
LAG-3 Inhibitors: Novel Immune Checkpoint Inhibitors Changing the Landscape of Immunotherapy

Abstract
:1. Introduction
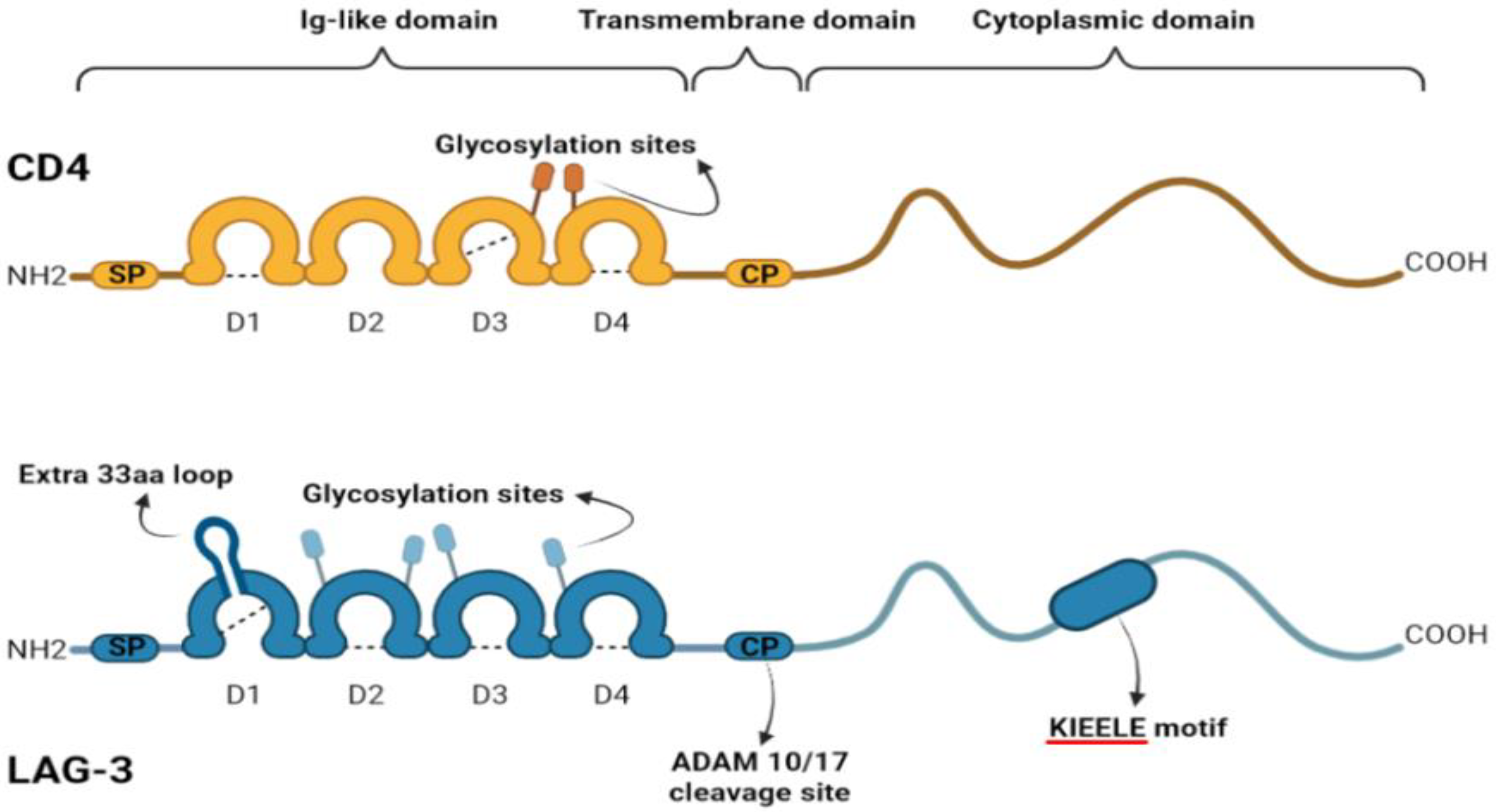
2. LAG-3 Structure and Regulation
3. Expression of LAG-3 on Immune Cells
4. Expression of LAG-3 on Tumor Cells
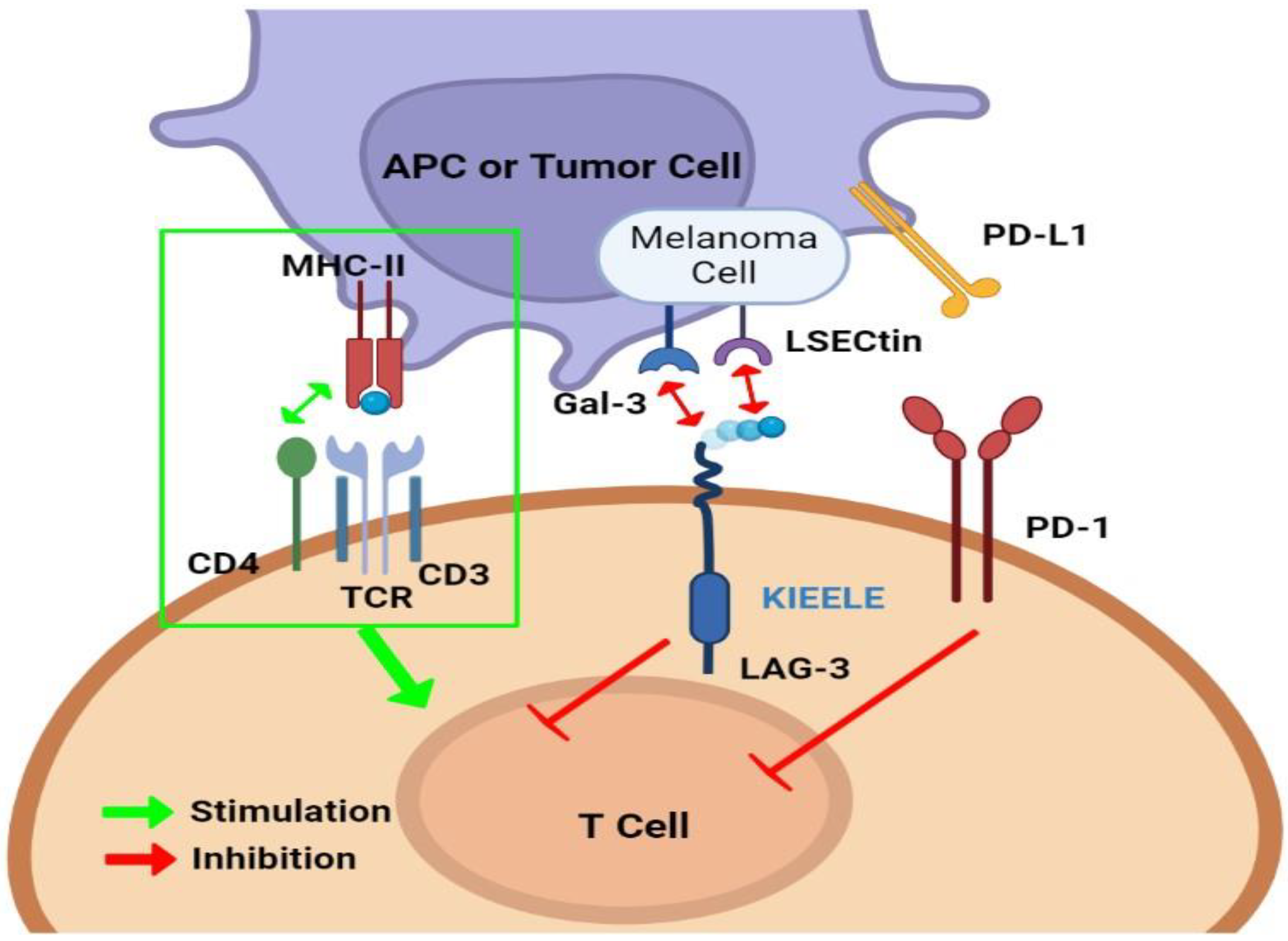
5. LAG-3 and Other Immune Checkpoints
6. Anti-LAG-3 Immunotherapy
6.1. LAG-3 Fusion Proteins
6.2. Anti-LAG3 Monoclonal Antibodies
6.3. Anti-LAG3 Bispecific Antibodies
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, N.M.; Simon, M.C. The Tumor Microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef] [PubMed]
- Son, B.; Lee, S.; Youn, H.; Kim, E.; Kim, W.; Youn, B. The Role of Tumor Microenvironment in Therapeutic Resistance. Oncotarget 2017, 8, 3933–3945. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Subramanian, S. Intrinsic Resistance of Solid Tumors to Immune Checkpoint Blockade Therapy. Cancer Res. 2017, 77, 817–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valiullina, A.K.; Zmievskaya, E.A.; Ganeeva, I.A.; Zhuravleva, M.N.; Garanina, E.E.; Rizvanov, A.A.; Petukhov, A.V.; Bulatov, E.R. Evaluation of CAR-T Cells’ Cytotoxicity against Modified Solid Tumor Cell Lines. Biomedicines 2023, 11, 626. [Google Scholar] [CrossRef]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. AAPS J. 2021, 23, 39. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Rivard, C.J.; Rozeboom, L.; Yu, H.; Ellison, K.; Kowalewski, A.; Zhou, C.; Hirsch, F.R. Lymphocyte-Activation Gene-3, an Important Immune Checkpoint in Cancer. Cancer Sci. 2016, 107, 1193–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, L.; Zhang, X.; Chen, F.; Pan, Q.; Phiphatwatchara, P.; Zeng, Y.; Chen, H. The Promising Immune Checkpoint LAG-3: From Tumor Microenvironment to Cancer Immunotherapy. Genes Cancer 2018, 9, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Zhu, C.; Kuchroo, V.K. Tim-3 and Its Role in Regulating Anti-Tumor Immunity. Immunol. Rev. 2017, 276, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Villarroel-Espindola, F.; Yu, X.; Datar, I.; Mani, N.; Sanmamed, M.; Velcheti, V.; Syrigos, K.; Toki, M.; Zhao, H.; Chen, L.; et al. Spatially Resolved and Quantitative Analysis of VISTA/PD-1H as a Novel Immunotherapy Target in Human Non-Small Cell Lung Cancer. Clin. Cancer Res. 2018, 24, 1562–1573. [Google Scholar] [CrossRef] [Green Version]
- Workman, C.J.; Dugger, K.J.; Vignali, D.A.A. Cutting Edge: Molecular Analysis of the Negative Regulatory Function of Lymphocyte Activation Gene-3. J. Immunol. 2002, 169, 5392–5395. [Google Scholar] [CrossRef] [Green Version]
- Andrews, L.P.; Marciscano, A.E.; Drake, C.G.; Vignali, D.A.A. LAG3 (CD223) as a Cancer Immunotherapy Target. Immunol. Rev. 2017, 276, 80–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruhashi, T.; Sugiura, D.; Okazaki, I.-M.; Okazaki, T. LAG-3: From Molecular Functions to Clinical Applications. J. Immunother. Cancer 2020, 8, e001014. [Google Scholar] [CrossRef]
- Triebel, F.; Jitsukawa, S.; Baixeras, E.; Roman-Roman, S.; Genevee, C.; Viegas-Pequignot, E.; Hercend, T. LAG-3, a Novel Lymphocyte Activation Gene Closely Related to CD4. J. Exp. Med. 1990, 171, 1393–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huard, B.; Mastrangeli, R.; Prigent, P.; Bruniquel, D.; Donini, S.; El-Tayar, N.; Maigret, B.; Dréano, M.; Triebel, F. Characterization of the Major Histocompatibility Complex Class II Binding Site on LAG-3 Protein. Proc. Natl. Acad. Sci. USA 1997, 94, 5744–5749. [Google Scholar] [CrossRef]
- Andreae, S.; Buisson, S.; Triebel, F. MHC Class II Signal Transduction in Human Dendritic Cells Induced by a Natural Ligand, the LAG-3 Protein (CD223). Blood 2003, 102, 2130–2137. [Google Scholar] [CrossRef] [Green Version]
- Hannier, S.; Tournier, M.; Bismuth, G.; Triebel, F. CD3/TCR Complex-Associated Lymphocyte Activation Gene-3 Molecules Inhibit CD3/TCR Signaling. J. Immunol. 1998, 161, 4058–4065. [Google Scholar] [CrossRef] [PubMed]
- Dumic, J.; Dabelic, S.; Flögel, M. Galectin-3: An Open-Ended Story. Biochim. Biophys. Acta 2006, 1760, 616–635. [Google Scholar] [CrossRef]
- Liu, W.; Tang, L.; Zhang, G.; Wei, H.; Cui, Y.; Guo, L.; Gou, Z.; Chen, X.; Jiang, D.; Zhu, Y.; et al. Characterization of a Novel C-Type Lectin-like Gene, LSECtin: Demonstration of Carbohydrate Binding and Expression in Sinusoidal Endothelial Cells of Liver and Lymph Node. J. Biol. Chem. 2004, 279, 18748–18758. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, M.V.; Drake, C.G. LAG-3 in Cancer Immunotherapy. Curr. Top. Microbiol. Immunol. 2011, 344, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Workman, C.J.; Cauley, L.S.; Kim, I.-J.; Blackman, M.A.; Woodland, D.L.; Vignali, D.A.A. Lymphocyte Activation Gene-3 (CD223) Regulates the Size of the Expanding T Cell Population Following Antigen Activation in Vivo. J. Immunol. 2004, 172, 5450–5455. [Google Scholar] [CrossRef]
- Workman, C.J.; Vignali, D.A.A. The CD4-Related Molecule, LAG-3 (CD223), Regulates the Expansion of Activated T Cells. Eur. J. Immunol. 2003, 33, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.M.; Brodsky, M.H.; Irving, B.A.; Levin, S.D.; Perlmutter, R.M.; Littman, D.R. Interaction of the Unique N-Terminal Region of Tyrosine Kinase P56lck with Cytoplasmic Domains of CD4 and CD8 Is Mediated by Cysteine Motifs. Cell 1990, 60, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.S.; Chalupny, J.; Whitney, J.A.; Hammond, C.; Amrein, K.E.; Kavathas, P.; Sefton, B.M.; Rose, J.K. Short Related Sequences in the Cytoplasmic Domains of CD4 and CD8 Mediate Binding to the Amino-Terminal Domain of the P56lck Tyrosine Protein Kinase. Mol. Cell. Biol. 1990, 10, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, Y.; Forbes, K.; Vignali, K.M.; Heale, B.S.; Saftig, P.; Hartmann, D.; Black, R.A.; Rossi, J.J.; Blobel, C.P.; et al. Metalloproteases Regulate T-Cell Proliferation and Effector Function via LAG-3. EMBO J. 2007, 26, 494–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, N.; Szlasa, W.; Jonderko, L.; Oślizło, M.; Kunachowicz, D.; Kulbacka, J.; Karłowicz-Bodalska, K. LAG-3 as a Potent Target for Novel Anticancer Therapies of a Wide Range of Tumors. Int. J. Mol. Sci. 2022, 23, 9958. [Google Scholar] [CrossRef]
- Annunziato, F.; Manetti, R.; Cosmi, L.; Galli, G.; Heusser, C.H.; Romagnani, S.; Maggi, E. Opposite Role for Interleukin-4 and Interferon-Gamma on CD30 and Lymphocyte Activation Gene-3 (LAG-3) Expression by Activated Naive T Cells. Eur. J. Immunol. 1997, 27, 2239–2244. [Google Scholar] [CrossRef]
- Bruniquel, D.; Borie, N.; Hannier, S.; Triebel, F. Regulation of Expression of the Human Lymphocyte Activation Gene-3 (LAG-3) Molecule, a Ligand for MHC Class II. Immunogenetics 1998, 48, 116–124. [Google Scholar] [CrossRef]
- Annunziato, F.; Manetti, R.; Tomasévic, I.; Guidizi, M.G.; Biagiotti, R.; Giannò, V.; Germano, P.; Mavilia, C.; Maggi, E.; Romagnani, S. Expression and Release of LAG-3-Encoded Protein by Human CD4+ T Cells Are Associated with IFN-Gamma Production. FASEB J. 1996, 10, 769–776. [Google Scholar] [CrossRef]
- Saleh, R.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Role of Epigenetic Modifications in Inhibitory Immune Checkpoints in Cancer Development and Progression. Front. Immunol. 2020, 11, 1469. [Google Scholar] [CrossRef]
- Goltz, D.; Gevensleben, H.; Dietrich, J.; Ellinger, J.; Landsberg, J.; Kristiansen, G.; Dietrich, D. Promoter Methylation of the Immune Checkpoint Receptor PD-1 (PDCD1) Is an Independent Prognostic Biomarker for Biochemical Recurrence-Free Survival in Prostate Cancer Patients Following Radical Prostatectomy. Oncoimmunology 2016, 5, e1221555. [Google Scholar] [CrossRef] [Green Version]
- Goltz, D.; Gevensleben, H.; Grünen, S.; Dietrich, J.; Kristiansen, G.; Landsberg, J.; Dietrich, D. PD-L1 (CD274) Promoter Methylation Predicts Survival in Patients with Acute Myeloid Leukemia. Leukemia 2017, 31, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Goltz, D.; Gevensleben, H.; Dietrich, J.; Schroeck, F.; de Vos, L.; Droege, F.; Kristiansen, G.; Schroeck, A.; Landsberg, J.; Bootz, F.; et al. PDCD1 (PD-1) Promoter Methylation Predicts Outcome in Head and Neck Squamous Cell Carcinoma Patients. Oncotarget 2017, 8, 41011–41020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Lv, D.; Cai, C.; Zhao, Z.; Wang, M.; Chen, W.; Liu, Y. A TP53-Associated Immune Prognostic Signature for the Prediction of Overall Survival and Therapeutic Responses in Muscle-Invasive Bladder Cancer. Front. Immunol. 2020, 11, 590618. [Google Scholar] [CrossRef]
- Querfeld, C.; Wu, X.; Sanchez, J.F.; Palmer, J.M.; Motevalli, A.; Zain, J.; Rosen, S.T. The miRNA Profile of Cutaneous T Cell Lymphoma Correlates with the Dysfunctional Immunophenotype of the Disease. Blood 2016, 128, 4132. [Google Scholar] [CrossRef]
- Laino, A.S.; Betts, B.C.; Veerapathran, A.; Dolgalev, I.; Sarnaik, A.; Quayle, S.N.; Jones, S.S.; Weber, J.S.; Woods, D.M. HDAC6 Selective Inhibition of Melanoma Patient T-Cells Augments Anti-Tumor Characteristics. J. Immunother. Cancer 2019, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.-R.; Li, N.; Bruno, T.C.; Forbes, K.; Brown, S.; Workman, C.; Drake, C.G.; Vignali, D.A.A. Differential Subcellular Localization of the Regulatory T-Cell Protein LAG-3 and the Coreceptor CD4. Eur. J. Immunol. 2010, 40, 1768–1777. [Google Scholar] [CrossRef] [Green Version]
- Shi, A.-P.; Tang, X.-Y.; Xiong, Y.-L.; Zheng, K.-F.; Liu, Y.-J.; Shi, X.-G.; Lv, Y.; Jiang, T.; Ma, N.; Zhao, J.-B. Immune Checkpoint LAG3 and Its Ligand FGL1 in Cancer. Front. Immunol. 2021, 12, 785091. [Google Scholar] [CrossRef]
- Bae, J.; Lee, S.J.; Park, C.-G.; Lee, Y.S.; Chun, T. Trafficking of LAG-3 to the Surface on Activated T Cells via Its Cytoplasmic Domain and Protein Kinase C Signaling. J. Immunol. 2014, 193, 3101–3112. [Google Scholar] [CrossRef] [Green Version]
- Martins, I.; Deshayes, F.; Baton, F.; Forget, A.; Ciechomska, I.; Sylla, K.; Aoudjit, F.; Charron, D.; Al-Daccak, R.; Alcaide-Loridan, C. Pathologic Expression of MHC Class II Is Driven by Mitogen-Activated Protein Kinases. Eur. J. Immunol. 2007, 37, 788–797. [Google Scholar] [CrossRef]
- Hemon, P.; Jean-Louis, F.; Ramgolam, K.; Brignone, C.; Viguier, M.; Bachelez, H.; Triebel, F.; Charron, D.; Aoudjit, F.; Al-Daccak, R.; et al. MHC Class II Engagement by Its Ligand LAG-3 (CD223) Contributes to Melanoma Resistance to Apoptosis. J. Immunol. 2011, 186, 5173–5183. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yu, H.; Rozeboom, L.; Rivard, C.J.; Ellison, K.; Dziadziuszko, R.; Suda, K.; Ren, S.; Wu, C.; Hou, L.; et al. LAG-3 Protein Expression in Non-Small Cell Lung Cancer and Its Relationship with PD-1/PD-L1 and Tumor-Infiltrating Lymphocytes. J. Thorac. Oncol. 2017, 12, 814–823. [Google Scholar] [CrossRef] [Green Version]
- Burugu, S.; Gao, D.; Leung, S.; Chia, S.K.; Nielsen, T.O. LAG-3+ Tumor Infiltrating Lymphocytes in Breast Cancer: Clinical Correlates and Association with PD-1/PD-L1+ Tumors. Ann. Oncol. 2017, 28, 2977–2984. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, J.; Gnjatic, S.; Mhawech-Fauceglia, P.; Beck, A.; Miller, A.; Tsuji, T.; Eppolito, C.; Qian, F.; Lele, S.; Shrikant, P.; et al. Tumor-Infiltrating NY-ESO-1-Specific CD8+ T Cells Are Negatively Regulated by LAG-3 and PD-1 in Human Ovarian Cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 7875–7880. [Google Scholar] [CrossRef]
- Li, F.-J.; Zhang, Y.; Jin, G.-X.; Yao, L.; Wu, D.-Q. Expression of LAG-3 Is Coincident with the Impaired Effector Function of HBV-Specific CD8(+) T Cell in HCC Patients. Immunol. Lett. 2013, 150, 116–122. [Google Scholar] [CrossRef]
- Giraldo, N.A.; Becht, E.; Pagès, F.; Skliris, G.; Verkarre, V.; Vano, Y.; Mejean, A.; Saint-Aubert, N.; Lacroix, L.; Natario, I.; et al. Orchestration and Prognostic Significance of Immune Checkpoints in the Microenvironment of Primary and Metastatic Renal Cell Cancer. Clin. Cancer Res. 2015, 21, 3031–3040. [Google Scholar] [CrossRef] [Green Version]
- Huard, B.; Tournier, M.; Hercend, T.; Triebel, F.; Faure, F. Lymphocyte-Activation Gene 3/Major Histocompatibility Complex Class II Interaction Modulates the Antigenic Response of CD4+ T Lymphocytes. Eur. J. Immunol. 1994, 24, 3216–3221. [Google Scholar] [CrossRef] [PubMed]
- Sittig, S.P.; Køllgaard, T.; Grønbæk, K.; Idorn, M.; Hennenlotter, J.; Stenzl, A.; Gouttefangeas, C.; Thor Straten, P. Clonal Expansion of Renal Cell Carcinoma-Infiltrating T Lymphocytes. Oncoimmunology 2013, 2, e26014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.; Meijers, R.; Xiong, Y.; Liu, J.H.; Sakihama, T.; Zhang, R.; Joachimiak, A.; Reinherz, E.L. Crystal Structure of the Human CD4 N-Terminal Two-Domain Fragment Complexed to a Class II MHC Molecule. Proc. Natl. Acad. Sci. USA 2001, 98, 10799–10804. [Google Scholar] [CrossRef]
- Farsam, V.; Hassan, Z.M.; Zavaran-Hosseini, A.; Noori, S.; Mahdavi, M.; Ranjbar, M. Antitumor and Immunomodulatory Properties of Artemether and Its Ability to Reduce CD4+ CD25+ FoxP3+ T Reg Cells in Vivo. Int. Immunopharmacol. 2011, 11, 1802–1808. [Google Scholar] [CrossRef]
- Park, H.J.; Kusnadi, A.; Lee, E.-J.; Kim, W.W.; Cho, B.C.; Lee, I.J.; Seong, J.; Ha, S.-J. Tumor-Infiltrating Regulatory T Cells Delineated by Upregulation of PD-1 and Inhibitory Receptors. Cell Immunol. 2012, 278, 76–83. [Google Scholar] [CrossRef]
- Huang, C.-T.; Workman, C.J.; Flies, D.; Pan, X.; Marson, A.L.; Zhou, G.; Hipkiss, E.L.; Ravi, S.; Kowalski, J.; Levitsky, H.I.; et al. Role of LAG-3 in Regulatory T Cells. Immunity 2004, 21, 503–513. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Dierich, A.; Benoist, C.; Mathis, D. Independent Modes of Natural Killing Distinguished in Mice Lacking Lag3. Science 1996, 272, 405–408. [Google Scholar] [CrossRef]
- Byun, H.-J.; Jung, W.-W.; Lee, D.-S.; Kim, S.; Kim, S.J.; Park, C.-G.; Chung, H.Y.; Chun, T. Proliferation of Activated CD1d-Restricted NKT Cells Is down-Modulated by Lymphocyte Activation Gene-3 Signaling via Cell Cycle Arrest in S Phase. Cell Biol. Int. 2007, 31, 257–262. [Google Scholar] [CrossRef]
- Szymczak-Workman, A.L.; Workman, C.J.; Vignali, D.A.A. Cutting Edge: Regulatory T Cells Do Not Require Stimulation through Their TCR to Suppress. J. Immunol. 2009, 182, 5188–5192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, S.D.; Shin, H.; Haining, W.N.; Zou, T.; Workman, C.J.; Polley, A.; Betts, M.R.; Freeman, G.J.; Vignali, D.A.A.; Wherry, E.J. Coregulation of CD8+ T Cell Exhaustion by Multiple Inhibitory Receptors during Chronic Viral Infection. Nat. Immunol. 2009, 10, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisielow, M.; Kisielow, J.; Capoferri-Sollami, G.; Karjalainen, K. Expression of Lymphocyte Activation Gene 3 (LAG-3) on B Cells Is Induced by T Cells. Eur. J. Immunol. 2005, 35, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Lino, A.C.; Dang, V.D.; Lampropoulou, V.; Welle, A.; Joedicke, J.; Pohar, J.; Simon, Q.; Thalmensi, J.; Baures, A.; Flühler, V.; et al. LAG-3 Inhibitory Receptor Expression Identifies Immunosuppressive Natural Regulatory Plasma Cells. Immunity 2018, 49, 120–133.e9. [Google Scholar] [CrossRef]
- Takaya, S.; Saito, H.; Ikeguchi, M. Upregulation of Immune Checkpoint Molecules, PD-1 and LAG-3, on CD4+ and CD8+ T Cells after Gastric Cancer Surgery. Yonago Acta Med. 2015, 58, 39–44. [Google Scholar] [PubMed]
- Chen, J.; Chen, Z. The Effect of Immune Microenvironment on the Progression and Prognosis of Colorectal Cancer. Med. Oncol. 2014, 31, 82. [Google Scholar] [CrossRef]
- Yang, Z.-Z.; Kim, H.J.; Villasboas, J.C.; Chen, Y.-P.; Price-Troska, T.; Jalali, S.; Wilson, M.; Novak, A.J.; Ansell, S.M. Expression of LAG-3 Defines Exhaustion of Intratumoral PD-1+ T Cells and Correlates with Poor Outcome in Follicular Lymphoma. Oncotarget 2017, 8, 61425–61439. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.-W.; Mao, L.; Yu, G.-T.; Bu, L.-L.; Ma, S.-R.; Liu, B.; Gutkind, J.S.; Kulkarni, A.B.; Zhang, W.-F.; Sun, Z.-J. LAG-3 Confers Poor Prognosis and Its Blockade Reshapes Antitumor Response in Head and Neck Squamous Cell Carcinoma. Oncoimmunology 2016, 5, e1239005. [Google Scholar] [CrossRef] [Green Version]
- Norström, M.M.; Rådestad, E.; Sundberg, B.; Mattsson, J.; Henningsohn, L.; Levitsky, V.; Uhlin, M. Progression of Benign Prostatic Hyperplasia Is Associated with Pro-Inflammatory Mediators and Chronic Activation of Prostate-Infiltrating Lymphocytes. Oncotarget 2016, 7, 23581–23593. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Liu, Z.; Rangelova, E.; Poiret, T.; Ambati, A.; Rane, L.; Xie, S.; Verbeke, C.; Dodoo, E.; Del Chiaro, M.; et al. Expansion of Tumor-Reactive T Cells from Patients with Pancreatic Cancer. J. Immunother. 2016, 39, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanik, E.L.; Kaunitz, G.J.; Cottrell, T.R.; Succaria, F.; McMiller, T.L.; Ascierto, M.L.; Esandrio, J.; Xu, H.; Ogurtsova, A.; Cornish, T.; et al. Association of HIV Status with Local Immune Response to Anal Squamous Cell Carcinoma: Implications for Immunotherapy. JAMA Oncol. 2017, 3, 974–978. [Google Scholar] [CrossRef]
- Donia, M.; Andersen, R.; Kjeldsen, J.W.; Fagone, P.; Munir, S.; Nicoletti, F.; Andersen, M.H.; Thor Straten, P.; Svane, I.M. Aberrant Expression of MHC Class II in Melanoma Attracts Inflammatory Tumor-Specific CD4+ T- Cells, Which Dampen CD8+ T-Cell Antitumor Reactivity. Cancer Res. 2015, 75, 3747–3759. [Google Scholar] [CrossRef] [Green Version]
- Camisaschi, C.; De Filippo, A.; Beretta, V.; Vergani, B.; Villa, A.; Vergani, E.; Santinami, M.; Cabras, A.D.; Arienti, F.; Triebel, F.; et al. Alternative Activation of Human Plasmacytoid DCs in Vitro and in Melanoma Lesions: Involvement of LAG-3. J. Investig. Dermatol. 2014, 134, 1893–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, M.; Tomaszowski, K.-H.; Marisetty, A.; Kong, L.-Y.; Wei, J.; Duna, M.; Blumberg, K.; Ji, X.; Jacobs, C.; Fuller, G.N.; et al. Profiling of Patients with Glioma Reveals the Dominant Immunosuppressive Axis Is Refractory to Immune Function Restoration. JCI Insight 2020, 5, e134386. [Google Scholar] [CrossRef] [PubMed]
- Mair, M.J.; Kiesel, B.; Feldmann, K.; Widhalm, G.; Dieckmann, K.; Wöhrer, A.; Müllauer, L.; Preusser, M.; Berghoff, A.S. LAG-3 Expression in the Inflammatory Microenvironment of Glioma. J. Neurooncol. 2021, 152, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Du, Q.; Jin, J.; Wei, Y.; Lu, Y.; Li, Q. LAG3 and Its Emerging Role in Cancer Immunotherapy. Clin. Transl. Med. 2021, 11, e365. [Google Scholar] [CrossRef]
- Harris-Bookman, S.; Mathios, D.; Martin, A.M.; Xia, Y.; Kim, E.; Xu, H.; Belcaid, Z.; Polanczyk, M.; Barberi, T.; Theodros, D.; et al. Expression of LAG-3 and Efficacy of Combination Treatment with Anti-LAG-3 and Anti-PD-1 Monoclonal Antibodies in Glioblastoma. Int. J. Cancer 2018, 143, 3201–3208. [Google Scholar] [CrossRef] [Green Version]
- Botticelli, A.; Zizzari, I.G.; Scagnoli, S.; Pomati, G.; Strigari, L.; Cirillo, A.; Cerbelli, B.; Di Filippo, A.; Napoletano, C.; Scirocchi, F.; et al. The Role of Soluble LAG3 and Soluble Immune Checkpoints Profile in Advanced Head and Neck Cancer: A Pilot Study. J. Pers. Med. 2021, 11, 651. [Google Scholar] [CrossRef]
- Jie, H.-B.; Gildener-Leapman, N.; Li, J.; Srivastava, R.M.; Gibson, S.P.; Whiteside, T.L.; Ferris, R.L. Intratumoral Regulatory T Cells Upregulate Immunosuppressive Molecules in Head and Neck Cancer Patients. Br. J. Cancer 2013, 109, 2629–2635. [Google Scholar] [CrossRef]
- Luo, Y.; Yang, Y.-C.; Shen, C.-K.; Ma, B.; Xu, W.-B.; Wang, Q.-F.; Zhang, Y.; Liao, T.; Wei, W.-J.; Wang, Y. Immune Checkpoint Protein Expression Defines the Prognosis of Advanced Thyroid Carcinoma. Front. Endocrinol. (Lausanne) 2022, 13, 859013. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wei, X.; Pan, Y.; Xu, J.; Si, Y.; Min, Z.; Yu, B. A New Risk Factor Indicator for Papillary Thyroid Cancer Based on Immune Infiltration. Cell Death Dis. 2021, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Giannini, R.; Moretti, S.; Ugolini, C.; Macerola, E.; Menicali, E.; Nucci, N.; Morelli, S.; Colella, R.; Mandarano, M.; Sidoni, A.; et al. Immune Profiling of Thyroid Carcinomas Suggests the Existence of Two Major Phenotypes: An ATC-Like and a PDTC-Like. J. Clin. Endocrinol. Metab. 2019, 104, 3557–3575. [Google Scholar] [CrossRef]
- Sun, H.; Dai, J.; Zhao, L.; Zhu, J.; Wang, H.; Chen, P.; Lu, H.; Chen, Q.; Zhang, Z. Lymphocyte Activation Gene-3 Is Associated with Programmed Death-Ligand 1 and Programmed Cell Death Protein 1 in Small Cell Lung Cancer. Ann. Transl. Med. 2021, 9, 1468. [Google Scholar] [CrossRef]
- Wei, T.; Zhang, J.; Qin, Y.; Wu, Y.; Zhu, L.; Lu, L.; Tang, G.; Shen, Q. Increased Expression of Immunosuppressive Molecules on Intratumoral and Circulating Regulatory T Cells in Non-Small-Cell Lung Cancer Patients. Am. J. Cancer Res. 2015, 5, 2190–2201. [Google Scholar]
- Ma, Q.-Y.; Huang, D.-Y.; Zhang, H.-J.; Wang, S.; Chen, X.-F. Function and Regulation of LAG3 on CD4+CD25- T Cells in Non-Small Cell Lung Cancer. Exp. Cell Res. 2017, 360, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Getz, G.; Wheeler, D.A.; Mardis, E.R.; McLellan, M.D.; Cibulskis, K.; Sougnez, C.; Greulich, H.; Muzny, D.M.; Morgan, M.B.; et al. Somatic Mutations Affect Key Pathways in Lung Adenocarcinoma. Nature 2008, 455, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Datar, I.; Sanmamed, M.F.; Wang, J.; Henick, B.S.; Choi, J.; Badri, T.; Dong, W.; Mani, N.; Toki, M.; Mejías, L.D.; et al. Expression Analysis and Significance of PD-1, LAG-3, and TIM-3 in Human Non-Small Cell Lung Cancer Using Spatially Resolved and Multiparametric Single-Cell Analysis. Clin. Cancer Res. 2019, 25, 4663–4673. [Google Scholar] [CrossRef]
- Xiong, D.; Pan, J.; Yin, Y.; Jiang, H.; Szabo, E.; Lubet, R.A.; Wang, Y.; You, M. Novel Mutational Landscapes and Expression Signatures of Lung Squamous Cell Carcinoma. Oncotarget 2018, 9, 7424–7441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stovgaard, E.S.; Kümler, I.; List-Jensen, K.; Roslind, A.; Christensen, I.J.; Høgdall, E.; Nielsen, D.; Balslev, E. Prognostic and Clinicopathologic Associations of LAG-3 Expression in Triple-Negative Breast Cancer. Appl. Immunohistochem. Mol. Morphol. 2022, 30, 62–71. [Google Scholar] [CrossRef]
- Sobottka, B.; Moch, H.; Varga, Z. Differential PD-1/LAG-3 Expression and Immune Phenotypes in Metastatic Sites of Breast Cancer. Breast Cancer Res. 2021, 23, 4. [Google Scholar] [CrossRef]
- Du, H.; Yi, Z.; Wang, L.; Li, Z.; Niu, B.; Ren, G. The Co-Expression Characteristics of LAG3 and PD-1 on the T Cells of Patients with Breast Cancer Reveal a New Therapeutic Strategy. Int. Immunopharmacol. 2020, 78, 106113. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, M.K.; Lambley, E.; Duraiswamy, J.; Dua, U.; Smith, C.; Elliott, S.; Gill, D.; Marlton, P.; Seymour, J.; Khanna, R. Expression of LAG-3 by Tumor-Infiltrating Lymphocytes Is Coincident with the Suppression of Latent Membrane Antigen-Specific CD8+ T-Cell Function in Hodgkin Lymphoma Patients. Blood 2006, 108, 2280–2289. [Google Scholar] [CrossRef] [Green Version]
- El Halabi, L.; Adam, J.; Gravelle, P.; Marty, V.; Danu, A.; Lazarovici, J.; Ribrag, V.; Bosq, J.; Camara-Clayette, V.; Laurent, C.; et al. Expression of the Immune Checkpoint Regulators LAG-3 and TIM-3 in Classical Hodgkin Lymphoma. Clin. Lymphoma Myeloma Leuk. 2021, 21, 257–266.e3. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jilisihan, B.; Wang, W.; Tang, Y.; Keyoumu, S. Soluble LAG3 Acts as a Potential Prognostic Marker of Gastric Cancer and Its Positive Correlation with CD8+T Cell Frequency and Secretion of IL-12 and INF-γ in Peripheral Blood. Cancer Biomark. 2018, 23, 341–351. [Google Scholar] [CrossRef]
- Lv, K.; Li, R.; Cao, Y.; Gu, Y.; Liu, X.; He, X.; Jin, K.; Fang, H.; Fei, Y.; Shi, M.; et al. Lymphocyte-Activation Gene 3 Expression Associates with Poor Prognosis and Immunoevasive Contexture in Epstein-Barr Virus-Positive and MLH1-Defective Gastric Cancer Patients. Int. J. Cancer 2021, 148, 759–768. [Google Scholar] [CrossRef]
- Okazaki, T.; Okazaki, I.; Wang, J.; Sugiura, D.; Nakaki, F.; Yoshida, T.; Kato, Y.; Fagarasan, S.; Muramatsu, M.; Eto, T.; et al. PD-1 and LAG-3 Inhibitory Co-Receptors Act Synergistically to Prevent Autoimmunity in Mice. J. Exp. Med. 2011, 208, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Okagawa, T.; Konnai, S.; Deringer, J.R.; Ueti, M.W.; Scoles, G.A.; Murata, S.; Ohashi, K.; Brown, W.C. Cooperation of PD-1 and LAG-3 Contributes to T-Cell Exhaustion in Anaplasma Marginale-Infected Cattle. Infect. Immun. 2016, 84, 2779–2790. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.-R.; Turnis, M.E.; Goldberg, M.V.; Bankoti, J.; Selby, M.; Nirschl, C.J.; Bettini, M.L.; Gravano, D.M.; Vogel, P.; Liu, C.L.; et al. Immune Inhibitory Molecules LAG-3 and PD-1 Synergistically Regulate T-Cell Function to Promote Tumoral Immune Escape. Cancer Res. 2012, 72, 917–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.-Y.; Eppolito, C.; Lele, S.; Shrikant, P.; Matsuzaki, J.; Odunsi, K. LAG3 and PD1 Co-Inhibitory Molecules Collaborate to Limit CD8+ T Cell Signaling and Dampen Antitumor Immunity in a Murine Ovarian Cancer Model. Oncotarget 2015, 6, 27359–27377. [Google Scholar] [CrossRef]
- Dempke, W.C.M.; Fenchel, K.; Uciechowski, P.; Dale, S.P. Second- and Third-Generation Drugs for Immuno-Oncology Treatment—The More the Better? Eur. J. Cancer 2017, 74, 55–72. [Google Scholar] [CrossRef]
- Baumeister, S.H.; Freeman, G.J.; Dranoff, G.; Sharpe, A.H. Coinhibitory Pathways in Immunotherapy for Cancer. Annu. Rev. Immunol. 2016, 34, 539–573. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yang, P.; Zhou, H.; Li, B.; Huang, X.; Meng, Q.; Wang, L.; Kijlstra, A. CD4+CD25+Tregs Express an Increased LAG-3 and CTLA-4 in Anterior Chamber-Associated Immune Deviation. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Boor, P.P.C.; Metselaar, H.J.; de Jonge, S.; Mancham, S.; van der Laan, L.J.W.; Kwekkeboom, J. Human Plasmacytoid Dendritic Cells Induce CD8+ LAG-3+ Foxp3+ CTLA-4+ Regulatory T Cells That Suppress Allo-Reactive Memory T Cells. Eur. J. Immunol. 2011, 41, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Chung, Y.-H. Construction, and in Vitro and in Vivo Analyses of Tetravalent Immunoadhesins. J. Microbiol. Biotechnol. 2012, 22, 1066–1076. [Google Scholar] [CrossRef] [Green Version]
- Bjoern, J.; Lyngaa, R.; Andersen, R.; Hölmich, L.R.; Hadrup, S.R.; Donia, M.; Svane, I.M. Influence of Ipilimumab on Expanded Tumour Derived T Cells from Patients with Metastatic Melanoma. Oncotarget 2017, 8, 27062–27074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avice, M.N.; Sarfati, M.; Triebel, F.; Delespesse, G.; Demeure, C.E. Lymphocyte Activation Gene-3, a MHC Class II Ligand Expressed on Activated T Cells, Stimulates TNF-Alpha and IL-12 Production by Monocytes and Dendritic Cells. J. Immunol. 1999, 162, 2748–2753. [Google Scholar] [CrossRef]
- Tawbi, H.A.; Schadendorf, D.; Lipson, E.J.; Ascierto, P.A.; Matamala, L.; Castillo Gutiérrez, E.; Rutkowski, P.; Gogas, H.J.; Lao, C.D.; De Menezes, J.J.; et al. Relatlimab and Nivolumab versus Nivolumab in Untreated Advanced Melanoma. N. Engl. J. Med. 2022, 386, 24–34. [Google Scholar] [CrossRef]
- Moreau, P.; Garfall, A.L.; van de Donk, N.W.C.J.; Nahi, H.; San-Miguel, J.F.; Oriol, A.; Nooka, A.K.; Martin, T.; Rosinol, L.; Chari, A.; et al. Teclistamab in Relapsed or Refractory Multiple Myeloma. N. Engl. J. Med. 2022, 387, 495–505. [Google Scholar] [CrossRef]
- Dickinson, M.J.; Carlo-Stella, C.; Morschhauser, F.; Bachy, E.; Corradini, P.; Iacoboni, G.; Khan, C.; Wróbel, T.; Offner, F.; Trněný, M.; et al. Glofitamab for Relapsed or Refractory Diffuse Large B-Cell Lymphoma. N. Engl. J. Med. 2022, 387, 2220–2231. [Google Scholar] [CrossRef] [PubMed]
- Dirix, L.; Triebel, F. AIPAC: A Phase IIb Study of Eftilagimod Alpha (IMP321 or LAG-3Ig) Added to Weekly Paclitaxel in Patients with Metastatic Breast Cancer. Future Oncol. 2019, 15, 1963–1973. [Google Scholar] [CrossRef]
- Legat, A.; Maby-El Hajjami, H.; Baumgaertner, P.; Cagnon, L.; Abed Maillard, S.; Geldhof, C.; Iancu, E.M.; Lebon, L.; Guillaume, P.; Dojcinovic, D.; et al. Vaccination with LAG-3Ig (IMP321) and Peptides Induces Specific CD4 and CD8 T-Cell Responses in Metastatic Melanoma Patients—Report of a Phase I/IIa Clinical Trial. Clin. Cancer Res. 2016, 22, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Brignone, C.; Grygar, C.; Marcu, M.; Schäkel, K.; Triebel, F. A Soluble Form of Lymphocyte Activation Gene-3 (IMP321) Induces Activation of a Large Range of Human Effector Cytotoxic Cells. J. Immunol. 2007, 179, 4202–4211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forster, M.; Felip, E.; Doger, B.; Pousa, A.L.; Carcereny, E.; Bajaj, P.; Church, M.; Peguero, J.; Roxburgh, P.; Triebel, F. 927P Initial Results from a Phase II Study (TACTI-002) of Eftilagimod Alpha (Soluble LAG-3 Protein) and Pembrolizumab as 2nd Line Treatment for PD-L1 Unselected Metastatic Head and Neck Cancer Patients. Ann. Oncol. 2020, 31, S667. [Google Scholar] [CrossRef]
- Mauro, G. Eftilagimod Alpha Combination Demonstrates Durable Responses in Frontline NSCLC. Target. Oncol. 2023, 11, 39. [Google Scholar]
- Forster, M.; Krebs, M.; Majem, M.; Peguero, J.; Clay, T.; Felip, E.; Iams, W.; Roxburgh, P.; Doger, B.; Bajaj, P.; et al. EP08.01-109 TACTI-002: A Phase II Study of Eftilagimod Alpha (Soluble LAG-3) & Pembrolizumab in 2nd Line PD-1/PD-L1 Refractory Metastatic NSCLC. J. Thorac. Oncol. 2022, 17, S395. [Google Scholar] [CrossRef]
- Ascierto, P.A.; Lipson, E.J.; Dummer, R.; Larkin, J.; Long, G.V.; Sanborn, R.E.; Chiarion-Sileni, V.; Dréno, B.; Dalle, S.; Schadendorf, D.; et al. Nivolumab and Relatlimab in Patients with Advanced Melanoma That Had Progressed on Anti-Programmed Death-1/Programmed Death Ligand 1 Therapy: Results from the Phase I/IIa RELATIVITY-020 Trial. J. Clin. Oncol. 2023, 41, 2724–2735. [Google Scholar] [CrossRef]
- Amaria, R.N.; Postow, M.; Burton, E.M.; Tetzlaff, M.T.; Ross, M.I.; Torres-Cabala, C.; Glitza, I.C.; Duan, F.; Milton, D.R.; Busam, K.; et al. Neoadjuvant Relatlimab and Nivolumab in Resectable Melanoma. Nature 2022, 611, 155–160. [Google Scholar] [CrossRef]
- Johnson, N.A.; Lavie, D.; Borchmann, P.; Gregory, G.; Herrera, A.F.; Minuk, L.; Vucinic, V.; Armand, P.; Avigdor, A.; Gasiorowski, R.; et al. Favezelimab (Anti–LAG-3) plus Pembrolizumab in Patients with Anti–PD-1–Naive Relapsed or Refractory (R/R) Classical Hodgkin Lymphoma (cHL): An Open-Label Phase 1/2 Study. JCO 2022, 40, 7516. [Google Scholar] [CrossRef]
- Timmerman, J.; Lavie, D.; Johnson, N.A.; Avigdor, A.; Borchmann, P.; Andreadis, C.; Bazargan, A.; Gregory, G.; Keane, C.; Inna, T.; et al. Favezelimab (Anti–LAG-3) plus Pembrolizumab in Patients with Relapsed or Refractory (R/R) Classical Hodgkin Lymphoma (cHL) after Anti–PD-1 Treatment: An Open-Label Phase 1/2 Study. JCO 2022, 40, 7545. [Google Scholar] [CrossRef]
- Hamid, O.; Wang, D.; Kim, T.M.; Kim, S.-W.; Lakhani, N.J.; Johnson, M.L.; Groisberg, R.; Papadopoulos, K.P.; Kaczmar, J.M.; Middleton, M.R.; et al. Clinical Activity of Fianlimab (REGN3767), a Human Anti-LAG-3 Monoclonal Antibody, Combined with Cemiplimab (Anti-PD-1) in Patients (Pts) with Advanced Melanoma. JCO 2021, 39, 9515. [Google Scholar] [CrossRef]
- Ren, Z.; Guo, Y.; Bai, Y.; Ying, J.; Meng, Z.; Chen, Z.; Gu, S.; Zhang, J.; Liang, J.; Hou, X.; et al. Tebotelimab, a PD-1/LAG-3 Bispecific Antibody, in Patients with Advanced Hepatocellular Carcinoma Who Had Failed Prior Targeted Therapy and/or Immunotherapy: An Open-Label, Single-Arm, Phase 1/2 Dose-Escalation and Expansion Study. JCO 2023, 41, 578. [Google Scholar] [CrossRef]
- Rohrberg, K.S.; Garralda, E.; Calvo, E.; Garcia, V.M.; Guidi, M.; Kraus, D.G.; McIntyre, C.; Kao, H.; Deak, L.C.; Michielin, F.; et al. 745P Clinical Activity, Safety, and PK/PD from the First in Human Study (NP41300) of RO7247669, a PD1-LAG3 Bispecific Antibody. Ann. Oncol. 2022, 33, S884–S885. [Google Scholar] [CrossRef]
- Wang-Gillam, A.; Plambeck-Suess, S.; Goedegebuure, P.; Simon, P.O.; Mitchem, J.B.; Hornick, J.R.; Sorscher, S.; Picus, J.; Suresh, R.; Lockhart, A.C.; et al. A Phase I Study of IMP321 and Gemcitabine as the Front-Line Therapy in Patients with Advanced Pancreatic Adenocarcinoma. Investig. New Drugs 2013, 31, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, V.; Khattak, A.; Haydon, A.; Eastgate, M.; Roy, A.; Prithviraj, P.; Mueller, C.; Brignone, C.; Triebel, F. Eftilagimod Alpha, a Soluble Lymphocyte Activation Gene-3 (LAG-3) Protein plus Pembrolizumab in Patients with Metastatic Melanoma. J. Immunother. Cancer 2020, 8, e001681. [Google Scholar] [CrossRef]
- Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; González-Rodríguez, A.P.; Payer, Á.R.; González-García, E.; López-Soto, A.; Gonzalez, S. LAG-3 Blockade with Relatlimab (BMS-986016) Restores Anti-Leukemic Responses in Chronic Lymphocytic Leukemia. Cancers 2021, 13, 2112. [Google Scholar] [CrossRef]
- Schuler, M.H.H.; Cuppens, K.; Ploenes, T.; Vanbockrijck, M.; Wiesweg, M.; Darwiche, K.; Schramm, A.; Maes, B.; Hegedus, B.; Schildhaus, H.-U.; et al. LBA37 A Randomized, Multicentric Phase II Study of Preoperative Nivolumab plus Relatlimab or Nivolumab in Patients with Resectable Non-Small Cell Lung Cancer (NEOpredict-Lung). Ann. Oncol. 2022, 33, S1404. [Google Scholar] [CrossRef]
- Bhagwat, B.; Cherwinski, H.; Sathe, M.; Seghezzi, W.; McClanahan, T.K.; de Waal Malefyt, R.; Willingham, A. Establishment of Engineered Cell-Based Assays Mediating LAG3 and PD1 Immune Suppression Enables Potency Measurement of Blocking Antibodies and Assessment of Signal Transduction. J. Immunol. Methods 2018, 456, 7–14. [Google Scholar] [CrossRef]
- Garralda, E.; Sukari, A.; Lakhani, N.J.; Patnaik, A.; Lou, Y.; Im, S.-A.; Golan, T.; Geva, R.; Wermke, M.; de Miguel, M.; et al. A First-in-Human Study of the Anti-LAG-3 Antibody Favezelimab plus Pembrolizumab in Previously Treated, Advanced Microsatellite Stable Colorectal Cancer. ESMO Open 2022, 7, 100639. [Google Scholar] [CrossRef] [PubMed]
- Schöffski, P.; Tan, D.S.W.; Martín, M.; Ochoa-de-Olza, M.; Sarantopoulos, J.; Carvajal, R.D.; Kyi, C.; Esaki, T.; Prawira, A.; Akerley, W.; et al. Phase I/II Study of the LAG-3 Inhibitor Ieramilimab (LAG525) ± Anti-PD-1 Spartalizumab (PDR001) in Patients with Advanced Malignancies. J. Immunother. Cancer 2022, 10, e003776. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Asch, A.S.; Hamad, N.; Weickhardt, A.; Tomaszewska-Kiecana, M.; Dlugosz-Danecka, M.; Pylypenko, H.; Bahadur, S.; Ulahannan, S.; Koucheki, J.; et al. A Phase 1, Open-Label Study of MGD013, a Bispecific DART® Molecule Binding PD-1 and LAG-3 in Patients with Relapsed or Refractory Diffuse Large B-Cell Lymphoma. Blood 2020, 136, 21–22. [Google Scholar] [CrossRef]
- Patel, M.; Luke, J.; Hamilton, E.; Chmielowski, B.; Blumenschein, G.; Kindler, H.; Bahadur, S.; Santa-Maria, C.; Koucheki, J.; Sun, J.; et al. 313 A Phase 1 Evaluation of Tebotelimab, a Bispecific PD-1 x LAG-3 DART® Molecule, in Combination with Margetuximab in Patients with Advanced HER2+ Neoplasms. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Drug | Class | Treatment | Phase | Population | No of Pts | Outcome | AEs |
|---|---|---|---|---|---|---|---|
| Eftilagimod alpha (IMP321) [106] | LAG-3 fusion protein | Eftilagimod alpha + pembrolizumab | II | 2nd line PD-L1 unselected HNSCC | 18 | ORR: 39% | Cough (29%) Asthenia (24%) Dyspnea (18%) |
| Eftilagimod alpha (IMP321) [107] | LAG-3 fusion protein | Eftilagimod alpha + pembrolizumab | II | 1st line metastatic NSCLC | 110 | ORR (ITT): 40.4% ORR (TPS < 1%): 31% ORR (TPS 1–49%): 45% ORR (TPS > 50%): 55% | Pruritus (20%) Asthenia (19%) Rash (13%) Diarrhea (11%) |
| Eft-ilagimod alpha (IMP321) [108] | LAG-3 fusion protein | Eftilagimod alpha + pembrolizumab | II | 2nd line NSCLC refractor to PD1/PD-L1 inhibitors | 36 | Median OS: 9.7 months | |
| Relatlimab (RELATIVITY-020) [109] | LAG-3 monoclonal antibody | Relatlimab + nivolumab | I/II | Metastatic melanoma progressing after PD-1/PD-L1 inhibitors | 582 D1:354 D2:164 | ORR: D1: 12% D2: 9% | Grade 3 or more: D1: 15% D2: 13% |
| Relatlimab (RELATIVITY-047) [100] | LAG-3 monoclonal antibody | Relatlimab + nivolumab vs. nivolumab | II/III | Untreated advanced melanoma | 714 | mPFS: 10.1 vs. 4.6 months | Grade 3 or more: 19% in combination vs. 10% in monotherapy |
| Relatlimab [110] | LAG-3 monoclonal antibody | Relatlimab + nivolumab | II | Neoadjuvant treatment in operable melanoma | 60 | pCR: 57% overall PR: 70% | No grade 3 AEs |
| Favezelimab [111] | LAG-3 monoclonal antibody | Favezelimab + pembrolizumab | I/II | PD-1 inhibitor naïve R/R cHL | 30 | ORR: 73% mPFS: 19 months | Grade 3: 20% |
| Favezelimab [112] | LAG-3 monoclonal antibody | Favezelimab + pembrolizumab | I/II | Prior PD-1 inhibitor R/R cHL | 29 | ORR: 31% mPFS: 9 months | Grade 3: 18% |
| Fianlimab [113] | LAG-3 monoclonal antibody | Fianlimab + cemiplimab | I | Advanced melanoma | 33 | ORR: 64% in PD-1/PD-L1i-naïve ORR: 13% in PD-1/PD-L1i-resistant | Grade 3 or more: 35% |
| Tebotelimab [114] | LAG-3/PD-1 bispecific antibody | Tebotelimab | I/II | Advanced HCC | 63 | ORR: 13% in ICI-naïve 3% in ICI-refractory | Grade 3 or more: 19% |
| RO7247669 [115] | LAG-3/PD-1 bispecific antibody | RO7247669 | I | Advanced and/or metastatic solid tumors | 35 | ORR: 17% | Grade 3 or more: 17% |
| Reference | Drugs | Phase | N | Population | Primary Endpoint |
|---|---|---|---|---|---|
| NCT05002569 (RELATIVITY-098) | Nivolumab + relatlimab vs. nivolumab | 3 | 1050 | Adjuvant therapy after resection of stage III-IV melanoma | RFS |
| NCT05625399 (RELATIVITY-127) | Sc nivolumab + relatlimab vs. IV nivolumab + relatlimab | 3 | 814 | Previously untreated metastatic or unresectable melanoma | ORR |
| NCT05328908 (RELATIVITY-123) | Nivolumab + relatlimab vs. regorafenib or TAS-102 | 3 | 700 | Later-lines of metastatic CRC | OS in PD-L1 > 1 OS |
| NCT05337137 (RELATIVITY-106) | Nivolumab + relatlimab + bevacizumab | 1/2 | 162 | Treatment-naïve advanced or metastatic HCC | DLT PFS |
| NCT04205552 (NEOpredict) | Nivolumab alone or + relatlimab | 2 | 90 | Neoadjuvant in resectable NSCLC | Feasibility |
| NCT05704647 | Nivolumab + relatlimab | 2 | 30 | Melanoma with active brain metastases | AEs |
| NCT05148546 (NESCIO) | Nivolumab alone or + ipilimumab or relatlimab | 2 | 69 | Neoadjuvant in clear cell RCC at risk for recurrence or distant metastases | PRR |
| NCT03610711 (REACTION) | Nivolumab alone or + relatlimab | 1/2 | 21 | Advanced esophagogastric cancer | Change in infiltrating CD8+ T-cell density |
| NCT04552223 | Nivolumab + relatlimab | 2 | 27 | Metastatic uveal melanoma | ORR |
| NCT05418972 (Neo ReNi II) | Nivolumab + relatlimab | 2 | 20 | Neoadjuvant, high-risk, stage II melanoma | PRR |
| NCT03743766 | Nivolumab + relatlimab | 2 | 42 | Metastatic melanoma naïve to prior immunotherapy | ORR |
| NCT03607890 | Nivolumab + relatlimab | 2 | 42 | Advanced mismatch repair deficient cancers resistant to prior PD-(L)1 inhibitor | ORR |
| NCT04658147 | Nivolumab with or without relatlimab | 1 | 20 | Perioperative potentially resectable HCC | % of patients who complete pre-op treatment |
| NCT04095208 (CONGRATS) | Nivolumab + relatlimab | 2 | 67 | Advanced or metastatic soft-tissue sarcoma | ORR, DCR |
| NCT04913922 (AARON) | Nivolumab + relatlimab + 5-Azacytidine | 2 | 30 | R/R AML, untreated older AML patients | MTD, DLT, ORR |
| NCT05176483 (STELLAR-002) | XL092 + nivolumab + ipilimumab or relatlimab | 1 | 1078 | Unresectable advanced or metastatic solid tumors | AEs ORR PFS OS |
| NCT03623854 | Nivolumab + relatlimab | 2 | 20 | Advanced chordoma | ORR |
| NCT05255601 (RELATIVITY-069) | Nivolumab + relatlimab | 1/2 | 68 | pediatric and young adult with R/R cHL and NHL | DLT MTD |
| NCT05704933 | Nivolumab + ipilimumab or relatlimab | 1 | 24 | Surgically resectable melanoma brain metastases | Feasibility comparison of immune cell population |
| NCT05428007 | Sarilumab + ipilimumab + nivolumab + relatlimab | 2 | 69 | Unresectable stage III or stage IV melanoma | irAE ORR |
| NCT04204837 | Nivolumab alone or + relatlimab | 2 | 61 | LA or metastatic SCC of the skin | ORR |
| NCT03521830 | Nivolumab alone or + relatlimab or ipilimumab | 2 | 40 | LA or metastatic basal cell carcinoma | ORR |
| NCT03642067 | Nivolumab + relatlimab | 2 | 96 | MSS advanced CRC | ORR |
| NCT03026140 (NICHE) | Nivolumab + ipilimumab or relatlimab | 2 | 268 | Neoadjuvant combination in early stage CRC | AEs DFS |
| NCT04623775 | Nivolumab + relatlimab + chemotherapy vs. nivolumab + chemotherapy | 2 | 420 | First-line in stage IV or recurrent NSCLC | TRAE ORR |
| NCT04080804 | Nivolumab alone or in combination with relatlimab or ipilimumab | 2 | 60 | Neoadjuvant in LA resectable HNSCC | AEs |
| Reference | Drugs | Phase | N | Population | Primary Endpoint |
|---|---|---|---|---|---|
| NCT04811027 (TACTI-003) | Eftilagimod alpha + pembrolizumab | 2 | 154 | First-line: unresectable R/M HNSCC | ORR |
| NCT04252768 (AIPAC-002) | Eftilagimod alpha + paclitaxel | 1 | 24 | HR+ metastatic breast cancer | Safety and tolerability |
| NCT05747794 (AIPAC-003) | Eftilagimod alpha or placebo + paclitaxel | 3 | 849 | HER2-neg/low metastatic breast cancer | OS, Aes, OBD |
| NCT03252938 | Eftilagimod alpha | 1 | 45 | IT, IP, SC alone or in combination in advanced solid tumors | Feasibility rate |
| NCT03005782 | Fianlimab with or without REGN2810 (Anti-PD1) | 1 | 333 | Advanced malignancies | DLTs AEs Serious AEs |
| NCT05352672 | Fianlimab + cemiplimab vs. pembrolizumab | 3 | 1590 | Previously untreated unresectable LA or metastatic melanoma | PFS |
| NCT04140500 | RO7247669 (PD1-LAG3 bispecific antibody) | 1 | 320 | Advanced and/or metastatic solid tumors | DLTs, AEs, ORR, DCR, DOR, PFS |
| NCT05419388 | RO7247669 | 1/2 | 80 | Previously untreated unresectable or metastatic melanoma | PFS |
| NCT05645692 | RO7247669 +/− tiragolumab vs. atezolizumab | 2 | 240 | Previously untreated advanced or metastatic UC ineligible for platinum-containing chemotherapy | ORR |
| NCT04785820 | RO7247669 vs. RO7121661 (PD1-TIM3 bispecific antibody) vs. nivolumab | 2 | 210 | Relapsed or intolerant to platinum-containing regimens in A/M SCCE | OS |
| NCT05508867 (KEYFORM-008) | favezelimab + pembrolizumab vs. physician’s choice chemotherapy | 3 | 360 | PD-(L)1-refractory, R/R classical Hodgkin lymphoma | PFS |
| NCT05064059 (MK-4280A-007) | favezelimab + pembrolizumab vs. SOC | 3 | 432 | Previously treated metastatic PD-L1 positive CRC | OS |
| NCT03598608 (MK-4280-003) | Favezelimab + pembrolizumab | 1/2 | 174 | Hematologic malignancies | DLTs, AEs, treatment discontinuation due to AEs |
| NCT04938817 (MK-3475-B98/KEYNOTE-B98) | Pembrolizumab + favezelimab or quavonlimab | 1/2 | 80 | PD-(L)1 refractory extensive-stage SCLC | DLTs AEs TRAEs ORR |
| NCT05695898 | XmAb23104 (PD1-ICOS) + XmAb22841 (CTLA-4-LAG3) | 1/2 | 46 | Metastatic melanoma refractory to prior ICI with and without CNS disease | TEAEs, irAEs, DLTs |
| NCT04150965 | BMS-986016 + Pomalidomide + dexamethasone (Arm B) | 1/2 | 104 | Relapsed and/or refractory MM | ORR, AEs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, R.; Saleh, K.; Chahine, C.; Khoury, R.; Khalife, N.; Cesne, A.L. LAG-3 Inhibitors: Novel Immune Checkpoint Inhibitors Changing the Landscape of Immunotherapy. Biomedicines 2023, 11, 1878. https://doi.org/10.3390/biomedicines11071878
Ibrahim R, Saleh K, Chahine C, Khoury R, Khalife N, Cesne AL. LAG-3 Inhibitors: Novel Immune Checkpoint Inhibitors Changing the Landscape of Immunotherapy. Biomedicines. 2023; 11(7):1878. https://doi.org/10.3390/biomedicines11071878
Chicago/Turabian StyleIbrahim, Rebecca, Khalil Saleh, Claude Chahine, Rita Khoury, Nadine Khalife, and Axel Le Cesne. 2023. "LAG-3 Inhibitors: Novel Immune Checkpoint Inhibitors Changing the Landscape of Immunotherapy" Biomedicines 11, no. 7: 1878. https://doi.org/10.3390/biomedicines11071878