
REST Is Not Resting: REST/NRSF in Health and Disease

1
School of Life Sciences, Liaoning University, Shenyang 110036, China
2
Department of Stem Cells and Regenerative Medicine, Key Laboratory of Cell Biology, National Health Commission of China, and Key Laboratory of Medical Cell Biology, Ministry of Education of China, China Medical University, Shenyang 110122, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Biomolecules 2023, 13(10), 1477; https://doi.org/10.3390/biom13101477
Submission received: 11 June 2023
/
Revised: 26 September 2023
/
Accepted: 29 September 2023
/
Published: 2 October 2023
{kind=link}
{kind=link}
Abstract
:Chromatin modifications play a crucial role in the regulation of gene expression. The repressor element-1 (RE1) silencing transcription factor (REST), also known as neuron-restrictive silencer factor (NRSF) and X2 box repressor (XBR), was found to regulate gene transcription by binding to chromatin and recruiting chromatin-modifying enzymes. Earlier studies revealed that REST plays an important role in the development and disease of the nervous system, mainly by repressing the transcription of neuron-specific genes. Subsequently, REST was found to be critical in other tissues, such as the heart, pancreas, skin, eye, and vascular. Dysregulation of REST was also found in nervous and non-nervous system cancers. In parallel, multiple strategies to target REST have been developed. In this paper, we provide a comprehensive summary of the research progress made over the past 28 years since the discovery of REST, encompassing both physiological and pathological aspects. These insights into the effects and mechanisms of REST contribute to an in-depth understanding of the transcriptional regulatory mechanisms of genes and their roles in the development and progression of disease, with a view to discovering potential therapeutic targets and intervention strategies for various related diseases.
1. Introduction
The repressor element-1 (RE1) silencing transcription factor (REST), also known as neuron-restrictive silencer factor (NRSF) and X2 box repressor (XBR), is a transcriptional repressor first reported in 1995. REST binds to the RE1 motif, also named neural restrictive silencing element (NRSE), and recruits corepressor complexes [1,2]. Initially identified as a regulator of neuron-specific genes in nonneuronal tissues, REST has been found to play crucial roles in neuronal development, function, and related diseases. Over nearly three decades of research, REST has been revealed to have diverse functions in other tissues such as the heart, pancreas, skin, eye, and vascular, as well as in various cancers. In this comprehensive review, we systematically summarize the research progress of REST, with a view to providing valuable insight into exploring the function of REST and the potential treatment of related diseases.
1.1. Gene and Protein Structure
The full-length human REST contains a DNA binding domain (DBD), an N-terminal repressor domain (NRD), and a C-terminal repressor domain (CRD) (Figure 1). REST is highly conserved in mammals, and there are various mRNA alternative splice variants predictive of multiple protein isoforms. Studies showed that REST might produce at least 45 mRNA variants encoding REST4, REST1, RESTC, and small peptides [3]. Interestingly, Chen and Miller noted that the variable splicing of REST produces more complex protein isoforms than expected. The detections using different primers, probes, or antibodies may target different variants, and knockout (KO) or knockdown of REST can be effective against one or a few, but not all, of the specific variants. Variable splicing of REST was considered to be underestimated in some previous studies, which may have led to inconsistent data in different studies and data misinterpretation [3].
1.2. Nuclear Location
The nuclear location of transcription factors is essential for its modulatory function. Classically, REST binds to DNA and recruits coregulators to modulate gene expression upon entering the nucleus. The deletion of residues 512–522 in the full-length REST resulted in cytosol location. However, the isoform REST4, which lacks this putative nuclear localization signal (NLS), was localized to the nucleus, while another isoform REST1, lacking zinc finger 5, was localized in the cytosol [4]. Via deletion and mutation, the NLS around zinc finger 5 was verified to be the sole NLS within REST [5]. In addition, the nuclear envelope protein REST/NRSF-interacting Lin-11, Isl-1, and Mec-3 (LIM) domain protein (RILP) served as an essential nuclear receptor for REST [6].
1.3. DNA Binding
The DBD containing eight noncanonical Cys2-His2 zinc fingers (zinc finger 1–8 from N-terminus to C-terminus in Figure 1B) is crucial for DNA binding [2]. The deletion of zinc finger 3 or 5 abolished the DNA binding activity of REST, and the segment of amino acids 196–207 is essential for DNA binding [7]. Generally, each Cys2-His2 zinc finger domain binds three nucleotides within DNA, and a point mutation that changes a Cys residue to an Arg residue disrupts the Cys2-His2 zinc finger. The single mutation in zinc finger 7 and combinations of mutations in any two of zinc finger 6–8 decreases DNA binding, while a single mutation in zinc finger 6 or 8 has little effect [4]. Further, the residue Cys397 plays an important role in the global folding of multiple zinc fingers domain [8].
Initially, a homologous DNA element named NRSE within SCG10, SCN2A2, and SYN1 genes was discovered to repress their transcription via REST binding in nonneuronal cells [9,10,11]. Using biochemical, bioinformatic, and high-throughput sequencing approaches, the canonical and noncanonical potential NRSE motifs within more than 3000 genes have been characterized in vertebrates, including humans, mice, rats, and fugu [12,13,14,15,16,17,18], after the cloning of complementary DNA for REST [1,2]. These investigations have provided valuable insights into the wide-ranging presence of NRSE motifs across various species. The regulatory effects of REST on the expression of target genes are dependent on specific physiological and pathological contexts, and hundreds of target genes have been validated and studied.
Concurrently, several bioinformatics approaches were developed to facilitate the analysis of REST binding. For example, Cistematic [19] and CodingMotif [20], which were employed for predicting transcription factor binding sites. In addition, there are tools such as Site Identification from Short Sequence Reads (SISSRs) [21], CisGenome [22], quantitative enrichment of sequence tags (QuEST) [23], USeq [24], Motif Identification for ChIP-Seq Analysis (MICSA) [25], Genomic Regions Enrichment of Annotations Tool (GREAT) [26], and NEXT-peak [27] that have been developed specifically for the analysis of high-throughput chromatin immunoprecipitation (ChIP) data. These computational tools assist researchers in identifying and characterizing REST binding sites in a genome-wide context, providing valuable insights into the regulatory mechanisms and functional implications of REST.
1.4. Coregulator Recruitment
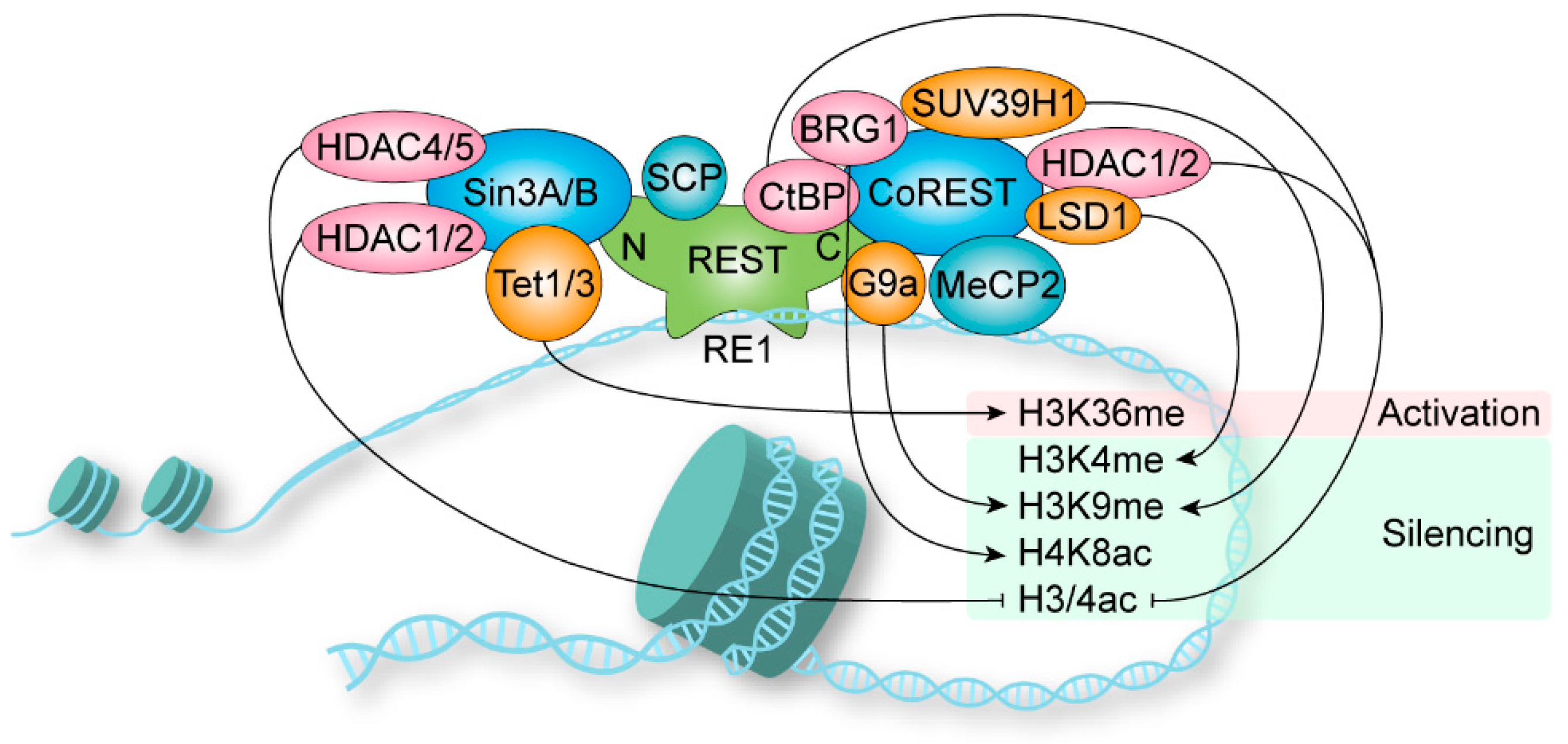
The transcriptional regulation mediated by REST requires the assembly of multimeric complexes [28]. Sin3 is constitutively required for REST repression, while CoREST is recruited for more specialized repressor functions [29] (Figure 2). The NRD of REST recruits Sin3A and Sin3B, which interact with HDAC1/2, HDAC4/5, and other adaptors to form a complex [30,31,32,33,34]. The HDACs remove acetyl groups from histone to silent chromatin. Nevertheless, the functions of the other adaptor proteins are largely not clear [35]. The short hydrophobic a-helix structure of NRD is captured in the cleft of the PAH1 domain of Sin3B, and PAH1 is highly conserved among Sin3A and Sin3B [36,37]. Further, the Sin3 complex is also involved in transcriptional activation [32]. Sin3A recruits Tet1 to the PAH1 domain, and Sin3 interaction domains in Tet1/Tet3 are highly conserved [38]. In the mouse retina, REST was found to mediate transcriptional activation via recruiting Tet3 to DNA for 5-hydroxymethylcytosine generation followed by NSD3-mediated H3K36 trimethylation, but no classical repressor complex protein was identified [39]. Thus, the dependency of REST-mediated gene activation on the Sin3-dependent remains enigmatic. In addition, REST recruits small CTD phosphatases (SCPs) to neuronal genes to reduce RNA polymerase II activity, and the responsible domain for SCP is still unclear [40].
The CRD containing a zinc finger (zinc finger 9) recruits CoREST as a beacon for the recruitment of repressors [41,42], including HDAC1/2, LSD1, BRG1, CtBP, G9a, SUV39H1, HP1 and MeCP2 [43]. HDACs remove acetyl groups on H3 and H4, and H3K9 deacetylation triggers localized domains of histone methyltransferase G9a-mediated H3K9 methylation [44]. The K9 residues methylated by G9a and another histone, H3K9 methyltransferase SUV39H1, are binding sites for heterochromatin protein 1 (HP1), which causes chromatin condensation [42]. Further, REST (amino acids 141–600) recruits G9a and interacts with Mediator via MED19/MED26 and MED12 subunits, respectively [45,46]. The histone demethylase LSD1 is involved in reducing H3K4 methylation [47]. The chromatin-remodeling enzyme BRG1 increases H4K8 acetylation, which stabilizes the interaction between REST and chromatin [48]. CtBP is a corepressor of REST, and the reduction of glycolysis-derived NADH promotes the recruitment of CtBP, leading to histone deacetylation [49,50]. In certain genes during neuron differentiation, methyl-CpG-binding protein MeCP2 and the CoREST repressor complex bind to methylated DNA, resulting in gene repression independently of REST [51].
2. REST in Nervous System
2.1. Embryogenesis and Neurogenesis
During mouse embryogenesis, REST was found to be required in repressing neuronal gene expression in both non-neural and neural precursor tissues [52]. The postnatal development of the hippocampus, especially the formation of the subgranular zone (SGZ), was influenced by conditional knockout (cKO) of REST in mice [53]. During early development, the loss of REST induces global hypermethylation [54,55], and the deregulation of REST was found to be an early event in Down syndrome (DS) [56,57]. In neural progenitor cells, REST was under the regulation of Wnt signaling, which controls various aspects of early development [58,59]. In zebrafish, REST serves as a regulator of hedgehog signaling, and it is required for gastrulation and neurogenesis during embryogenesis [60,61]. Additionally, REST is required for the ectodermal cell fates in Xenopus [62]. In addition, REST was involved in the inhibition of neurogenesis in dorsal root ganglion (DRG) by NO [63]. However, zebrafish with REST mutation generated by zinc-finger nuclease targeting underwent largely normal neurogenesis [64], and they have behavioral abnormalities such as erratic swimming and abnormal spatial preferences in adults and hypoactivity in larva [65]. In REST cKO mice, REST plays a role in the suppression of neuron genes in cultured neuronal and non-neuronal cells. However, target gene expression was not altered upon REST KO in vivo and the mice are apparently normal and grow into adults [66]. Thus, REST plays important roles in embryo development and neurogenesis [67], and its absence could be partially compensated, with the mechanisms involved being unclear.
2.2. Neuronal Differentiation
REST is a critical regulator of differentiation in embryonic stem cells (ESCs), neural stem cells (NSCs), and mature cell types [18,51]. In ESCs, a single REST allele disruption reduced alkaline phosphatase activity and several pluripotency-associated genes expression [68]. However, partial or complete loss of REST could not abrogate differentiation potential as reflected by marker gene expression [69]. REST knockdown in mouse ESCs could induce neuronal lineage differentiation [69], and its knockdown in human ESCs increased cell survival and the expression of mesendoderm differentiation markers [70]. Further, REST-mediated antenatal-arsenic-induced NSC function dysregulation led to deficits in differentiation in adults [71].
The repression of neuron-specific genes by REST in nonneuronal tissues highlights its crucial function during neuronal differentiation. Using ChIP-seq data mining, REST was found to mediate more than 2000 gene silencing in ESCs but not in ESC-derived neurons [72]. REST is critical for maintaining the NSCs pool, and the activation of REST target genes by REST-VP16 transgene was sufficient to induce neuronal differentiation in NSCs [73,74]. The nuclear REST was reduced during neuronal differentiation of embryonic cells, and its target genes N-methyl-D-aspartate receptor subunit 1 (NR1) and tyrosine hydroxylase (TH) were upregulated [75,76]. Further, REST plays stage-specific roles during NSC-mediated astrocyte (AS) and oligodendrocyte (OL) lineage specification, OL lineage maturation, and myelination [77,78].
The master regulatory effects of REST offer a promising strategy for the induction of neuronal differentiation. REST silencing induces neuronal differentiation of human mesenchymal stem cells (MSCs) and rat hippocampal progenitors (AHPs), and secretogranin II (Scg2) was a critical secreted REST target responsible for non-cell-autonomous phenotype [79,80]. Recently, the neuronal differentiation of human MSCs was achieved using Crispr/Cas9 targeting REST [81].
2.3. Neuronal Survival
REST expression is typically low in differentiated neurons, whereas its elevation frequently exhibits protective effects. For instance, the mice with neural crest-specific KO of REST presented aberrations of the myenteric plexus, leading to neonatal lethality [82]. In differentiated neural cells, overexpression of REST promoted proliferation and disrupted neurosecretion via the REST-TSC2-β-catenin signaling pathway [83]. BMP/RA-inducible neural-specific protein 1 (BRINP1) was a negative regulator of cell cycle progression, and its promotor activity was inhibited by the RE1 motif within its promotor [84]. REST inhibited BAF53a to quit mitosis by targeting miR-9* and miR-124 in post-mitotic neurons [85]. In addition, BNDF transcription was regulated by REST and its competitive isoform REST4 to regulate various neuronal cell behaviors, including cell survival [86]. In addition, ethanol was found to induce NR2B expression in fetal cortical neurons by regulating REST [87]. The augment of REST binding activity contributes to the depression of differentiation and neurogenesis induced by ethanol exposure [88,89]. REST protects the developing brain from the detrimental effects of ethanol, indicating it as a potential target for fetal alcohol syndrome treatment [90].
The expression of REST in SH-SY5Y cells was induced by polychlorinated biphenyls (PCBs), and ERK2/Sp1/Sp3/REST signaling mediated the neuronal toxicity of PCBs [91]. REST was increased during monosodium-glutamate-induced excitotoxicity in rats [92]. REST could protect cath.-a-differentiated (CAD) neuronal cells from Mn-induced toxicity by enhancing the expression of the dopamine-synthesizing enzyme tyrosine hydroxylase (TH) [93]. The dual roles of REST in the mediation of neuroprotection and neurotoxicity suggest REST as a critical regulator in neural survival.
2.4. Neuronal Transmission and Synaptic Plasticity
REST maintains neuronal function balance by dynamically regulating the synaptic efficiency. To facilitate neurite outgrowth, REST restricted the cell adhesion molecule L1 in the embryonic nervous system [94] and positively regulated the exocytosis of enlargeosomes [95]. However, the constitutive expression of REST led to axon guidance mistakes in the spinal cord of chicken embryos, with the depression of the target genes N-tubulin and Ng-CAM [96]. Syn1 was negatively regulated by REST to modulate synaptogenesis and neurotransmitter release [97], and Sec6 has a REST binding site and function on synapse formation and synaptic plasticity [98]. Further, the synaptic function is altered in an inflammatory environment involved in various neurodegenerative pathologies [99]. The activation of REST was found to mediate the synaptic scaling in the presence of neuroinflammation induced by IL-1β [100]. REST repressed target genes, including NPAS4 and BNDF, to regulate inhibitory synapse gene expression, thereby maintaining the excitatory/inhibitory balance in neurons [101,102]. Additionally, in NMB cells, REST inhibited VGF indicating its involvement in hippocampal synaptic activity [103].
REST protects neurons from hyperexcitability via modulating the transcriptional activity of voltage-gated ion channel genes. REST elevated by hyperactivity inhibited Nav1.2 expression to maintain homeostasis, and silencing of REST disrupted this homeostatic response [104]. Another major sodium channel, SCN8A (Nav1.6), was also found to be under the control of REST [105]. Various Ca2+ signaling was repressed by REST differentially [106]. The functional expression of P/Q-type channels was increased by REST silencing in GN11 neuron cells [107], while the promotor activity of N-type Ca2+ channel alpha 1B was not influenced by REST in NS20Y cells [108]. Recently, REST was found to control spatial K buffering in primary cortical astrocytes, impairing neuronal activity [109].
Consistently, REST presented distinct repressive actions on the transcriptional activity of neurotransmitter and receptor genes. The expression of choline acetyltransferase (ChAT) was restricted by REST to neuronal cells [110]. REST controlled tryptophan hydroxylase 2 (TPH2) expression to modulate the synthesis of neuronal serotonin [111]. The discovered receptors under the control of REST include neural-specific m4 muscarinic acetylcholine receptor (mAChR) [112,113], gamma2 subunits of the type A receptors for gamma-aminobutyric acid (GABA) [114], N-methyl-D-aspartate (NMDA) receptor subunit type I (NR1) [115], 5-HT1A receptor [116], AMPA glutamate receptor subunit GluR2 [117], and adrenergic α2C receptor (ADRA2C) [118].
2.5. Pain
REST regulates μ-opioid receptor (MOR) transcriptional activity [119,120,121], contributing to MOR agonist remifentanil inducing postoperative hyperalgesia [122]. PACAP was regulated by REST to mediate neuropathic pain after spinal nerve injury [123]. The negative regulation of NR2B by REST was involved in bone cancer pain [124]. The repression of MOR by REST reduced morphine analgesia in sarcoma-induced bone cancer pain [125]. REST mediated diabetic neuropathic pain in db/db mice [126]. REST also plays a major role in the acute-to-chronic pain transition upon nerve injury [127]. REST KO reverted injury-induced hyperalgesia, and REST regulates peripheral somatosensory neuron programming, leading to chronic pain [128]. REST silenced K(v)4.3 in DRG nerve injury [129]. Nerve injury elevated REST, which silenced MOP and Na(v)1.8 epigenetically [130]. REST enhances some pain, and targeting REST to release its inhibition of target genes provides a novel perspective for analgesia.
2.6. Neuroendocrine
The cocaine- and amphetamine-regulated transcript (CART) peptide is under strict control of REST in neuroendocrine cells [131]. The neurosecretory process involving dense-core vesicles (DCVs) was governed by REST in PC12 and astrocyte cells [132,133]. The depression of REST by Kaposi’s sarcoma-associated herpesvirus (KSHV) latent open reading frame K12 (ORFK12) gene (kaposin A) regulated neuroendocrine gene expression in infected endothelial cells [134]. Further, the proprotein convertase PC2 was regulated in nonneuroendocrine cells [135]. Interestingly, REST could repress the transcription of corticotropin releasing hormone (CRH) and also enhance the transcription via an NRSE-independent mechanism [136].
2.7. Intelligence and Memory
Compared with mice, human-specific genes with REST occupancy were enriched in learning and memory functions [137,138]. In early life adversity (ELA) rats, REST contributes to the memory deficits and its blockage rescued spatial memory [139]. Additionally, in APP/PS1 mice, REST is essential for the maintenance of memory performance during aging [140]. The elevated REST by Pb explore inhibited SV2C affected synaptic plasticity, leading to impairment of learning and memory [141]. During the memory impairment induced by a flame retardant PBDE-209, REST was decreased and the inhibition of NR1 was released. CDRI-08, the ethanolic extract of a nootropic plant Bacopa monnieri, could attenuate the impairment and restore the binding of REST to NR1 promotor [142]. Interestingly, the early acute exposure to PBDE-209 interferes with the regulation of NMDAR1 by (cAMP-response element-binding protein) CREB and REST, however, long-term effects persist only in young males that could be associated with cognitive impairment [143]. Further, the reasoning skill were not affected by the VNTR and rs2228991 variants of REST in a young Chinese Han population [144].
2.8. Aging and Alzheimer’s Disease
The REST-RE1 machinery is well maintained during aging in rats [145]. By regulating autophagy and senescence in neurons, REST plays a protective role during physiological aging [146]. The homologous gene of REST in C. elegans, Spr3, is crucial for lifespan regulation, as its mutation and silencing led to shortened lifespan [147]. REST was elevated during neuronal senescence induced by high glucose and palmitic acid [148]. Upon peripheral nerve injury, REST was elevated in young mice but not in aged mice [149]. In Alzheimer’s disease (AD), the abnormally high REST affected ChAT expression, which is a major biochemical disorder in AD [150,151]. In human cortical and hippocampal neurons, REST was elevated during aging, but it was lost in mild cognitive impairment and AD. REST protects neurons from oxidative stress and amyloid β-protein toxicity [152]. Moreover, REST levels were also found to be associated with cognitive preservation and longevity [152].
2.9. Parkinson’s Disease
Similar to AD, REST enters the nucleus of aged dopaminergic neurons, and it is mostly absent in neurons of Parkinson’s disease (PD) [153]. REST and REST4 were increased in the neurotoxin 1-methyl-4-phenyl-pyridinium ion (MPP+) induced PD cell model [154]. REST KO mice presented higher sensitivity to the dopaminergic neurotoxin MPTP [155], while the neuroprotective effects of Trichostatin A (TSA) were mediated by REST in MPTP PD models [156]. REST deficiency aggravated dopaminergic neurodegeneration and impaired neurogenesis in mice with MPTP-induced PD [157,158]. Taken together, these findings suggest that REST was a negative modulator of neurogenesis and a pro-survival factor in the context of PD.
2.10. Huntington’s Disease
In Huntington’s disease (HD), the increased binding of REST to RE1 reduced its target gene expression. The binding was regulated by huntingtin, and the control was lost in HD, leading to the loss of neuronal gene expression [159,160,161]. The blockage of REST binding rescued BDNF [162]. Huntingtin and REST formed a complex to facilitate REST translocation to the nucleus [163]. The transcriptional regulation of REST by huntingtin interacting protein 1 protein interactor (HIPPI) contributed to the deregulation of transcription in HD [164]. The exon-3 skipping triggered by antisense oligonucleotides (ASOs) treatment rescued neuronal genes [165]. Additionally, the inhibition of mGluR5 could correct REST signaling via Wnt pathway [166]. Thus, targeting REST is a potential treatment for HD.
2.11. Epilepsy
The role of REST in epilepsy is controversial, and in the literature, opposite views coexist. The elevated REST during aging was abolished in patients with drug-resistant mesial temporal lobe epilepsy (MTLE) [167]. REST cKO exhibited dramatically accelerated seizure progression and prolonged afterdischarge duration [168]. However, several investigations suggested that REST is proepileptogenic. REST cKO mice showed higher resistance to convulsions induced by PTZ [169]. The interference with the chromatin binding of REST rescued spatial memory impaired by febrile status epilepticus [170]. In kainic acid-induced status epilepticus, the repression of the HCN1 channel was mediated by REST binding [171], and mild inactivation of REST reduces susceptibility [172]. The antiepileptic effect of 2-deoxy-d-glucose (2DG), which inhibits BDNF and its receptor TrkB to reduce the progression of kindling, was mediated by REST [50], and this effect was abolished in REST cKO mice [173]. The mutation of RILP could block its interaction with REST, contributing to the progressive myoclonus epilepsy progression [174]. REST and REST4 controlled the expression of preprotachykinin-A (TAC1), which encodes the neuropeptide substance P [175,176,177]. REST mediated the contribution of Sit1 to epileptogenesis [178]. Sirt1 reduced miR-124, which targeted REST mRNA [179]. These conflicting data suggest that REST plays a role in the occurrence and development of epilepsy, and further research is needed to understand the mechanism of regulatory differences in different models.
2.12. Ischemia
In ischemic damage to neurons, REST appears more like a mediator. Global ischemia induced REST expression to repress GluR2 and MOR-1, and REST knockdown protected neurons from ischemia-induced death [180,181,182,183]. Moreover, the elevated REST antagonized the CREB signaling on CART activation, leading to increased cell death [184]. Pyrvinium pamoate, an activator of casein kinase 1, can suppress REST expression and rescue neurons that would otherwise undergo cell death [185]. In addition, the neuron-specific miR-132 was a REST target mediating neuronal death upon ischemia [186]. It was discovered that the protective effects of melatonin on cerebral ischemia-reperfusion injury were mediated by miR-26a-5p-NRSF and JAK2-STAT3 pathway [187].
2.13. Psychiatric Disorders
REST contributes to genome-wide epigenetic changes and is implicated in schizophrenia [188]. The aberrant increase of REST at the Grin2b promoter caused deficient synaptic physiology and PFC-dependent cognitive dysfunction, a hallmark of schizophrenia [189]. THP2 is the rate-limiting enzyme for 5-HT, and it is under the regulation of REST [190]. The mood-stabilizing drugs lithium and sodium valproate have been shown to regulate REST expression [191,192]. miR-9 targeted REST and PDYN, and miR-9 was targeted by REST [193]. REST was elevated in care-augmented rats, and its target CRH was inhibited [194]. The compound C737 mimics the helical structure of REST to regulate target genes involved in neuronal function and pain modulation and reduce stress-induced weight loss in female Tupaia [195]. REST signaling contributes to stress resilience [196]. The improvement in diabetes mellitus-related cognitive impairment by 9-PAHSA was mediated by REST [197]. Repression of the aberrantly increased activity of REST might be beneficial in the prevention and treatment of psychiatric disorders.
3. REST in Other Systems
3.1. Heart
REST regulates heart development, structure, and function [198]. During embryonic heart development, the elevated REST inhibited an embryonic cardiac gene HCN4 [199,200,201,202]. The training-induced REST contributes to the pathological heart rate adaptation to exercise training by reducing HCN4 [203]. REST chronically represses T channels in cardiomyocytes [204]. REST also maintains low alpha 1H expression [205] and regulates CACNA1H encoding alpha-subunit of Cav3.2 [206], thereby regulating aldosterone and cortisol production, which are independent predictors of mortality risk for heart failure patients. Gαo, an inhibitory G protein encoded by GNAO1, was regulated by REST. In several mouse models of heart failure, Gαo is decreased in the ventricles, leading to increased surface sarcolemmal L-type Ca2+ channel activity and impairing Ca2+ handling in ventricular myocytes, resulting in cardiac dysfunction [207]. REST contributes to atrial fibrillation-associated remodeling of APD and K+ channel expression in cardiomyocytes [208]. In cardiac hypertrophy, depressed REST releases ANP and BNP [33,209]. ANP was under the regulation of REST in ventricular myocytes and is involved in the endothelin 1-induced increase of ANP [210]. Additionally, the dominant-negative REST induces hypertrophy by targeting TRP1 and regulating calcium entry [211]. REST is negatively regulated by Zfp90 development of cardiac dysfunction [212]. The important role of REST in cardiac physiology and pathology suggests it as a potential target for the treatment of heart diseases.
3.2. Pancreas
REST is absent in insulinoma, insulin- and glucagon-producing cells [213]. Similar to neuronal differentiation, REST is expressed in pancreas progenitors and decreased in differentiated endocrine cells [214]. RE1 motif was found within various genes involved in pancreas development, including pdx-1, Beta2/NeuroD, and pax4 [215]. Cx36 is also controlled by REST in insulin-producing cells [216]. In addition, the rs2518719 SNP could alter the RE1 motif involving pancreatic neuroendocrine tumor progression [217]. It was revealed that the REST silencing could induce human amniotic fluid-derived stem cells (hAFSCs) and bone marrow-derived mesenchymal stem cells (bmMSCs) to differentiate into insulin-producing cells [218,219]. The similar pattern of REST regulation in the pancreas to that of neurons inspired us to focus on their similarities and differences to help build upon the well-studied foundations of REST in the nervous system and provide insights into its role in pancreatic development and function.
3.3. Skin
REST is expressed in skin keratinocytes, and NRP1 was found to be under its regulation to promote cell migration by blocking the inhibition of semaphorin 3A [220]. During the early neural crest stage, REST is essential for melanoblast development [221]. In melanoma, the decreased REST/RE1 activity leads to aberrant metabotropic glutamate receptor 1 (mGluR1) expression, whose ectopic expression in mouse melanocytes was sufficient to induce melanoma development [222]. These findings provide valuable insights into the functions of REST in the skin, although much more research is needed to fully understand its role in skin biology and pathology.
3.4. Eye
The eye, being a specialized structure derived from the skin, also relies on the proper regulation of REST. In REST cKO mice generated by the Sox1-Cre allele and the floxed REST gene, REST was excised in the early neural tissues, including the lens and retinal primordia. The slightly reduced proliferation of lens epithelial cells after birth was observed, and vacuoles formed without inducing cell apoptosis. The augmented Notch signaling and the reduced lens fiber regulator genes may contribute [223]. The REST-like gene in Drosophila was named Chn, and it promotes D1 for photoreceptor cell development [224]. This further emphasizes the importance of REST as a regulator in the development and function of eye structures. Collectively, these findings indicate the role of REST in lens formation and the maintenance of the overall structure and function of the eye.
3.5. Vascular
In vascular tissue, REST was found to regulate the proliferation of vascular smooth muscle cells by targeting the K(Ca)3.1 (IKCa) potassium channel. REST was downregulated when there was cellular proliferation, indicating an inverse relationship with IKCa [225]. In addition, REST declines in the medial smooth muscle of atherosclerotic carotids, and REST overexpression inhibits smooth muscle migration. Inhibition of REST and overexpression of IKCa determine smooth muscle and matrix content of atherosclerotic lesions [226].
4. REST in Cancer
4.1. Nervous System Cancer
The dysregulation of REST was found in various nervous system cancers, often playing promoting roles. In neuroblastoma cells, REST was found to be reduced during differentiation [227]. REST was downregulated in mouse neuroblastoma, leading to increased neurite length [228]. It was found that the loss of lncRNA neuroblastoma-associated transcript-1 (NBAT-1) contributed to aggressive neuroblastoma by activating REST [229]. REST is highly expressed in glioma, and its interference inhibits tumors [230]. REST mediated the inhibition of proliferation of glioma by pioglitazone [231]. In medulloblastoma, REST promotes cancer progression via epigenetic modification and AKT activation [232]. The abnormal expression of REST in NSCs blocks their differentiation and promotes medulloblastoma formation [233]. The transient expression of a competitive form of REST, REST-VP16, inhibited medulloblastoma [234,235]. Additionally, inhibiting REST via LSD1 inhibition inhibited medulloblastoma migration [236]. Moreover, the elevated REST promotes vascular growth via autocrine and paracrine [237]. The frequent neuron-specific splicing of REST is involved in neuroblastoma progression [238], while REST activity was not affected by its mutations [239]. In certain cell lines, such as NS20Y and NIE115, REST4 has been identified as the major transcript [240]. Taken together, these findings highlight REST as a potential therapeutic target for nervous system cancers, offering opportunities for developing novel treatment approaches aimed at disrupting REST-mediated tumorigenic processes.
4.2. Non-Nervous System Cancer
The regulation pattern of REST varies in different types of lung cancers. In small-cell lung cancer cells (SCLCs), A splice variant of REST with a 50-bp insert predicting a truncated REST was highly expressed [241]. The decreased REST permits its target gene expression, including glycine receptor alpha1 (GLRA1) subunit, migration-promoting gene ITPKA [242,243,244], leading to de-regulation of AKT activation, promoting malignant progression [245]. The altered REST splicing was found to mediate the anti-tumor effects of SRRM4 targeting in SCLC [246]. Further, miR-9 inhibits the proliferation and migration of lung squamous cell carcinoma cells by blocking REST/EGFR signaling [247]. However, in neuroendocrine lung cancer, REST mediates neuroendocrine to non-neuroendocrine fate switch to play a pro-tumorigenic role [248]. REST-regulated SCG3 provides a sensitive prognostic biomarker for neuroendocrine lung cancer [249]. Similarly, in human non-small cell lung carcinoma cells (NSCLCs), depression of REST targets contributes to tumorigenicity [250]. REST also mediates nicotine-mediated invasion and migration [251]. REST is a potential marker for the diagnosis of lung cancer types and a potential target for the treatment of some types of lung cancers. These findings suggest that REST can serve as a potential marker for diagnosing different types of lung cancer and as a potential therapeutic target for specific lung cancer subtypes.
Unlike in nervous system cancers, the loss of REST repression occurs in most non-nervous system cancers. In colorectal cancer, REST plays a suppressive role. A frameshift mutation of REST encodes a dominantly acting truncation activating PI3K signaling [252], and rs968697 polymorphism in the HMGA2 gene influences REST binding [253]. In hepatocellular carcinoma (HCC), REST was found to be elevated in 27 of 49 human tissue samples, and it plays a key role in promoting diethylnitrosamine (DEN)-induced HCC initiation [254]. However, another study revealed that REST is decreased in cholangiocellular carcinoma but not in HCC when compared with normal liver tissues [255]. In breast cancer, REST was decreased, contributing to pathogenesis [256]. Additionally, REST is required for beta-estradiol (E2) stimulation of the cell cycle [257]. In Ewing sarcoma, EWSR1 interacted with REST, and EWSR1 reduced genes are a subset of neuronal genes that contain RE1 [258]. In uterine fibroids, REST represses GPR10 in the normal myometrium, and that the loss of REST in fibroids permits GPR10 expression [259]. In oral cancer, REST plays an important role in the cell survival by regulating the mTOR signaling pathway [260]. In the human prostate carcinoma cell line LNCaP, REST repressed IB1/JIP-1 to regulate the neuronal phenotype and JNK signaling pathway [261]. Furthermore, altered REST splicing was also found to mediate the anti-tumor effects of SRRM4 targeting in prostate cancer cells [246]. A recent study revealed REST diminution in premalignant lesions, squamous cell carcinoma (SCC), endocervical adenocarcinoma (ADC), and the cancerous cell lines SiHa and HeLa [262]. These investigations suggest that REST may be a hub that extensively regulates tumor occurrence and development and can serve as a diagnostic and therapeutic target. These investigations suggest that REST may serve as a central regulator in tumor occurrence and development, making it a potential diagnostic and therapeutic target in various types of cancer.
5. Targeting REST
Considering the important role of REST in health and disease, multiple strategies have been developed to inhibit or activate REST. Small interfering RNA (siRNA) is widely used to knockdown gene expression, and siRNA targeting REST could inhibit GBM cells and ovarian cancer cells [263,264,265]. In addition, the siRNA delivered by PEI@Fe3O4 nanoparticles induced MSC differentiation into insulin-producing cells [266]. In contrast to siRNA, the nuclear-localized small modulatory double-stranded RNA (smRNA) encoding NRSE sequences has been developed to activate REST target genes via smRNA/REST interaction, promoting neuronal differentiation [267,268]. Another approach involves the use of the recombinant transcription factor REST-VP16, which was constructed by replacing the repressor domains of REST with the activation domain of a viral activator VP16. It could induce neuronal differentiation in NSCs, myoblasts, and possibly other stem cells [74,269,270]. Small molecules have also been identified as REST modulators. For instance, the small molecule mS-11 was designed based on the mSin3-binding helix structure of REST. It inhibits mSin3-REST binding to reverse pain and autism in mice models [271,272,273]. The quinolone-like compound 91 (C91) was discovered by virtual screening. It reduces mSIN3b nuclear entry and occupancy at NRSE within BNDF to restore BDNF levels in HD cells [274]. Sertraline, chlorprothixene, and chlorpromazine were also found to interact with the REST-binding site of mSin3 to inhibit medulloblastoma [275]. Further, three 2-aminothiazole derivatives were found to modulate NRSE silencing activity [276]. Through high-throughput screening with luciferase-assay measuring REST activity, a benzoimidazole-5-carboxamide derivative X5050 was found to induce REST degradation, other than REST expression, splicing, or NRSE binding [277]. In addition, the small molecule compound C737 mimicking the helical structure of REST reduced stress-induced weight loss in female Tupaia [195]. The optogenetic approach mediated REST inhibition was achieved using the fusion of the inhibitory domains that impair REST-DNA binding or recruitment of the cofactor mSin3a to the light-sensitive Avena sativa light-oxygen-voltage-sensing 2-phototrophin 1 to modulate REST target genes [278]. Recently, an approach combining biochemical experiments and molecular dynamics simulations was applied to develop ad hoc designed RNA binding proteins to specifically target and dock to REST mRNA [279]. These diverse regulatory strategies provide a solid foundation for further investigations and drug development targeting REST, opening up novel possibilities for therapeutic interventions in REST-related conditions.
6. Concluding Remarks
REST is a key transcription regulator that primarily governs gene transcription via the modification of chromatin. REST plays vital roles in both physiological and pathological processes within the nervous system as well as other non-nervous systems. The imbalance of REST dynamic regulation often leads to dysplasia or diseases. Therefore, it is necessary to conduct in-depth research on its involvement in different diseases and the underlying mechanisms is imperative to gain novel insights into potential interventions for related diseases.
The patterns of interaction between REST and its target genes, as well as the mechanisms underlying gene transcription regulation, have been identified by current research. The dynamic regulation of REST expression and gene targeting during development and in various diseases remains largely unknown. However, advancements in technologies such as single cell sequencing, spatial transcriptome, structural biology, and artificial intelligence hold promise in acquiring more relevant information. Integrating these findings with cellular and molecular biology technologies will strongly enhance our understanding of the mechanism of transcription factors, including REST.
REST plays a pivotal role in early development, and its regulation exhibits robustness with upstream bias. While many investigations have focused on understanding the regulation of target genes by REST, there is still much to be discovered regarding the upstream regulatory mechanisms governing REST transcription, translation, and activity. Unraveling these aspects would contribute to the construction of comprehensive gene regulatory networks centered around REST.
Different antibodies target different antigenic epitopes, and different REST isoforms are post-translationally modified, making it difficult to discriminate between non-specific bands and REST isoforms on the basis of band position in Western blotting data. Simply cropping out bands that do not match the expected molecular weight is likely to miss a lot of important information. In addition, the use of currently unavailable REST antibodies in some studies might lead to non-reproducible or misleading results due to, for example, poor specificity [3]. Therefore, the specificity of the REST antibody and the multiple bands in the blotting are strongly recommended to be cautioned in cropping and analysis.
In conclusion, this review highlights the mechanisms of REST and the progress made in understanding its roles in health and disease. It aims to provide assistance in comprehending the mode of action and exploring potential interventions for the related diseases. REST is a fascinating gene that merits further investigation, and we eagerly anticipate more interdisciplinary research results and breakthroughs in the field of REST.
Author Contributions
Conceptualization, D.Z.; writing—original draft preparation, L.J., Y.L. and D.Z.; writing—review and editing, Y.W. and Y.H.; funding acquisition, D.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant numbers 81703102 and 82073419.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chong, J.A.; Tapia-Ramirez, J.; Kim, S.; Toledo-Aral, J.J.; Zheng, Y.; Boutros, M.C.; Altshuller, Y.M.; Frohman, M.A.; Kraner, S.D.; Mandel, G. REST: A mammalian silencer protein that restricts sodium channel gene expression to neurons. Cell 1995, 80, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Schoenherr, C.J.; Anderson, D.J. The neuron-restrictive silencer factor (NRSF): A coordinate repressor of multiple neuron-specific genes. Science 1995, 267, 1360–1363. [Google Scholar] [CrossRef]
- Chen, G.L.; Miller, G.M. Alternative REST Splicing Underappreciated. Eneuro 2018, 5, 1–6. [Google Scholar] [CrossRef]
- Shimojo, M.; Lee, J.H.; Hersh, L.B. Role of zinc finger domains of the transcription factor neuron-restrictive silencer factor/repressor element-1 silencing transcription factor in DNA binding and nuclear localization. J. Biol. Chem. 2001, 276, 13121–13126. [Google Scholar] [CrossRef] [PubMed]
- Shimojo, M. Characterization of the nuclear targeting signal of REST/NRSF. Neurosci. Lett. 2006, 398, 161–166. [Google Scholar] [CrossRef]
- Shimojo, M.; Hersh, L.B. Characterization of the REST/NRSF-interacting LIM domain protein (RILP): Localization and interaction with REST/NRSF. J. Neurochem. 2006, 96, 1130–1138. [Google Scholar] [CrossRef]
- Lee, J.H.; Shimojo, M.; Chai, Y.G.; Hersh, L.B. Studies on the interaction of REST4 with the cholinergic repressor element-1/neuron restrictive silencer element. Brain Res. Mol. Brain Res. 2000, 80, 88–98. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, W.; Shen, J.; Tong, X.; Yang, Z.; Shen, Z.; Lan, W.; Wu, H.; Cao, C. Cysteine 397 plays important roles in the folding of the neuron-restricted silencer factor/RE1-silencing transcription factor. Biochem. Biophys. Res. Commun. 2011, 414, 309–314. [Google Scholar] [CrossRef]
- Kraner, S.D.; Chong, J.A.; Tsay, H.J.; Mandel, G. Silencing the type II sodium channel gene: A model for neural-specific gene regulation. Neuron 1992, 9, 37–44. [Google Scholar] [CrossRef]
- Mori, N.; Schoenherr, C.; Vandenbergh, D.J.; Anderson, D.J. A common silencer element in the SCG10 and type II Na+ channel genes binds a factor present in nonneuronal cells but not in neuronal cells. Neuron 1992, 9, 45–54. [Google Scholar] [CrossRef]
- Li, L.; Suzuki, T.; Mori, N.; Greengard, P. Identification of a functional silencer element involved in neuron-specific expression of the synapsin I gene. Proc. Natl. Acad. Sci. USA 1993, 90, 1460–1464. [Google Scholar] [CrossRef]
- Schoenherr, C.J.; Paquette, A.J.; Anderson, D.J. Identification of potential target genes for the neuron-restrictive silencer factor. Proc. Natl. Acad. Sci. USA 1996, 93, 9881–9886. [Google Scholar] [CrossRef]
- Bruce, A.W.; Donaldson, I.J.; Wood, I.C.; Yerbury, S.A.; Sadowski, M.I.; Chapman, M.; Gottgens, B.; Buckley, N.J. Genome-wide analysis of repressor element 1 silencing transcription factor/neuron-restrictive silencing factor (REST/NRSF) target genes. Proc. Natl. Acad. Sci. USA 2004, 101, 10458–10463. [Google Scholar] [CrossRef]
- Sun, Y.M.; Greenway, D.J.; Johnson, R.; Street, M.; Belyaev, N.D.; Deuchars, J.; Bee, T.; Wilde, S.; Buckley, N.J. Distinct profiles of REST interactions with its target genes at different stages of neuronal development. Mol. Biol. Cell 2005, 16, 5630–5638. [Google Scholar] [CrossRef]
- Johnson, D.S.; Mortazavi, A.; Myers, R.M.; Wold, B. Genome-wide mapping of in vivo protein-DNA interactions. Science 2007, 316, 1497–1502. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, M.; Niu, G.; Cheng, Y.; Fei, J. Genome-wide identification of target genes repressed by the zinc finger transcription factor REST/NRSF in the HEK 293 cell line. Acta Biochim. Biophys. Sin. 2009, 41, 1008–1017. [Google Scholar] [CrossRef]
- Sedaghat, Y.; Bui, H.H.; Mazur, C.; Monia, B.P. Identification of REST-regulated genes and pathways using a REST-targeted antisense approach. Nucleic Acid Ther. 2013, 23, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Teh, C.H.; Kunarso, G.; Wong, K.Y.; Srinivasan, G.; Cooper, M.L.; Volta, M.; Chan, S.S.; Lipovich, L.; Pollard, S.M.; et al. REST regulates distinct transcriptional networks in embryonic and neural stem cells. PLoS Biol. 2008, 6, e256. [Google Scholar] [CrossRef]
- Mortazavi, A.; Leeper Thompson, E.C.; Garcia, S.T.; Myers, R.M.; Wold, B. Comparative genomics modeling of the NRSF/REST repressor network: From single conserved sites to genome-wide repertoire. Genome Res. 2006, 16, 1208–1221. [Google Scholar] [CrossRef]
- Ding, Y.; Lorenz, W.A.; Chuang, J.H. CodingMotif: Exact determination of overrepresented nucleotide motifs in coding sequences. BMC Bioinform. 2012, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Jothi, R.; Cuddapah, S.; Barski, A.; Cui, K.; Zhao, K. Genome-wide identification of in vivo protein-DNA binding sites from ChIP-Seq data. Nucleic Acids Res. 2008, 36, 5221–5231. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Jiang, H.; Ma, W.; Johnson, D.S.; Myers, R.M.; Wong, W.H. An integrated software system for analyzing ChIP-chip and ChIP-seq data. Nat. Biotechnol. 2008, 26, 1293–1300. [Google Scholar] [CrossRef]
- Valouev, A.; Johnson, D.S.; Sundquist, A.; Medina, C.; Anton, E.; Batzoglou, S.; Myers, R.M.; Sidow, A. Genome-wide analysis of transcription factor binding sites based on ChIP-Seq data. Nat. Methods 2008, 5, 829–834. [Google Scholar] [CrossRef]
- Nix, D.A.; Courdy, S.J.; Boucher, K.M. Empirical methods for controlling false positives and estimating confidence in ChIP-Seq peaks. BMC Bioinform. 2008, 9, 523. [Google Scholar] [CrossRef]
- Boeva, V.; Surdez, D.; Guillon, N.; Tirode, F.; Fejes, A.P.; Delattre, O.; Barillot, E. De novo motif identification improves the accuracy of predicting transcription factor binding sites in ChIP-Seq data analysis. Nucleic Acids Res. 2010, 38, e126. [Google Scholar] [CrossRef]
- McLean, C.Y.; Bristor, D.; Hiller, M.; Clarke, S.L.; Schaar, B.T.; Lowe, C.B.; Wenger, A.M.; Bejerano, G. GREAT improves functional interpretation of cis-regulatory regions. Nat. Biotechnol. 2010, 28, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.K.; Jayatillake, R.V.; Spouge, J.L. NEXT-peak: A normal-exponential two-peak model for peak-calling in ChIP-seq data. BMC Genom. 2013, 14, 349. [Google Scholar] [CrossRef]
- Yu, H.B.; Johnson, R.; Kunarso, G.; Stanton, L.W. Coassembly of REST and its cofactors at sites of gene repression in embryonic stem cells. Genome Res. 2011, 21, 1284–1293. [Google Scholar] [CrossRef]
- Grimes, J.A.; Nielsen, S.J.; Battaglioli, E.; Miska, E.A.; Speh, J.C.; Berry, D.L.; Atouf, F.; Holdener, B.C.; Mandel, G.; Kouzarides, T. The co-repressor mSin3A is a functional component of the REST-CoREST repressor complex. J. Biol. Chem. 2000, 275, 9461–9467. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Myers, S.J.; Dingledine, R. Transcriptional repression by REST: Recruitment of Sin3A and histone deacetylase to neuronal genes. Nat. Neurosci. 1999, 2, 867–872. [Google Scholar] [CrossRef]
- Roopra, A.; Sharling, L.; Wood, I.C.; Briggs, T.; Bachfischer, U.; Paquette, A.J.; Buckley, N.J. Transcriptional repression by neuron-restrictive silencer factor is mediated via the Sin3-histone deacetylase complex. Mol. Cell Biol. 2000, 20, 2147–2157. [Google Scholar] [CrossRef]
- Adams, G.E.; Chandru, A.; Cowley, S.M. Co-repressor, co-activator and general transcription factor: The many faces of the Sin3 histone deacetylase (HDAC) complex. Biochem. J. 2018, 475, 3921–3932. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Kuwahara, K.; Harada, M.; Takahashi, N.; Yasuno, S.; Adachi, Y.; Kawakami, R.; Nakanishi, M.; Tanimoto, K.; Usami, S.; et al. Class II HDACs mediate CaMK-dependent signaling to NRSF in ventricular myocytes. J. Mol. Cell. Cardiol. 2006, 41, 1010–1022. [Google Scholar] [CrossRef]
- Naruse, Y.; Aoki, T.; Kojima, T.; Mori, N. Neural restrictive silencer factor recruits mSin3 and histone deacetylase complex to repress neuron-specific target genes. Proc. Natl. Acad. Sci. USA 1999, 96, 13691–13696. [Google Scholar] [CrossRef]
- Ooi, L.; Wood, I.C. Chromatin crosstalk in development and disease: Lessons from REST. Nat. Rev. Genet. 2007, 8, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Uda-Tochio, H.; Murai, K.; Mori, N.; Nishimura, Y. The neural repressor NRSF/REST binds the PAH1 domain of the Sin3 corepressor by using its distinct short hydrophobic helix. J. Mol. Biol. 2005, 354, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Higo, J.; Nishimura, Y.; Nakamura, H. A free-energy landscape for coupled folding and binding of an intrinsically disordered protein in explicit solvent from detailed all-atom computations. J. Am. Chem. Soc. 2011, 133, 10448–10458. [Google Scholar] [CrossRef]
- Chandru, A.; Bate, N.; Vuister, G.W.; Cowley, S.M. Sin3A recruits Tet1 to the PAH1 domain via a highly conserved Sin3-Interaction Domain. Sci. Rep. 2018, 8, 14689. [Google Scholar] [CrossRef] [PubMed]
- Perera, A.; Eisen, D.; Wagner, M.; Laube, S.K.; Kunzel, A.F.; Koch, S.; Steinbacher, J.; Schulze, E.; Splith, V.; Mittermeier, N.; et al. TET3 is recruited by REST for context-specific hydroxymethylation and induction of gene expression. Cell Rep. 2015, 11, 283–294. [Google Scholar] [CrossRef]
- Yeo, M.; Lee, S.K.; Lee, B.; Ruiz, E.C.; Pfaff, S.L.; Gill, G.N. Small CTD phosphatases function in silencing neuronal gene expression. Science 2005, 307, 596–600. [Google Scholar] [CrossRef]
- Andres, M.E.; Burger, C.; Peral-Rubio, M.J.; Battaglioli, E.; Anderson, M.E.; Grimes, J.; Dallman, J.; Ballas, N.; Mandel, G. CoREST: A functional corepressor required for regulation of neural-specific gene expression. Proc. Natl. Acad. Sci. USA 1999, 96, 9873–9878. [Google Scholar] [CrossRef] [PubMed]
- Lunyak, V.V.; Burgess, R.; Prefontaine, G.G.; Nelson, C.; Sze, S.H.; Chenoweth, J.; Schwartz, P.; Pevzner, P.A.; Glass, C.; Mandel, G.; et al. Corepressor-dependent silencing of chromosomal regions encoding neuronal genes. Science 2002, 298, 1747–1752. [Google Scholar] [CrossRef] [PubMed]
- Ballas, N.; Mandel, G. The many faces of REST oversee epigenetic programming of neuronal genes. Curr. Opin. Neurobiol. 2005, 15, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Roopra, A.; Qazi, R.; Schoenike, B.; Daley, T.J.; Morrison, J.F. Localized domains of G9a-mediated histone methylation are required for silencing of neuronal genes. Mol. Cell 2004, 14, 727–738. [Google Scholar] [CrossRef]
- Ding, N.; Tomomori-Sato, C.; Sato, S.; Conaway, R.C.; Conaway, J.W.; Boyer, T.G. MED19 and MED26 are synergistic functional targets of the RE1 silencing transcription factor in epigenetic silencing of neuronal gene expression. J. Biol. Chem. 2009, 284, 2648–2656. [Google Scholar] [CrossRef]
- Ding, N.; Zhou, H.; Esteve, P.O.; Chin, H.G.; Kim, S.; Xu, X.; Joseph, S.M.; Friez, M.J.; Schwartz, C.E.; Pradhan, S.; et al. Mediator links epigenetic silencing of neuronal gene expression with x-linked mental retardation. Mol. Cell 2008, 31, 347–359. [Google Scholar] [CrossRef]
- Shi, Y.; Lan, F.; Matson, C.; Mulligan, P.; Whetstine, J.R.; Cole, P.A.; Casero, R.A.; Shi, Y. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell 2004, 119, 941–953. [Google Scholar] [CrossRef]
- Ooi, L.; Belyaev, N.D.; Miyake, K.; Wood, I.C.; Buckley, N.J. BRG1 chromatin remodeling activity is required for efficient chromatin binding by repressor element 1-silencing transcription factor (REST) and facilitates REST-mediated repression. J. Biol. Chem. 2006, 281, 38974–38980. [Google Scholar] [CrossRef]
- Shi, Y.; Sawada, J.; Sui, G.; Affarel, B.; Whetstine, J.R.; Lan, F.; Ogawa, H.; Luke, M.P.; Nakatani, Y.; Shi, Y. Coordinated histone modifications mediated by a CtBP co-repressor complex. Nature 2003, 422, 735–738. [Google Scholar] [CrossRef]
- Garriga-Canut, M.; Schoenike, B.; Qazi, R.; Bergendahl, K.; Daley, T.J.; Pfender, R.M.; Morrison, J.F.; Ockuly, J.; Stafstrom, C.; Sutula, T.; et al. 2-Deoxy-D-glucose reduces epilepsy progression by NRSF-CtBP-dependent metabolic regulation of chromatin structure. Nat. Neurosci. 2006, 9, 1382–1387. [Google Scholar] [CrossRef]
- Ballas, N.; Grunseich, C.; Lu, D.D.; Speh, J.C.; Mandel, G. REST and its corepressors mediate plasticity of neuronal gene chromatin throughout neurogenesis. Cell 2005, 121, 645–657. [Google Scholar] [CrossRef]
- Chen, Z.F.; Paquette, A.J.; Anderson, D.J. NRSF/REST is required in vivo for repression of multiple neuronal target genes during embryogenesis. Nat. Genet. 1998, 20, 136–142. [Google Scholar] [CrossRef]
- Wang, Y.C.; Liu, P.; Yue, L.Y.; Huang, F.; Xu, Y.X.; Zhu, C.Q. NRSF deficiency leads to abnormal postnatal development of dentate gyrus and impairment of progenitors in subgranular zone of hippocampus. Hippocampus 2021, 31, 935–9563. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Lee, Y.K.; Lim, Y.C.; Zheng, Z.; Lin, X.M.; Ng, D.P.; Holbrook, J.D.; Law, H.Y.; Kwek, K.Y.; Yeo, G.S.; et al. Global DNA hypermethylation in down syndrome placenta. PLoS Genet. 2013, 9, e1003515. [Google Scholar] [CrossRef]
- El Hajj, N.; Dittrich, M.; Bock, J.; Kraus, T.F.; Nanda, I.; Muller, T.; Seidmann, L.; Tralau, T.; Galetzka, D.; Schneider, E.; et al. Epigenetic dysregulation in the developing down syndrome cortex. Epigenetics 2016, 11, 563–578. [Google Scholar] [CrossRef] [PubMed]
- Canzonetta, C.; Mulligan, C.; Deutsch, S.; Ruf, S.; O’Doherty, A.; Lyle, R.; Borel, C.; Lin-Marq, N.; Delom, F.; Groet, J.; et al. DYRK1A-dosage imbalance perturbs NRSF/REST levels, deregulating pluripotency and embryonic stem cell fate in Down syndrome. Am. J. Hum. Genet. 2008, 83, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Lepagnol-Bestel, A.M.; Zvara, A.; Maussion, G.; Quignon, F.; Ngimbous, B.; Ramoz, N.; Imbeaud, S.; Loe-Mie, Y.; Benihoud, K.; Agier, N.; et al. DYRK1A interacts with the REST/NRSF-SWI/SNF chromatin remodelling complex to deregulate gene clusters involved in the neuronal phenotypic traits of Down syndrome. Hum. Mol. Genet. 2009, 18, 1405–1414. [Google Scholar] [CrossRef]
- Nishihara, S.; Tsuda, L.; Ogura, T. The canonical Wnt pathway directly regulates NRSF/REST expression in chick spinal cord. Biochem. Biophys. Res. Commun. 2003, 311, 55–63. [Google Scholar] [CrossRef]
- Li, H.; Malbon, C.C.; Wang, H.Y. Gene profiling of Frizzled-1 and Frizzled-2 signaling: Expression of G-protein-coupled receptor chimeras in mouse F9 teratocarcinoma embryonal cells. Mol. Pharmacol. 2004, 65, 45–55. [Google Scholar] [CrossRef]
- Gates, K.P.; Mentzer, L.; Karlstrom, R.O.; Sirotkin, H.I. The transcriptional repressor REST/NRSF modulates hedgehog signaling. Dev. Biol. 2010, 340, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ren, J.; Wang, Z.; Yao, J.; Fei, J. NRSF/REST is required for gastrulation and neurogenesis during zebrafish development. Acta Biochim. Biophys. Sin. 2012, 44, 385–393. [Google Scholar] [CrossRef]
- Olguin, P.; Oteiza, P.; Gamboa, E.; Gomez-Skarmeta, J.L.; Kukuljan, M. RE-1 silencer of transcription/neural restrictive silencer factor modulates ectodermal patterning during Xenopus development. J. Neurosci. 2006, 26, 2820–2829. [Google Scholar] [CrossRef]
- Arora, D.K.; Cosgrave, A.S.; Howard, M.R.; Bubb, V.; Quinn, J.P.; Thippeswamy, T. Evidence of postnatal neurogenesis in dorsal root ganglion: Role of nitric oxide and neuronal restrictive silencer transcription factor. J. Mol. Neurosci. 2007, 32, 97–107. [Google Scholar] [CrossRef]
- Kok, F.O.; Taibi, A.; Wanner, S.J.; Xie, X.; Moravec, C.E.; Love, C.E.; Prince, V.E.; Mumm, J.S.; Sirotkin, H.I. Zebrafish rest regulates developmental gene expression but not neurogenesis. Development 2012, 139, 3838–3848. [Google Scholar] [CrossRef]
- Moravec, C.E.; Li, E.; Maaswinkel, H.; Kritzer, M.F.; Weng, W.; Sirotkin, H.I. Rest mutant zebrafish swim erratically and display atypical spatial preferences. Behav. Brain Res. 2015, 284, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Hara, A.; Era, T.; Kunisada, T.; Yamada, Y. Genetic ablation of Rest leads to in vitro-specific derepression of neuronal genes during neurogenesis. Development 2012, 139, 667–677. [Google Scholar] [CrossRef]
- Cheng, Y.; Yin, Y.; Zhang, A.; Bernstein, A.M.; Kawaguchi, R.; Gao, K.; Potter, K.; Gilbert, H.Y.; Ao, Y.; Ou, J.; et al. Transcription factor network analysis identifies REST/NRSF as an intrinsic regulator of CNS regeneration in mice. Nat. Commun. 2022, 13, 4418. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Kagalwala, M.N.; Parker-Thornburg, J.; Adams, H.; Majumder, S. REST maintains self-renewal and pluripotency of embryonic stem cells. Nature 2008, 453, 223–227. [Google Scholar] [CrossRef]
- Gupta, S.K.; Gressens, P.; Mani, S. NRSF downregulation induces neuronal differentiation in mouse embryonic stem cells. Differentiation 2009, 77, 19–28. [Google Scholar] [CrossRef]
- Thakore-Shah, K.; Koleilat, T.; Jan, M.; John, A.; Pyle, A.D. REST/NRSF Knockdown Alters Survival, Lineage Differentiation and Signaling in Human Embryonic Stem Cells. PLoS ONE 2015, 10, e0145280. [Google Scholar] [CrossRef]
- Tyler, C.R.; Labrecque, M.T.; Solomon, E.R.; Guo, X.; Allan, A.M. Prenatal arsenic exposure alters REST/NRSF and microRNA regulators of embryonic neural stem cell fate in a sex-dependent manner. Neurotoxicol. Teratol. 2017, 59, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Satoh, J.; Kawana, N.; Yamamoto, Y. ChIP-Seq Data Mining: Remarkable Differences in NRSF/REST Target Genes between Human ESC and ESC-Derived Neurons. Bioinform. Biol. Insights 2013, 7, 357–368. [Google Scholar] [CrossRef]
- Gao, Z.; Ure, K.; Ding, P.; Nashaat, M.; Yuan, L.; Ma, J.; Hammer, R.E.; Hsieh, J. The master negative regulator REST/NRSF controls adult neurogenesis by restraining the neurogenic program in quiescent stem cells. J. Neurosci. 2011, 31, 9772–9786. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Kameoka, S.; Lentz, S.; Majumder, S. Activation of REST/NRSF target genes in neural stem cells is sufficient to cause neuronal differentiation. Mol. Cell. Biol. 2004, 24, 8018–8025. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Zhuang, Z.; Liu, A.; Chai, Y.; Hoffman, P.W. The role of the RE1 element in activation of the NR1 promoter during neuronal differentiation. J. Neurochem. 2003, 86, 992–1005. [Google Scholar] [CrossRef]
- Kim, S.M.; Yang, J.W.; Park, M.J.; Lee, J.K.; Kim, S.U.; Lee, Y.S.; Lee, M.A. Regulation of human tyrosine hydroxylase gene by neuron-restrictive silencer factor. Biochem. Biophys. Res. Commun. 2006, 346, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Abrajano, J.J.; Qureshi, I.A.; Gokhan, S.; Zheng, D.; Bergman, A.; Mehler, M.F. Differential deployment of REST and CoREST promotes glial subtype specification and oligodendrocyte lineage maturation. PLoS ONE 2009, 4, e7665. [Google Scholar] [CrossRef]
- Kohyama, J.; Sanosaka, T.; Tokunaga, A.; Takatsuka, E.; Tsujimura, K.; Okano, H.; Nakashima, K. BMP-induced REST regulates the establishment and maintenance of astrocytic identity. J. Cell Biol. 2010, 189, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, Y.; Lv, Y.; Zhang, S.; Chen, L.; Bai, C.; Nan, X.; Yue, W.; Pei, X. NRSF silencing induces neuronal differentiation of human mesenchymal stem cells. Exp. Cell Res. 2008, 314, 2257–2265. [Google Scholar] [CrossRef]
- Kim, H.J.; Denli, A.M.; Wright, R.; Baul, T.D.; Clemenson, G.D.; Morcos, A.S.; Zhao, C.; Schafer, S.T.; Gage, F.H.; Kagalwala, M.N. REST Regulates Non-Cell-Autonomous Neuronal Differentiation and Maturation of Neural Progenitor Cells via Secretogranin II. J. Neurosci. 2015, 35, 14872–14884. [Google Scholar] [CrossRef]
- Zhao, X.; Glass, Z.; Chen, J.; Yang, L.; Kaplan, D.L.; Xu, Q. mRNA Delivery Using Bioreducible Lipidoid Nanoparticles Facilitates Neural Differentiation of Human Mesenchymal Stem Cells. Adv. Healthc. Mater. 2021, 10, e2000938. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Hara, A.; Oomori, Y.; Shimizu, Y.; Yamada, Y.; Kunisada, T. Neonatal lethality of neural crest cell-specific Rest knockout mice is associated with gastrointestinal distension caused by aberrations of myenteric plexus. Genes Cells 2014, 19, 723–742. [Google Scholar] [CrossRef] [PubMed]
- Tomasoni, R.; Negrini, S.; Fiordaliso, S.; Klajn, A.; Tkatch, T.; Mondino, A.; Meldolesi, J.; D’Alessandro, R. A signaling loop of REST, TSC2 and beta-catenin governs proliferation and function of PC12 neural cells. J. Cell Sci. 2011, 124, 3174–3186. [Google Scholar] [CrossRef]
- Toshiyuki, N.; Ichiro, M. Molecular mechanisms regulating cell type specific expression of BMP/RA Inducible Neural-specific Protein-1 that suppresses cell cycle progression: Roles of NRSF/REST and DNA methylation. Brain Res. Mol. Brain Res. 2004, 125, 47–59. [Google Scholar] [CrossRef]
- Yoo, A.S.; Staahl, B.T.; Chen, L.; Crabtree, G.R. MicroRNA-mediated switching of chromatin-remodelling complexes in neural development. Nature 2009, 460, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, A.; Yamada, T.; Sasagawa, S.; Naruse, Y.; Mori, N.; Tsuda, M. REST4-mediated modulation of REST/NRSF-silencing function during BDNF gene promoter activation. Biochem. Biophys. Res. Commun. 2002, 290, 415–420. [Google Scholar] [CrossRef]
- Qiang, M.; Rani, C.S.; Ticku, M.K. Neuron-restrictive silencer factor regulates the N-methyl-D-aspartate receptor 2B subunit gene in basal and ethanol-induced gene expression in fetal cortical neurons. Mol. Pharmacol. 2005, 67, 2115–2125. [Google Scholar] [CrossRef] [PubMed]
- Tateno, M.; Ukai, W.; Hashimoto, E.; Ikeda, H.; Saito, T. Implication of increased NRSF/REST binding activity in the mechanism of ethanol inhibition of neuronal differentiation. J. Neural Transm. 2006, 113, 283–293. [Google Scholar] [CrossRef]
- Tateno, M.; Saito, T. Biological studies on alcohol-induced neuronal damage. Psychiatry Investig. 2008, 5, 21–27. [Google Scholar] [CrossRef]
- Cai, L.; Bian, M.; Liu, M.; Sheng, Z.; Suo, H.; Wang, Z.; Huang, F.; Fei, J. Ethanol-induced neurodegeneration in NRSF/REST neuronal conditional knockout mice. Neuroscience 2011, 181, 196–205. [Google Scholar] [CrossRef]
- Formisano, L.; Guida, N.; Laudati, G.; Boscia, F.; Esposito, A.; Secondo, A.; Di Renzo, G.; Canzoniero, L.M. Extracellular signal-related kinase 2/specificity protein 1/specificity protein 3/repressor element-1 silencing transcription factor pathway is involved in Aroclor 1254-induced toxicity in SH-SY5Y neuronal cells. J. Neurosci. Res. 2015, 93, 167–177. [Google Scholar] [CrossRef]
- Rivera-Cervantes, M.C.; Castaneda-Arellano, R.; Castro-Torres, R.D.; Gudino-Cabrera, G.; Feria y Velasco, A.I.; Camins, A.; Beas-Zarate, C. P38 MAPK inhibition protects against glutamate neurotoxicity and modifies NMDA and AMPA receptor subunit expression. J. Mol. Neurosci. 2015, 55, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.; Rizor, A.; Son, D.S.; Aschner, M.; Lee, E. The transcription factor REST up-regulates tyrosine hydroxylase and antiapoptotic genes and protects dopaminergic neurons against manganese toxicity. J. Biol. Chem. 2020, 295, 3040–3054. [Google Scholar] [CrossRef]
- Kallunki, P.; Edelman, G.M.; Jones, F.S. Tissue-specific expression of the L1 cell adhesion molecule is modulated by the neural restrictive silencer element. J. Cell Biol. 1997, 138, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Schulte, C.; Racchetti, G.; D’Alessandro, R.; Meldolesi, J. A new form of neurite outgrowth sustained by the exocytosis of enlargeosomes expressed under the control of REST. Traffic 2010, 11, 1304–1314. [Google Scholar] [CrossRef] [PubMed]
- Paquette, A.J.; Perez, S.E.; Anderson, D.J. Constitutive expression of the neuron-restrictive silencer factor (NRSF)/REST in differentiating neurons disrupts neuronal gene expression and causes axon pathfinding errors in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 12318–12323. [Google Scholar] [CrossRef]
- Schoch, S.; Cibelli, G.; Thiel, G. Neuron-specific gene expression of synapsin I. Major role of a negative regulatory mechanism. J. Biol. Chem. 1996, 271, 3317–3323. [Google Scholar] [CrossRef]
- Chin, L.S.; Weigel, C.; Li, L. Transcriptional regulation of gene expression of sec6, a component of mammalian exocyst complex at the synapse. Brain Res. Mol. Brain Res. 2000, 79, 127–137. [Google Scholar] [CrossRef]
- Pozzi, D.; Menna, E.; Canzi, A.; Desiato, G.; Mantovani, C.; Matteoli, M. The Communication Between the Immune and Nervous Systems: The Role of IL-1β in Synaptopathies. Front. Mol. Neurosci. 2018, 11, 111. [Google Scholar] [CrossRef]
- Buffolo, F.; Petrosino, V.; Albini, M.; Moschetta, M.; Carlini, F.; Floss, T.; Kerlero de Rosbo, N.; Cesca, F.; Rocchi, A.; Uccelli, A.; et al. Neuroinflammation induces synaptic scaling through IL-1β-mediated activation of the transcriptional repressor REST/NRSF. Cell Death Dis. 2021, 12, 180. [Google Scholar] [CrossRef]
- Bersten, D.C.; Wright, J.A.; McCarthy, P.J.; Whitelaw, M.L. Regulation of the neuronal transcription factor NPAS4 by REST and microRNAs. Biochim. Biophys. Acta 2014, 1839, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Prestigio, C.; Ferrante, D.; Marte, A.; Romei, A.; Lignani, G.; Onofri, F.; Valente, P.; Benfenati, F.; Baldelli, P. REST/NRSF drives homeostatic plasticity of inhibitory synapses in a target-dependent fashion. eLife 2021, 10, e69058. [Google Scholar] [CrossRef]
- Moon, S.M.; Kim, J.S.; Park, B.R.; Kim, D.K.; Kim, S.G.; Kim, H.J.; Chun, H.S.; Lee, B.K.; Kim, C.S. Transcriptional regulation of the neuropeptide VGF by the neuron-restrictive silencer factor/neuron-restrictive silencer element. NeuroReport 2015, 26, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, D.; Lignani, G.; Ferrea, E.; Contestabile, A.; Paonessa, F.; D’Alessandro, R.; Lippiello, P.; Boido, D.; Fassio, A.; Meldolesi, J.; et al. REST/NRSF-mediated intrinsic homeostasis protects neuronal networks from hyperexcitability. EMBO J. 2013, 32, 2994–3007. [Google Scholar] [CrossRef]
- Drews, V.L.; Shi, K.; de Haan, G.; Meisler, M.H. Identification of evolutionarily conserved, functional noncoding elements in the promoter region of the sodium channel gene SCN8A. Mamm. Genome 2007, 18, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Ariano, P.; Zamburlin, P.; D’Alessandro, R.; Meldolesi, J.; Lovisolo, D. Differential repression by the transcription factor REST/NRSF of the various Ca2+ signalling mechanisms in pheochromocytoma PC12 cells. Cell Calcium 2010, 47, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Antoniotti, S.; Ruffinatti, F.A.; Torriano, S.; Luganini, A.; D’Alessandro, R.; Lovisolo, D. REST levels affect the functional expression of voltage dependent calcium channels and the migratory activity in immortalized GnRH neurons. Neurosci. Lett. 2016, 629, 19–25. [Google Scholar] [CrossRef]
- Kim, D.S. No effect of expression of neuron-restrictive silencer factor (NRSF) protein on N-type Ca2+ channel alpha 1B gene promoter activity in NS20Y cells. Mol. Cells 1998, 8, 600–605. [Google Scholar]
- Centonze, E.; Marte, A.; Albini, M.; Rocchi, A.; Cesca, F.; Chiacchiaretta, M.; Floss, T.; Baldelli, P.; Ferroni, S.; Benfenati, F.; et al. Neuron-restrictive silencer factor/repressor element 1-silencing transcription factor (NRSF/REST) controls spatial K(+) buffering in primary cortical astrocytes. J. Neurochem. 2023, 165, 701–721. [Google Scholar] [CrossRef]
- Lonnerberg, P.; Schoenherr, C.J.; Anderson, D.J.; Ibanez, C.F. Cell type-specific regulation of choline acetyltransferase gene expression. Role of the neuron-restrictive silencer element and cholinergic-specific enhancer sequences. J. Biol. Chem. 1996, 271, 33358–33365. [Google Scholar] [CrossRef]
- Nawa, Y.; Kaneko, H.; Oda, M.; Tsubonoya, M.; Hiroi, T.; Gentile, M.T.; Colucci-D’Amato, L.; Takahashi, R.; Matsui, H. Functional characterization of the neuron-restrictive silencer element in the human tryptophan hydroxylase 2 gene expression. J. Neurochem. 2017, 142, 827–840. [Google Scholar] [CrossRef]
- Wood, I.C.; Roopra, A.; Buckley, N.J. Neural specific expression of the m4 muscarinic acetylcholine receptor gene is mediated by a RE1/NRSE-type silencing element. J. Biol. Chem. 1996, 271, 14221–14225. [Google Scholar] [CrossRef]
- Mieda, M.; Haga, T.; Saffen, D.W. Expression of the rat m4 muscarinic acetylcholine receptor gene is regulated by the neuron-restrictive silencer element/repressor element 1. J. Biol. Chem. 1997, 272, 5854–5860. [Google Scholar] [CrossRef]
- Mu, W.; Burt, D.R. Transcriptional regulation of GABAA receptor gamma2 subunit gene. Brain Res. Mol. Brain Res. 1999, 67, 137–147. [Google Scholar] [CrossRef]
- Okamoto, S.; Sherman, K.; Lipton, S.A. Absence of binding activity of neuron-restrictive silencer factor is necessary, but not sufficient for transcription of NMDA receptor subunit type 1 in neuronal cells. Brain Res. Mol. Brain Res. 1999, 74, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Lemonde, S.; Rogaeva, A.; Albert, P.R. Cell type-dependent recruitment of trichostatin A-sensitive repression of the human 5-HT1A receptor gene. J. Neurochem. 2004, 88, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Brene, S.; Messer, C.; Okado, H.; Hartley, M.; Heinemann, S.F.; Nestler, E.J. Regulation of GluR2 promoter activity by neurotrophic factors via a neuron-restrictive silencer element. Eur. J. Neurosci. 2000, 12, 1525–1533. [Google Scholar] [CrossRef]
- Lee, K.S.; Chatterjee, P.; Choi, E.Y.; Sung, M.K.; Oh, J.; Won, H.; Park, S.M.; Kim, Y.J.; Yi, S.V.; Choi, J.K. Selection on the regulation of sympathetic nervous activity in humans and chimpanzees. PLoS Genet. 2018, 14, e1007311. [Google Scholar] [CrossRef]
- Kim, C.S.; Hwang, C.K.; Song, K.Y.; Choi, H.S.; Kim, D.K.; Law, P.Y.; Wei, L.N.; Loh, H.H. Novel function of neuron-restrictive silencer factor (NRSF) for posttranscriptional regulation. Biochim. Biophys. Acta 2008, 1783, 1835–1846. [Google Scholar] [CrossRef]
- Kim, C.S.; Hwang, C.K.; Choi, H.S.; Song, K.Y.; Law, P.Y.; Wei, L.N.; Loh, H.H. Neuron-restrictive silencer factor (NRSF) functions as a repressor in neuronal cells to regulate the mu opioid receptor gene. J. Biol. Chem. 2004, 279, 46464–46473. [Google Scholar] [CrossRef]
- Kim, C.S.; Choi, H.S.; Hwang, C.K.; Song, K.Y.; Lee, B.K.; Law, P.Y.; Wei, L.N.; Loh, H.H. Evidence of the neuron-restrictive silencer factor (NRSF) interaction with Sp3 and its synergic repression to the mu opioid receptor (MOR) gene. Nucleic Acids Res. 2006, 34, 6392–6403. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Shi, L.; Zhou, Y.; He, J.; Zhang, W.; Gu, X.; Zhang, J.; Ma, Z. Changes of Mu-opioid receptor and neuron-restrictive silencer factor in periaquductal gray in mouse models of remifentanil-induced postoperative hyperalgesia. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2014, 39, 901–906. [Google Scholar] [CrossRef]
- Shudo, Y.; Shimojo, M.; Fukunaga, M.; Ito, S. Pituitary adenylate cyclase-activating polypeptide is regulated by alternative splicing of transcriptional repressor REST/NRSF in nerve injury. Life Sci. 2015, 143, 174–181. [Google Scholar] [CrossRef]
- Wang, D.; Yu, J. Negative regulation of REST on NR2B in spinal cord contributes to the development of bone cancer pain in mice. Oncotarget 2016, 7, 85564–85572. [Google Scholar] [CrossRef]
- Zhu, C.; Tang, J.; Ding, T.; Chen, L.; Wang, W.; Mei, X.P.; He, X.T.; Wang, W.; Zhang, L.D.; Dong, Y.L.; et al. Neuron-restrictive silencer factor-mediated downregulation of mu-opioid receptor contributes to the reduced morphine analgesia in bone cancer pain. Pain 2017, 158, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Xiao-Die, X.; Xiao-Hong, W.; Cheng-Feng, H.; Zhong-Yu, Y.; Jian-Tao, W.; Hou-Guang, Z.; Jing-Chun, G. Increased NRSF/REST in anterior cingulate cortex contributes to diabetes-related neuropathic pain. Biochem. Biophys. Res. Commun. 2020, 527, 785–790. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, S.R.; Chen, H.; Pan, H.L. RE1-silencing transcription factor controls the acute-to-chronic neuropathic pain transition and Chrm2 receptor gene expression in primary sensory neurons. J. Biol. Chem. 2018, 293, 19078–19091. [Google Scholar] [CrossRef]
- Zhang, F.; Gigout, S.; Liu, Y.; Wang, Y.; Hao, H.; Buckley, N.J.; Zhang, H.; Wood, I.C.; Gamper, N. Repressor element 1-silencing transcription factor drives the development of chronic pain states. Pain 2019, 160, 2398–2408. [Google Scholar] [CrossRef] [PubMed]
- Uchida, H.; Sasaki, K.; Ma, L.; Ueda, H. Neuron-restrictive silencer factor causes epigenetic silencing of Kv4.3 gene after peripheral nerve injury. Neuroscience 2010, 166, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Uchida, H.; Ma, L.; Ueda, H. Epigenetic gene silencing underlies C-fiber dysfunctions in neuropathic pain. J. Neurosci. 2010, 30, 4806–4814. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Q.; Yang, Y.; Lv, Y.; Chen, L.; Bai, C.; Nan, X.; Wang, Y.; Pei, X. Regulatory role of neuron-restrictive silencing factor in the specific expression of cocaine- and amphetamine-regulated transcript gene. J. Neurochem. 2008, 106, 1314–1324. [Google Scholar] [CrossRef]
- D’Alessandro, R.; Klajn, A.; Stucchi, L.; Podini, P.; Malosio, M.L.; Meldolesi, J. Expression of the neurosecretory process in PC12 cells is governed by REST. J. Neurochem. 2008, 105, 1369–1383. [Google Scholar] [CrossRef]
- Prada, I.; Marchaland, J.; Podini, P.; Magrassi, L.; D’Alessandro, R.; Bezzi, P.; Meldolesi, J. REST/NRSF governs the expression of dense-core vesicle gliosecretion in astrocytes. J. Cell Biol. 2011, 193, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Valiya Veettil, M.; Krishna, G.; Roy, A.; Ghosh, A.; Dutta, D.; Kumar, B.; Chakraborty, S.; Anju, T.R.; Sharma-Walia, N.; Chandran, B. Kaposi’s Sarcoma-Associated Herpesvirus Infection Induces the Expression of Neuroendocrine Genes in Endothelial Cells. J. Virol. 2020, 94, e01692-19. [Google Scholar] [CrossRef] [PubMed]
- Mbikay, M.; Raffin-Sanson, M.L.; Sirois, F.; Kalenga, L.; Chretien, M.; Seidah, N.G. Characterization of a repressor element in the promoter region of proprotein convertase 2 (PC2) gene. Brain Res. Mol. Brain Res. 2002, 102, 35–47. [Google Scholar] [CrossRef]
- Seth, K.A.; Majzoub, J.A. Repressor element silencing transcription factor/neuron-restrictive silencing factor (REST/NRSF) can act as an enhancer as well as a repressor of corticotropin-releasing hormone gene transcription. J. Biol. Chem. 2001, 276, 13917–13923. [Google Scholar] [CrossRef] [PubMed]
- Rockowitz, S.; Zheng, D. Significant expansion of the REST/NRSF cistrome in human versus mouse embryonic stem cells: Potential implications for neural development. Nucleic Acids Res. 2015, 43, 5730–5743. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, D.; Rockowitz, S.; Zheng, D. Divergence and rewiring of regulatory networks for neural development between human and other species. Neurogenesis 2016, 3, e1231495. [Google Scholar] [CrossRef]
- Bolton, J.L.; Schulmann, A.; Garcia-Curran, M.M.; Regev, L.; Chen, Y.; Kamei, N.; Shao, M.; Singh-Taylor, A.; Jiang, S.; Noam, Y.; et al. Unexpected Transcriptional Programs Contribute to Hippocampal Memory Deficits and Neuronal Stunting after Early-Life Adversity. Cell Rep. 2020, 33, 108511. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, X.; Li, D.; Fang, R.; Wang, Z.; Yun, D.; Wang, M.; Wang, J.; Dong, H.; Fei, Z.; et al. NRSF regulates age-dependently cognitive ability and its conditional knockout in APP/PS1 mice moderately alters ad-like pathology. Hum. Mol. Genet. 2022, 32, 2558–2575. [Google Scholar] [CrossRef]
- Yang, M.; Li, Y.; Hu, L.; Luo, D.; Zhang, Y.; Xiao, X.; Li, G.; Zhang, L.; Zhu, G. Lead exposure inhibits expression of SV2C through NRSF. Toxicology 2018, 398–399, 23–30. [Google Scholar] [CrossRef]
- Verma, P.; Gupta, R.K.; Gandhi, B.S.; Singh, P. CDRI-08 Attenuates REST/NRSF-Mediated Expression of NMDAR1 Gene in PBDE-209-Exposed Mice Brain. Evid. Based Complement. Alternat Med. 2015, 2015, 403840. [Google Scholar] [CrossRef]
- Gupta, P.; Gupta, R.K.; Gandhi, B.S.; Singh, P. Differential binding of CREB and REST/NRSF to NMDAR1 promoter is associated with the sex-selective cognitive deficit following postnatal PBDE-209 exposure in mice. Environ. Sci. Pollut. Res. Int. 2023. ahead-of-print. [Google Scholar] [CrossRef]
- Gong, P.; Zhang, F.; Lei, X.; Wu, X.; Chen, D.; Zhang, W.; Zhang, K.; Zheng, A.; Gao, X. No observable relationship between the 12 genes of nervous system and reasoning skill in a young Chinese Han population. Cell Mol. Neurobiol. 2011, 31, 519–526. [Google Scholar] [CrossRef]
- Mori, N.; Mizuno, T.; Murai, K.; Nakano, I.; Yamashita, H. Effect of age on the gene expression of neural-restrictive silencing factor NRSF/REST. Neurobiol. Aging 2002, 23, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, A.; Carminati, E.; De Fusco, A.; Kowalska, J.A.; Floss, T.; Benfenati, F. REST/NRSF deficiency impairs autophagy and leads to cellular senescence in neurons. Aging Cell 2021, 20, e13471. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Sun, R.; Yao, M.; Chen, W.; Wang, Z.; Fei, J. A C-terminal truncated mutation of spr-3 gene extends lifespan in Caenorhabditis elegans. Acta Biochim. Biophys. Sin. 2013, 45, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.J.; He, C.F.; Zhou, R.Y.; Xu, X.D.; Xiang, L.X.; Wang, J.T.; Wang, X.R.; Zhou, H.G.; Guo, J.C. High glucose and palmitic acid induces neuronal senescence by NRSF/REST elevation and the subsequent mTOR-related autophagy suppression. Mol. Brain 2022, 15, 61. [Google Scholar] [CrossRef] [PubMed]
- Obata, H.; Naito, K.; Kikui, A.; Nakamura, S.; Suzuki, K.; Kawakita, S.; Suzuki, T.; Goto, K.; Nagura, N.; Sugiyama, Y.; et al. Age-related differences for expression of the nerve-specific proteins after peripheral nerve injury. Exp. Ther. Med. 2022, 24, 682. [Google Scholar] [CrossRef]
- Gonzalez-Castaneda, R.E.; Sanchez-Gonzalez, V.J.; Flores-Soto, M.; Vazquez-Camacho, G.; Macias-Islas, M.A.; Ortiz, G.G. Neural restrictive silencer factor and choline acetyltransferase expression in cerebral tissue of Alzheimer’s Disease patients: A pilot study. Genet. Mol. Biol. 2013, 36, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Orta-Salazar, E.; Aguilar-Vazquez, A.; Martinez-Coria, H.; Luquin-De Anda, S.; Rivera-Cervantes, M.; Beas-Zarate, C.; Feria-Velasco, A.; Diaz-Cintra, S. REST/NRSF-induced changes of ChAT protein expression in the neocortex and hippocampus of the 3xTg-AD mouse model for Alzheimer’s disease. Life Sci. 2014, 116, 83–89. [Google Scholar] [CrossRef]
- Lu, T.; Aron, L.; Zullo, J.; Pan, Y.; Kim, H.; Chen, Y.; Yang, T.H.; Kim, H.M.; Drake, D.; Liu, X.S.; et al. REST and stress resistance in ageing and Alzheimer’s disease. Nature 2014, 507, 448–454. [Google Scholar] [CrossRef]
- Kawamura, M.; Sato, S.; Matsumoto, G.; Fukuda, T.; Shiba-Fukushima, K.; Noda, S.; Takanashi, M.; Mori, N.; Hattori, N. Loss of nuclear REST/NRSF in aged-dopaminergic neurons in Parkinson’s disease patients. Neurosci. Lett. 2019, 699, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Cai, L.; Liang, M.; Huang, Y.; Gao, H.; Lu, S.; Fei, J.; Huang, F. Alteration of NRSF expression exacerbating 1-methyl-4-phenyl-pyridinium ion-induced cell death of SH-SY5Y cells. Neurosci. Res. 2009, 65, 236–244. [Google Scholar] [CrossRef]
- Yu, M.; Suo, H.; Liu, M.; Cai, L.; Liu, J.; Huang, Y.; Xu, J.; Wang, Y.; Zhu, C.; Fei, J.; et al. NRSF/REST neuronal deficient mice are more vulnerable to the neurotoxin MPTP. Neurobiol. Aging 2013, 34, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Suo, H.; Wang, P.; Tong, J.; Cai, L.; Liu, J.; Huang, D.; Huang, L.; Wang, Z.; Huang, Y.; Xu, J.; et al. NRSF is an essential mediator for the neuroprotection of trichostatin A in the MPTP mouse model of Parkinson’s disease. Neuropharmacology 2015, 99, 67–78. [Google Scholar] [CrossRef]
- Huang, D.; Li, Q.; Wang, Y.; Liu, Z.; Wang, Z.; Li, H.; Wang, J.; Su, J.; Ma, Y.; Yu, M.; et al. Brain-specific NRSF deficiency aggravates dopaminergic neurodegeneration and impairs neurogenesis in the MPTP mouse model of Parkinson’s disease. Aging 2019, 11, 3280–3297. [Google Scholar] [CrossRef]
- Li, H.; Liu, Z.; Wu, Y.; Chen, Y.; Wang, J.; Wang, Z.; Huang, D.; Wang, M.; Yu, M.; Fei, J.; et al. The deficiency of NRSF/REST enhances the pro-inflammatory function of astrocytes in a model of Parkinson’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165590. [Google Scholar] [CrossRef]
- Zuccato, C.; Tartari, M.; Crotti, A.; Goffredo, D.; Valenza, M.; Conti, L.; Cataudella, T.; Leavitt, B.R.; Hayden, M.R.; Timmusk, T.; et al. Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes. Nat. Genet. 2003, 35, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Marullo, M.; Valenza, M.; Mariotti, C.; Di Donato, S.; Cattaneo, E.; Zuccato, C. Analysis of the repressor element-1 silencing transcription factor/neuron-restrictive silencer factor occupancy of non-neuronal genes in peripheral lymphocytes from patients with Huntington’s disease. Brain Pathol. 2010, 20, 96–105. [Google Scholar] [CrossRef]
- Schiffer, D.; Caldera, V.; Mellai, M.; Conforti, P.; Cattaneo, E.; Zuccato, C. Repressor element-1 silencing transcription factor (REST) is present in human control and Huntington’s disease neurones. Neuropathol. Appl. Neurobiol. 2014, 40, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.; Belyaev, N.; Conforti, P.; Ooi, L.; Tartari, M.; Papadimou, E.; MacDonald, M.; Fossale, E.; Zeitlin, S.; Buckley, N.; et al. Widespread disruption of repressor element-1 silencing transcription factor/neuron-restrictive silencer factor occupancy at its target genes in Huntington’s disease. J. Neurosci. 2007, 27, 6972–6983. [Google Scholar] [CrossRef]
- Shimojo, M. Huntingtin regulates RE1-silencing transcription factor/neuron-restrictive silencer factor (REST/NRSF) nuclear trafficking indirectly through a complex with REST/NRSF-interacting LIM domain protein (RILP) and dynactin p150 Glued. J. Biol. Chem. 2008, 283, 34880–34886. [Google Scholar] [CrossRef] [PubMed]
- Datta, M.; Bhattacharyya, N.P. Regulation of RE1 protein silencing transcription factor (REST) expression by HIP1 protein interactor (HIPPI). J. Biol. Chem. 2011, 286, 33759–33769. [Google Scholar] [CrossRef]
- Chen, G.L.; Ma, Q.; Goswami, D.; Shang, J.; Miller, G.M. Modulation of nuclear REST by alternative splicing: A potential therapeutic target for Huntington’s disease. J. Cell. Mol. Med. 2017, 21, 2974–2984. [Google Scholar] [CrossRef] [PubMed]
- de Souza, J.M.; Abd-Elrahman, K.S.; Ribeiro, F.M.; Ferguson, S.S.G. mGluR5 regulates REST/NRSF signaling through N-cadherin/beta-catenin complex in Huntington’s disease. Mol. Brain 2020, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Navarrete-Modesto, V.; Orozco-Suarez, S.; Alonso-Vanegas, M.; Feria-Romero, I.A.; Rocha, L. REST/NRSF transcription factor is overexpressed in hippocampus of patients with drug-resistant mesial temporal lobe epilepsy. Epilepsy Behav. 2019, 94, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.L.; Cheng, X.; Cai, L.; Tan, G.H.; Xu, L.; Feng, X.Y.; Lu, T.J.; Xiong, H.; Fei, J.; Xiong, Z.Q. Conditional deletion of NRSF in forebrain neurons accelerates epileptogenesis in the kindling model. Cereb. Cortex 2011, 21, 2158–2165. [Google Scholar] [CrossRef]
- Liu, M.; Sheng, Z.; Cai, L.; Zhao, K.; Tian, Y.; Fei, J. Neuronal conditional knockout of NRSF decreases vulnerability to seizures induced by pentylenetetrazol in mice. Acta Biochim. Biophys. Sin. 2012, 44, 476–482. [Google Scholar] [CrossRef]
- Patterson, K.P.; Barry, J.M.; Curran, M.M.; Singh-Taylor, A.; Brennan, G.; Rismanchi, N.; Page, M.; Noam, Y.; Holmes, G.L.; Baram, T.Z. Enduring Memory Impairments Provoked by Developmental Febrile Seizures Are Mediated by Functional and Structural Effects of Neuronal Restrictive Silencing Factor. J. Neurosci. 2017, 37, 3799–3812. [Google Scholar] [CrossRef]
- McClelland, S.; Flynn, C.; Dubé, C.; Richichi, C.; Zha, Q.; Ghestem, A.; Esclapez, M.; Bernard, C.; Baram, T.Z. Neuron-restrictive silencer factor-mediated hyperpolarization-activated cyclic nucleotide gated channelopathy in experimental temporal lobe epilepsy. Ann. Neurol. 2011, 70, 454–464. [Google Scholar] [CrossRef]
- Carminati, E.; Buffolo, F.; Rocchi, A.; Michetti, C.; Cesca, F.; Benfenati, F. Mild Inactivation of RE-1 Silencing Transcription Factor (REST) Reduces Susceptibility to Kainic Acid-Induced Seizures. Front. Cell. Neurosci. 2019, 13, 580. [Google Scholar] [CrossRef]
- Hu, X.L.; Cheng, X.; Fei, J.; Xiong, Z.Q. Neuron-restrictive silencer factor is not required for the antiepileptic effect of the ketogenic diet. Epilepsia 2011, 52, 1609–1616. [Google Scholar] [CrossRef]
- Bassuk, A.G.; Wallace, R.H.; Buhr, A.; Buller, A.R.; Afawi, Z.; Shimojo, M.; Miyata, S.; Chen, S.; Gonzalez-Alegre, P.; Griesbach, H.L.; et al. A homozygous mutation in human PRICKLE1 causes an autosomal-recessive progressive myoclonus epilepsy-ataxia syndrome. Am. J. Hum. Genet. 2008, 83, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.P.; Bubb, V.J.; Marshall-Jones, Z.V.; Coulson, J.M. Neuron restrictive silencer factor as a modulator of neuropeptide gene expression. Regul. Pept. 2002, 108, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Spencer, E.M.; Chandler, K.E.; Haddley, K.; Howard, M.R.; Hughes, D.; Belyaev, N.D.; Coulson, J.M.; Stewart, J.P.; Buckley, N.J.; Kipar, A.; et al. Regulation and role of REST and REST4 variants in modulation of gene expression in in vivo and in vitro in epilepsy models. Neurobiol. Dis. 2006, 24, 41–52. [Google Scholar] [CrossRef]
- Gillies, S.; Haddley, K.; Vasiliou, S.; Bubb, V.J.; Quinn, J.P. The human neurokinin B gene, TAC3, and its promoter are regulated by Neuron Restrictive Silencing Factor (NRSF) transcription factor family. Neuropeptides 2009, 43, 333–340. [Google Scholar] [CrossRef]
- Hall, A.M.; Brennan, G.P.; Nguyen, T.M.; Singh-Taylor, A.; Mun, H.S.; Sargious, M.J.; Baram, T.Z. The Role of Sirt1 in Epileptogenesis. Eneuro 2017, preprint. [Google Scholar] [CrossRef] [PubMed]
- Brennan, G.P.; Dey, D.; Chen, Y.; Patterson, K.P.; Magnetta, E.J.; Hall, A.M.; Dube, C.M.; Mei, Y.T.; Baram, T.Z. Dual and Opposing Roles of MicroRNA-124 in Epilepsy Are Mediated through Inflammatory and NRSF-Dependent Gene Networks. Cell Rep. 2016, 14, 2402–2412. [Google Scholar] [CrossRef] [PubMed]
- Calderone, A.; Jover, T.; Noh, K.M.; Tanaka, H.; Yokota, H.; Lin, Y.; Grooms, S.Y.; Regis, R.; Bennett, M.V.; Zukin, R.S. Ischemic insults derepress the gene silencer REST in neurons destined to die. J. Neurosci. 2003, 23, 2112–2121. [Google Scholar] [CrossRef]
- Formisano, L.; Noh, K.M.; Miyawaki, T.; Mashiko, T.; Bennett, M.V.; Zukin, R.S. Ischemic insults promote epigenetic reprogramming of mu opioid receptor expression in hippocampal neurons. Proc. Natl. Acad. Sci. USA 2007, 104, 4170–4175. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.M.; Geng, L.J.; Shi, X.Y.; Zhang, C.G.; Wang, S.Y.; Zhang, G.M. By up-regulating mu- and delta-opioid receptors, neuron-restrictive silencer factor knockdown promotes neurological recovery after ischemia. Oncotarget 2017, 8, 101012–101025. [Google Scholar] [CrossRef] [PubMed]
- He, C.F.; Xue, W.J.; Xu, X.D.; Wang, J.T.; Wang, X.R.; Feng, Y.; Zhou, H.G.; Guo, J.C. Knockdown of NRSF Alleviates Ischemic Brain Injury and Microvasculature Defects in Diabetic MCAO Mice. Front. Neurol. 2022, 13, 869220. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, S.; Yuan, L.; Yang, Y.; Zhang, B.; Liu, Q.; Chen, L.; Yue, W.; Li, Y.; Pei, X. Neuron-restrictive silencer factor (NRSF) represses cocaine- and amphetamine-regulated transcript (CART) transcription and antagonizes cAMP-response element-binding protein signaling through a dual NRSE mechanism. J. Biol. Chem. 2012, 287, 42574–42587. [Google Scholar] [CrossRef]
- Kaneko, N.; Hwang, J.Y.; Gertner, M.; Pontarelli, F.; Zukin, R.S. Casein kinase 1 suppresses activation of REST in insulted hippocampal neurons and halts ischemia-induced neuronal death. J. Neurosci. 2014, 34, 6030–6039. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Kaneko, N.; Noh, K.M.; Pontarelli, F.; Zukin, R.S. The gene silencing transcription factor REST represses miR-132 expression in hippocampal neurons destined to die. J. Mol. Biol. 2014, 426, 3454–3466. [Google Scholar] [CrossRef]
- Yang, B.; Zang, L.E.; Cui, J.W.; Zhang, M.Y.; Ma, X.; Wei, L.L. Melatonin Plays a Protective Role by Regulating miR-26a-5p-NRSF and JAK2-STAT3 Pathway to Improve Autophagy, Inflammation and Oxidative Stress of Cerebral Ischemia-Reperfusion Injury. Drug Des. Dev. Ther. 2020, 14, 3177–3188. [Google Scholar] [CrossRef] [PubMed]
- Loe-Mie, Y.; Lepagnol-Bestel, A.M.; Maussion, G.; Doron-Faigenboim, A.; Imbeaud, S.; Delacroix, H.; Aggerbeck, L.; Pupko, T.; Gorwood, P.; Simonneau, M.; et al. SMARCA2 and other genome-wide supported schizophrenia-associated genes: Regulation by REST/NRSF, network organization and primate-specific evolution. Hum. Mol. Genet. 2010, 19, 2841–2857. [Google Scholar] [CrossRef] [PubMed]
- Gulchina, Y.; Xu, S.J.; Snyder, M.A.; Elefant, F.; Gao, W.J. Epigenetic mechanisms underlying NMDA receptor hypofunction in the prefrontal cortex of juvenile animals in the MAM model for schizophrenia. J. Neurochem. 2017, 143, 320–333. [Google Scholar] [CrossRef]
- Patel, P.D.; Bochar, D.A.; Turner, D.L.; Meng, F.; Mueller, H.M.; Pontrello, C.G. Regulation of tryptophan hydroxylase-2 gene expression by a bipartite RE-1 silencer of transcription/neuron restrictive silencing factor (REST/NRSF) binding motif. J. Biol. Chem. 2007, 282, 26717–26724. [Google Scholar] [CrossRef]
- Ishii, T.; Hashimoto, E.; Ukai, W.; Tateno, M.; Yoshinaga, T.; Saito, S.; Sohma, H.; Saito, T. Lithium-induced suppression of transcription repressor NRSF/REST: Effects on the dysfunction of neuronal differentiation by ethanol. Eur. J. Pharmacol. 2008, 593, 36–43. [Google Scholar] [CrossRef]
- Warburton, A.; Savage, A.L.; Myers, P.; Peeney, D.; Bubb, V.J.; Quinn, J.P. Molecular signatures of mood stabilisers highlight the role of the transcription factor REST/NRSF. J. Affect. Disord. 2015, 172, 63–73. [Google Scholar] [CrossRef]
- Henriksson, R.; Backman, C.M.; Harvey, B.K.; Kadyrova, H.; Bazov, I.; Shippenberg, T.S.; Bakalkin, G. PDYN, a gene implicated in brain/mental disorders, is targeted by REST in the adult human brain. Biochim. Biophys. Acta 2014, 1839, 1226–1232. [Google Scholar] [CrossRef]
- Korosi, A.; Shanabrough, M.; McClelland, S.; Liu, Z.W.; Borok, E.; Gao, X.B.; Horvath, T.L.; Baram, T.Z. Early-life experience reduces excitation to stress-responsive hypothalamic neurons and reprograms the expression of corticotropin-releasing hormone. J. Neurosci. 2010, 30, 703–713. [Google Scholar] [CrossRef]
- Hai-Ying, C.; Nagano, K.; Ezzikouri, S.; Yamaguchi, C.; Kayesh, M.E.; Rebbani, K.; Kitab, B.; Nakano, H.; Kouji, H.; Kohara, M.; et al. Establishment of an intermittent cold stress model using Tupaia belangeri and evaluation of compound C737 targeting neuron-restrictive silencer factor. Exp. Anim. 2016, 65, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Singh-Taylor, A.; Molet, J.; Jiang, S.; Korosi, A.; Bolton, J.L.; Noam, Y.; Simeone, K.; Cope, J.; Chen, Y.; Mortazavi, A.; et al. NRSF-dependent epigenetic mechanisms contribute to programming of stress-sensitive neurons by neonatal experience, promoting resilience. Mol. Psychiatry 2018, 23, 648–657. [Google Scholar] [CrossRef]
- Wen, X.H.; Guo, Q.L.; Guo, J.C. Effect of 9—PAHSA on cognitive dysfunction in diabetic mice and its possible mechanism. Biochem. Biophys. Res. Commun. 2020, 524, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, K.; Saito, Y.; Takano, M.; Arai, Y.; Yasuno, S.; Nakagawa, Y.; Takahashi, N.; Adachi, Y.; Takemura, G.; Horie, M.; et al. NRSF regulates the fetal cardiac gene program and maintains normal cardiac structure and function. EMBO J. 2003, 22, 6310–6321. [Google Scholar] [CrossRef]
- Kuratomi, S.; Kuratomi, A.; Kuwahara, K.; Ishii, T.M.; Nakao, K.; Saito, Y.; Takano, M. NRSF regulates the developmental and hypertrophic changes of HCN4 transcription in rat cardiac myocytes. Biochem. Biophys. Res. Commun. 2007, 353, 67–73. [Google Scholar] [CrossRef]
- Schweizer, P.A.; Yampolsky, P.; Malik, R.; Thomas, D.; Zehelein, J.; Katus, H.A.; Koenen, M. Transcription profiling of HCN-channel isotypes throughout mouse cardiac development. Basic. Res. Cardiol. 2009, 104, 621–629. [Google Scholar] [CrossRef]
- Zhang, D.; Wu, B.; Wang, P.; Wang, Y.; Lu, P.; Nechiporuk, T.; Floss, T.; Greally, J.M.; Zheng, D.; Zhou, B. Non-CpG methylation by DNMT3B facilitates REST binding and gene silencing in developing mouse hearts. Nucleic Acids Res. 2017, 45, 3102–3115. [Google Scholar] [CrossRef]
- Liu, J.; Liu, S.; Gao, H.; Han, L.; Chu, X.; Sheng, Y.; Shou, W.; Wang, Y.; Liu, Y.; Wan, J.; et al. Genome-wide studies reveal the essential and opposite roles of ARID1A in controlling human cardiogenesis and neurogenesis from pluripotent stem cells. Genome Biol. 2020, 21, 169. [Google Scholar] [CrossRef]
- D’Souza, A.; Bucchi, A.; Johnsen, A.B.; Logantha, S.J.; Monfredi, O.; Yanni, J.; Prehar, S.; Hart, G.; Cartwright, E.; Wisloff, U.; et al. Exercise training reduces resting heart rate via downregulation of the funny channel HCN4. Nat. Commun. 2014, 5, 3775. [Google Scholar] [CrossRef]
- Koyama, R.; Mannic, T.; Ito, J.; Amar, L.; Zennaro, M.C.; Rossier, M.F.; Maturana, A.D. MicroRNA-204 Is Necessary for Aldosterone-Stimulated T-Type Calcium Channel Expression in Cardiomyocytes. Int. J. Mol. Sci. 2018, 19, 2941. [Google Scholar] [CrossRef] [PubMed]
- Rossier, M.F. The Cardiac Mineralocorticoid Receptor (MR): A Therapeutic Target Against Ventricular Arrhythmias. Front. Endocrinol. 2021, 12, 694758. [Google Scholar] [CrossRef] [PubMed]
- Somekawa, S.; Imagawa, K.; Naya, N.; Takemoto, Y.; Onoue, K.; Okayama, S.; Takeda, Y.; Kawata, H.; Horii, M.; Nakajima, T.; et al. Regulation of aldosterone and cortisol production by the transcriptional repressor neuron restrictive silencer factor. Endocrinology 2009, 150, 3110–3117. [Google Scholar] [CrossRef] [PubMed]
- Inazumi, H.; Kuwahara, K.; Nakagawa, Y.; Kuwabara, Y.; Numaga-Tomita, T.; Kashihara, T.; Nakada, T.; Kurebayashi, N.; Oya, M.; Nonaka, M.; et al. NRSF-GNAO1 Pathway Contributes to the Regulation of Cardiac Ca2+ Homeostasis. Circ. Res. 2022, 130, 234–248. [Google Scholar] [CrossRef]
- Lugenbiel, P.; Govorov, K.; Syren, P.; Rahm, A.K.; Wieder, T.; Wunsch, M.; Weiberg, N.; Manolova, E.; Gramlich, D.; Rivinius, R.; et al. Epigenetic regulation of cardiac electrophysiology in atrial fibrillation: HDAC2 determines action potential duration and suppresses NRSF in cardiomyocytes. Basic Res. Cardiol. 2021, 116, 13. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, E.; Saito, Y.; Kuwahara, K.; Harada, M.; Miyamoto, Y.; Hamanaka, I.; Kajiyama, N.; Takahashi, N.; Izumi, T.; Kawakami, R.; et al. Fibronectin signaling stimulates BNP gene transcription by inhibiting neuron-restrictive silencer element-dependent repression. Cardiovasc. Res. 2002, 53, 451–459. [Google Scholar] [CrossRef]
- Kuwahara, K.; Saito, Y.; Ogawa, E.; Takahashi, N.; Nakagawa, Y.; Naruse, Y.; Harada, M.; Hamanaka, I.; Izumi, T.; Miyamoto, Y.; et al. The neuron-restrictive silencer element-neuron-restrictive silencer factor system regulates basal and endothelin 1-inducible atrial natriuretic peptide gene expression in ventricular myocytes. Mol. Cell Biol. 2001, 21, 2085–2097. [Google Scholar] [CrossRef]
- Ohba, T.; Watanabe, H.; Takahashi, Y.; Suzuki, T.; Miyoshi, I.; Nakayama, S.; Satoh, E.; Iino, K.; Sasano, H.; Mori, Y.; et al. Regulatory role of neuron-restrictive silencing factor in expression of TRPC1. Biochem. Biophys. Res. Commun. 2006, 351, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Hata, L.; Murakami, M.; Kuwahara, K.; Nakagawa, Y.; Kinoshita, H.; Usami, S.; Yasuno, S.; Fujiwara, M.; Kuwabara, Y.; Minami, T.; et al. Zinc-finger protein 90 negatively regulates neuron-restrictive silencer factor-mediated transcriptional repression of fetal cardiac genes. J. Mol. Cell. Cardiol. 2011, 50, 972–981. [Google Scholar] [CrossRef]
- Atouf, F.; Czernichow, P.; Scharfmann, R. Expression of neuronal traits in pancreatic beta cells. Implication of neuron-restrictive silencing factor/repressor element silencing transcription factor, a neuron-restrictive silencer. J. Biol. Chem. 1997, 272, 1929–1934. [Google Scholar] [CrossRef]
- Martin, D.; Kim, Y.H.; Sever, D.; Mao, C.A.; Haefliger, J.A.; Grapin-Botton, A. REST represses a subset of the pancreatic endocrine differentiation program. Dev. Biol. 2015, 405, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Kemp, D.M.; Lin, J.C.; Habener, J.F. Regulation of Pax4 paired homeodomain gene by neuron-restrictive silencer factor. J. Biol. Chem. 2003, 278, 35057–35062. [Google Scholar] [CrossRef]
- Martin, D.; Tawadros, T.; Meylan, L.; Abderrahmani, A.; Condorelli, D.F.; Waeber, G.; Haefliger, J.A. Critical role of the transcriptional repressor neuron-restrictive silencer factor in the specific control of connexin36 in insulin-producing cell lines. J. Biol. Chem. 2003, 278, 53082–53089. [Google Scholar] [CrossRef]
- Campa, D.; Capurso, G.; Pastore, M.; Talar-Wojnarowska, R.; Milanetto, A.C.; Landoni, L.; Maiello, E.; Lawlor, R.T.; Malecka-Panas, E.; Funel, N.; et al. Common germline variants within the CDKN2A/2B region affect risk of pancreatic neuroendocrine tumors. Sci. Rep. 2016, 6, 39565. [Google Scholar] [CrossRef]
- Li, B.; Wang, S.; Liu, H.; Liu, D.; Zhang, J.; Zhang, B.; Yao, H.; Lv, Y.; Wang, R.; Chen, L.; et al. Neuronal restrictive silencing factor silencing induces human amniotic fluid-derived stem cells differentiation into insulin-producing cells. Stem Cells Dev. 2011, 20, 1223–1231. [Google Scholar] [CrossRef]
- Li, H.T.; Jiang, F.X.; Shi, P.; Zhang, T.; Liu, X.Y.; Lin, X.W.; Pang, X.N. In vitro reprogramming of rat bone marrow-derived mesenchymal stem cells into insulin-producing cells by genetically manipulating negative and positive regulators. Biochem. Biophys. Res. Commun. 2012, 420, 793–798. [Google Scholar] [CrossRef]
- Kurschat, P.; Bielenberg, D.; Rossignol-Tallandier, M.; Stahl, A.; Klagsbrun, M. Neuron restrictive silencer factor NRSF/REST is a transcriptional repressor of neuropilin-1 and diminishes the ability of semaphorin 3A to inhibit keratinocyte migration. J. Biol. Chem. 2006, 281, 2721–2729. [Google Scholar] [CrossRef]
- Aoki, H.; Hara, A.; Kunisada, T. White spotting phenotype induced by targeted REST disruption during neural crest specification to a melanocyte cell lineage. Genes Cells 2015, 20, 439–449. [Google Scholar] [CrossRef]
- Lee, H.J.; Wall, B.A.; Wangari-Talbot, J.; Chen, S. Regulation of mGluR1 expression in human melanocytes and melanoma cells. Biochim. Biophys. Acta 2012, 1819, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Ogino, H.; Tomita, H.; Hara, A.; Kunisada, T. Disruption of Rest Leads to the Early Onset of Cataracts with the Aberrant Terminal Differentiation of Lens Fiber Cells. PLoS ONE 2016, 11, e0163042. [Google Scholar] [CrossRef]
- Tsuda, L.; Kaido, M.; Lim, Y.M.; Kato, K.; Aigaki, T.; Hayashi, S. An NRSF/REST-like repressor downstream of Ebi/SMRTER/Su(H) regulates eye development in Drosophila. EMBO J. 2006, 25, 3191–3202. [Google Scholar] [CrossRef]
- Cheong, A.; Bingham, A.J.; Li, J.; Kumar, B.; Sukumar, P.; Munsch, C.; Buckley, N.J.; Neylon, C.B.; Porter, K.E.; Beech, D.J.; et al. Downregulated REST transcription factor is a switch enabling critical potassium channel expression and cell proliferation. Mol. Cell 2005, 20, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Tharp, D.L.; Bowles, D.K. KCa3.1 Inhibition Decreases Size and Alters Composition of Atherosclerotic Lesions Induced by Low, Oscillatory Flow. Artery Res. 2021, 27, 93–100. [Google Scholar] [CrossRef]
- Nishimura, E.; Sasaki, K.; Maruyama, K.; Tsukada, T.; Yamaguchi, K. Decrease in neuron-restrictive silencer factor (NRSF) mRNA levels during differentiation of cultured neuroblastoma cells. Neurosci. Lett. 1996, 211, 101–104. [Google Scholar] [CrossRef]
- Lepagnol-Bestel, A.M.; Maussion, G.; Ramoz, N.; Moalic, J.M.; Gorwood, P.; Simonneau, M. Nrsf silencing induces molecular and subcellular changes linked to neuronal plasticity. NeuroReport 2007, 18, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Mitra, S.; Subhash, S.; Hertwig, F.; Kanduri, M.; Mishra, K.; Fransson, S.; Ganeshram, A.; Mondal, T.; Bandaru, S.; et al. The risk-associated long noncoding RNA NBAT-1 controls neuroblastoma progression by regulating cell proliferation and neuronal differentiation. Cancer Cell 2014, 26, 722–737. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Crisafulli, L.; Caldera, V.; Tortoreto, M.; Brilli, E.; Conforti, P.; Zunino, F.; Magrassi, L.; Schiffer, D.; Cattaneo, E. REST controls self-renewal and tumorigenic competence of human glioblastoma cells. PLoS ONE 2012, 7, e38486. [Google Scholar] [CrossRef]
- Ren, H.; Gao, Z.; Wu, N.; Zeng, L.; Tang, X.; Chen, X.; Liu, Z.; Zhang, W.; Wang, L.; Li, Z. Expression of REST4 in human gliomas in vivo and influence of pioglitazone on REST in vitro. Biochem. Biophys. Res. Commun. 2015, 463, 504–509. [Google Scholar] [CrossRef]
- Dobson, T.H.W.; Tao, R.H.; Swaminathan, J.; Maegawa, S.; Shaik, S.; Bravo-Alegria, J.; Sharma, A.; Kennis, B.; Yang, Y.; Callegari, K.; et al. Transcriptional repressor REST drives lineage stage-specific chromatin compaction at Ptch1 and increases AKT activation in a mouse model of medulloblastoma. Sci. Signal 2019, 12, eaan8680. [Google Scholar] [CrossRef]
- Su, X.; Gopalakrishnan, V.; Stearns, D.; Aldape, K.; Lang, F.F.; Fuller, G.; Snyder, E.; Eberhart, C.G.; Majumder, S. Abnormal expression of REST/NRSF and Myc in neural stem/progenitor cells causes cerebellar tumors by blocking neuronal differentiation. Mol. Cell. Biol. 2006, 26, 1666–1678. [Google Scholar] [CrossRef]
- Lawinger, P.; Venugopal, R.; Guo, Z.S.; Immaneni, A.; Sengupta, D.; Lu, W.; Rastelli, L.; Marin Dias Carneiro, A.; Levin, V.; Fuller, G.N.; et al. The neuronal repressor REST/NRSF is an essential regulator in medulloblastoma cells. Nat. Med. 2000, 6, 826–831. [Google Scholar] [CrossRef]
- Fuller, G.N.; Su, X.; Price, R.E.; Cohen, Z.R.; Lang, F.F.; Sawaya, R.; Majumder, S. Many human medulloblastoma tumors overexpress repressor element-1 silencing transcription (REST)/neuron-restrictive silencer factor, which can be functionally countered by REST-VP16. Mol. Cancer Ther. 2005, 4, 343–349. [Google Scholar] [CrossRef]
- Callegari, K.; Maegawa, S.; Bravo-Alegria, J.; Gopalakrishnan, V. Pharmacological inhibition of LSD1 activity blocks REST-dependent medulloblastoma cell migration. Cell Commun. Signal 2018, 16, 60. [Google Scholar] [CrossRef]
- Shaik, S.; Maegawa, S.; Haltom, A.R.; Wang, F.; Xiao, X.; Dobson, T.; Sharma, A.; Yang, Y.; Swaminathan, J.; Kundra, V.; et al. REST promotes ETS1-dependent vascular growth in medulloblastoma. Mol. Oncol. 2021, 15, 1486–1506. [Google Scholar] [CrossRef]
- Palm, K.; Metsis, M.; Timmusk, T. Neuron-specific splicing of zinc finger transcription factor REST/NRSF/XBR is frequent in neuroblastomas and conserved in human, mouse and rat. Brain Res. Mol. Brain Res. 1999, 72, 30–39. [Google Scholar] [CrossRef]
- Blom, T.; Tynninen, O.; Puputti, M.; Halonen, M.; Paetau, A.; Haapasalo, H.; Tanner, M.; Nupponen, N.N. Molecular genetic analysis of the REST/NRSF gene in nervous system tumors. Acta Neuropathol. 2006, 112, 483–490. [Google Scholar] [CrossRef]
- Lee, J.H.; Chai, Y.G.; Hersh, L.B. Expression patterns of mouse repressor element-1 silencing transcription factor 4 (REST4) and its possible function in neuroblastoma. J. Mol. Neurosci. 2000, 15, 205–214. [Google Scholar] [CrossRef]
- Coulson, J.M.; Edgson, J.L.; Woll, P.J.; Quinn, J.P. A splice variant of the neuron-restrictive silencer factor repressor is expressed in small cell lung cancer: A potential role in derepression of neuroendocrine genes and a useful clinical marker. Cancer Res. 2000, 60, 1840–1844. [Google Scholar] [PubMed]
- Gurrola-Diaz, C.; Lacroix, J.; Dihlmann, S.; Becker, C.M.; von Knebel Doeberitz, M. Reduced expression of the neuron restrictive silencer factor permits transcription of glycine receptor alpha1 subunit in small-cell lung cancer cells. Oncogene 2003, 22, 5636–5645. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.B.; Seitz, R.; Gorzella, A.; Heister, A.; Doeberitz, M.; Becker, C.M. Relaxation of glycine receptor and onconeural gene transcription control in NRSF deficient small cell lung cancer cell lines. Brain Res. Mol. Brain Res. 2004, 120, 173–181. [Google Scholar] [CrossRef]
- Chang, L.; Schwarzenbach, H.; Meyer-Staeckling, S.; Brandt, B.; Mayr, G.W.; Weitzel, J.M.; Windhorst, S. Expression Regulation of the Metastasis-Promoting Protein InsP3-Kinase-A in Tumor Cells. Mol. Cancer Res. 2011, 9, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Kreisler, A.; Strissel, P.L.; Strick, R.; Neumann, S.B.; Schumacher, U.; Becker, C.M. Regulation of the NRSF/REST gene by methylation and CREB affects the cellular phenotype of small-cell lung cancer. Oncogene 2010, 29, 5828–5838. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Oda, C.; Mishima, K.; Tsuji, I.; Obika, S.; Shimojo, M. An antisense amido-bridged nucleic acid gapmer oligonucleotide targeting SRRM4 alters REST splicing and exhibits anti-tumor effects in small cell lung cancer and prostate cancer cells. Cancer Cell Int. 2023, 23, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xu, K.; Leng, X.; Han, Y.; Fang, Q. miRNA-9 Inhibits Proliferation and Migration of Lung Squamous Cell Carcinoma Cells by Regulating NRSF/EGFR. Technol. Cancer Res. Treat. 2020, 19, 1533033820945807. [Google Scholar] [CrossRef]
- Lim, J.S.; Ibaseta, A.; Fischer, M.M.; Cancilla, B.; O’Young, G.; Cristea, S.; Luca, V.C.; Yang, D.; Jahchan, N.S.; Hamard, C.; et al. Intratumoural heterogeneity generated by Notch signalling promotes small-cell lung cancer. Nature 2017, 545, 360–364. [Google Scholar] [CrossRef]
- Moss, A.C.; Jacobson, G.M.; Walker, L.E.; Blake, N.W.; Marshall, E.; Coulson, J.M. SCG3 transcript in peripheral blood is a prognostic biomarker for REST-deficient small cell lung cancer. Clin. Cancer Res. 2009, 15, 274–283. [Google Scholar] [CrossRef]
- Watanabe, H.; Mizutani, T.; Haraguchi, T.; Yamamichi, N.; Minoguchi, S.; Yamamichi-Nishina, M.; Mori, N.; Kameda, T.; Sugiyama, T.; Iba, H. SWI/SNF complex is essential for NRSF-mediated suppression of neuronal genes in human nonsmall cell lung carcinoma cell lines. Oncogene 2006, 25, 470–479. [Google Scholar] [CrossRef]
- Nair, S.; Bora-Singhal, N.; Perumal, D.; Chellappan, S. Nicotine-mediated invasion and migration of non-small cell lung carcinoma cells by modulating STMN3 and GSPT1 genes in an ID1-dependent manner. Mol. Cancer 2014, 13, 173. [Google Scholar] [CrossRef] [PubMed]
- Westbrook, T.F.; Martin, E.S.; Schlabach, M.R.; Leng, Y.; Liang, A.C.; Feng, B.; Zhao, J.J.; Roberts, T.M.; Mandel, G.; Hannon, G.J.; et al. A genetic screen for candidate tumor suppressors identifies REST. Cell 2005, 121, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, X. HMGA2 rs968697 T > C polymorphism is associated with the risk of colorectal cancer. Nucleosides Nucleotides Nucleic Acids 2021, 40, 821–828. [Google Scholar] [CrossRef]
- Ma, M.; Zhou, Y.; Sun, R.; Shi, J.; Tan, Y.; Yang, H.; Zhang, M.; Shen, R.; Xu, L.; Wang, Z.; et al. STAT3 and AKT signaling pathways mediate oncogenic role of NRSF in hepatocellular carcinoma. Acta Biochim. Biophys. Sin. 2020, 52, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, S.; Zhang, H.; Zhang, X.; Guo, D.; Zhang, J. NRSF/REST levels are decreased in cholangiocellular carcinoma but not hepatocellular carcinoma compared with normal liver tissues: A tissue microarray study. Oncol. Lett. 2018, 15, 6592–6598. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Pan, G.; Zheng, G.; Wu, X.; Ren, H.; Liu, Y.; Wen, J. Expression and functions of the repressor element 1 (RE-1)-silencing transcription factor (REST) in breast cancer. J. Cell. Biochem. 2010, 110, 968–974. [Google Scholar] [CrossRef]
- Bronson, M.W.; Hillenmeyer, S.; Park, R.W.; Brodsky, A.S. Estrogen coordinates translation and transcription, revealing a role for NRSF in human breast cancer cells. Mol. Endocrinol. 2010, 24, 1120–1135. [Google Scholar] [CrossRef]
- Sankar, S.; Gomez, N.C.; Bell, R.; Patel, M.; Davis, I.J.; Lessnick, S.L.; Luo, W. EWS and RE1-Silencing Transcription Factor Inhibit Neuronal Phenotype Development and Oncogenic Transformation in Ewing Sarcoma. Genes Cancer 2013, 4, 213–223. [Google Scholar] [CrossRef]
- Varghese, B.V.; Koohestani, F.; McWilliams, M.; Colvin, A.; Gunewardena, S.; Kinsey, W.H.; Nowak, R.A.; Nothnick, W.B.; Chennathukuzhi, V.M. Loss of the repressor REST in uterine fibroids promotes aberrant G protein-coupled receptor 10 expression and activates mammalian target of rapamycin pathway. Proc. Natl. Acad. Sci. USA 2013, 110, 2187–2192. [Google Scholar] [CrossRef]
- Cho, E.; Moon, S.M.; Park, B.R.; Kim, D.K.; Lee, B.K.; Kim, C.S. NRSF/REST regulates the mTOR signaling pathway in oral cancer cells. Oncol. Rep. 2015, 33, 1459–1464. [Google Scholar] [CrossRef]
- Tawadros, T.; Martin, D.; Abderrahmani, A.; Leisinger, H.J.; Waeber, G.; Haefliger, J.A. IB1/JIP-1 controls JNK activation and increased during prostatic LNCaP cells neuroendocrine differentiation. Cell Signal 2005, 17, 929–939. [Google Scholar] [CrossRef]
- Cortes-Sarabia, K.; Alarcon-Romero, L.D.C.; Mendoza-Catalan, M.A.; Carpio-Pedroza, J.C.; Castaneda-Saucedo, E.; Ortuno-Pineda, C. REST/NRSF Silencing Modifies Neuronal Gene Expression in siRNA-Treated HeLa Cells: A Preliminary Exploration in the Search for Neuronal Biomarkers of Cervical Cancer. Medicina 2023, 59, 537. [Google Scholar] [CrossRef]
- Zhang, D.; Li, Y.; Wang, R.; Li, Y.; Shi, P.; Kan, Z.; Pang, X. Inhibition of REST Suppresses Proliferation and Migration in Glioblastoma Cells. Int. J. Mol. Sci. 2016, 17, 664. [Google Scholar] [CrossRef]
- Deng, P.; Zuo, Y.; Feng, S.; Li, Z.; Chen, W.; Li, H.; Wang, X. Knockdown of NRSF inhibits cell proliferation of ovarian cancer via activating Hippo pathway. Life Sci. 2018, 215, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Degirmenci, V.; Xin, H.; Li, Y.; Wang, L.; Chen, J.; Hu, X.; Zhang, D. PEI-Coated Fe3O4 Nanoparticles Enable Efficient Delivery of Therapeutic siRNA Targeting REST into Glioblastoma Cells. Int. J. Mol. Sci. 2018, 19, 2230. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, D.; Zhang, T.; Zhao, F.; Lang, H.; Lin, X.; Pang, X. The differentiation of human MSCs derived from adipose and amniotic tissues into insulin-producing cells, induced by PEI@Fe3O4 nanoparticles-mediated NRSF and SHH silencing. Int. J. Mol. Med. 2018, 42, 2831–2838. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, T.; Hsieh, J.; Nakashima, K.; Taira, K.; Gage, F.H. A small modulatory dsRNA specifies the fate of adult neural stem cells. Cell 2004, 116, 779–793. [Google Scholar] [CrossRef]
- Kuwabara, T.; Hsieh, J.; Nakashima, K.; Warashina, M.; Taira, K.; Gage, F.H. The NRSE smRNA specifies the fate of adult hippocampal neural stem cells. Nucleic Acids Symp. Ser. 2005, 49, 87–88. [Google Scholar] [CrossRef]
- Immaneni, A.; Lawinger, P.; Zhao, Z.; Lu, W.; Rastelli, L.; Morris, J.H.; Majumder, S. REST-VP16 activates multiple neuronal differentiation genes in human NT2 cells. Nucleic Acids Res. 2000, 28, 3403–3410. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kameoka, S.; Gopalakrishnan, V.; Aldape, K.D.; Pan, Z.Z.; Lang, F.F.; Majumder, S. Conversion of myoblasts to physiologically active neuronal phenotype. Genes Dev. 2004, 18, 889–900. [Google Scholar] [CrossRef]
- Ueda, H.; Kurita, J.I.; Neyama, H.; Hirao, Y.; Kouji, H.; Mishina, T.; Kasai, M.; Nakano, H.; Yoshimori, A.; Nishimura, Y. A mimetic of the mSin3-binding helix of NRSF/REST ameliorates abnormal pain behavior in chronic pain models. Bioorg. Med. Chem. Lett. 2017, 27, 4705–4709. [Google Scholar] [CrossRef]
- Kawase, H.; Ago, Y.; Naito, M.; Higuchi, M.; Hara, Y.; Hasebe, S.; Tsukada, S.; Kasai, A.; Nakazawa, T.; Mishina, T.; et al. mS-11, a mimetic of the mSin3-binding helix in NRSF, ameliorates social interaction deficits in a prenatal valproic acid-induced autism mouse model. Pharmacol. Biochem. Behav. 2019, 176, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Higo, J.; Takashima, H.; Fukunishi, Y.; Yoshimori, A. Generalized-ensemble method study: A helix-mimetic compound inhibits protein-protein interaction by long-range and short-range intermolecular interactions. J. Comput. Chem. 2021, 42, 956–969. [Google Scholar] [CrossRef]
- Conforti, P.; Zuccato, C.; Gaudenzi, G.; Ieraci, A.; Camnasio, S.; Buckley, N.J.; Mutti, C.; Cotelli, F.; Contini, A.; Cattaneo, E. Binding of the repressor complex REST-mSIN3b by small molecules restores neuronal gene transcription in Huntington’s disease models. J. Neurochem. 2013, 127, 22–35. [Google Scholar] [CrossRef]
- Kurita, J.I.; Hirao, Y.; Nakano, H.; Fukunishi, Y.; Nishimura, Y. Sertraline, chlorprothixene, and chlorpromazine characteristically interact with the REST-binding site of the corepressor mSin3, showing medulloblastoma cell growth inhibitory activities. Sci. Rep. 2018, 8, 13763. [Google Scholar] [CrossRef]
- Leone, S.; Mutti, C.; Kazantsev, A.; Sturlese, M.; Moro, S.; Cattaneo, E.; Rigamonti, D.; Contini, A. SAR and QSAR study on 2-aminothiazole derivatives, modulators of transcriptional repression in Huntington’s disease. Bioorg. Med. Chem. 2008, 16, 5695–5703. [Google Scholar] [CrossRef] [PubMed]
- Charbord, J.; Poydenot, P.; Bonnefond, C.; Feyeux, M.; Casagrande, F.; Brinon, B.; Francelle, L.; Auregan, G.; Guillermier, M.; Cailleret, M.; et al. High throughput screening for inhibitors of REST in neural derivatives of human embryonic stem cells reveals a chemical compound that promotes expression of neuronal genes. Stem Cells 2013, 31, 1816–1828. [Google Scholar] [CrossRef] [PubMed]
- Paonessa, F.; Criscuolo, S.; Sacchetti, S.; Amoroso, D.; Scarongella, H.; Pecoraro Bisogni, F.; Carminati, E.; Pruzzo, G.; Maragliano, L.; Cesca, F.; et al. Regulation of neural gene transcription by optogenetic inhibition of the RE1-silencing transcription factor. Proc. Natl. Acad. Sci. USA 2016, 113, E91–E100. [Google Scholar] [CrossRef]
- Criscuolo, S.; Gatti Iou, M.; Merolla, A.; Maragliano, L.; Cesca, F.; Benfenati, F. Engineering REST-Specific Synthetic PUF Proteins to Control Neuronal Gene Expression: A Combined Experimental and Computational Study. ACS Synth. Biol. 2020, 9, 2039–2054. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Gene structure and protein structure of REST. (A) Human REST is located in Chromosome 4q12, and it has five exons. The exon 5 was found to be mutually exclusive of exon 4. (B) The full-length human REST consists of 1097 aa, and there is a DNA binding domain (DBD), an N-terminal repressor domain (NRD), and a C-terminal repressor domain (CRD) for its regulatory activities. ZF indicates zinc finger. (C) The protein structure was predicated using Alphafold2.
Figure 1.
Gene structure and protein structure of REST. (A) Human REST is located in Chromosome 4q12, and it has five exons. The exon 5 was found to be mutually exclusive of exon 4. (B) The full-length human REST consists of 1097 aa, and there is a DNA binding domain (DBD), an N-terminal repressor domain (NRD), and a C-terminal repressor domain (CRD) for its regulatory activities. ZF indicates zinc finger. (C) The protein structure was predicated using Alphafold2.

Figure 2.
REST modulates gene activation via histone modification. REST binds to the RE1 motif within target genes and recruits Sin3A/B and CoREST as beacons for the recruitment of coregulators to form regulatory complexes. The chromatin was silenced via histone 3/4 deacetylation by HDACs, H3K9 methylation by G9a and SUV39H1, and H3K4 demethylation by LSD1 and H4K8 acetylation by BRG1. Further, the H3K36 methylation mediated by Tet1/3 might activate gene transcription.
Figure 2.
REST modulates gene activation via histone modification. REST binds to the RE1 motif within target genes and recruits Sin3A/B and CoREST as beacons for the recruitment of coregulators to form regulatory complexes. The chromatin was silenced via histone 3/4 deacetylation by HDACs, H3K9 methylation by G9a and SUV39H1, and H3K4 demethylation by LSD1 and H4K8 acetylation by BRG1. Further, the H3K36 methylation mediated by Tet1/3 might activate gene transcription.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jin, L.; Liu, Y.; Wu, Y.; Huang, Y.; Zhang, D. REST Is Not Resting: REST/NRSF in Health and Disease. Biomolecules 2023, 13, 1477. https://doi.org/10.3390/biom13101477
AMA Style
Jin L, Liu Y, Wu Y, Huang Y, Zhang D. REST Is Not Resting: REST/NRSF in Health and Disease. Biomolecules. 2023; 13(10):1477. https://doi.org/10.3390/biom13101477
Chicago/Turabian StyleJin, Lili, Ying Liu, Yifan Wu, Yi Huang, and Dianbao Zhang. 2023. "REST Is Not Resting: REST/NRSF in Health and Disease" Biomolecules 13, no. 10: 1477. https://doi.org/10.3390/biom13101477
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.