
Alteration of Mitochondrial DNA Copy Number and Increased Expression Levels of Mitochondrial Dynamics-Related Genes in Sjögren’s Syndrome

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Recruitment
2.2. Mitochondrial DNA Copy Number Evaluation
2.3. mRNA Isolation and Expression Analysis
2.4. Statistical Analysis
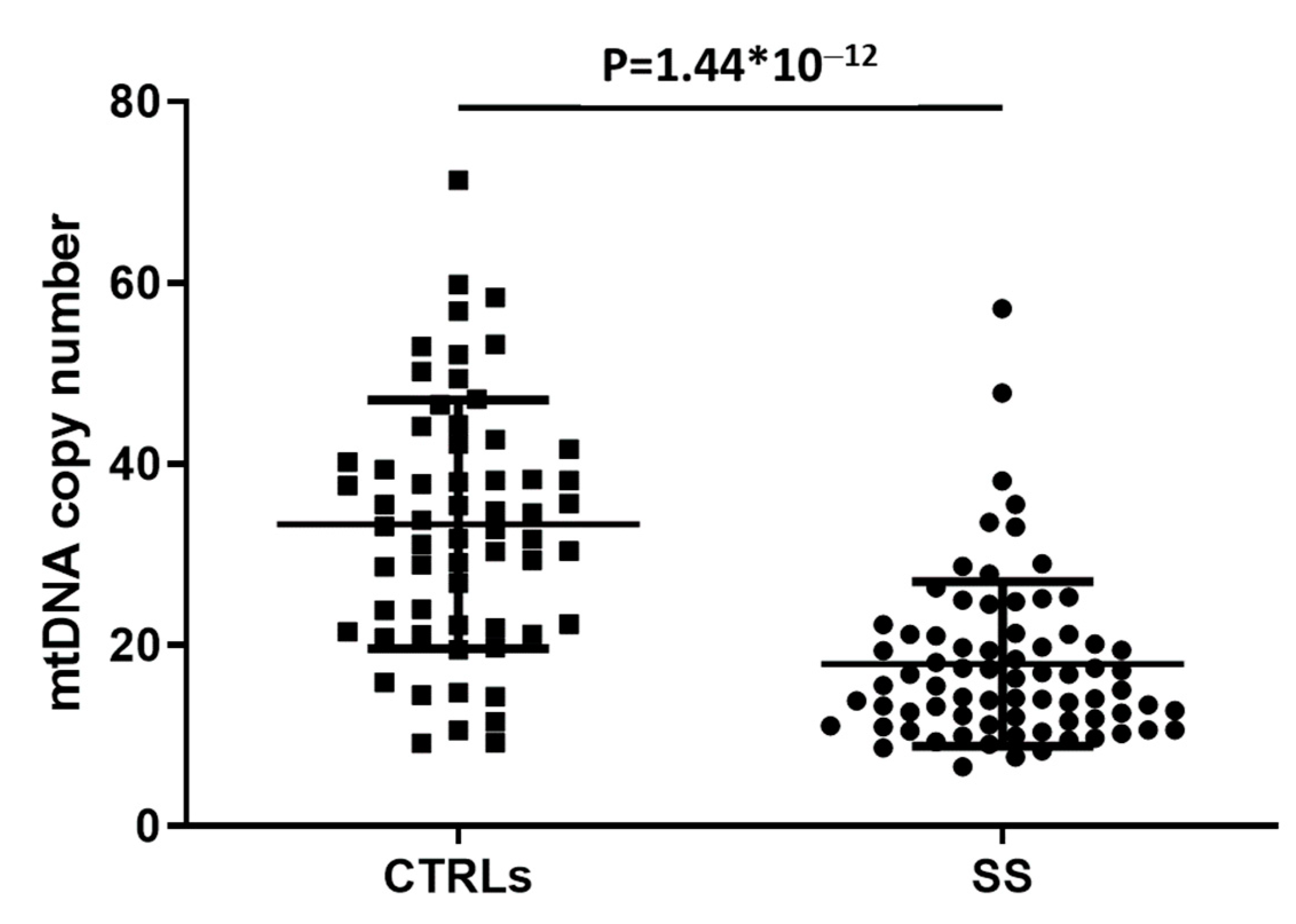
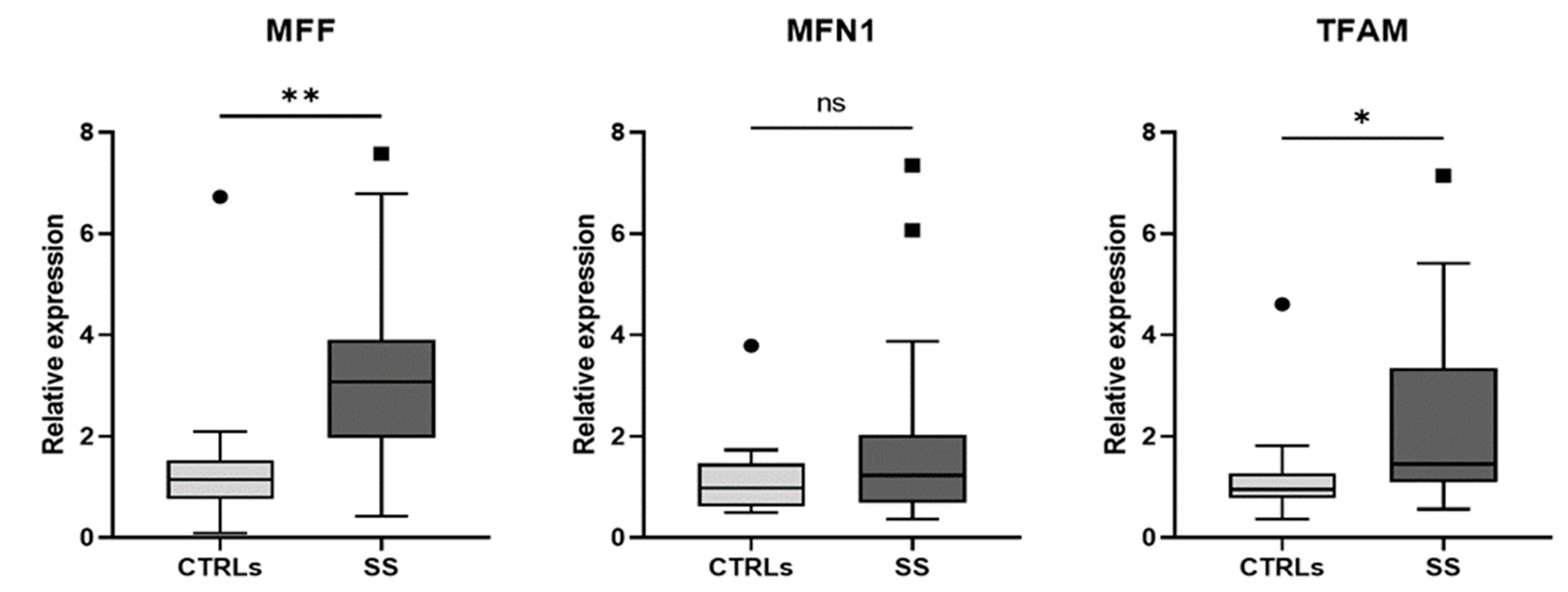
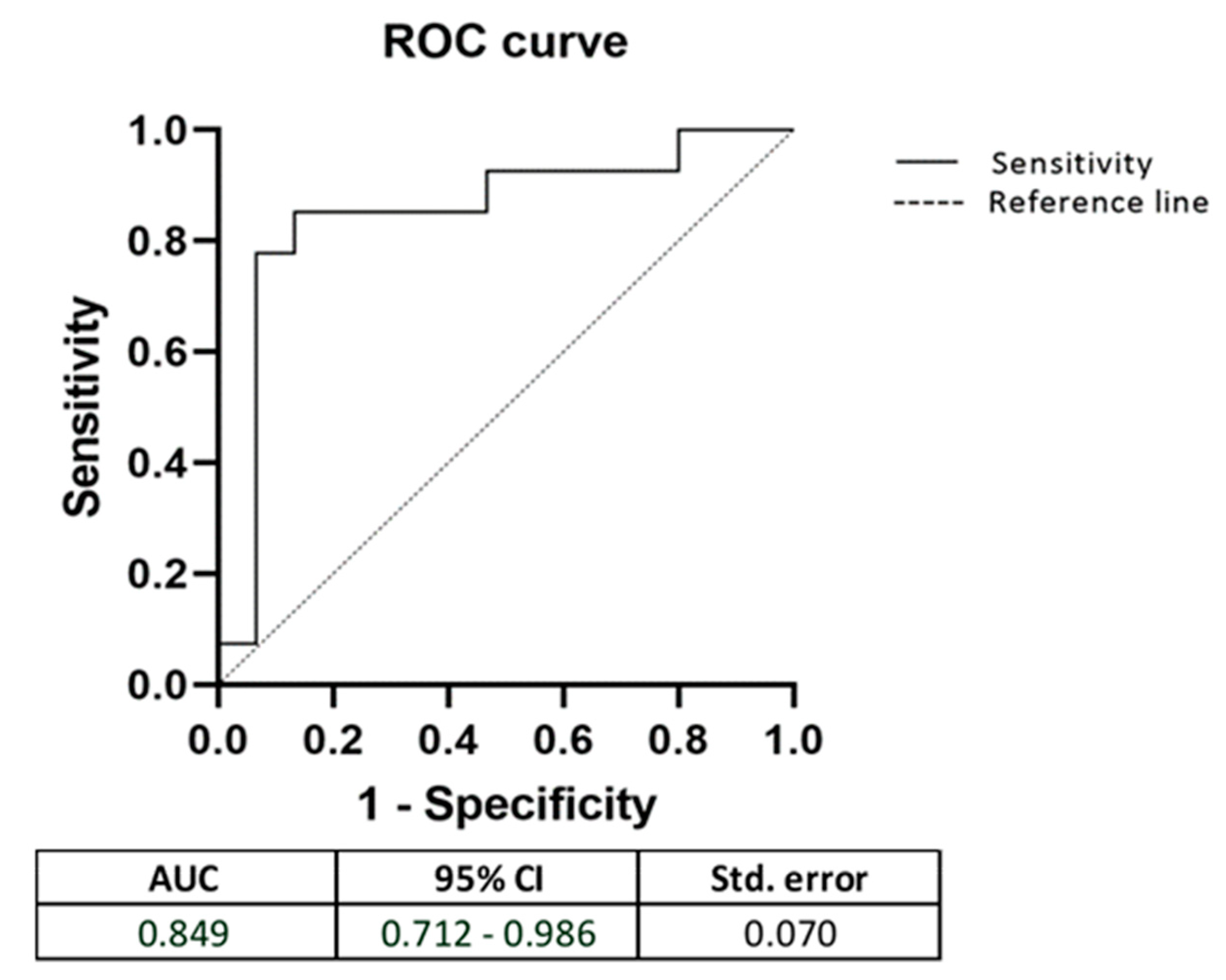
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mariette, X.; Criswell, L.A. Primary Sjögren’s Syndrome. N. Engl. J. Med. 2018, 378, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Imgenberg-Kreuz, J.; Rasmussen, A.; Sivils, K.; Nordmark, G. Genetics and epigenetics in primary Sjögren’s syndrome. Rheumatology 2021, 60, 2085–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimessi, A.; Previati, M.; Nigro, F.; Wieckowski, M.R.; Pinton, P. Mitochondrial reactive oxygen species and inflammation: Molecular mechanisms, diseases and promising therapies. Int. J. Biochem. Cell Biol. 2016, 81, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Sandhir, R.; Halder, A.; Sunkaria, A. Mitochondria as a centrally positioned hub in the innate immune response. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1090–1097. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, A.; Tiwari-Pandey, R.; Pandey, N.R. Mitochondria: The indispensable players in innate immunity and guardians of the inflammatory response. J. Cell Commun. Signal. 2019, 13, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Gergely, P., Jr.; Grossman, C.; Niland, B.; Puskas, F.; Neupane, H.; Allam, F.; Banki, K.; Phillips, P.E.; Perl, A. Mitochondrial hyperpolarization and ATP depletion in patients with systemic lupus erythematosus. Arthritis Rheum. 2002, 46, 175–190. [Google Scholar] [CrossRef]
- Ghafourifar, P.; Mousavizadeh, K.; Parihar, M.S.; Nazarewicz, R.R.; Parihar, A.; Zenebe, W.J. Mitochondria in multiple sclerosis. Front. Biosci. 2008, 13, 3116–3126. [Google Scholar] [CrossRef] [Green Version]
- Fearon, U.; Canavan, M.; Biniecka, M.; Veale, D.J. Hypoxia, mitochondrial dysfunction and synovial invasiveness in rheumatoid arthritis. Nat. Rev. Rheumatol. 2016, 12, 385–397. [Google Scholar] [CrossRef]
- Maiese, K.; Morhan, S.D.; Chong, Z.Z. Oxidative stress biology and cell injury during type 1 and type 2 diabetes mellitus. Curr. Neurovasc. Res. 2007, 4, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Sivitz, W.I.; Yorek, M.A. Mitochondrial dysfunction in diabetes: From molecular mechanisms to functional significance and therapeutic opportunities. Antioxid. Redox Signal. 2010, 12, 537–577. [Google Scholar] [CrossRef]
- Becker, Y.; Loignon, R.C.; Julien, A.S.; Marcoux, G.; Allaeys, I.; Lévesque, T.; Rollet-Labelle, E.; Benk-Fortin, H.; Cloutier, N.; Melki, I.; et al. Anti-mitochondrial autoantibodies in systemic lupus erythematosus and their association with disease manifestations. Sci. Rep. 2019, 9, 4530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colapietro, F.; Lleo, A.; Generali, E. Antimitochondrial Antibodies: From Bench to Bedside. Clin. Rev. Allergy Immunol. 2021, 29, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ryo, K.; Yamada, H.; Nakagawa, Y.; Tai, Y.; Obara, K.; Inoue, H.; Mishima, K.; Saito, I. Possible involvement of oxidative stress in salivary gland of patients with Sjogren’s syndrome. Pathobiology 2006, 73, 252–260. [Google Scholar] [CrossRef]
- Norheim, K.B.; Jonsson, G.; Harboe, E.; Hanasand, M.; Gøransson, L.; Omdal, R. Oxidative stress, as measured by protein oxidation, is increased in primary Sjøgren’s syndrome. Free Radic. Res. 2012, 46, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, A.; Kurien, B.T.; Scofield, R.H. Autoantibodies in Sjögren’s Syndrome. Rheum. Dis. Clin. N. Am. 2016, 42, 419–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scofield, R.H.; Fayyaz, A.; Kurien, B.T.; Koelsch, K.A. Prognostic value of Sjögren’s syndrome autoantibodies. J. Lab. Precis. Med. 2018, 3, 10. [Google Scholar] [CrossRef]
- Chan, D.C. Fusion and fission: Interlinked processes critical for mitochondrial health. Annu. Rev. Genet. 2012, 46, 265–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial fission, fusion, and stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [Green Version]
- Kang, I.; Chu, C.T.; Kaufman, B.A. The mitochondrial transcription factor TFAM in neurodegeneration: Emerging evidence and mechanisms. FEBS Lett. 2018, 592, 793–811. [Google Scholar] [CrossRef] [Green Version]
- Noack, H.; Bednarek, T.; Heidler, J.; Ladig, R.; Holtz, J.; Szibor, M. TFAM-dependent and independent dynamics of mtDNA levels in C2C12 myoblasts caused by redox stress. Biochim. Biophys. Acta 2006, 1760, 141–150. [Google Scholar] [CrossRef]
- Ekstrand, M.I.; Falkenberg, M.; Rantanen, A.; Park, C.B.; Gaspari, M.; Hultenby, K.; Rustin, P.; Gustafsson, C.M.; Larsson, N.G. Mitochondrial transcription factor A regulates mtDNA copy number in mammals. Hum. Mol. Genet. 2004, 13, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunkel, G.H.; Chaturvedi, P.; Tyagi, S.C. Mitochondrial pathways to cardiac recovery: TFAM. Heart Fail Rev. 2016, 21, 499–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrera, M.J.; Aguilera, S.; Castro, I.; Carvajal, P.; Jara, D.; Molina, C.; González, S.; González, M.J. Dysfunctional mitochondria as critical players in the inflammation of autoimmune diseases: Potential role in Sjögren’s syndrome. Autoimmun. Rev. 2021, 20, 102867. [Google Scholar] [CrossRef]
- Shiboski, C.H.; Shiboski, S.C.; Seror, R.; Criswell, L.A.; Labetoulle, M.; Lietman, T.M.; Rasmussen, A.; Scofield, H.; Vitali, C.; Bowman, S.J.; et al. 2016 American College of Rheumatology/European League Against Rheumatism classification criteria for primary Sjögren’s syndrome: A consensus and data-driven methodology involving three international patient cohorts. Ann. Rheum. Dis. 2017, 76, 9–16. [Google Scholar] [CrossRef]
- Rooney, J.P.; Ryde, I.T.; Sanders, L.H.; Howlett, E.H.; Colton, M.D.; Germ, K.E.; Mayer, G.D.; Greenamyre, J.T.; Meyer, J.N. PCR based determination of mitochondrial DNA copy number in multiple species. Methods Mol. Biol. 2015, 1241, 23–38. [Google Scholar]
- Xing, J.; Chen, M.; Wood, C.G.; Lin, J.; Spitz, M.R.; Ma, J.; Amos, C.I.; Shields, P.G.; Benowitz, N.L.; Gu, J.; et al. Mitochondrial DNA content: Its genetic heritability and association with renal cell carcinoma. J. Natl. Cancer Inst. 2008, 100, 1104–1112. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.; Shahane, A. The epidemiology of Sjögren’s syndrome. Clin. Epidemiol. 2014, 6, 247–255. [Google Scholar]
- Pagano, G.; Talamanca, A.A.; Castello, G.; Cordero, M.D.; d’Ischia, M.; Gadaleta, M.N.; Pallardó, F.V.; Petrović, S.; Tiano, L.; Zatterale, A. Oxidative stress and mitochondrial dysfunction across broad-ranging pathologies: Toward mitochondria-targeted clinical strategies. Oxid. Med. Cell Longev. 2014, 2014, 541230. [Google Scholar] [CrossRef]
- Liu, A.; Kage, F.; Higgs, H.N. Mff oligomerization is required for Drp1 activation and synergy with actin filaments during mitochondrial division. Mol. Biol. Cell 2021, 32, ar5. [Google Scholar] [CrossRef]
- Otera, H.; Wang, C.; Cleland, M.M.; Setoguchi, K.; Yokota, S.; Youle, R.J.; Mihara, K. Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells. J. Cell Biol. 2010, 191, 1141–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shutt, T.; Geoffrion, M.; Milne, R.; McBride, H.M. The intracellular redox state is a core determinant of mitochondrial fusion. EMBO Rep. 2012, 10, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorzano, A.; Liesa, M.; Sebastián, D.; Segalés, J.; Palacín, M. Mitochondrial fusion proteins: Dual regulators of morphology and metabolism. Semin. Cell Dev. Biol. 2010, 21, 566–574. [Google Scholar] [CrossRef]
- Ishihara, N.; Otera, H.; Oka, T.; Mihara, K. Regulation and physiologic functions of GTPases in mitochondrial fusion and fission in mammals. Antioxid. Redox Signal. 2013, 19, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, Y.; Hu, J.; Wu, Y.; Yang, J.; Fan, H.; Li, L.; Luo, D.; Ye, Y.; Gao, Y.; et al. A Link Between Mitochondrial Dysfunction and the Immune Microenvironment of Salivary Glands in Primary Sjogren’s Syndrome. Front. Immunol. 2022, 13, 845209. [Google Scholar] [CrossRef]
- Xu, S.; Zhong, M.; Zhang, L.; Wang, Y.; Zhou, Z.; Hao, Y.; Zhang, W.; Yang, X.; Wei, A.; Pei, L.; et al. Overexpression of Tfam protects mitochondria against beta-amyloid-induced oxidative damage in SH-SY5Y cells. FEBS J. 2009, 276, 3800–3809. [Google Scholar] [CrossRef]
- Aguirre-Rueda, D.; Guerra-Ojeda, S.; Aldasoro, M.; Iradi, A.; Obrador, E.; Ortega, A.; Mauricio, M.D.; Vila, J.M.; Valles, S.L. Astrocytes protect neurons from Aβ1-42 peptide-induced neurotoxicity increasing TFAM and PGC-1 and decreasing PPAR-γ and SIRT-1. Int. J. Med. Sci. 2015, 12, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Bonekamp, N.A.; Jiang, M.; Motori, E.; Garcia Villegas, R.; Koolmeister, C.; Atanassov, I.; Mesaros, A.; Park, C.B.; Larsson, N.G. High levels of TFAM repress mammalian mitochondrial DNA transcription in vivo. Life Sci. Alliance 2021, 4, e202101034. [Google Scholar] [CrossRef]
- Zhao, Y.; Peng, C.; Zhang, J.; Lai, R.; Zhang, X.; Guo, Z. Mitochondrial Displacement Loop Region SNPs Modify Sjögren’s Syndrome Development by Regulating Cytokines Expression in Female Patients. Front. Genet. 2022, 13, 847521. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| SS (n = 74) | SS Subgroup (n = 27) | |
|---|---|---|
| Sex (% of females) | 92.2 | 88.5 |
| Age (mean ± SD) | 59.5 ± 11.09 | 58.79 ± 10.33 |
| Age at diagnosis (mean ± SD) | 51.1 ± 11.65 | 54.14 ± 9.76 |
| Xerophthalmia (%) | 98.6 | 96.4 |
| Xerostomia (%) | 90.5 | 85.7 |
| Salivary gland swelling (%) | 21.6 | 3.6 |
| Arthritis (%) | 14.9 | 14.3 |
| Lymphoma (%) | 10.8 | 7.1 |
| ANA (%) | 91.9 | 82.1 |
| Anti-SSA (%) | 86.5 | 64.3 |
| Anti-SSB (%) | 60.8 | 53.6 |
| Hypergammaglobulinemia (%) | 41.9 | 39.3 |
| Rheumatoid factor (%) | 41.1 | 23.1 |
| Leukopenia (%) | 23.0 | 28.6 |
| Hypocomplementemia (%) | 5.5 | 14.3 |
| Monoclonal component (%) | 11.1 | 14.8 |
| Cryoglobulins (%) | 4.2 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Benedittis, G.; Latini, A.; Colafrancesco, S.; Priori, R.; Perricone, C.; Novelli, L.; Borgiani, P.; Ciccacci, C. Alteration of Mitochondrial DNA Copy Number and Increased Expression Levels of Mitochondrial Dynamics-Related Genes in Sjögren’s Syndrome. Biomedicines 2022, 10, 2699. https://doi.org/10.3390/biomedicines10112699
De Benedittis G, Latini A, Colafrancesco S, Priori R, Perricone C, Novelli L, Borgiani P, Ciccacci C. Alteration of Mitochondrial DNA Copy Number and Increased Expression Levels of Mitochondrial Dynamics-Related Genes in Sjögren’s Syndrome. Biomedicines. 2022; 10(11):2699. https://doi.org/10.3390/biomedicines10112699
Chicago/Turabian StyleDe Benedittis, Giada, Andrea Latini, Serena Colafrancesco, Roberta Priori, Carlo Perricone, Lucia Novelli, Paola Borgiani, and Cinzia Ciccacci. 2022. "Alteration of Mitochondrial DNA Copy Number and Increased Expression Levels of Mitochondrial Dynamics-Related Genes in Sjögren’s Syndrome" Biomedicines 10, no. 11: 2699. https://doi.org/10.3390/biomedicines10112699