

Recognition of Arboviruses by the Mosquito Immune System



Department of Biochemistry and Molecular Biology, College of Agriculture, Biotechnology & Natural Resources, University of Nevada, Reno, NV 89557, USA
*
Author to whom correspondence should be addressed.
Biomolecules 2023, 13(7), 1159; https://doi.org/10.3390/biom13071159
Submission received: 16 June 2023
/
Revised: 12 July 2023
/
Accepted: 18 July 2023
/
Published: 21 July 2023
(This article belongs to the Special Issue New Insight into Vector Borne Diseases)
Abstract
:Arthropod-borne viruses (arboviruses) pose a significant threat to both human and animal health worldwide. These viruses are transmitted through the bites of mosquitoes, ticks, sandflies, or biting midges to humans or animals. In humans, arbovirus infection often results in mild flu-like symptoms, but severe disease and death also occur. There are few vaccines available, so control efforts focus on the mosquito population and virus transmission control. One area of research that may enable the development of new strategies to control arbovirus transmission is the field of vector immunology. Arthropod vectors, such as mosquitoes, have coevolved with arboviruses, resulting in a balance of virus replication and vector immune responses. If this balance were disrupted, virus transmission would likely be reduced, either through reduced replication, or even through enhanced replication, resulting in mosquito mortality. The first step in mounting any immune response is to recognize the presence of an invading pathogen. Recent research advances have been made to tease apart the mechanisms of arbovirus detection by mosquitoes. Here, we summarize what is known about arbovirus recognition by the mosquito immune system, try to generate a comprehensive picture, and highlight where there are still gaps in our current understanding.
1. Introduction
Mosquitoes transmit the causative agents for many human infectious diseases that result in significant morbidity and mortality globally, especially throughout tropical and subtropical regions. These insect vectors transmit many medically important arthropod-borne viruses (arboviruses) belonging to a wide variety of virus families, with the majority of arboviruses belonging to Bunyavirales, Flaviviridae, and Togaviridae [1]. Depending on the virus, symptomatic arbovirus infections may result in high fever, joint pain, encephalitis, hemorrhagic fever, coma, seizures, and sometimes death [2]. Arboviral diseases have undergone significant emergence and resurgence over the last 20 years, including epidemics of chikungunya virus (CHIKV) and Zika virus (ZIKV) [3]. Although this is the result of a variety of factors, anthropogenic impacts are considered to be primary drivers for the increasing incidence of arbovirus infections. These catalysts include rapid growth in global population, excessive urbanization, the disruption of natural habitats, booming international travel, and climate change [4,5]. Controlling mosquito populations as well as virus transmission itself are major strategies to reduce the burden of arboviral diseases. In order to develop novel strategies to interrupt arbovirus transmission, there is a strong need to further elucidate the interactions between arboviruses and their mosquito vectors.
Arboviruses must be highly efficient at crossing the barriers present in their mosquito vectors to be transmitted [6]. Following a viremic blood meal, viruses have to pass through the midgut infection barrier to propagate in the midgut epithelium [6]. The viruses must then escape from the midgut tissue and systemically replicate in other tissues. Finally, viruses need to spread to and replicate in the salivary glands. The following blood feed can then transfer infectious saliva to a new host [6]. The natural transmission cycles of arboviruses rarely result in any discernible loss in fitness or behavioral changes to their mosquito vectors. Viruses tend to persist throughout the lives of infected mosquitoes, which allows for efficient viral transmission [7]. Numerous studies have suggested that the activation of cellular and humoral immune responses limit the replication of arboviruses in mosquitoes in vitro and in vivo (reviewed in [7,8,9,10]). Yet, one of the least understood aspects of antiviral immunity in mosquitoes is how viruses are initially detected by the immune system.
Innate immune responses are activated by pattern recognition receptors (PRRs), which recognize pathogen-associated molecular patterns (PAMPs) [11,12]. Mosquito antiviral immune responses (Figure 1) include several conserved signal transduction pathways that lead to antiviral effector gene expression and restrict arboviral replication. These pathways include the Janus kinase–signal transducer and activator of transcription (JAK/STAT), Toll, Mitogen-activated protein kinases (MAPK), and immune deficiency (Imd) pathways. While these transcriptional responses have been shown to limit viral replication in some contexts, the RNA interference (RNAi) pathway is considered the central antiviral mechanism in mosquitoes [13,14]. In addition to these pathways, mosquitoes possess other defenses, such as phagocytosis and melanization [15]. In most situations, PRRs are responsible for either starting signaling cascades or acting as opsonins. Any detailed understanding of PRRs and virus sensing is still quite scarce in mosquitoes, and knowledge obtained from other dipterans, such as Drosophila melanogaster, is often used as the most likely indicator for mosquito immune mechanisms. However, mosquitoes vary quite substantially in their life cycles and physiology and may share some immune mechanisms, but may also have evolved different mechanisms to sense and defend against viruses. This review will compile the current knowledge of how viral PAMPs are sensed by mosquitoes and what we know and suspect from other organisms, and we will highlight potential gaps in our current understanding of mosquitoes’ immune sensing of viruses.
2. Dicer2 and DExD/H RNA Helicases
Antiviral immunity, and the recognition of viruses in particular, through the RNAi pathway is the most well-understood of all the pathways involved in the arthropod antiviral immune response. The RNAi pathway is one of the major innate immune pathways that control virus infection in insects. Insect RNAi comprises three distinct pathways mediated by either short interfering RNAs (siRNAs), microRNAs (miRNA), or PIWI-interacting RNAs (piRNAs) [16]. Viral recognition through and the activation of the piRNA and miRNA pathways remains unclear, but virus-derived piRNAs are generated in multiple mosquito species to a number of viruses [17,18,19], and in silico analysis indicates that cellular mosquito miRNAs may recognize and bind directly to viral genomes [20]. Further, the exogenous siRNA pathway is clearly activated during virus infection in mosquitoes (and other arthropods) in response to arbovirus infection. This subject has been reviewed extensively in the past [14] so we will not go into detail here. The only step we would like to draw attention to is the first step in the RNAi response, which is viral double-stranded RNA (dsRNA) recognition [21].
Viral dsRNA is a major PAMP in the immune response of any organism. Mammalian Rig-I-like receptors (RLRs) are well-established sensors of viral dsRNA that promote an interferon response [22,23,24,25]. In insects, the endonuclease Dicer-2 (Dcr-2) is related to mammalian RLRs and also recognizes viral dsRNA, but processes it into virus-derived siRNAs [26,27]. The correct dsRNA processing and RNAi functioning of Drosophila and mosquito Dcr-2 is dependent on a DExD/H RNA helicase domain [28,29,30,31]. Proteins with this domain fall into a large superfamily of RNA helicases [32,33]. Out of all the DExD/H RNA helicases, Drosophila Dcr-2’s helicase domain is most closely related to the RLR helicase domain, and is also required for the upregulation of the vago gene following infection with Drosophila C virus (DCV) and Sindbis virus (SINV) [34]. Vago is a small cysteine-rich secreted polypeptide that acts like a cytokine, comparable to an interferon response [34]. Importantly, Vago was crucial for controlling DCV replication in Drosophila fat bodies [34]. These findings suggested an RNAi-independent role of Dcr-2 in insect antiviral immunity that resembles RLR signaling in mammals [35]. It was also observed that the recombinant Flock House virus protein B2, which binds dsRNA, and thus, inhibits Dcr-2 function, reduced vago expression in flies following infection with DCV, further implicating that Dcr-2 signaling through vago expression plays an antiviral role in response to dsRNA [34,36]. Similar to human RLRs [37], the helicase domain of Dcr-2 in flies and mosquitoes was found to be responsible for sensing defective viral genomes, which feed into viral DNA synthesis and boost the siRNA response [35,38]. Whether there is intracellular immune signaling associated with this sensing is unknown.
In mosquitoes, Dcr-2 has been shown to activate the NF-κB transcription factor Rel2 in Culex quinquefasciatus Hsu cells in response to West Nile virus (WNV) infection [39,40]. Following the activation of Rel2, the expression of vago was upregulated, leading to the activation of the JAK/STAT pathway (see Section 5) and the expression of antiviral genes [39,40]. This finding established Vago as an antiviral cytokine-like molecule in mosquitoes. Vago expression is also induced in Aedes albopictus cells after dengue virus (DENV) infection, although this was not shown to be dependent on Dcr-2 [40]. In addition, vago upregulation was not observed in two studies that treated Aedes aegypti Aag2 cells with a dsRNA mimic, poly(I:C) [41,42]. However, vago upregulation may require nucleic acid features that are unique to viral dsRNA or may depend on the exact site of virus replication [34]. Altogether, there is evidence that dsRNA acts as a viral PAMP for mosquito Dcr-2 sensing that leads to the activation of RNAi-independent immune signaling pathways, but it remains to be seen whether this is conserved across mosquito species and which viruses are impacted by this immune response.
Mosquitoes also encode DExD/H RNA helicases that differ phylogenetically from the RLR subfamily, including DEAD- and DEAH-box helicases [43,44]. Many of these are responsible for sensing viral infection in other organisms [45,46,47,48] and some have been implicated in antiviral responses in insects (Table 1). For instance, in Drosophila, DDX56 binds CHIKV RNA and limits infection [49]. Taschuk et al. also found five other DEAD-box helicases in Drosophila that were antiviral against SINV [49]. Recent efforts have also implicated select Ae. aegypti helicases in the response to virus infection. The Ae. aegypti DEAD-box helicase ME31B, an ortholog of mammalian DDX6, was recently shown to bind the subgenomic flavivirus RNA (sfRNA) of WNV and ZIKV and to exhibit antiviral activity against these viruses in Ae. aegypti Aag2 cells [50]. However, ME31B binding to sfRNA was not shown to initiate an immune response. It was proposed that sfRNA binding actually enhances viral replication by sequestering ME31B, thereby counteracting its antiviral role [50]. In Drosophila, ME31B is antiviral against La Crosse virus (LACV) and Rift Valley fever virus (RVFV) via its canonical role of depleting the available pool of host capped mRNAs available for cap snatching [51]. In humans, DDX6 was found to be a co-sensor with RIG-I for influenza virus and enterovirus RNA that mediated the antiviral interferon response [52,53]. DDX6 has also been implicated in a proviral state for several arboviruses in human cells [48]. While ME31B has clearly been indicated in mosquito and fly antiviral responses, it remains unclear whether it acts as a classic PRR and, if so, which downstream antiviral responses are activated.
Another DEAD-box helicase in Drosophila, Rm62, binds bunyaviral RNA and restricts LACV and RVFV replication but not SINV or vesicular stomatitis virus (VSV) in cells and flies [54]. The binding of RVFV RNA by the human ortholog DDX17 appears not to trigger interferon signaling, but restricts replication directly [54]. The Ae. aegypti ortholog of Rm62 was found to bind CHIKV nsP3 in Aag2 cells [55], but the consequences of this interaction need to be investigated further. The antiviral effect of human DDX17 has so far only been suggested to be a result of viral RNA degradation, either alone or as a co-factor with the zinc-finger antiviral protein (ZAP) [54,56]. The putative role of mosquito Rm62 in antiviral defenses and virus sensing remains unclear, but existing evidence warrants further investigation.
The Ae. aegypti ortholog of mammalian DHX15, a DEAH-box helicase, is antiviral against CHIKV, although this appears to be dependent on its function in regulating glycolysis as opposed to sensing virus infection directly [57]. However, there is considerable evidence that human DHX15 acts as a PRR for dsRNA in response to virus infection [46]. In addition to DHX15, Machado et al. identified two other broadly acting antiviral DExD/H-box helicases in mosquitoes [57]. One of these is an ortholog of human DHX9, which is known to sense viral nucleic acids and can lead to a pro- or antiviral outcome depending on the virus and host context [46].
Another Ae. aegypti DExH domain-containing protein, Spindle-E, is antiviral against two alphaviruses in Aag2 cells [58]. Spindle-E also contains a Tudor domain and was thus hypothesized to play a role in the RNAi response, like other Tudor domain-containing proteins [59]. Yet, its antiviral activity was independent of the RNAi response, suggesting other antiviral mechanisms that may include dsRNA sensing and immune signaling [58].
Considering their ability to sense viral nucleic acids and proteins in other organisms, and current evidence from mosquitoes, DExD/H-box RNA helicases may represent an important and understudied class of PRRs in mosquitoes that warrant additional research.
3. The IMD Pathway
The immunodeficiency (IMD) pathway is an inducible signaling pathway that leads to the activation of the NF-κB transcription factor Rel2 (Relish in Drosophila) and subsequent antimicrobial peptide (AMP) production [60,61,62]. It has been well studied for its involvement in non-viral pathogen defenses, which typically are mediated through the peptidoglycan-recognition protein (PGRP) recognition of bacteria and fungi in Drosophila [63,64,65]. In Anopheles gambiae, the IMD pathway regulates Plasmodium and bacterial infection [66]. For Plasmodium defense, PGRP-LC, PGRP-LA, and PGRP-S2/PGRP-S3 all play a role in Anopheles coluzzi, while PGRP-LB promotes permissiveness to infection [67]. Rel2 and other IMD pathway components protect Ae. aegypti from Gram-negative and Gram-positive bacteria [62,68]. The antiviral properties of the Drosophila IMD pathway have also been demonstrated [69]. Conflicting reports about whether the IMD pathway plays an antiviral role in mosquitoes currently exist (reviewed in [8]). It is also worth noting that Rel2 may not be strictly linked to just the IMD pathway. As previously described, Dcr-2 can also activate Rel2, seemingly independently of IMD [34,39,40].
It has been found that microbiota present in the midgut of Ae. aegypti increase Rel2 levels [70]. Thus, studying the putative antiviral effects of Rel2 may be confounded by changes in the bacterial microbiome, since blood feeding can trigger nutrient-induced growth in microbiota and the associated Rel2 upregulation [70]. IMD’s involvement in the antiviral response may, therefore, depend, in part, on bacterial PRRs. In line with this, it has been shown that heat-killed bacteria can restore the IMD-dependent antiviral defense of flies that have been treated with antibiotics [71]. Additionally, pgrp-lc mutant flies have increased ZIKV and DCV loads after virus challenge, further confirming the role of bacterial sensing in antiviral responses [71,72].
Yet, some direct virus-induced activation and upregulation of IMD pathway components has been observed. In Ae. aegypti mosquitoes, blood-fed SINV increased Rel2 expression compared to a non-infectious control, suggesting the presence of a virus-specific receptor that can upregulate an important component of the IMD pathway [70]. Further, Sansone et al. demonstrated that DCV protection in flies is dependent on both bacterial and viral stimulation of the IMD pathway [71]. Another study showed that the knockdown of PGRP-LC had no effect on the replication of a SINV replicon in flies, while other IMD pathway components did have an effect [73]. However, Barletta et al. depleted Caspar, the negative regulator of Rel2, as a method of activating the IMD pathway, and this led to decreased microbiota and increased SINV levels [70]. These findings further highlight the importance of bacterial contributions and microbiome balance in antiviral immunity [70]. Finally, one of the previously mentioned studies using poly(I:C) to mimic dsRNA stimulation showed the upregulation of genes downstream of IMD and Rel2 signaling in Aag2 cells [41]. While dsRNA sensing may not signal directly through IMD, it can be sensed by other proteins (e.g., Dcr-2) that activate Rel2, bypassing the canonical IMD pathway. In line with this, a recent paper showed that the RVFV infection of Ae. aegypti Aag2 cells can prime immune responses, resulting in increased expression of certain bacterial PRRs and an enhanced response to bacterial challenge [74]. These data indicate that RVFV infection is indeed sensed by the mosquito immune system, resulting in increased IMD pathway sensitivity.
4. Toll Receptors
In mammals, the function of Toll-like receptors (TLRs) as PRRs for a wide array of ligand types from many different pathogens is relatively well understood [75]. Mammalian TLRs can sense a variety of microbial PAMPs, including those of viruses. TLR3 is perhaps the most studied in terms of virus infection as its ectodomain binds dsRNA, leading to a pro-inflammatory immune response [76,77,78]. It has now been documented that TLRs 2, 3, 6, 7, 8, 9, and 10 all recognize viruses to some extent, either through foreign nucleic acid sensing or the detection of viral surface proteins [75,79,80].
The canonical immune responsive Toll pathway in Drosophila functions through the detection of a PAMP by a PRR that triggers cleavage of the extracellular ligand Spaetzle, which then binds to and activates the transmembrane Toll-1 receptor [81,82]. Similar to the IMD pathway, this cascade, in turn, leads to the activation of the NF-κB transcription factors Dorsal and Dif in Drosophila, or Rel1A/B in mosquitoes, and AMP expression [83,84,85]. Bacterial and fungal PRRs that are known to activate Spaetzle and Spaetzle-like proteins include Gram-negative binding protein 1, PGRP SA, and Gram-positive specific serine protease [86,87]. A Toll receptor in Ae. aegypti, Toll5a, recently had its structure solved and was shown to bind to Spaetzle1C [88]. Spaetzle-like proteins themselves have been shown to contribute to antiviral immunity against several viruses in Drosophila, often by upregulating AMP expression [89,90,91]. However, it is still unclear which PRR is responsible for the cleavage of Spaetzle-like proteins in response to viral infection in flies or mosquitoes. Nonetheless, several studies have shown other Toll pathway components to be antiviral in An. gambiae and Ae. aegypti, although specific Toll receptors or ligands have not been discovered [92,93,94].
Drosophila Toll-1 and Toll-7 have been shown to bind VSV directly, which actually resulted in the induction of the autophagy pathway [89,95]. A Toll receptor from the insect vector Laodelphax striatellus bound Rice-Stripe virus nucleocapsid protein and contributed to antiviral defenses, although this did not seem to be dependent on other canonical Toll-pathway components either [96]. Additionally, Toll-7 was required for the autophagy-dependent restriction of RVFV in Drosophila [97]. However, others have found that Toll-7 is not required for Drosophila resistance to VSV and autophagy plays a minor role [98]. Toll-1 and Toll-7 binding to VSV and Spaetzle-like proteins did, however, result in AMP promoter activation [89]. Further complicating the determination of the source of activation and which pathway is induced, a study found that, in Ae. aegypti, Wolbachia-mediated reactive oxygen species activated the Toll pathway and led to the upregulation of AMPs and the inhibition of DENV replication [99].
The Toll gene family has expanded in most mosquito species compared Drosophila [61,100]. For example, two mosquito species have experienced Toll-9 gene duplications [100]. In addition, mosquitoes have gained two new clades, Toll10 and 11, while losing two others, Toll2 and 3. Inferences of function based on homology alone may not be sufficient for these Toll genes, especially those that have undergone taxa-specific expansion. There is a need to determine Toll gene function experimentally [101]. Of course, homology inquiries have been used as launching points for successful studies. For example, based on in-depth phylogenetic analysis and homology modeling of human TLR4, Bombyx mori Toll-9 was studied and shown to recognize lipopolysaccharides [102]. Another recent study found that a dsRNA binding site of TLR3 is conserved in Ae. aegypti Toll6 (AaToll6) [42]. Importantly, AaToll6 was upregulated in Aag2 cells after treatment with poly(I:C) [42]. Together, these findings were used to suggest that AaToll6 is a PRR-recognizing dsRNA. In another study, however, poly(I:C) did not activate the Rel1A promoter region in Aag2 cells, and instead, induced Rel2-dependent expression [41]. The discrepancy in these two studies may be due, in part, to experimental differences, such as the method of poly(I:C) delivery [103]. For instance, in mammals, TLR3 primarily senses dsRNA in endosomes while RLRs sense it in the cytoplasm [104]. The delivery and location of dsRNA within the cell may thus impact the type of immune response that is elicited. Overall, evidence for AaToll6 or any other Toll receptor directly interacting with a viral PAMPs in response to virus infection in mosquitoes is still lacking. It is likely that, in some cases, upstream viral PRRs are responsible for activating the Toll pathway in mosquitoes, perhaps through Spaetzle.
5. The JAK/STAT Pathway
In Drosophila, the JAK/STAT signaling pathway begins with the activation of Unpaired (Upd) ligands (Upd, Upd2, Upd3) that signal through the receptor Domeless (Dome). How (and if) Upd ligands are activated in response to viral infection is still unknown, but it may be mediated by insulin signaling (see Section 6) [105]. Ultimately, the pathway concludes with the activation of the transcription factor STAT and downstream antiviral defenses, depending on the virus/insect combination (reviewed in [84]). In mosquitoes, Upd ligands have not yet been identified [8]. However, the cytokine-like molecule Vago was shown to activate the JAK/STAT pathway in Cx. Quinquefasciatus Hsu cells, with downstream antiviral effects on WNV replication [39]. As discussed in Section 2, vago expression was upregulated after WNV infection via Dcr-2/Rel2 signaling, implicating Dcr-2 as a PRR for JAK/STAT pathway activation. However, the receptor Dome was not necessary for anti-WNV action, prompting the question of whether another JAK/STAT receptor exists [39]. Recently, a study on kuruma shrimp (Marsupenaeus japonicus) identified integrin as a receptor for Vago that was essential for antiviral activity against white spot syndrome virus (WSSV) [106]. This study also identified ficolin as the antiviral effector molecule upregulated following Vago-mediated JAK/STAT signaling that bound the virus and promoted its clearing [106]. However, in this instance, Dcr-2’s involvement in vago upregulation was not investigated. It is unknown what other mosquito PRRs are responsible for activating the JAK/STAT pathway in response to viral infection, although recent work has identified some possible mechanisms (see Section 7). Insulin may also play a role (see Section 6).
6. MAPKs
MAPKs are involved in signaling cascades that have recently emerged as components of mosquito antiviral immunity (reviewed in [8]). In mammals, a diverse set of stimuli can activate MAPKs, which, in turn, regulate the activity of target proteins, including transcription factors [107]. Two conventional MAPKs include extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK), which are conserved in mosquitoes [108]. Many reports have now demonstrated that the ERK pathway is induced by virus infection in mosquitoes, serves an antiviral role, and, importantly, can be enhanced by the mammalian insulin present in a bloodmeal [109,110,111]. Ahlers et al. showed that Upd and JAK/STAT pathway activation was dependent on insulin-mediated ERK pathway activation [110]. The microbiota-dependent stimulation of the IMD pathway, discussed above, was also shown to lead to the activation of the ERK pathway in flies [71]. These reports point towards the stimulation of ERK being largely dependent on non-viral stimuli.
Interestingly, Liu et al. showed that silencing JNK in Ae. Albopictus C6/36 cells had no impact on DENV2 RNA levels, while another study saw increased DENV2, ZIKV, and CHIKV RNA levels after silencing Kayak, a JNK pathway core component, in Ae. Aegypti salivary glands [109,112]. This discrepancy may be due to a species-specific response and/or represent a difference between cell-culture and in vivo studies. Nonetheless, how the JNK pathway is activated remains largely unknown. In Drosophila, the JNK pathway can be activated through the PGRP-LC recognition of peptidoglycan [113]. However, similar activation is not observed in C6/36 cells [114]. Chowdhury et al. proposed that the antiviral JNK pathway activity they observed was due to infection-induced oxidative stress [112]. Whether this is the extent of JNK activation in response to viral infection or whether any direct virus sensing can activate JNK remains to be seen.
7. TEPs and LRRs
The complement system has long been shown to be a crucial component of vertebrate innate immunity [115]. At its core, the C3 protein, one of the most abundant circulating plasma proteins, is involved in all three complement activation pathways, which all lead to opsonization and contribute to the clearing of pathogens [116,117]. C3, along with additional complement factors, is part of the thioester-containing protein (TEP) family. These proteins share a conserved and highly reactive thioester CGEQ amino acid motif [118,119]. In vertebrates, there are two subfamilies of TEPs—complement factors and the closely related α2-macroglobulins, which also serve a role in innate immunity [120]. An additional TEP subfamily specific to insects was described through studies on Drosophila. These complement-like insect TEPs (iTEPs) were identified based on high sequence similarity to the C3/α2-macroglobulin family [121]. One of the roles of these iTEPs may be as PRRs upstream of the Toll pathway, as indicated by reduced Toll signaling after microbial infection in flies lacking all four of the secreted iTEPs [122].
A mosquito iTEP, An. gambiae TEP1 (AgTEP1), has been characterized in detail and shown to be of high structural and functional similarity to vertebrate C3 [123]. Functional analysis showed the ability of AgTEP1 to recognize, directly bind, and promote the clearing of Gram-negative bacteria and Plasmodium parasites [124,125]. The antiviral activity of iTEPs was showcased in mosquitoes through the silencing of Ae. aegypti TEP1 (AaTEP1), which led to a significant increase in WNV and DENV2 load [126]. A more recent study confirmed AaTEP1 to have anti-DENV activity in Ae. aegypti mosquitoes and suggested that this activity was the result of enhanced AMP production [127]. Interestingly, in these TEP1-depleted transgenic mosquitoes, rel2 expression, but not rel1 expression, was suppressed compared to wild-type mosquitoes following a bloodmeal [127]. These results indicate a possible link between AaTEP1 and the IMD pathway, although Rel2 expression can be driven by enhanced microbiota growth after a bloodmeal [70] (see Section 3 above). These results also further highlight the complex interactions between the mosquito immune response, the microbiome, and virus replication. Another TEP, AaTEP20, impacted ZIKV replication and AMP levels when silenced in mosquitoes [112]. In none of these studies, however, were AaTEPs investigated directly for their role in sensing viruses.
Insects all possess a varying number of iTEP genes, possibly suggesting an evolutionary expansion ba sed on pathogen-specific recognition needs [128]. Similarly, a repertoire of host proteins with a leucine-rich repeat (LRR) motif contribute to the recognition of variable PAMPs in animals and plants and even comprise the foundation for adaptive immunity in agnathans [129,130]. This LRR domain is conserved across many families of immune proteins and species [131]. Copies of these motifs form the structural basis of mammalian TLRs, facilitate TLR pathogen binding in humans, and are also present in mosquito and tick Toll receptors [132,133,134,135]. The AgTEP1 binding of Plasmodium was found to be dependent on two LRR immune proteins (LRIM 1 and 2) [129,136,137]. In silico analyses later revealed many LRR-containing genes in mosquito disease vectors that share similar features with LRIM 1 and 2, and therefore, were proposed to represent a diversified set of PRRs in mosquitoes [138]. Differential gene expression analysis showed two LRR-containing genes to be downregulated compared to other immune genes after infection with ZIKV and CHIKV in Ae. aegypti [139]. In addition, an LRR-containing gene was enriched upon challenge with DENV in a genetic DENV-refractory strain of Ae. aegypti, a change that perhaps confers DENV restriction [139,140].
An RNAi screen revealed the Drosophila macroglobulin/complement-related subfamily (DmMCR) as another functionally significant class of TEPs in insects. DmMCR was found to bind Candida albicans and promote phagocytosis in Drosophila [141]. Although often grouped under iTEPs, phylogenetic analysis reveals unique structural properties of MCRs [119]. Most notably, DmMCR (AKA DmTEP6) and its mosquito orthologs AgMCR (AKA AgTEP13) and AaMCR (AKA AaTEP13) all have a mutated thioester motif [61,128,142]. In contrast to iTEPs, MCRs are highly conserved among arthropods [61]. Together, these differences suggest a mechanism of pathogen recognition separate from iTEPs. AaMCR was shown to bind the E protein of DENV via an adaptor molecule, an Ae. aegypti homolog of scavenger receptor-C (AaSR-C) (see Section 8), suggesting another mechanism of arbovirus recognition by mosquito TEPs and related proteins [142]. The recognition of DENV by AaSR-C and AaMCR was also shown to regulate the expression of AMPs exerting anti-DENV activity, directly linking virus recognition to an antiviral signal transduction immune response [142]. Importantly, the AMPs regulated by the AaMCR/AaSR-C system share an overlap with Toll and JAK/STAT-regulated genes [142,143].
8. Scavenger Receptors
Scavenger receptors are a large family of heterogeneous molecules that are well-established to recognize danger-associated molecular patterns and PAMPs in other organisms, as well as carry out functions such as binding lipoproteins [144] (reviewed in [145,146]). They have been divided into 10 classes based on structural homology [147,148]. Class-C SRs consist of one member that was first described in Drosophila and is absent in mammals [148,149]. Since its discovery, it has become recognized as a PRR, primarily for Gram-negative and Gram-positive bacteria [150]. However, as stated above, an Ae. aegypti homologue was discovered and found to bind directly to DENV [142]. Studies on other arthropods have also identified SR-C orthologs, including in the kuruma shrimp, where it binds WSSV and promotes its clearance [151]. Another type of SR, class-A1, was recently described and shown to bind CHIKV’s NSP1 via Co-IP in mice, which facilitated virus control by inducing autophagy [152]. In addition, the E2 protein of multiple alphaviruses can be bound by the mouse scavenger receptor SR-A6 (MARCO) to induce the clearing of a circulating virus independent of the complement system or natural antibodies [153].
Multiple scavenger receptors are conserved across mosquito species genomes, and it would be worth investigating whether any have antiviral activity [100,154]. So far, only a few SRs besides DmSRC have been studied in Drosophila, and only in the context of antibacterial immunity (reviewed in [12]). It is important to note, however, that there are also cases of viruses using SRs to facilitate cellular entry [145]. For example, a high-density lipoprotein receptor, SR-B1, is responsible for internalizing ZIKV and DENV by binding the NS1 of both viruses in human Huh7 and Ae. albopictus C6/36 cells [155].
Besides promoting the autophagy of infected cells and being involved in the phagocytosis of microbes, including viruses [145,151], the overexpression of SR-C in B. mori was recently found to upregulate Toll pathway components and AMPs [156]. Indeed, it has been known for some time that SRs interact with TLRs in mammals as co-receptors required for NF-κB signaling in response to bacterial infections [157,158,159]. Additionally, SR-A1 is expressed on the surface of human epithelial cells and can bind dsRNA to mediate its uptake and downstream signaling [160]. Later, it was also shown that SR-A1 is needed for the RLR and TLR3 sensing of dsRNA [161]. In this case, SR-A1 is thought to act as a gatekeeper that delivers dsRNA to the established receptors inside the cell [161]. Ultimately, it was shown that SR-A1 is necessary for the TLR3-mediated antiviral response to hepatitis C virus [162]. Mosquitoes encode five class-A SRs, representing an interesting avenue for future research [100].
9. Lectins
Calcium-type (C-type) lectins (CTLs) describe a large group of proteins that bind carbohydrates in a calcium-dependent manner, although many CTL domains (CTLDs) have been found in proteins that do not bind sugar and/or depend on calcium [163,164]. CTLD-containing proteins are known to play a key role in the vertebrate immune system by acting as PRRs for a multitude of pathogens [165]. Mannose binding lectin (MBL), a secreted CTLD-containing protein, is responsible for recognizing and binding to N-linked glycans on WNV structural proteins in mice [166]. In addition, an ELISA assay showed MBL’s ability to bind DENV virions in humans as a crucial step in viral clearance [167].
In insects, two novel CTLs derived from the housefly, Musca domestica, exhibited significant antiviral activity in lepidopteran S9 cells [168]. However, while some mosquito CTLs (mosCTL) have antibacterial [169] and antifungal [170] properties, evidence of any mosCTL antiviral activity is lacking. In contrast, as has been seen above with other innate immune receptors, most mosCTLs that have been studied for their involvement in viral immunity act to facilitate infection [171], suggesting that viruses have evolved to hijack these proteins traditionally involved in immunity. In 2010, Cheng et al. identified mosquito galactose-specific binding CTL-1 (mosGCTL-1) in Aedes mosquitoes as crucial for WNV infection after gene silencing, based on an RNAi screen of human proteins, and found that blood feeding with mosGCTL-1 antisera reduced vector competence [172,173]. WNV attachment to cells via mosGCTL-1 also required the membrane-bound mosPTP-1, an ortholog of human CD45 [172]. A similar effect with a Cx. quinquefasciatus mosGCTL-1 homolog was seen [172]. However, a more recent study failed to interfere with WNV infection using anti-mosGCTL-1 antibodies in Culex pipiens [174]. Additional mosCTLs have been shown to bind the viral envelopes of DENV and JEV, promoting their entry into mosquito cells [175,176]. Further advances in Ae. aegypti genome annotation have uncovered over 50 CTLs, a significant portion of which have unknown function in the mosquito immune response to viral infection [171]. It is thus entirely possible that other mosCTLs exhibit antiviral activity through virus binding and viral PAMP sensing.
10. Phenoloxidase Activity
Melanization is a key innate immune mechanism in mosquitoes and other invertebrates that is managed by the highly regulated phenoloxidase (PO) enzyme and results in hemocytes’ production of melanin, promoting pathogen clearance [177,178]. However, in some cases, immune defense is achieved without the production of melanin [179]. In some insect species, namely the moth Manduca sexta, the serine protease cascade that leads to PO activation has been studied at length [178,180]. Molecules involved in the early stages of PO activation have thus been elucidated. The hemolymph protease 14, for instance, binds bacteria directly and senses fungal infection through beta-1,3-glucan recognition proteins (GRP) [181,182,183,184,185]. Two CTLs in Drosophila seem to act as bacterial PRRs that promote melanization [186]. In a genetically selected strain of An. gambiae, melanization is responsible for refractoriness to Plasmodium, mediated by TEP1 activity [187]. PRRs leading to PO activation have been identified in mosquitoes, including a mosquito GRP capable of binding multiple pathogens in Armigeres subalbatus [188].
The activation of the PO pathway by insects in response to virus infection is also well documented. An early study showed that Heliothis virescens larval hemolymph demonstrated antiviral activity towards Sindbis virus in a PO- and melanization-dependent manner [189]. Virucidal activity against a baculovirus was a consequence of H2O2 produced from the PO activation cascade in Heliothis virescens [190]. Interestingly, a poly DNA virus carried by the parasitoid wasp Microplitis demolitor encodes a protein, Egf1.0, that acts as a serine proteinase inhibitor (serpin) to block the PO cascade, although this may be responsible for blocking anti-parasite activity as opposed to antiviral PO activity [191]. There are now numerous documented pathogen factors that act to block proPO activation in invertebrates (reviewed in [179]). Together, these findings point to a crucial role of PO activity in insect immunity, with its antiviral roles less clearly defined.
In conditioned media from Ae. albopictus U4.4 cells, a PO cascade resulted in decreased viability of the arbovirus Semliki Forest Virus (SFV), and this decrease was correlated with increased melanization activity [192]. In the same study, recombinant SFV expressing Egf1.0 inhibited U4.4 PO activity and enhanced virus spread in cell culture and in Ae. aegypti mosquitoes [192]. Whether TEPs, lectins, or other receptors are responsible for sensing viral infection in mosquitoes and triggering the proPO activation cascade is unknown. In Drosophila, crosstalk between the IMD, Toll, and PO pathways has been observed in response to bacterial challenge, further complicating the interpretation of studies into all of these antimicrobial pathways [193,194].
11. Conclusions and Future Research
Over the last two decades, significant advances have been made in the field of virus recognition in mosquitoes, encompassing studies on a variety of innate signaling pathways and humoral responses. Despite these efforts, a comprehensive picture with clear connections from the initial virus detection to the signaling and clarification of pathway crosstalk is still missing. Some information is derived from Drosophila, some from Ae. aegypti, and some from Cx. quinquefasciatus or other mosquitoes. Similarly, most studies understandably focus on one specific virus as opposed to employing a more broadly comparative approach. Few studies have looked at the first steps of virus recognition—which mosquito PRRs detect which viral PAMPs and trigger mosquito immune responses. There is a need for comparative immunology studies across both Aedes spp. and Culex spp. mosquitoes and multiple arbovirus families. We have compiled the various potential mechanisms by which viruses may be sensed in mosquitoes, highlighted potential crosstalk between pathways, and discussed the complex nature of microbiome-mediated immunity versus virus-induced immunity. All of the immune responses outlined above require further studies to improve our current understanding of how antiviral responses are elicited and how these mechanisms differ between various viruses and mosquito species. With the advancement of bioengineering techniques, including gene editing in mosquitoes [195,196,197] and progress in gene drive strategies [198,199], it has become more evident that we need to fully understand the fundamental virus–mosquito interactions to design the best possible targets for gene editing. Disruption of the immune system has already been shown to increase disease phenotypes in mosquitoes following infection [200], while transgenic activation of the JAK/STAT pathway reduced DENV virus replication in vivo [201]. However, the better our understanding of mosquito immune responses, the better we can predict the success and limitations of gene targets for arbovirus transmission control.
Author Contributions
Writing—original draft preparation, B.C.P., E.W., T.Z.B.T. and C.R.; writing—review and editing, B.C.P., E.W., T.Z.B.T. and C.R.; visualization, B.C.P. and E.W.; supervision, C.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
Figure 1 was generated using BioRender.com (accessed on 16 June 2023) under agreement number AR25HWCLPK.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vasilakis, N.; Gubler, D.J. Arboviruses: Molecular Biology, Evolution and Control; Caister Academic Press: Poole, UK, 2016; pp. 1–410. [Google Scholar]
- Lee, H.; Halverson, S.; Ezinwa, N. Mosquito-Borne Diseases. Prim. Care 2018, 45, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Higgs, S.; Vanlandingham, D.L. Emergence and re-emergence of mosquito-borne arboviruses. Curr. Opin. Virol. 2019, 34, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.A.; Stewart-Ibarra, A.M.; Estallo, E.L. Climate change and viral emergence: Evidence from Aedes-borne arboviruses. Curr. Opin. Virol. 2020, 40, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Ebel, G.D. How Do Virus-Mosquito Interactions Lead to Viral Emergence? Trends Parasitol. 2018, 34, 310–321. [Google Scholar] [CrossRef]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Antiviral Immunity and Virus-Mediated Antagonism in Disease Vector Mosquitoes. Trends Microbiol. 2018, 26, 447–461. [Google Scholar] [CrossRef]
- Tikhe, C.V.; Dimopoulos, G. Mosquito antiviral immune pathways. Dev. Comp. Immunol. 2021, 116, 103964. [Google Scholar] [CrossRef]
- Lee, W.S.; Webster, J.A.; Madzokere, E.T.; Stephenson, E.B.; Herrero, L.J. Mosquito antiviral defense mechanisms: A delicate balance between innate immunity and persistent viral infection. Parasites Vectors 2019, 12, 165. [Google Scholar] [CrossRef] [Green Version]
- Ruckert, C.; Bell-Sakyi, L.; Fazakerley, J.K.; Fragkoudis, R. Antiviral responses of arthropod vectors: An update on recent advances. Virusdisease 2014, 25, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Su, F.; Li, Q.; Zhang, J.; Li, Y.; Tang, T.; Hu, Q.; Yu, X.Q. Pattern recognition receptors in Drosophila immune responses. Dev. Comp. Immunol. 2020, 102, 103468. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.D.; Olson, K.E. The role of RNA interference (RNAi) in arbovirus-vector interactions. Viruses 2015, 7, 820–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, C.D. Mosquito RNAi is the major innate immune pathway controlling arbovirus infection and transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso-Jaime, V.; Tikhe, C.V.; Dong, S.; Dimopoulos, G. The Role of Mosquito Hemocytes in Viral Infections. Viruses 2022, 14, 2088. [Google Scholar] [CrossRef]
- Olson, K.E.; Blair, C.D. Arbovirus-mosquito interactions: RNAi pathway. Curr. Opin. Virol. 2015, 15, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Altinli, M.; Leggewie, M.; Schulze, J.; Gyanwali, R.; Badusche, M.; Sreenu, V.B.; Fuss, J.; Schnettler, E. Antiviral RNAi Response in Culex quinquefasciatus-Derived HSU Cells. Viruses 2023, 15, 436. [Google Scholar] [CrossRef]
- Varjak, M.; Leggewie, M.; Schnettler, E. The antiviral piRNA response in mosquitoes? J. Gen. Virol. 2018, 99, 1551–1562. [Google Scholar] [CrossRef]
- Walsh, E.; Torres, T.Z.B.; Ruckert, C. Culex Mosquito Piwi4 Is Antiviral against Two Negative-Sense RNA Viruses. Viruses 2022, 14, 2758. [Google Scholar] [CrossRef]
- Yen, P.S.; Chen, C.H.; Sreenu, V.; Kohl, A.; Failloux, A.B. Assessing the Potential Interactions between Cellular miRNA and Arboviral Genomic RNA in the Yellow Fever Mosquito, Aedes aegypti. Viruses 2019, 11, 540. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef]
- Paro, S.; Imler, J.L.; Meignin, C. Sensing viral RNAs by Dicer/RIG-I like ATPases across species. Curr. Opin. Immunol. 2015, 32, 106–113. [Google Scholar] [CrossRef] [Green Version]
- de Faria, I.J.; Olmo, R.P.; Silva, E.G.; Marques, J.T. dsRNA sensing during viral infection: Lessons from plants, worms, insects, and mammals. J. Interferon Cytokine Res. 2013, 33, 239–253. [Google Scholar] [CrossRef]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Blair, C.D.; Olson, K.E. Mosquito immune responses to arbovirus infections. Curr. Opin. Insect Sci. 2014, 3, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoa, N.T.; Keene, K.M.; Olson, K.E.; Zheng, L. Characterization of RNA interference in an Anopheles gambiae cell line. Insect Biochem. Mol. Biol. 2003, 33, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Nakahara, K.; Pham, J.W.; Kim, K.; He, Z.; Sontheimer, E.J.; Carthew, R.W. Distinct roles for Drosophila Dicer-1 and Dicer-2 in the siRNA/miRNA silencing pathways. Cell 2004, 117, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Donelick, H.M.; Talide, L.; Bellet, M.; Aruscavage, P.J.; Lauret, E.; Aguiar, E.; Marques, J.T.; Meignin, C.; Bass, B.L. In vitro studies provide insight into effects of Dicer-2 helicase mutations in Drosophila melanogaster. RNA 2020, 26, 1847–1861. [Google Scholar] [CrossRef]
- Kandasamy, S.K.; Zhu, L.; Fukunaga, R. The C-terminal dsRNA-binding domain of Drosophila Dicer-2 is crucial for efficient and high-fidelity production of siRNA and loading of siRNA to Argonaute2. RNA 2017, 23, 1139–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gestuveo, R.J.; Parry, R.; Dickson, L.B.; Lequime, S.; Sreenu, V.B.; Arnold, M.J.; Khromykh, A.A.; Schnettler, E.; Lambrechts, L.; Varjak, M.; et al. Mutational analysis of Aedes aegypti Dicer 2 provides insights into the biogenesis of antiviral exogenous small interfering RNAs. PLoS Pathog. 2022, 18, e1010202. [Google Scholar] [CrossRef]
- Luo, D.; Kohlway, A.; Pyle, A.M. Duplex RNA activated ATPases (DRAs): Platforms for RNA sensing, signaling and processing. RNA Biol. 2013, 10, 111–120. [Google Scholar] [CrossRef]
- Tanner, N.K.; Linder, P. DExD/H box RNA helicases: From generic motors to specific dissociation functions. Mol. Cell 2001, 8, 251–262. [Google Scholar] [CrossRef]
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Hoffmann, J.A.; Imler, J.L. The DExD/H-box helicase Dicer-2 mediates the induction of antiviral activity in drosophila. Nat. Immunol. 2008, 9, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Baldaccini, M.; Pfeffer, S. Untangling the roles of RNA helicases in antiviral innate immunity. PLoS Pathog. 2021, 17, e1010072. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.A.; Lee, J.H.; Chapados, B.R.; Debler, E.W.; Schneemann, A.; Williamson, J.R. Dual modes of RNA-silencing suppression by Flock House virus protein B2. Nat. Struct. Mol. Biol. 2005, 12, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Sanchez David, R.Y.; Combredet, C.; Sismeiro, O.; Dillies, M.A.; Jagla, B.; Coppee, J.Y.; Mura, M.; Guerbois Galla, M.; Despres, P.; Tangy, F.; et al. Comparative analysis of viral RNA signatures on different RIG-I-like receptors. Elife 2016, 5, e11275. [Google Scholar] [CrossRef]
- Poirier, E.Z.; Goic, B.; Tomé-Poderti, L.; Frangeul, L.; Boussier, J.; Gausson, V.; Blanc, H.; Vallet, T.; Loyd, H.; Levi, L.I.; et al. Dicer-2-Dependent Generation of Viral DNA from Defective Genomes of RNA Viruses Modulates Antiviral Immunity in Insects. Cell Host Microbe 2018, 23, 353–365.e358. [Google Scholar] [CrossRef] [Green Version]
- Paradkar, P.N.; Trinidad, L.; Voysey, R.; Duchemin, J.B.; Walker, P.J. Secreted Vago restricts West Nile virus infection in Culex mosquito cells by activating the Jak-STAT pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 18915–18920. [Google Scholar] [CrossRef]
- Paradkar, P.N.; Duchemin, J.B.; Voysey, R.; Walker, P.J. Dicer-2-dependent activation of Culex Vago occurs via the TRAF-Rel2 signaling pathway. PLoS Negl. Trop. Dis. 2014, 8, e2823. [Google Scholar] [CrossRef] [Green Version]
- Russell, T.A.; Ayaz, A.; Davidson, A.D.; Fernandez-Sesma, A.; Maringer, K. Imd pathway-specific immune assays reveal NF-κB stimulation by viral RNA PAMPs in Aedes aegypti Aag2 cells. PLoS Negl. Trop. Dis. 2021, 15, e0008524. [Google Scholar] [CrossRef]
- Angleró-Rodríguez, Y.I.; Tikhe, C.V.; Kang, S.; Dimopoulos, G. Aedes aegypti Toll pathway is induced through dsRNA sensing in endosomes. Dev. Comp. Immunol. 2021, 122, 104138. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Ding, S.C.; Vela, A.; Kohlway, A.; Lindenbach, B.D.; Pyle, A.M. Structural insights into RNA recognition by RIG-I. Cell 2011, 147, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Linder, P. Dead-box proteins: A family affair—Active and passive players in RNP-remodeling. Nucleic Acids Res. 2006, 34, 4168–4180. [Google Scholar] [CrossRef] [PubMed]
- Ullah, R.; Li, J.; Fang, P.; Xiao, S.; Fang, L. DEAD/H-box helicases:Anti-viral and pro-viral roles during infections. Virus Res. 2022, 309, 198658. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, B.; Goujon, C. DExH/D-box helicases at the frontline of intrinsic and innate immunity against viral infections. J. Gen. Virol. 2022, 103, 001766. [Google Scholar] [CrossRef]
- Ali, M.A.M. DEAD-box RNA helicases: The driving forces behind RNA metabolism at the crossroad of viral replication and antiviral innate immunity. Virus Res. 2021, 296, 198352. [Google Scholar] [CrossRef]
- Taschuk, F.; Cherry, S. DEAD-Box Helicases: Sensors, Regulators, and Effectors for Antiviral Defense. Viruses 2020, 12, 181. [Google Scholar] [CrossRef] [Green Version]
- Taschuk, F.; Tapescu, I.; Moy, R.H.; Cherry, S. DDX56 Binds to Chikungunya Virus RNA To Control Infection. mBio 2020, 11, 10–1128. [Google Scholar] [CrossRef]
- Göertz, G.P.; van Bree, J.W.M.; Hiralal, A.; Fernhout, B.M.; Steffens, C.; Boeren, S.; Visser, T.M.; Vogels, C.B.F.; Abbo, S.R.; Fros, J.J.; et al. Subgenomic flavivirus RNA binds the mosquito DEAD/H-box helicase ME31B and determines Zika virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2019, 116, 19136–19144. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, K.C.; McLane, L.M.; Maqbool, T.; Panda, D.; Gordesky-Gold, B.; Cherry, S. A genome-wide RNAi screen reveals that mRNA decapping restricts bunyaviral replication by limiting the pools of Dcp2-accessible targets for cap-snatching. Genes Dev. 2013, 27, 1511–1525. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Cheng, M.; Liu, B.; Yuan, M.; Chen, D.; Wang, Y.; Wu, Z. DEAD-Box Helicase DDX6 Facilitated RIG-I-Mediated Type-I Interferon Response to EV71 Infection. Front. Cell Infect Microbiol. 2021, 11, 725392. [Google Scholar] [CrossRef] [PubMed]
- Núñez, R.D.; Budt, M.; Saenger, S.; Paki, K.; Arnold, U.; Sadewasser, A.; Wolff, T. The RNA Helicase DDX6 Associates with RIG-I to Augment Induction of Antiviral Signaling. Int. J. Mol. Sci. 2018, 19, 1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moy, R.H.; Cole, B.S.; Yasunaga, A.; Gold, B.; Shankarling, G.; Varble, A.; Molleston, J.M.; tenOever, B.R.; Lynch, K.W.; Cherry, S. Stem-loop recognition by DDX17 facilitates miRNA processing and antiviral defense. Cell 2014, 158, 764–777. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Srivastava, P.; Mathur, K.; Shrinet, J.; Dubey, S.K.; Chinnappan, M.; Kaur, I.; Nayak, D.; Chattopadhyay, S.; Bhatnagar, R.K.; et al. Chikungunya virus non-structural protein nsP3 interacts with Aedes aegypti DEAD-box helicase RM62F. Virusdisease 2021, 32, 657–665. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, G.; Lv, F.; Wang, X.; Ji, X.; Xu, Y.; Sun, J.; Wu, L.; Zheng, Y.T.; Gao, G. Zinc-finger antiviral protein inhibits HIV-1 infection by selectively targeting multiply spliced viral mRNAs for degradation. Proc. Natl. Acad. Sci. USA 2011, 108, 15834–15839. [Google Scholar] [CrossRef] [PubMed]
- Rosendo Machado, S.; Qu, J.; Koopman, W.J.H.; Miesen, P. The DEAD-box RNA helicase Dhx15 controls glycolysis and arbovirus replication in Aedes aegypti mosquito cells. PLoS Pathog. 2022, 18, e1010694. [Google Scholar] [CrossRef]
- Varjak, M.; Dietrich, I.; Sreenu, V.B.; Till, B.E.; Merits, A.; Kohl, A.; Schnettler, E. Spindle-E Acts Antivirally Against Alphaviruses in Mosquito Cells. Viruses 2018, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Lasko, P. Tudor domain. Curr. Biol. 2010, 20, R666–R667. [Google Scholar] [CrossRef] [Green Version]
- Myllymäki, H.; Valanne, S.; Rämet, M. The Drosophila imd signaling pathway. J. Immunol. 2014, 192, 3455–3462. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary dynamics of immune-related genes and pathways in disease-vector mosquitoes. Science 2007, 316, 1738–1743. [Google Scholar] [CrossRef] [Green Version]
- Antonova, Y.; Alvarez, K.S.; Kim, Y.J.; Kokoza, V.; Raikhel, A.S. The role of NF-kappaB factor REL2 in the Aedes aegypti immune response. Insect Biochem. Mol. Biol. 2009, 39, 303–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Yano, T.; Aggarwal, K.; Lim, J.H.; Ueda, K.; Oshima, Y.; Peach, C.; Erturk-Hasdemir, D.; Goldman, W.E.; Oh, B.H.; et al. PGRP-LC and PGRP-LE have essential yet distinct functions in the drosophila immune response to monomeric DAP-type peptidoglycan. Nat. Immunol. 2006, 7, 715–723. [Google Scholar] [CrossRef]
- Choe, K.M.; Werner, T.; Stöven, S.; Hultmark, D.; Anderson, K.V. Requirement for a peptidoglycan recognition protein (PGRP) in Relish activation and antibacterial immune responses in Drosophila. Science 2002, 296, 359–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottar, M.; Gobert, V.; Michel, T.; Belvin, M.; Duyk, G.; Hoffmann, J.A.; Ferrandon, D.; Royet, J. The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 2002, 416, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Meister, S.; Kanzok, S.M.; Zheng, X.L.; Luna, C.; Li, T.R.; Hoa, N.T.; Clayton, J.R.; White, K.P.; Kafatos, F.C.; Christophides, G.K.; et al. Immune signaling pathways regulating bacterial and malaria parasite infection of the mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2005, 102, 11420–11425. [Google Scholar] [CrossRef]
- Gendrin, M.; Turlure, F.; Rodgers, F.H.; Cohuet, A.; Morlais, I.; Christophides, G.K. The Peptidoglycan Recognition Proteins PGRPLA and PGRPLB Regulate Anopheles Immunity to Bacteria and Affect Infection by Plasmodium. J. Innate Immun. 2017, 9, 333–342. [Google Scholar] [CrossRef]
- Magalhaes, T.; Leandro, D.C.; Ayres, C.F. Knock-down of REL2, but not defensin A, augments Aedes aegypti susceptibility to Bacillus subtilis and Escherichia coli. Acta Trop. 2010, 113, 167–173. [Google Scholar] [CrossRef]
- Costa, A.; Jan, E.; Sarnow, P.; Schneider, D. The Imd pathway is involved in antiviral immune responses in Drosophila. PLoS ONE 2009, 4, e7436. [Google Scholar] [CrossRef] [Green Version]
- Barletta, A.B.; Nascimento-Silva, M.C.; Talyuli, O.A.; Oliveira, J.H.; Pereira, L.O.; Oliveira, P.L.; Sorgine, M.H. Microbiota activates IMD pathway and limits Sindbis infection in Aedes aegypti. Parasit Vectors 2017, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Sansone, C.L.; Cohen, J.; Yasunaga, A.; Xu, J.; Osborn, G.; Subramanian, H.; Gold, B.; Buchon, N.; Cherry, S. Microbiota-Dependent Priming of Antiviral Intestinal Immunity in Drosophila. Cell Host Microbe 2015, 18, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gordesky-Gold, B.; Leney-Greene, M.; Weinbren, N.L.; Tudor, M.; Cherry, S. Inflammation-Induced, STING-Dependent Autophagy Restricts Zika Virus Infection in the Drosophila Brain. Cell Host Microbe 2018, 24, 57–68.e53. [Google Scholar] [CrossRef] [Green Version]
- Avadhanula, V.; Weasner, B.P.; Hardy, G.G.; Kumar, J.P.; Hardy, R.W. A novel system for the launch of alphavirus RNA synthesis reveals a role for the Imd pathway in arthropod antiviral response. PLoS Pathog. 2009, 5, e1000582. [Google Scholar] [CrossRef] [Green Version]
- Laureti, M.; Lee, R.X.; Bennett, A.; Wilson, L.A.; Sy, V.E.; Kohl, A.; Dietrich, I. Rift Valley Fever Virus Primes Immune Responses in Aedes aegypti Cells. Pathogens 2023, 12, 563. [Google Scholar] [CrossRef]
- Sartorius, R.; Trovato, M.; Manco, R.; D’Apice, L.; De Berardinis, P. Exploiting viral sensing mediated by Toll-like receptors to design innovative vaccines. NPJ Vaccines 2021, 6, 127. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Botos, I.; Wang, Y.; Leonard, J.N.; Shiloach, J.; Segal, D.M.; Davies, D.R. Structural basis of toll-like receptor 3 signaling with double-stranded RNA. Science 2008, 320, 379–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lester, S.N.; Li, K. Toll-like receptors in antiviral innate immunity. J. Mol. Biol. 2014, 426, 1246–1264. [Google Scholar] [CrossRef]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Yip, T.F.; Yan, S.; Jin, D.Y.; Wei, H.L.; Guo, R.T.; Peiris, J.S.M. Recognition of Double-Stranded RNA and Regulation of Interferon Pathway by Toll-Like Receptor 10. Front. Immunol. 2018, 9, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carty, M.; Guy, C.; Bowie, A.G. Detection of Viral Infections by Innate Immunity. Biochem. Pharmacol. 2021, 183, 114316. [Google Scholar] [CrossRef]
- Valanne, S.; Wang, J.H.; Ramet, M. The Drosophila Toll signaling pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Gobert, V.; Gottar, M.; Matskevich, A.A.; Rutschmann, S.; Royet, J.; Belvin, M.; Hoffmann, J.A.; Ferrandon, D. Dual activation of the Drosophila toll pathway by two pattern recognition receptors. Science 2003, 302, 2126–2130. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, S.A.; Wasserman, S.A. Conventional and non-conventional Drosophila Toll signaling. Dev. Comp. Immunol. 2014, 42, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Rosendo Machado, S.; van der Most, T.; Miesen, P. Genetic determinants of antiviral immunity in dipteran insects—Compiling the experimental evidence. Dev. Comp. Immunol. 2021, 119, 104010. [Google Scholar] [CrossRef]
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.M.; Hoffmann, J.A. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996, 86, 973–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Chamy, L.; Leclerc, V.; Caldelari, I.; Reichhart, J.M. Sensing of ‘danger signals’ and pathogen-associated molecular patterns defines binary signaling pathways ‘upstream’ of Toll. Nat. Immunol. 2008, 9, 1165–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Weber, A.N.; Atilano, M.L.; Filipe, S.R.; Gay, N.J.; Ligoxygakis, P. Sensing of Gram-positive bacteria in Drosophila: GNBP1 is needed to process and present peptidoglycan to PGRP-SA. Embo J. 2006, 25, 5005–5014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saucereau, Y.; Wilson, T.H.; Tang, M.C.K.; Moncrieffe, M.C.; Hardwick, S.W.; Chirgadze, D.Y.; Soares, S.G.; Marcaida, M.J.; Gay, N.J.; Gangloff, M. Structure and dynamics of Toll immunoreceptor activation in the mosquito Aedes aegypti. Nat. Commun. 2022, 13, 5110. [Google Scholar] [CrossRef]
- Chowdhury, M.; Li, C.F.; He, Z.; Lu, Y.; Liu, X.S.; Wang, Y.F.; Ip, Y.T.; Strand, M.R.; Yu, X.Q. Toll family members bind multiple Spätzle proteins and activate antimicrobial peptide gene expression in Drosophila. J. Biol. Chem. 2019, 294, 10172–10181. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Á, G.; Naylor, H.; Esteves, S.S.; Pais, I.S.; Martins, N.E.; Teixeira, L. The Toll-dorsal pathway is required for resistance to viral oral infection in Drosophila. PLoS Pathog. 2014, 10, e1004507. [Google Scholar] [CrossRef] [Green Version]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The Toll pathway is important for an antiviral response in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Dimopoulos, G. The Toll immune signaling pathway control conserved anti-dengue defenses across diverse Ae. aegypti strains and against multiple dengue virus serotypes. Dev. Comp. Immunol. 2010, 34, 625–629. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef]
- Waldock, J.; Olson, K.E.; Christophides, G.K. Anopheles gambiae antiviral immune response to systemic O’nyong-nyong infection. PLoS Neglected Trop. Dis. 2012, 6, e1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamoto, M.; Moy, R.H.; Xu, J.; Bambina, S.; Yasunaga, A.; Shelly, S.S.; Gold, B.; Cherry, S. Virus recognition by Toll-7 activates antiviral autophagy in Drosophila. Immunity 2012, 36, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.J.; Lu, G.; Qi, Y.H.; Zhang, Y.; Zhang, X.D.; Huang, H.J.; Zhuo, J.C.; Sun, Z.T.; Yan, F.; Chen, J.P.; et al. Activation of Toll Immune Pathway in an Insect Vector Induced by a Plant Virus. Front. Immunol. 2020, 11, 613957. [Google Scholar] [CrossRef] [PubMed]
- Moy, R.H.; Gold, B.; Molleston, J.M.; Schad, V.; Yanger, K.; Salzano, M.V.; Yagi, Y.; Fitzgerald, K.A.; Stanger, B.Z.; Soldan, S.S.; et al. Antiviral autophagy restrictsRift Valley fever virus infection and is conserved from flies to mammals. Immunity 2014, 40, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Lamiable, O.; Arnold, J.; de Faria, I.; Olmo, R.P.; Bergami, F.; Meignin, C.; Hoffmann, J.A.; Marques, J.T.; Imler, J.L. Analysis of the Contribution of Hemocytes and Autophagy to Drosophila Antiviral Immunity. J. Virol. 2016, 90, 5415–5426. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef]
- Bartholomay, L.C.; Waterhouse, R.M.; Mayhew, G.F.; Campbell, C.L.; Michel, K.; Zou, Z.; Ramirez, J.L.; Das, S.; Alvarez, K.; Arensburger, P.; et al. Pathogenomics of Culex quinquefasciatus and meta-analysis of infection responses to diverse pathogens. Science 2010, 330, 88–90. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.F.; Torres, A.Q.; Jardim, R.; Mesquita, R.D.; Schama, R. Evolution of Toll, Spatzle and MyD88 in insects: The problem of the Diptera bias. BMC Genom. 2021, 22, 562. [Google Scholar] [CrossRef]
- Zhang, R.; Li, X.; Zhang, J.; Li, Y.; Wang, Y.; Song, Y.; Ren, F.; Yi, H.; Deng, X.; Zhong, Y.; et al. Toll9 from Bombyx mori functions as a pattern recognition receptor that shares features with Toll-like receptor 4 from mammals. Proc. Natl. Acad. Sci. USA 2021, 118, e2103021118. [Google Scholar] [CrossRef] [PubMed]
- Cheung, Y.P.; Park, S.; Pagtalunan, J.; Maringer, K. The antiviral role of NF-κB-mediated immune responses and their antagonism by viruses in insects. J. Gen. Virol. 2022, 103, 001741. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B.; Eidenschenk, C.; Crozat, K.; Imler, J.L.; Takeuchi, O.; Hoffmann, J.A.; Akira, S. Genetic analysis of resistance to viral infection. Nat. Rev. Immunol. 2007, 7, 753–766. [Google Scholar] [CrossRef]
- Harrison, D.A.; McCoon, P.E.; Binari, R.; Gilman, M.; Perrimon, N. Drosophila unpaired encodes a secreted protein that activates the JAK signaling pathway. Genes. Dev. 1998, 12, 3252–3263. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Zhao, B.R.; Zhang, H.; You, Y.L.; Li, F.; Wang, X.W. Interferon functional analog activates antiviral Jak/Stat signaling through integrin in an arthropod. Cell Rep. 2021, 36, 109761. [Google Scholar] [CrossRef]
- Chang, L.; Karin, M. Mammalian MAP kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef]
- Horton, A.A.; Wang, B.; Camp, L.; Price, M.S.; Arshi, A.; Nagy, M.; Nadler, S.A.; Faeder, J.R.; Luckhart, S. The mitogen-activated protein kinome from Anopheles gambiae: Identification, phylogeny and functional characterization of the ERK, JNK and p38 MAP kinases. BMC Genom. 2011, 12, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.Q.; Chen, S.Q.; Bai, H.Q.; Wei, Q.M.; Zhang, S.N.; Chen, C.; Zhu, Y.H.; Yi, T.W.; Guo, X.P.; Chen, S.Y.; et al. The Ras/ERK signaling pathway couples antimicrobial peptides to mediate resistance to dengue virus in Aedes mosquitoes. PLoS Negl. Trop. Dis. 2020, 14, e0008660. [Google Scholar] [CrossRef] [PubMed]
- Ahlers, L.R.H.; Trammell, C.E.; Carrell, G.F.; Mackinnon, S.; Torrevillas, B.K.; Chow, C.Y.; Luckhart, S.; Goodman, A.G. Insulin Potentiates JAK/STAT Signaling to Broadly Inhibit Flavivirus Replication in Insect Vectors. Cell Rep. 2019, 29, 1946–1960.e5. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Hopkins, K.; Sabin, L.; Yasunaga, A.; Subramanian, H.; Lamborn, I.; Gordesky-Gold, B.; Cherry, S. ERK signaling couples nutrient status to antiviral defense in the insect gut. Proc. Natl. Acad. Sci. USA 2013, 110, 15025–15030. [Google Scholar] [CrossRef]
- Chowdhury, A.; Modahl, C.M.; Tan, S.T.; Wong Wei Xiang, B.; Missé, D.; Vial, T.; Kini, R.M.; Pompon, J.F. JNK pathway restricts DENV2, ZIKV and CHIKV infection by activating complement and apoptosis in mosquito salivary glands. PLoS Pathog. 2020, 16, e1008754. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, A.E.; Walker, A.J.; Goodbourn, S. Regulation of transcription of the Aedes albopictus cecropin A1 gene: A role for p38 mitogen-activated protein kinase. Insect Biochem. Mol. Biol. 2011, 41, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Ricklin, D.; Reis, E.S.; Mastellos, D.C.; Gros, P.; Lambris, J.D. Complement component C3—The “Swiss Army Knife” of innate immunity and host defense. Immunol. Rev. 2016, 274, 33–58. [Google Scholar] [CrossRef] [Green Version]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.; Baxter, R. The structure and function of thioester-containing proteins in arthropods. Biophys. Rev. 2014, 6, 261–272. [Google Scholar] [CrossRef]
- Blandin, S.; Levashina, E.A. Thioester-containing proteins and insect immunity. Mol. Immunol. 2004, 40, 903–908. [Google Scholar] [CrossRef]
- Rehman, A.A.; Ahsan, H.; Khan, F.H. α-2-Macroglobulin: A physiological guardian. J. Cell Physiol. 2013, 228, 1665–1675. [Google Scholar] [CrossRef]
- Lagueux, M.; Perrodou, E.; Levashina, E.A.; Capovilla, M.; Hoffmann, J.A. Constitutive expression of a complement-like protein in toll and JAK gain-of-function mutants of Drosophila. Proc. Natl. Acad. Sci. USA 2000, 97, 11427–11432. [Google Scholar] [CrossRef]
- Dostalova, A.; Rommelaere, S.; Poidevin, M.; Lemaitre, B. Thioester-containing proteins regulate the Toll pathway and play a role in Drosophila defence against microbial pathogens and parasitoid wasps. BMC Biol. 2017, 15, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, R.H.; Chang, C.I.; Chelliah, Y.; Blandin, S.; Levashina, E.A.; Deisenhofer, J. Structural basis for conserved complement factor-like function in the antimalarial protein TEP1. Proc. Natl. Acad. Sci. USA 2007, 104, 11615–11620. [Google Scholar] [CrossRef] [PubMed]
- Blandin, S.; Shiao, S.H.; Moita, L.F.; Janse, C.J.; Waters, A.P.; Kafatos, F.C.; Levashina, E.A. Complement-like protein TEP1 is a determinant of vectorial capacity in the malaria vector Anopheles gambiae. Cell 2004, 116, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Levashina, E.A.; Moita, L.F.; Blandin, S.; Vriend, G.; Lagueux, M.; Kafatos, F.C. Conserved role of a complement-like protein in phagocytosis revealed by dsRNA knockout in cultured cells of the mosquito, Anopheles gambiae. Cell 2001, 104, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Liu, L.; Wang, P.; Zhang, Y.; Zhao, Y.O.; Colpitts, T.M.; Feitosa, F.; Anderson, J.F.; Fikrig, E. An in vivo transfection approach elucidates a role for Aedes aegypti thioester-containing proteins in flaviviral infection. PLoS ONE 2011, 6, e22786. [Google Scholar] [CrossRef] [Green Version]
- Weng, S.C.; Li, H.H.; Li, J.C.; Liu, W.L.; Chen, C.H.; Shiao, S.H. A Thioester-Containing Protein Controls Dengue Virus Infection in Aedes aegypti Through Modulating Immune Response. Front. Immunol. 2021, 12, 670122. [Google Scholar] [CrossRef]
- Christophides, G.K.; Zdobnov, E.; Barillas-Mury, C.; Birney, E.; Blandin, S.; Blass, C.; Brey, P.T.; Collins, F.H.; Danielli, A.; Dimopoulos, G.; et al. Immunity-related genes and gene families in Anopheles gambiae. Science 2002, 298, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Nürnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: Striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef]
- Pancer, Z.; Amemiya, C.T.; Ehrhardt, G.R.; Ceitlin, J.; Gartland, G.L.; Cooper, M.D. Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nature 2004, 430, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Ng, A.; Xavier, R.J. Leucine-rich repeat (LRR) proteins: Integrators of pattern recognition and signaling in immunity. Autophagy 2011, 7, 1082–1084. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N. Tick-Virus Interactions: Toll Sensing. Front. Cell Infect Microbiol. 2017, 7, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botos, I.; Segal, D.M.; Davies, D.R. The structural biology of Toll-like receptors. Structure 2011, 19, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imler, J.L.; Zheng, L. Biology of Toll receptors: Lessons from insects and mammals. J. Leukoc. Biol. 2004, 75, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.K.; Mullen, G.E.; Leifer, C.A.; Mazzoni, A.; Davies, D.R.; Segal, D.M. Leucine-rich repeats and pathogen recognition in Toll-like receptors. Trends Immunol. 2003, 24, 528–533. [Google Scholar] [CrossRef]
- Fraiture, M.; Baxter, R.H.; Steinert, S.; Chelliah, Y.; Frolet, C.; Quispe-Tintaya, W.; Hoffmann, J.A.; Blandin, S.A.; Levashina, E.A. Two mosquito LRR proteins function as complement control factors in the TEP1-mediated killing of Plasmodium. Cell Host Microbe 2009, 5, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Povelones, M.; Waterhouse, R.M.; Kafatos, F.C.; Christophides, G.K. Leucine-rich repeat protein complex activates mosquito complement in defense against Plasmodium parasites. Science 2009, 324, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, R.M.; Povelones, M.; Christophides, G.K. Sequence-structure-function relations of the mosquito leucine-rich repeat immune proteins. BMC Genom. 2010, 11, 531. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Alto, B.W.; Shin, D. Transcriptional Profile of Aedes aegypti Leucine-Rich Repeat Proteins in Response to Zika and Chikungunya Viruses. Int. J. Mol. Sci. 2019, 20, 615. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.; Jupatanakul, N.; Ramirez, J.L.; Kang, S.; Romero-Vivas, C.M.; Mohammed, H.; Dimopoulos, G. Transcriptomic profiling of diverse Aedes aegypti strains reveals increased basal-level immune activation in dengue virus-refractory populations and identifies novel virus-vector molecular interactions. PLoS Negl. Trop. Dis. 2013, 7, e2295. [Google Scholar] [CrossRef] [Green Version]
- Stroschein-Stevenson, S.L.; Foley, E.; O’Farrell, P.H.; Johnson, A.D. Identification of Drosophila gene products required for phagocytosis of Candida albicans. PLoS Biol. 2006, 4, e4. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Liu, Y.; Zhang, X.; Wang, J.; Li, Z.; Pang, X.; Wang, P.; Cheng, G. Complement-related proteins control the flavivirus infection of Aedes aegypti by inducing antimicrobial peptides. PLoS Pathog. 2014, 10, e1004027. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Morton, J.C., Jr.; Ramirez, J.L.; Souza-Neto, J.A.; Dimopoulos, G. The entomopathogenic fungus Beauveria bassiana activate toll and JAK-STAT pathway-controlled effector genes and anti-dengue activity in Aedes aegypti. Insect Biochem. Mol. Biol. 2012, 42, 126–132. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, J.L.; Ho, Y.K.; Basu, S.K.; Brown, M.S. Binding site on macrophages that mediates uptake and degradation of acetylated low density lipoprotein, producing massive cholesterol deposition. Proc. Natl. Acad. Sci. USA 1979, 76, 333–337. [Google Scholar] [CrossRef]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Taban, Q.; Mumtaz, P.T.; Masoodi, K.Z.; Haq, E.; Ahmad, S.M. Scavenger receptors in host defense: From functional aspects to mode of action. Cell Commun. Signal 2022, 20, 2. [Google Scholar] [CrossRef]
- Zani, I.A.; Stephen, S.L.; Mughal, N.A.; Russell, D.; Homer-Vanniasinkam, S.; Wheatcroft, S.B.; Ponnambalam, S. Scavenger receptor structure and function in health and disease. Cells 2015, 4, 178–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plüddemann, A.; Neyen, C.; Gordon, S. Macrophage scavenger receptors and host-derived ligands. Methods 2007, 43, 207–217. [Google Scholar] [CrossRef]
- Pearson, A.; Lux, A.; Krieger, M. Expression cloning of dSR-CI, a class C macrophage-specific scavenger receptor from Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1995, 92, 4056–4060. [Google Scholar] [CrossRef] [PubMed]
- Rämet, M.; Pearson, A.; Manfruelli, P.; Li, X.; Koziel, H.; Göbel, V.; Chung, E.; Krieger, M.; Ezekowitz, R.A. Drosophila scavenger receptor CI is a pattern recognition receptor for bacteria. Immunity 2001, 15, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.C.; Shi, X.Z.; Yang, H.T.; Sun, J.J.; Xu, L.; Wang, X.W.; Zhao, X.F.; Wang, J.X. Scavenger Receptor C Mediates Phagocytosis of White Spot Syndrome Virus and Restricts Virus Proliferation in Shrimp. PLoS Pathog. 2016, 12, e1006127. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Geng, T.; Yang, G.; Ma, J.; Wang, L.; Ketkar, H.; Yang, D.; Lin, T.; Hwang, J.; Zhu, S.; et al. Macrophage scavenger receptor 1 controls Chikungunya virus infection through autophagy in mice. Commun. Biol. 2020, 3, 556. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, K.S.; Davenport, B.J.; Haist, K.C.; McCarthy, M.K.; May, N.A.; Robison, A.; Ruckert, C.; Ebel, G.D.; Morrison, T.E. Discrete viral E2 lysine residues and scavenger receptor MARCO are required for clearance of circulating alphaviruses. Elife 2019, 8, e49163. [Google Scholar] [CrossRef] [PubMed]
- Nichols, Z.; Vogt, R.G. The SNMP/CD36 gene family in Diptera, Hymenoptera and Coleoptera: Drosophila melanogaster, D. pseudoobscura, Anopheles gambiae, Aedes aegypti, Apis mellifera, and Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 398–415. [Google Scholar] [CrossRef] [Green Version]
- Alcalá, A.C.; Maravillas, J.L.; Meza, D.; Ramirez, O.T.; Ludert, J.E.; Palomares, L.A. Dengue Virus NS1 Uses Scavenger Receptor B1 as a Cell Receptor in Cultured Cells. J. Virol. 2022, 96, e0166421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Shen, L.; Wang, X.; Yang, H.; Zhang, X.; Pan, G.; Li, C.; Ji, H.; Abbas, M.N.; Cui, H. Scavenger receptor C regulates antimicrobial peptide expression by activating toll signaling in silkworm, Bombyx mori. Int. J. Biol. Macromol. 2021, 191, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhao, T.; Ai, K.; Li, H.; Jiang, X.; Li, C.; Wang, Q.; Yang, J.; Zhang, R. Role of scavenger receptor from Octopus ocellatus as a co-receptor of Toll-like receptor in initiation of TLR-NF-κB signaling during anti-bacterial response. Dev. Comp. Immunol. 2018, 84, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ha, T.; Liu, L.; Wang, X.; Gao, M.; Kelley, J.; Kao, R.; Williams, D.; Li, C. Scavenger receptor A (SR-A) is required for LPS-induced TLR4 mediated NF-κB activation in macrophages. Biochim. Biophys. Acta 2012, 1823, 1192–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiel, E.; Alonso, A.; Uematsu, S.; Akira, S.; Poynter, M.E.; Berwin, B. Pivotal Advance: Toll-like receptor regulation of scavenger receptor-A-mediated phagocytosis. J. Leukoc. Biol. 2009, 85, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Limmon, G.V.; Arredouani, M.; McCann, K.L.; Corn Minor, R.A.; Kobzik, L.; Imani, F. Scavenger receptor class-A is a novel cell surface receptor for double-stranded RNA. FASEB J. 2008, 22, 159–167. [Google Scholar] [CrossRef]
- DeWitte-Orr, S.J.; Collins, S.E.; Bauer, C.M.; Bowdish, D.M.; Mossman, K.L. An accessory to the ‘Trinity’: SR-As are essential pathogen sensors of extracellular dsRNA, mediating entry and leading to subsequent type I IFN responses. PLoS Pathog. 2010, 6, e1000829. [Google Scholar] [CrossRef] [Green Version]
- Dansako, H.; Yamane, D.; Welsch, C.; McGivern, D.R.; Hu, F.; Kato, N.; Lemon, S.M. Class A scavenger receptor 1 (MSR1) restricts hepatitis C virus replication by mediating toll-like receptor 3 recognition of viral RNAs produced in neighboring cells. PLoS Pathog. 2013, 9, e1003345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-type lectin superfamily in the immune system. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Pees, B.; Yang, W.; Zárate-Potes, A.; Schulenburg, H.; Dierking, K. High Innate Immune Specificity through Diversified C-Type Lectin-Like Domain Proteins in Invertebrates. J. Innate Immun. 2016, 8, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Hoving, J.C.; Wilson, G.J.; Brown, G.D. Signalling C-type lectin receptors, microbial recognition and immunity. Cell Microbiol. 2014, 16, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, A.; Pinto, A.K.; Schwaeble, W.J.; Diamond, M.S. The lectin pathway of complement activation contributes to protection from West Nile virus infection. Virology 2011, 412, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Avirutnan, P.; Hauhart, R.E.; Marovich, M.A.; Garred, P.; Atkinson, J.P.; Diamond, M.S. Complement-mediated neutralization of dengue virus requires mannose-binding lectin. mBio 2011, 2, 10–1128. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Fang, N.N.; Zheng, Y.; Liu, K.Y.; Mao, B.; Kong, L.N.; Chen, Y.; Ai, H. Identification and characterization of two novel C-type lectins from the larvae of housefly, Musca domestica L. Arch. Insect. Biochem. Physiol. 2018, 98, e21467. [Google Scholar] [CrossRef]
- Shi, X.Z.; Kang, C.J.; Wang, S.J.; Zhong, X.; Beerntsen, B.T.; Yu, X.Q. Functions of Armigeres subalbatus C-type lectins in innate immunity. Insect Biochem. Mol. Biol. 2014, 52, 102–114. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Wang, Y.; Li, F.; Liu, J.; Sun, Y.; Wu, J. Cloning and characterization of a mannose binding C-type lectin gene from salivary gland of Aedes albopictus. Parasit Vectors 2014, 7, 337. [Google Scholar] [CrossRef] [Green Version]
- Adelman, Z.N.; Myles, K.M. The C-Type Lectin Domain Gene Family in Aedes aegypti and Their Role in Arbovirus Infection. Viruses 2018, 10, 367. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Cox, J.; Wang, P.; Krishnan, M.N.; Dai, J.; Qian, F.; Anderson, J.F.; Fikrig, E. A C-type lectin collaborates with a CD45 phosphatase homolog to facilitate West Nile virus infection of mosquitoes. Cell 2010, 142, 714–725. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, M.N.; Ng, A.; Sukumaran, B.; Gilfoy, F.D.; Uchil, P.D.; Sultana, H.; Brass, A.L.; Adametz, R.; Tsui, M.; Qian, F.; et al. RNA interference screen for human genes associated with West Nile virus infection. Nature 2008, 455, 242–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakhshi, H.; Fazlalipour, M.; Dadgar-Pakdel, J.; Zakeri, S.; Raz, A.; Failloux, A.B.; Dinparast Djadid, N. Developing a Vaccine to Block West Nile Virus Transmission: In Silico Studies, Molecular Characterization, Expression, and Blocking Activity of Culex pipiens mosGCTL-1. Pathogens 2021, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, F.; Liu, J.; Xiao, X.; Zhang, S.; Qin, C.; Xiang, Y.; Wang, P.; Cheng, G. Transmission-blocking antibodies against mosquito C-type lectins for dengue prevention. PLoS Pathog. 2014, 10, e1003931. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Qian, Y.; Jung, Y.S.; Zhou, B.; Cao, R.; Shen, T.; Shao, D.; Wei, J.; Ma, Z.; Chen, P.; et al. mosGCTL-7, a C-Type Lectin Protein, Mediates Japanese Encephalitis Virus Infection in Mosquitoes. J. Virol. 2017, 91, e01348-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, B.M.; Li, J.; Chen, C.C.; Nappi, A.J. Melanization immune responses in mosquito vectors. Trends Parasitol. 2005, 21, 192–199. [Google Scholar] [CrossRef]
- Cerenius, L.; Lee, B.L.; Soderhall, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. Immune properties of invertebrate phenoloxidases. Dev. Comp. Immunol. 2021, 122, 104098. [Google Scholar] [CrossRef]
- Cerenius, L.; Soderhall, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, H. Prophenoloxidase activation and antimicrobial peptide expression induced by the recombinant microbe binding protein of Manduca sexta. Insect Biochem. Mol. Biol. 2017, 83, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, D.; Garcia, B.L.; Kanost, M.R. Initiating protease with modular domains interacts with β-glucan recognition protein to trigger innate immune response in insects. Proc. Natl. Acad. Sci. USA 2015, 112, 13856–13861. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, H. Binding properties of the regulatory domains in Manduca sexta hemolymph proteinase-14, an initiation enzyme of the prophenoloxidase activation system. Dev. Comp. Immunol. 2010, 34, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jiang, H. Interaction of beta-1,3-glucan with its recognition protein activates hemolymph proteinase 14, an initiation enzyme of the prophenoloxidase activation system in Manduca sexta. J. Biol. Chem. 2006, 281, 9271–9278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, C.; Wang, Y.; Guo, X.; Hartson, S.; Jiang, H. A pattern recognition serine proteinase triggers the prophenoloxidase activation cascade in the tobacco hornworm, Manduca sexta. J. Biol. Chem. 2004, 279, 34101–34106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, J.; Ling, E.; Yu, X.Q. Drosophila C-type lectins enhance cellular encapsulation. Mol. Immunol. 2007, 44, 2541–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, F.H.; Sakai, R.K.; Vernick, K.D.; Paskewitz, S.; Seeley, D.C.; Miller, L.H.; Collins, W.E.; Campbell, C.C.; Gwadz, R.W. Genetic selection of a Plasmodium-refractory strain of the malaria vector Anopheles gambiae. Science 1986, 234, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fuchs, J.F.; Infanger, L.C.; Rocheleau, T.A.; Hillyer, J.F.; Chen, C.C.; Christensen, B.M. Mosquito innate immunity: Involvement of beta 1,3-glucan recognition protein in melanotic encapsulation immune responses in Armigeres subalbatus. Mol. Biochem. Parasitol. 2005, 139, 65–73. [Google Scholar] [CrossRef]
- Ourth, D.D.; Renis, H.E. Antiviral melanization reaction of Heliothis virescens hemolymph against DNA and RNA viruses in vitro. Comp. Biochem. Physiol. B 1993, 105, 719–723. [Google Scholar] [CrossRef]
- Shelby, K.S.; Popham, H.J. Plasma phenoloxidase of the larval tobacco budworm, Heliothis virescens, is virucidal. J. Insect Sci. 2006, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.H.; Strand, M.R. A novel polydnavirus protein inhibits the insect prophenoloxidase activation pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 19267–19272. [Google Scholar] [CrossRef]
- Rodriguez-Andres, J.; Rani, S.; Varjak, M.; Chase-Topping, M.E.; Beck, M.H.; Ferguson, M.C.; Schnettler, E.; Fragkoudis, R.; Barry, G.; Merits, A.; et al. Phenoloxidase activity acts as a mosquito innate immune response against infection with Semliki Forest virus. PLoS Pathog. 2012, 8, e1002977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligoxygakis, P.; Pelte, N.; Ji, C.; Leclerc, V.; Duvic, B.; Belvin, M.; Jiang, H.; Hoffmann, J.A.; Reichhart, J.M. A serpin mutant links Toll activation to melanization in the host defence of Drosophila. Embo J. 2002, 21, 6330–6337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takehana, A.; Yano, T.; Mita, S.; Kotani, A.; Oshima, Y.; Kurata, S. Peptidoglycan recognition protein (PGRP)-LE and PGRP-LC act synergistically in Drosophila immunity. Embo J. 2004, 23, 4690–4700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.E.; Mavica, J.; Shackleford, L.; Flis, I.; Fochler, S.; Basu, S.; Alphey, L. CRISPR/Cas9 gene editing in the West Nile Virus vector, Culex quinquefasciatus Say. PLoS ONE 2019, 14, e0224857. [Google Scholar] [CrossRef]
- Chaverra-Rodriguez, D.; Macias, V.M.; Hughes, G.L.; Pujhari, S.; Suzuki, Y.; Peterson, D.R.; Kim, D.; McKeand, S.; Rasgon, J.L. Targeted delivery of CRISPR-Cas9 ribonucleoprotein into arthropod ovaries for heritable germline gene editing. Nat. Commun. 2018, 9, 3008. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Li, M.; McMeniman, C.J.; Akbari, O.S. CRISPR-Mediated Genome Engineering in Aedes aegypti. Methods Mol. Biol. 2022, 2509, 23–51. [Google Scholar] [CrossRef]
- Raban, R.R.; Marshall, J.M.; Akbari, O.S. Progress towards engineering gene drives for population control. J. Exp. Biol. 2020, 223, jeb208181. [Google Scholar] [CrossRef] [Green Version]
- Bier, E. Gene drives gaining speed. Nat. Rev. Genet. 2022, 23, 5–22. [Google Scholar] [CrossRef]
- Samuel, G.H.; Pohlenz, T.; Dong, Y.; Coskun, N.; Adelman, Z.N.; Dimopoulos, G.; Myles, K.M. RNA interference is essential to modulating the pathogenesis of mosquito-borne viruses in the yellow fever mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2023, 120, e2213701120. [Google Scholar] [CrossRef]
- Jupatanakul, N.; Sim, S.; Anglero-Rodriguez, Y.I.; Souza-Neto, J.; Das, S.; Poti, K.E.; Rossi, S.L.; Bergren, N.; Vasilakis, N.; Dimopoulos, G. Engineered Aedes aegypti JAK/STAT Pathway-Mediated Immunity to Dengue Virus. PLoS Neglected Trop. Dis. 2017, 11, e0005187. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Mosquito antiviral immune pathways. An overview of the different antiviral defense strategies available to mosquitoes and the potential receptors that act as pattern recognition receptors to initiate them. We focus on receptors for which there is experimental evidence for their putative role in sensing virus infection, including Drosophila melanogaster and mosquitoes. Receptors that are suspected or shown to be responsible for sensing viruses directly and initiating an antiviral response are colored in red. Protein receptors that may facilitate viral entry are colored in blue. Question marks denote strong candidate receptors or signaling pathways that so far lack any direct evidence for the receptor function in mosquitoes. Distinct pathways are in bold/underlined. Dotted arrows represent canonical pathway crosstalk. Created using BioRender.com (accessed on 16 June 2023).
Figure 1.
Mosquito antiviral immune pathways. An overview of the different antiviral defense strategies available to mosquitoes and the potential receptors that act as pattern recognition receptors to initiate them. We focus on receptors for which there is experimental evidence for their putative role in sensing virus infection, including Drosophila melanogaster and mosquitoes. Receptors that are suspected or shown to be responsible for sensing viruses directly and initiating an antiviral response are colored in red. Protein receptors that may facilitate viral entry are colored in blue. Question marks denote strong candidate receptors or signaling pathways that so far lack any direct evidence for the receptor function in mosquitoes. Distinct pathways are in bold/underlined. Dotted arrows represent canonical pathway crosstalk. Created using BioRender.com (accessed on 16 June 2023).


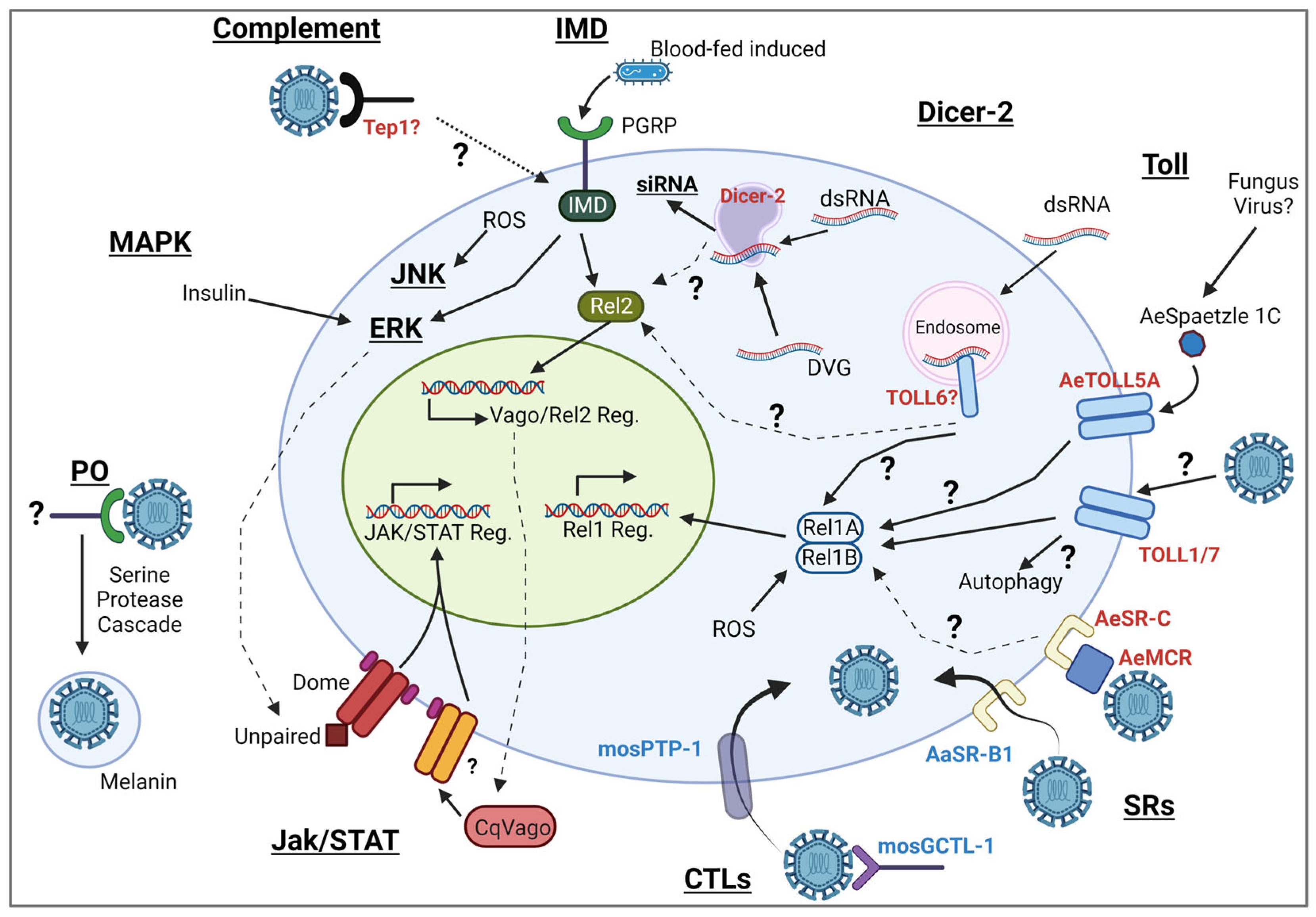{kind=link}
Table 1.
Ae. aegypti DExD/DExH-box helicases with putative PRR or antiviral activity.
| Name/ Accession | Motif II Sequence | Antiviral Activity | Binding Partner | Human Ortholog |
|---|---|---|---|---|
| ME31B (AAEL008500) | DEAD | Yes | sfRNA | DDX63 # |
| Rm62 * (AAEL008738) | DEAD | Unknown | CHIKV nsP3 | DDX17 # |
| DHX15 (AAEL004419) | DEAH | Yes | Unknown | DHX15 # |
| AAEL008728 | DETD | Yes | Unknown | DDX24 |
| AAEL004859 | DEIH | Yes | Unknown | DHX9 # |
| Spindle E (AAEL013235) | DEIH | Yes | Unknown | TDRD9 |
* Antiviral PRR in Drosophila. # Antiviral PRR in mammals.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Prince, B.C.; Walsh, E.; Torres, T.Z.B.; Rückert, C. Recognition of Arboviruses by the Mosquito Immune System. Biomolecules 2023, 13, 1159. https://doi.org/10.3390/biom13071159
AMA Style
Prince BC, Walsh E, Torres TZB, Rückert C. Recognition of Arboviruses by the Mosquito Immune System. Biomolecules. 2023; 13(7):1159. https://doi.org/10.3390/biom13071159
Chicago/Turabian StylePrince, Brian C., Elizabeth Walsh, Tran Zen B. Torres, and Claudia Rückert. 2023. "Recognition of Arboviruses by the Mosquito Immune System" Biomolecules 13, no. 7: 1159. https://doi.org/10.3390/biom13071159
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.