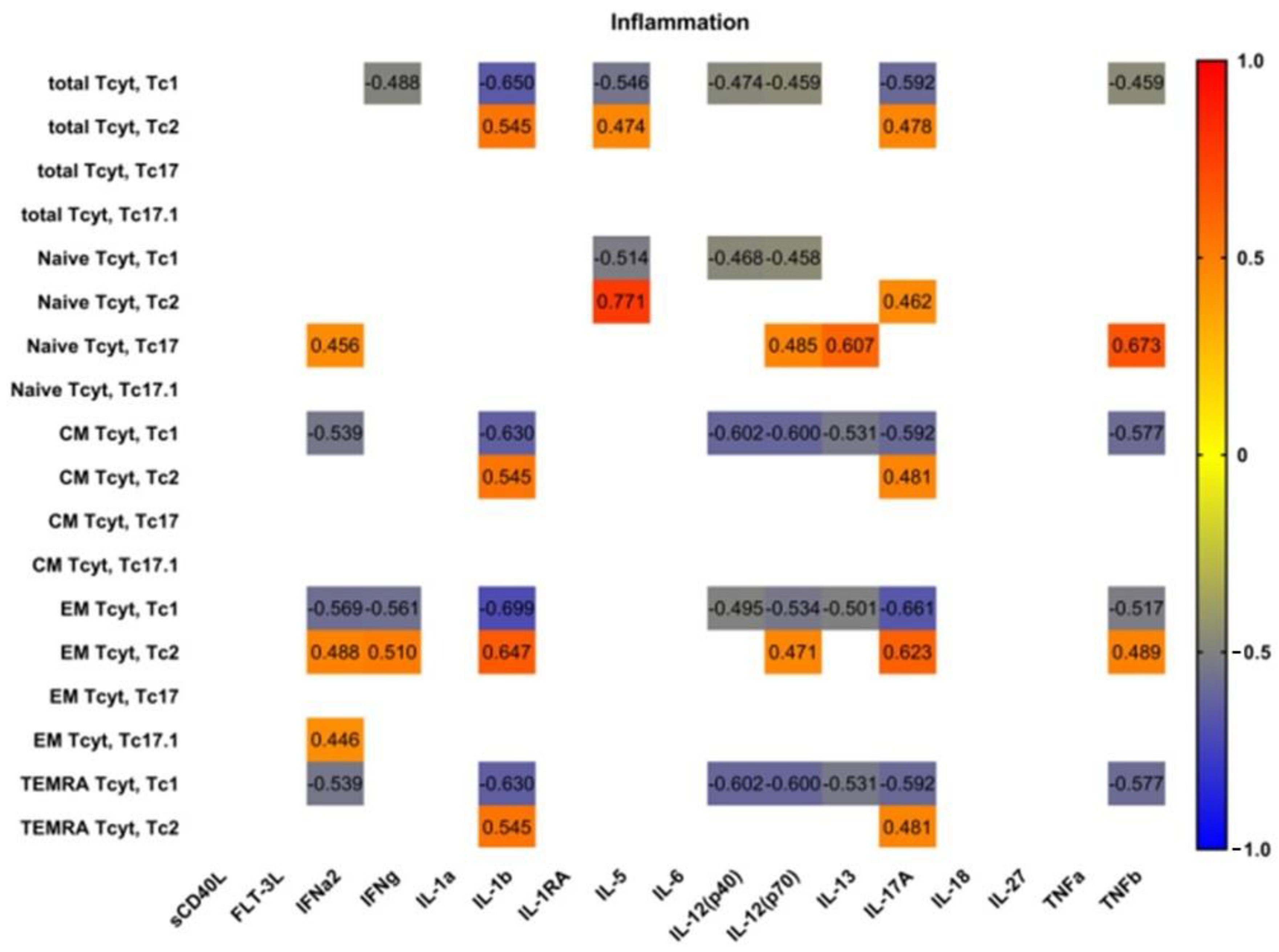
3.5. Blood Level of ‘Polarized’ CD8+ T Cell Subsets and Correlations with Cytokines and Chemokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Firestein, G.S.; Budd, R.C.; Gabriel, S.E.; McInnes, I.B.; O’Dell, J.R. Kelley and Firestein’s Textbook of Rheumatology, 10th ed.; Elsevier Health Sciences: Philadelphia, PA, USA, 2017; pp. 1221–1244. [Google Scholar]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjögren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.A.; Brown, R.M.; Bowman, S.J.; Barone, F. A review of salivary gland histopathology in primary Sjögren’s syndrome with a focus on its potential as a clinical trials. Ann. Rheum. Dis. 2015, 74, 1645–1650. [Google Scholar] [CrossRef] [PubMed]
- Gwenny, M.; Verstappen, F.G.; Kroese, M.; Bootsma, H. T cells in primary Sjögren’s syndrome: Targets for early intervention. Rheumatology 2021, 60, 30883098. [Google Scholar]
- Kassan, S.S.; Moutsopoulos, H.M. Clinical Manifestations and Early Diagnosis of Sjögren Syndrome. Arch. Intern. Med. 2004, 164, 1275–1284. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, J.; Tian, J.; Wang, S. CD8+ T Lymphocytes: Crucial Players in Sjögren’s Syndrome. Front. Immunol. 2021, 11, 602823. [Google Scholar] [CrossRef]
- Barr, J.Y.; Wang, X.; Meyerholz, D.K.; Lieberman, S.M. CD8 T cells contribute to lacrimal gland pathology in the nonobese diabetic mouse model of Sjögren syndrome. Immunol. Cell Biol. 2017, 95, 684–694. [Google Scholar] [CrossRef]
- Mingueneau, M.; Boudaoud, S.; Haskett, S.; Reynolds, T.L.; Nocturne, G.; Norton, E.; Zhang, X.; Constant, M.; Park, D.; Wang, W. Cytometry by time-of-flight immunophenotyping identifies a blood Sjögren’s signature correlating with disease activity and glandular inflammation. J. Allergy Clin. Immunol. 2016, 137, 1809–1821. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, T.; Fujita, H.; Tsubota, K.; Saito, K.; Tsuzaka, K.; Abe, T.; Takeuchi, T. Preferential localization of CD8+ alpha E beta 7+ T cells around acinar epithelial cells with apoptosis in patients with Sjögren’s syndrome. J. Immunol. 1999, 163, 2226–2235. [Google Scholar] [CrossRef]
- Kaneko, N.; Chen, H.; Perugino, C.A.; Maehara, T.; Munemura, R.; Yokomizo, S.; Sameshima, J.; Diefenbach, T.J.; Premo, K.R.; Chinju, A.; et al. Cytotoxic CD8+ T cells may be drivers of tissue destruction in Sjögren’s syndrome. Sci. Rep. 2022, 12, 15427. [Google Scholar] [CrossRef]
- Zhang, X.; Schaumburg, C.S.; Coursey, T.G.; Siemasko, K.F.; Volpe, E.A.; Gandhi, N.B.; Li, D.-Q.; Niederkorn, J.Y.; Stern, M.E.; Pflugfelder, S.C.; et al. CD8+ cells regulate the T helper-17 re-sponse in an experimental murine model of Sjogren syndrome. Mucosal Immunol. 2014, 7, 417–427. [Google Scholar] [CrossRef]
- Shiboski, C.H.; Shiboski, S.C.; Seror, R.; Criswell, L.A.; Labetoulle, M.; Lietman, T.M.; Rasmussen, A.; Scofield, H.; Vitali, C.; Bowman, S.J.; et al. ACR-EULAR Classification Criteria for primary Sjögren’s Syndrome: A Consensus and Data-Driven Methodology Involving Three International Patient Cohorts. Arthritis Rheumatol. 2017, 69, 35–45. [Google Scholar] [CrossRef]
- Seror, R.; Bowman, S.J.; Brito-Zeron, P.; Theander, E.; Bootsma, H.; Tzioufas, A.; Gottenberg, J.-E.; Ramos-Casals, M.; Dörner, T.; Ravaud, P.; et al. EULAR Sjogren’s syndrome disease activity index (ESSDAI): A user guide. RMD Open 2015, 1, e000022. [Google Scholar] [CrossRef] [PubMed]
- Seror, R.; Ravaud, P.; Mariette, X.; Bootsma, H.; Theander, E.; Hansen, A.; Ramos-Casals, M.; Dörner, T.; Bombardieri, S.; Hachulla, E.; et al. EULAR Sjögren’s Syndrome Patient Reported Index (ESSPRI): Development of a consensus patient index for primary Sjögren’s syndrome. Ann. Rheum. Dis. 2011, 70, 968–972. [Google Scholar] [CrossRef]
- Navazesh, M.; Kumar, S.K. Measuring salivary flow. J. Am. Dent. Assoc. 2008, 139, 35S–40S. [Google Scholar] [CrossRef]
- Kudryavtsev, I.V.; Arsentieva, N.A.; Korobova, Z.R.; Isakov, D.V.; Rubinstein, A.A.; Batsunov, O.K.; Khamitova, I.V.; Kuznetsova, R.N.; Savin, T.V.; Akisheva, T.V.; et al. Heterogenous CD8+ T Cell Maturation and ‘Polarization’ in Acute and Convalescent COVID-19 Patients. Viruses 2022, 14, 1906. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, O.; Golovkin, A.; Zaikova, E.; Aquino, A.; Bezrukikh, V.; Melnik, O.; Vasilieva, E.; Karonova, T.; Kudryavtsev, I.; Shlyakhto, E. Cytokine Storm Signature in Patients with Moderate and Severe COVID-19. Int. J. Mol. Sci. 2022, 23, 8879. [Google Scholar] [CrossRef]
- Sallusto, F.; Lenig, D.; Förster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing po-tentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef]
- Romero, P.; Zippelius, A.; Kurth, I.; Pittet, M.J.; Touvrey, C.; Iancu, E.M.; Corthesy, P.; Devevre, E.; Speiser, D.E.; Rufer, N. Four Functionally Distinct Populations of Human Effector-Memory CD8+ T Lymphocytes. Perspect. Surg. 2007, 178, 4112–4119. [Google Scholar] [CrossRef]
- Rufer, N.; Zippelius, A.; Batard, P.; Pittet, M.J.; Kurth, I.; Corthesy, P.; Cerottini, J.-C.; Leyvraz, S.; Roosnek, E.; Nabholz, M.; et al. Ex Vivo characterization of human CD8+ T subsets with distinct replicative history and partial effector functions. Blood 2003, 102, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Larbi, A.; Derhovanessian, E.; Özcelik, D.; Naumova, E.; Pawelec, G. Multiparameter flow cytometric analysis of CD4 and CD8 T cell subsets in young and old people. Immun. Ageing 2008, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Loyal, L.; Warth, S.; Jürchott, K.; Mölder, F.; Nikolaou, C.; Babel, N.; Nienen, M.; Durlanik, S.; Stark, R.; Kruse, B.; et al. SLAMF7 and IL-6R define distinct cytotoxic versus helper memory CD8+ T cells. Nat. Commun. 2020, 11, 6357. [Google Scholar] [CrossRef] [PubMed]
- Trombke, J.; Loyal, L.; Braun, J.; Pleyer, U.; Thiel, A.; Pohlmann, D. Analysis of peripheral inflammatory T cell subsets and their effector function in patients with Birdshot Retinochoroiditis. Sci. Rep. 2021, 11, 8604. [Google Scholar] [CrossRef] [PubMed]
- Gschwandtner, M.; Derler, R.; Midwood, K.S. More Than Just Attractive: How CCL2 Influences Myeloid Cell Behavior beyond Chemotaxis. Front. Immunol. 2019, 10, 2759. [Google Scholar] [CrossRef]
- Chang, T.-T.; Chen, C.; Chen, J.-W. CCL7 as a novel inflammatory mediator in cardiovascular disease, diabetes mellitus, and kidney disease. Cardiovasc. Diabetol. 2022, 21, 185. [Google Scholar] [CrossRef]
- Schaller, T.H.; Batich, K.A.; Suryadevara, C.M.; Desai, R.; Sampson, J.H. Chemokines as adjuvants for immunotherapy: Implications for immune activation with CCL3. Expert. Rev. Clin. Immunol. 2017, 13, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, F.; Romagnani, C.; Romagnani, S. The 3 major types of innate and adaptive cell-mediated effector immunity. J. Allergy Clin. Immunol. 2015, 135, 626–635. [Google Scholar] [CrossRef]
- Martin-Gutierrez, L.; Peng, J.; Thompson, N.L.; Robinson, G.A.; Naja, M.; Peckham, H.; Wu, W.; J’bari, H.; Ahwireng, N.; Waddington, K.E.; et al. Stratification of Patients with Sjögren’s Syndrome and Patients with Systemic Lupus Erythematosus According to Two Shared Immune Cell Signatures, with Potential Therapeutic Implications. Arthritis Rheumatol. 2021, 73, 1626–1637. [Google Scholar] [CrossRef]
- Narkeviciute, I.; Sudzius, G.; Mieliauskaite, D.; Mackiewicz, Z.; Butrimiene, I.; Viliene, R.; Dumalakiene, I. Are cytotoxic effector cells changes in peripheral blood of patients with Sjögren’s syndrome related to persistent virus infection: Suggestions and conun-drums. Cell Immunol. 2016, 310, 123–130. [Google Scholar] [CrossRef]
- Sudzius, G.; Mieliauskaite, D.; Siaurys, A.; Viliene, R.; Butrimiene, I.; Characiejus, D.; Dumalakiene, I. Distribution of Peripheral Lymphocyte Populations in Primary Sjögren’s Syndrome Patients. J. Immunol. Res. 2015, 2015, 854706. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, Y.; Wang, P.; Wang, Y.; Feng, Y.; Zhang, Y.; Wu, Z. Alterations of CD8+ T cells in the blood and salivary glands of patients with primary Sjögren’s syndrome. Clin. Rheumatol. 2023, 42, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Murata, O.; Suzuki, K.; Takeuchi, T. Thymus variants on imaging of patients with primary Sjögren’s syndrome and polymyo-sitis/dermatomyositis: Clinical and immunological significance. Immunol. Med. 2023, 46, 25–31. [Google Scholar] [CrossRef]
- Kobayashi, H.; Ozeki, Y.; Aida, S. Pulmonary and thymic lymphoid hyperplasia in primary Sjögren’s syndrome. Jpn. J. Radiol. 2009, 27, 107–110. [Google Scholar] [CrossRef]
- Kondo, K.; Miyoshi, T.; Sakiyama, S.; Shimosato, Y.; Monden, Y. Multilocular thymic cyst associated with Sjögren’s syndrome. Ann. Thorac. Surg. 2001, 72, 1367–1369. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Cai, H.; Li, Y.; Cui, Y. Thymic hyperplasia associated with primary Sjogren’s syndrome cured by thymectomy. J. Thorac. Dis. 2017, 9, E130–E132. [Google Scholar] [CrossRef] [PubMed]
- Minato, H.; Kinoshita, E.; Nakada, S.; Nojima, T.; Tanaka, M.; Usuda, K.; Sagawa, M.; Iwao, H.; Tanaka, M.; Doai, M.; et al. Thymic lymphoid hyperplasia with multilocular thymic cysts diagnosed before the Sjögren syndrome diagnosis. Diagn. Pathol. 2015, 10, 103. [Google Scholar] [CrossRef]
- Izumi, H.; Nobukawa, B.; Takahashi, K.; Kumasaka, T.; Miyamoto, H.; Yamazaki, A.; Sonobe, S.; Uekusa, T.; Suda, K. Multilocular thymic cyst associated with follicular hyperplasia: Clinicopathologic study of 4 resected cases. Hum. Pathol. 2005, 36, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Nizharadze, T.; Becker, N.B.; Höfer, T. Quantitating CD8+ T cell memory development. Trends Immunol. 2023, 44, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Mittrücker, H.-W.; Visekruna, A.; Huber, M. Heterogeneity in the Differentiation and Function of CD8+ T Cells. Arch. Immunol. Ther. Exp. 2014, 62, 449–458. [Google Scholar] [CrossRef]
- Zhou, J.; Kawai, T.; Yu, Q. Pathogenic role of endogenous TNF-α in the development of Sjögren’s-like si-aladenitis and secretory dysfunction in nonobese diabetic mice. Lab. Investig. 2017, 97, 458–467. [Google Scholar] [CrossRef]
- Moutsopoulos, N.M.; Katsifis, G.E.; Angelov, N.; Leakan, R.A.; Sankar, V.; Pillemer, S.; Wahl, S.M. Lack of efficacy of etanercept in Sjögren syndrome correlates with failed suppression of tumour necrosis factor alpha and systemic immune activation. Ann. Rheum. Dis. 2008, 67, 1437–1443. [Google Scholar] [CrossRef]
- Yoon, K.-C.; Park, C.-S.; You, I.-C.; Choi, H.-J.; Lee, K.-H.; Im, S.-K.; Park, H.-Y.; Pflugfelder, S.C. Expression of CXCL9, -10, -11, and CXCR3 in the Tear Film and Ocular Surface of Patients with Dry Eye Syndrome. Investig. Ophthalmol. Vis. Sci. 2010, 51, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Allo, V.C.M.; Hauk, V.; Sarbia, N.; Pinto, N.A.; Croci, D.O.; Dalotto-Moreno, T.; Morales, R.M.; Gatto, S.G.; Cocco, M.N.M.; Stupirski, J.C.; et al. Suppression of age-related salivary gland autoimmunity by glycosylation-dependent galectin-1-driven immune inhibitory circuits. Proc. Natl. Acad. Sci. USA 2020, 117, 6630–6639. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.S.; Ohashi, P.S. The Roles of CD8+ T Cell Subsets in Antitumor Immunity. Trends Cell Biol. 2020, 30, 695–704. [Google Scholar] [CrossRef]
- Cho, B.A.; Sim, J.H.; Park, J.A.; Kim, H.W.; Yoo, W.H.; Lee, S.H.; Lee, D.S.; Kang, J.S.; Hwang, Y.I.; Lee, W.J.; et al. Char-acterization of effector memory CD8+ T cells in the synovial fluid of rheumatoid arthritis. J. Clin. Immunol. 2012, 32, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.; Heink, S.; Pagenstecher, A.; Reinhard, K.; Ritter, J.; Visekruna, A.; Guralnik, A.; Bollig, N.; Jeltsch, K.; Heinemann, C.; et al. IL-17A secretion by CD8+ T cells supports Th17-mediated autoimmune encephalomyelitis. J. Clin. Investig. 2013, 123, 247–260. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sjögren’s Syndrome (n = 34) | Control Subjects (n = 34) | |
|---|---|---|
| Male/female ratio | 2/32 | 3/31 |
| Age, years, median (IQR) | 46 [41; 55] | 44 [39; 56] |
| Disease duration (years), median (IQR) | 8 [2.5; 11] | |
| ESSDAI, median (IQR) | 5 [2; 9] | |
| ESSPRI, median (IQR) | 6.7 [5.6; 7.3] | |
| Clinical manifestations/ESSDAI domain (percentage) | ||
| Constitutional domain | 5 (14.7) | |
| Lymphadenopathy and lymphoma domain | 6 (17.7) | |
| Glandular domain | 5 (14.7) | |
| Articular domain | 17 (50) | |
| Cutaneous domain | 5 (14.7) | |
| Pulmonary domain | 2 (5.8) | |
| Renal domain | 0 | |
| Muscular domain | 2 (5.8) | |
| Peripheral nervous system domain | 4 (11.6) | |
| Central nervous system domain | 0 | |
| Haematological domain | 14 (41.2) | |
| Biological domain Schirmer test, mm/5 min., median (IQR) Unstimulated sialometry, mL/15 min., median (IQR) | 3 (9.7) 3 [1; 9] 1.5 [1; 3] | |
| Laboratory manifestations (percentage) | ||
| Complement C3 levels below the lower limit of normal value | 6 (17.6) | |
| Complement C4 levels below the lower limit of normal value | 3 (8.7) | |
| Anti-DNA Ab | 4 (11.8) | |
| Rheumathoid factor | 10 (29.4) | |
| Ro-52 Ab | 20 (58.8) | |
| SSA Ab | 25 (73.5) | |
| SSB Ab | 10 (29.4) | |
| CENT B Ab | 2 (5.8) | |
| Medications | 28 (82.4) | |
| Prednisone dose, mg/day, median (IQR) | 3.75 [0.0; 10.0] | |
| Hydroxychloroquine (percentage) | 23 (67.7) | |
| Azathioprine (percentage) | 1 (2.9) | |
| Cyclosporine (percentage) | 1 (2.9) | |
| Mycophenolate mofetil (percentage) | 1 (2.9) | |
| Leflunomide (percentage) | 2 (5.8) | |
| Methotrexate (percentage) | 5 (14.7) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudryavtsev, I.; Benevolenskaya, S.; Serebriakova, M.; Grigor’yeva, I.; Kuvardin, E.; Rubinstein, A.; Golovkin, A.; Kalinina, O.; Zaikova, E.; Lapin, S.; et al. Circulating CD8+ T Cell Subsets in Primary Sjögren’s Syndrome. Biomedicines 2023, 11, 2778. https://doi.org/10.3390/biomedicines11102778
Kudryavtsev I, Benevolenskaya S, Serebriakova M, Grigor’yeva I, Kuvardin E, Rubinstein A, Golovkin A, Kalinina O, Zaikova E, Lapin S, et al. Circulating CD8+ T Cell Subsets in Primary Sjögren’s Syndrome. Biomedicines. 2023; 11(10):2778. https://doi.org/10.3390/biomedicines11102778
Chicago/Turabian StyleKudryavtsev, Igor, Stanislava Benevolenskaya, Maria Serebriakova, Irina Grigor’yeva, Evgeniy Kuvardin, Artem Rubinstein, Alexey Golovkin, Olga Kalinina, Ekaterina Zaikova, Sergey Lapin, and et al. 2023. "Circulating CD8+ T Cell Subsets in Primary Sjögren’s Syndrome" Biomedicines 11, no. 10: 2778. https://doi.org/10.3390/biomedicines11102778