
Preventive Effect of Epigallocatechin Gallate, the Main Component of Green Tea, on Acute Lung Injury Caused by Air Pollutants
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Animals
2.2. Urban Aerosol Particle Suspension
2.3. Treatment of Mice with Urban Aerosols, EGCG and GCG
2.4. Preparation of Bronchoalveolar Lavage Fluid (BALF) and Immunoblotting Analysis
2.5. In Vivo ROS Measurement by Imaging Analysis
2.6. Real-Time Reverse Transcription (RT) Polymerase Chain Reaction (PCR) Analysis
2.7. Cell Culture and In Vitro ROS Measurement
2.8. Statistical Analysis
3. Results
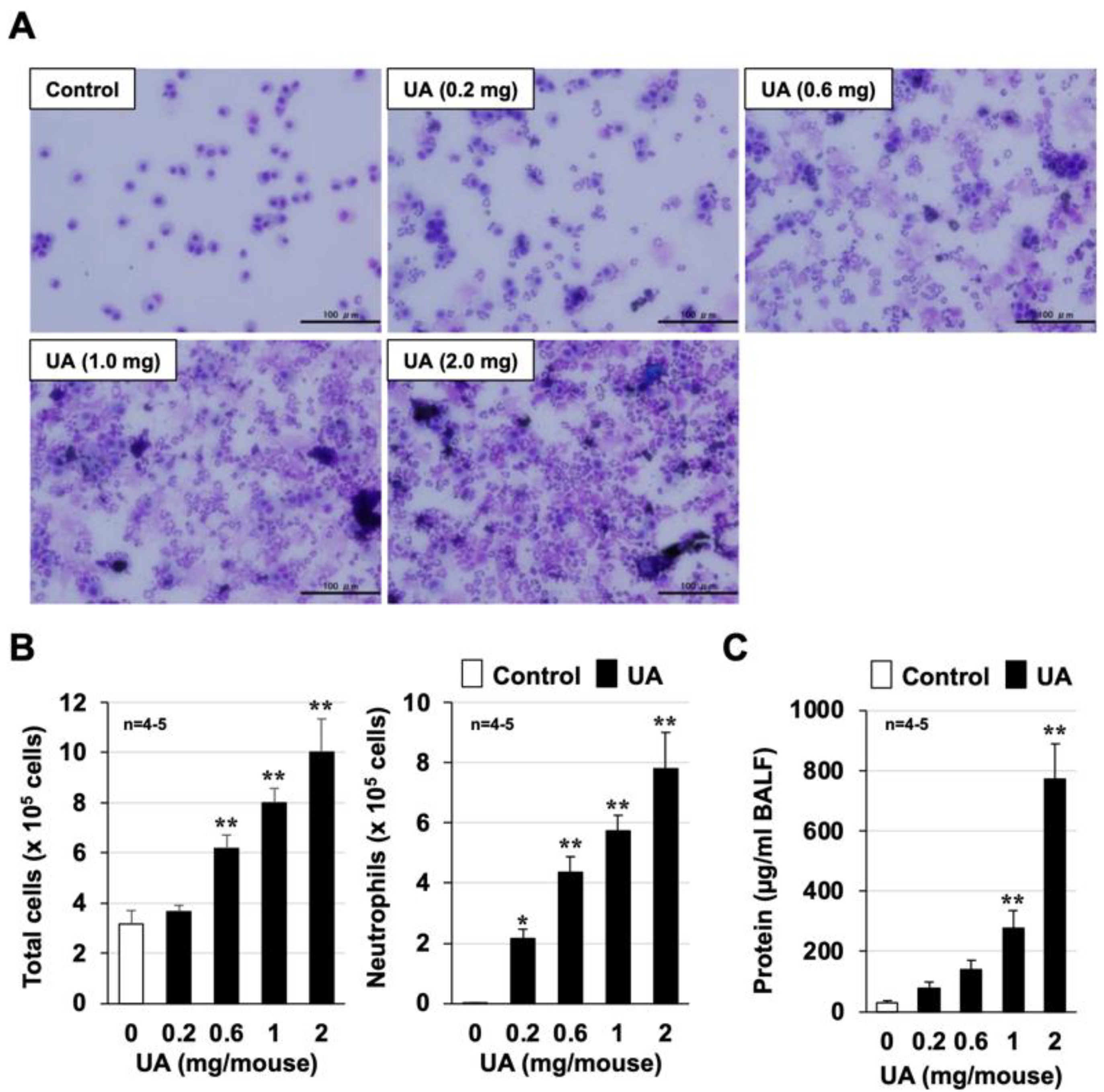
3.1. Mouse Model of Urban Aerosol-Induced Acute Lung Injury
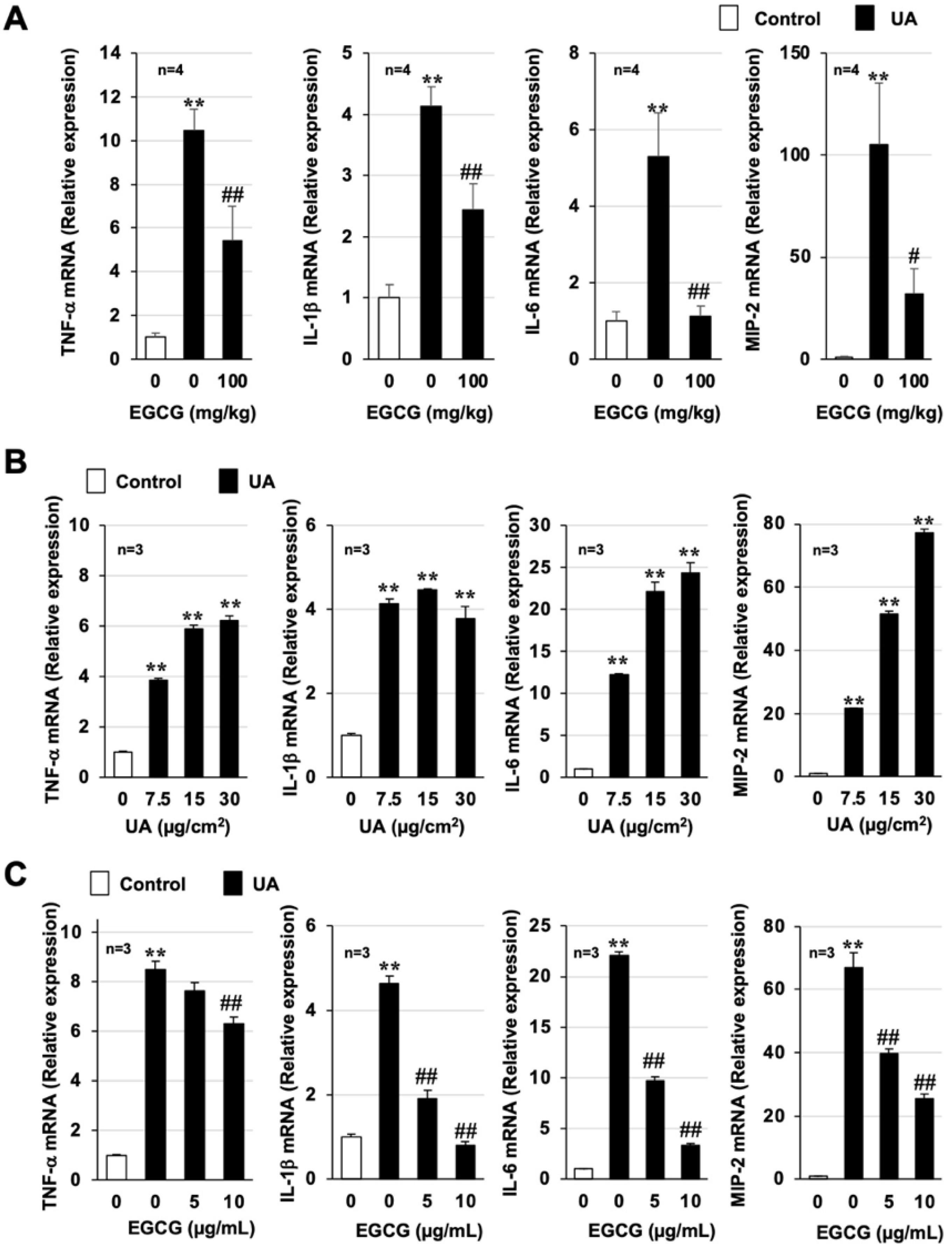
3.2. Effect of EGCG on Urban Aerosol-Induced Acute Lung Injury and Inflammatory Responses
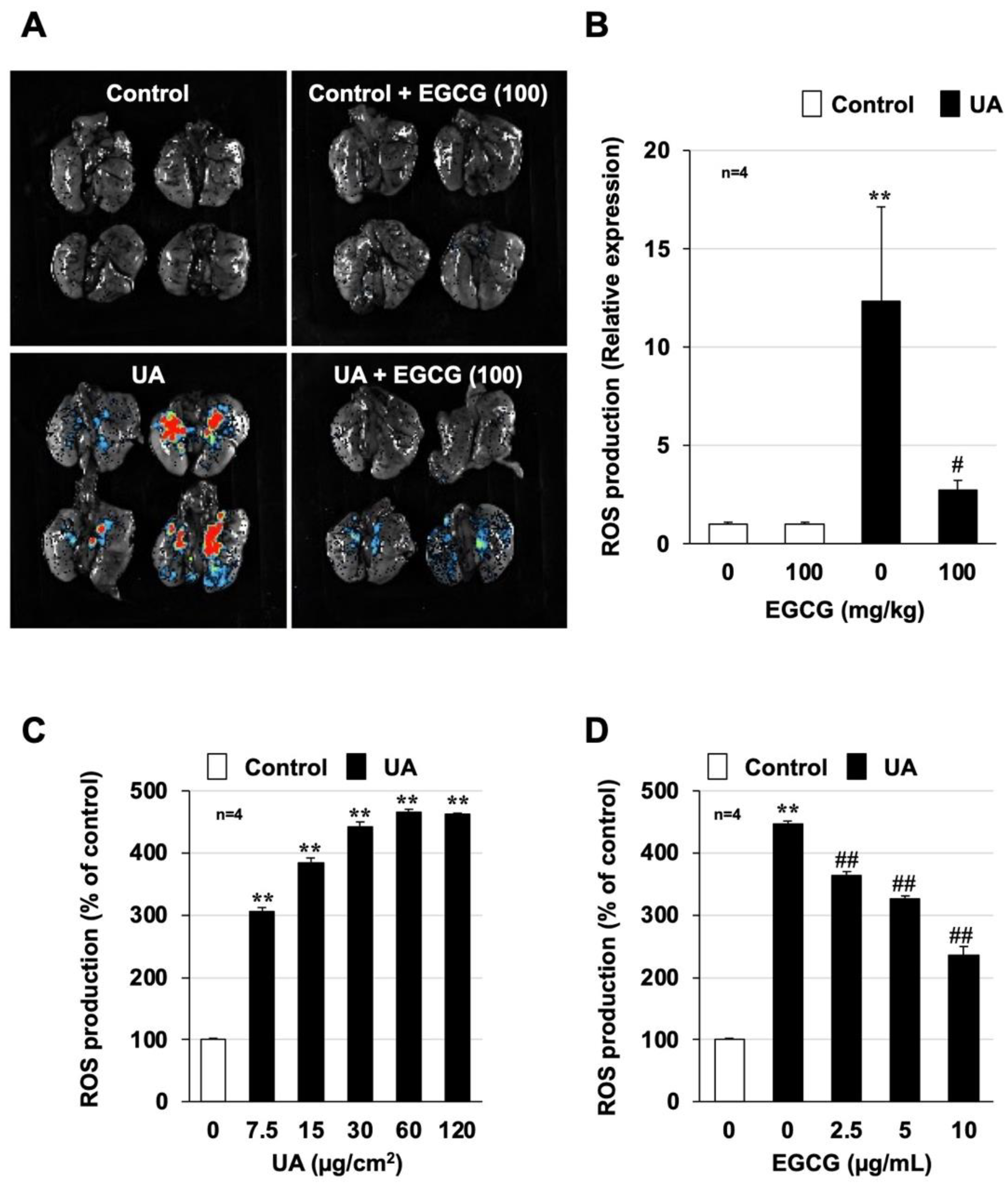
3.3. Effect of EGCG on Urban Aerosol-Induced Oxidative Stress
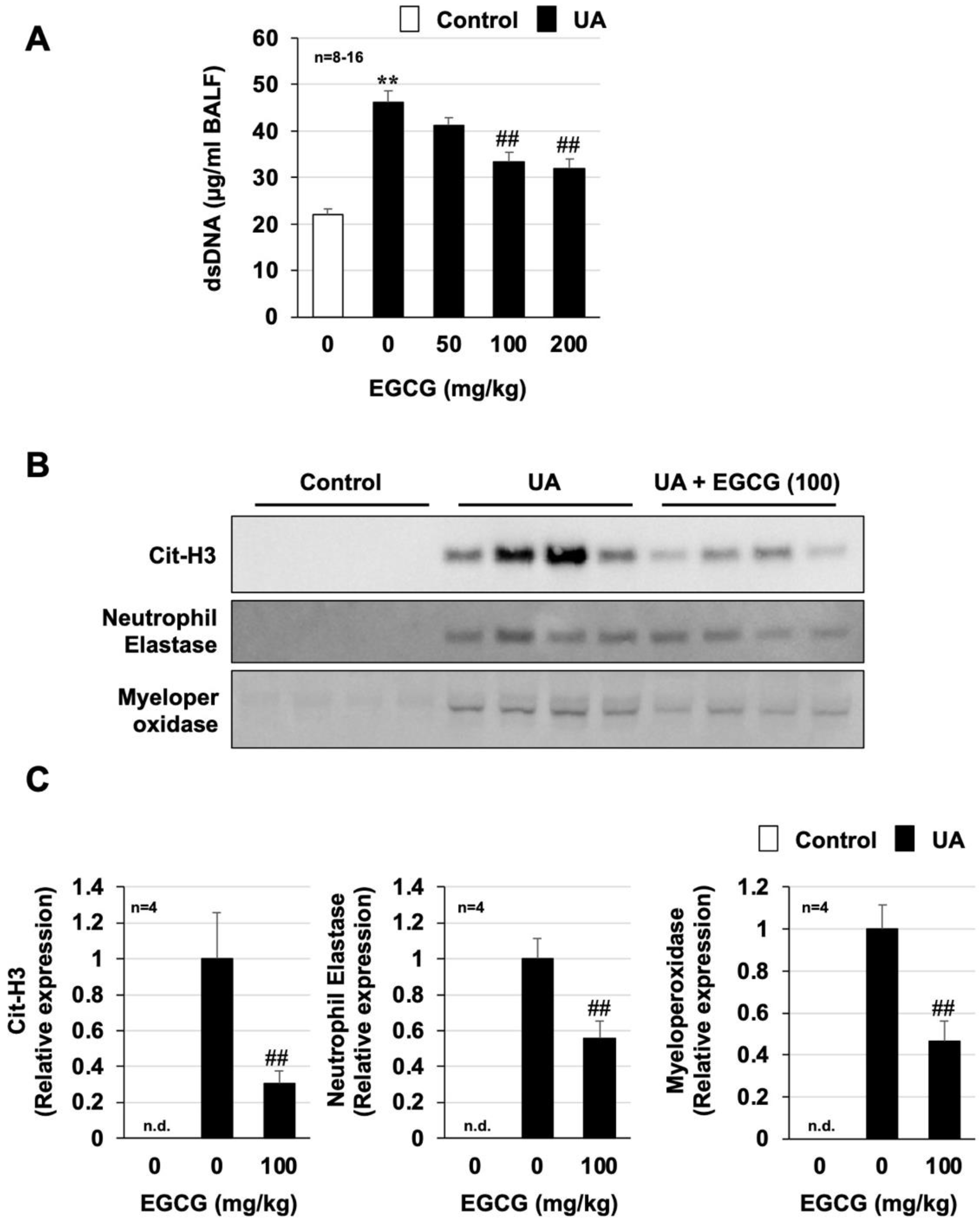
3.4. Effect of EGCG on Urban Aerosol-Induced NET Formation
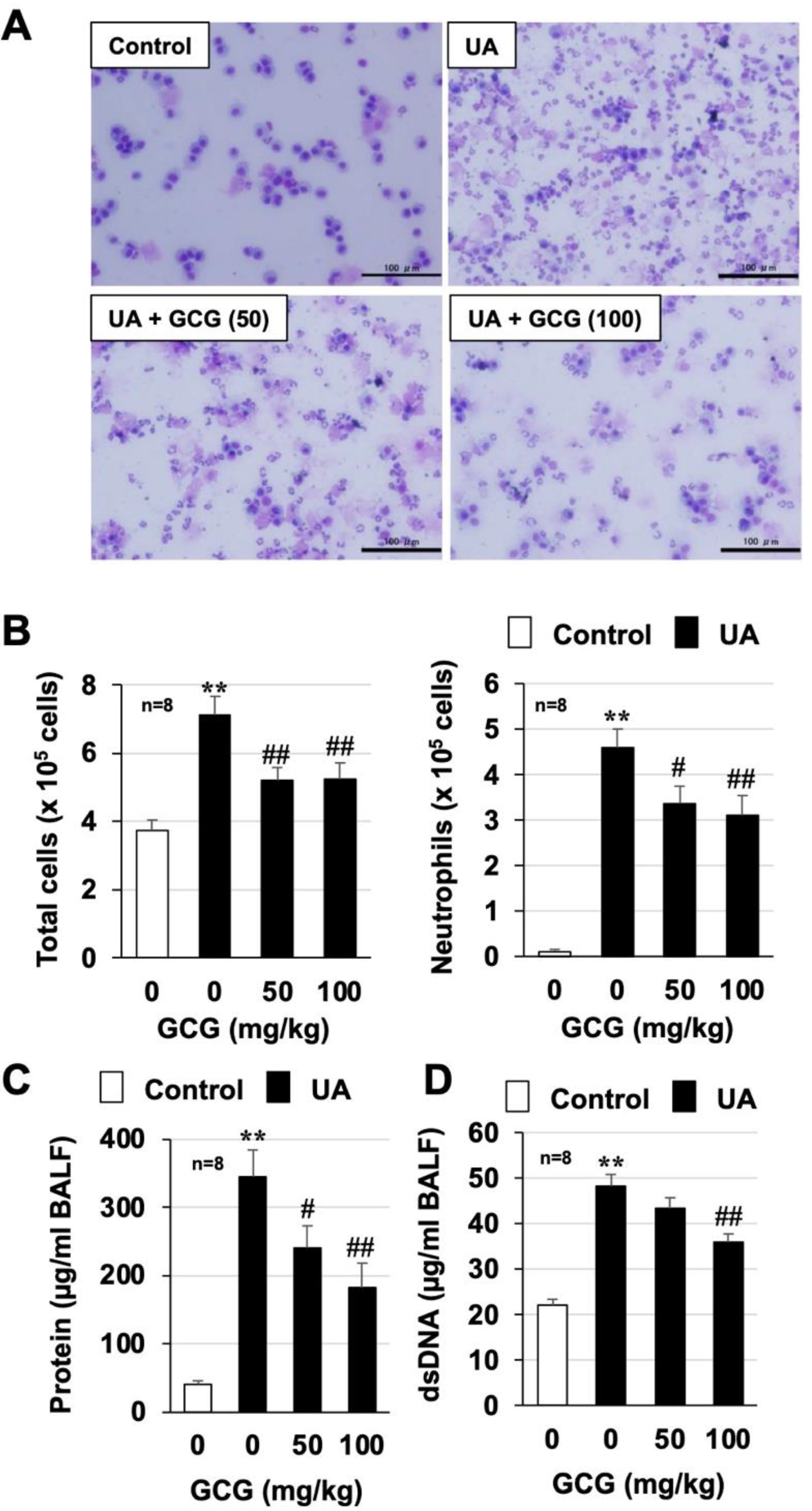
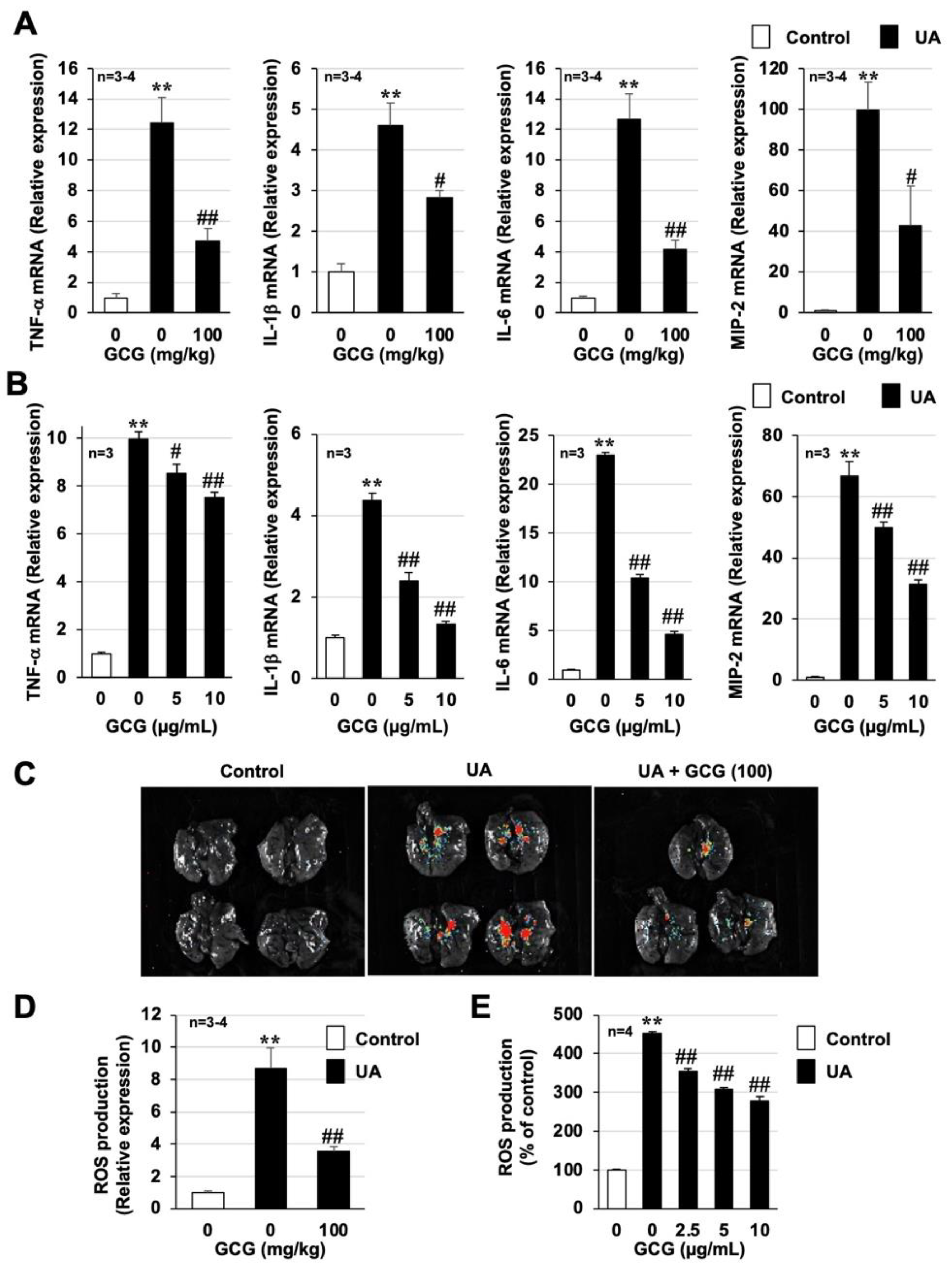
3.5. Effect of GCG on Urban Aerosol-Induced Acute Lung Injury and Inflammatory Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lelieveld, J.; Klingmuller, K.; Pozzer, A.; Poschl, U.; Fnais, M.; Daiber, A.; Munzel, T. Cardiovascular disease burden from ambient air pollution in Europe reassessed using novel hazard ratio functions. Eur. Heart. J. 2019, 40, 1590–1596. [Google Scholar] [CrossRef] [PubMed]
- Losacco, C.; Perillo, A. Particulate matter air pollution and respiratory impact on humans and animals. Environ. Sci. Pollut. Res. Int. 2018, 25, 33901–33910. [Google Scholar] [CrossRef]
- Beloconi, A.; Vounatsou, P. Substantial Reduction in Particulate Matter Air Pollution across Europe during 2006–2019: A Spatiotemporal Modeling Analysis. Environ. Sci. Technol. 2021, 55, 15505–15518. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.H.; Fan, Y.; Nguyen, T.V.; Shin, H.S.; Kim, H.T.; Song, C.H.; Chai, O.H. PM2.5 Exacerbates Oxidative Stress and Inflammatory Response through the Nrf2/NF-kappaB Signaling Pathway in OVA-Induced Allergic Rhinitis Mouse Model. Int. J. Mol. Sci. 2021, 22, 8173. [Google Scholar] [CrossRef]
- Li, J.; Li, H.; Li, H.; Guo, W.; An, Z.; Zeng, X.; Li, W.; Li, H.; Song, J.; Wu, W. Amelioration of PM2.5-induced lung toxicity in rats by nutritional supplementation with fish oil and Vitamin E. Respir. Res. 2019, 20, 76. [Google Scholar] [CrossRef]
- Liu, C.W.; Lee, T.L.; Chen, Y.C.; Liang, C.J.; Wang, S.H.; Lue, J.H.; Tsai, J.S.; Lee, S.W.; Chen, S.H.; Yang, Y.F.; et al. PM2.5-induced oxidative stress increases intercellular adhesion molecule-1 expression in lung epithelial cells through the IL-6/AKT/STAT3/NF-kappaB-dependent pathway. Part. Fibre Toxicol. 2018, 15, 4. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.I.; Kubota, M.; Shimoda, M.; Hayase, T.; Miyaguchi, M.; Kobayashi, N.; Ikeda, M.; Ishima, Y.; Kawahara, M. Thioredoxin-albumin fusion protein prevents urban aerosol-induced lung injury via suppressing oxidative stress-related neutrophil extracellular trap formation. Environ. Pollut. 2021, 268, 115787. [Google Scholar] [CrossRef]
- Li, W.; Wilker, E.H.; Dorans, K.S.; Rice, M.B.; Schwartz, J.; Coull, B.A.; Koutrakis, P.; Gold, D.R.; Keaney, J.F., Jr.; Lin, H.; et al. Short-Term Exposure to Air Pollution and Biomarkers of Oxidative Stress: The Framingham Heart Study. J. Am. Heart Assoc. 2016, 5, e002742. [Google Scholar] [CrossRef]
- Kochman, J.; Jakubczyk, K.; Antoniewicz, J.; Mruk, H.; Janda, K. Health Benefits and Chemical Composition of Matcha Green Tea: A Review. Molecules 2020, 26, 85. [Google Scholar] [CrossRef]
- Chacko, S.M.; Thambi, P.T.; Kuttan, R.; Nishigaki, I. Beneficial effects of green tea: A literature review. Chin. Med. 2010, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Fan, S.M.; Zhang, J. Epigallocatechin-3-gallate ameliorates lipopolysaccharide-induced acute lung injury by suppression of TLR4/NF-kappaB signaling activation. Braz. J. Med. Biol. Res. 2019, 52, e8092. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.J.; Liu, B.J.; Wang, C.L.; Wang, G.H.; Tian, Y.; Wang, S.H.; Li, J.; Li, P.Y.; Zhang, R.H.; Wei, D.; et al. Epigallocatechin-3-gallate inhibits TLR4 signaling through the 67-kDa laminin receptor and effectively alleviates acute lung injury induced by H9N2 swine influenza virus. Int. Immunopharmacol. 2017, 52, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Wu, C.J.; Chien, C.Y.; Chien, C.T. Green Tea Polyphenol Catechins Inhibit Coronavirus Replication and Potentiate the Adaptive Immunity and Autophagy-Dependent Protective Mechanism to Improve Acute Lung Injury in Mice. Antioxidants 2021, 10, 928. [Google Scholar] [CrossRef] [PubMed]
- Ishino, N.; Yanase, E.; Nakatsuka, S. Epimerization of tea catechins under weakly acidic and alkaline conditions. Biosci. Biotechnol. Biochem. 2010, 74, 875–877. [Google Scholar] [CrossRef]
- Kofink, M.; Papagiannopoulos, M.; Galensa, R. (-)-Catechin in cocoa and chocolate: Occurrence and analysis of an atypical flavan-3-ol enantiomer. Molecules 2007, 12, 1274–1288. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, Q.Y.; Tsang, D.; Huang, Y. Degradation of green tea catechins in tea drinks. J. Agric. Food Chem. 2001, 49, 477–482. [Google Scholar] [CrossRef]
- Mori, I.; Sun, Z.; Ukachi, M.; Nagano, K.; McLeod, C.W.; Cox, A.G.; Nishikawa, M. Development and certification of the new NIES CRM 28: Urban aerosols for the determination of multielements. Anal. Bioanal. Chem. 2008, 391, 1997–2003. [Google Scholar] [CrossRef]
- Tanaka, K.I.; Tamura, F.; Sugizaki, T.; Kawahara, M.; Kuba, K.; Imai, Y.; Mizushima, T. Evaluation of Lecithinized Superoxide Dismutase for the Prevention of Acute Respiratory Distress Syndrome in Animal Models. Am. J. Respir. Cell Mol. Biol. 2017, 56, 179–190. [Google Scholar] [CrossRef]
- Tanaka, K.I.; Shiota, S.; Sakakibara, O.; Shimoda, M.; Takafuji, A.; Takabatake, M.; Kadota, Y.; Kawakami, T.; Suzuki, S.; Kawahara, M. Exacerbation of Elastase-Induced Emphysema via Increased Oxidative Stress in Metallothionein-Knockout Mice. Biomolecules 2022, 12, 583. [Google Scholar] [CrossRef]
- Tanaka, K.I.; Shimoda, M.; Kubota, M.; Takafuji, A.; Kawahara, M.; Mizushima, T. Novel pharmacological effects of lecithinized superoxide dismutase on ischemia/reperfusion injury in the kidneys of mice. Life Sci. 2022, 288, 120164. [Google Scholar] [CrossRef]
- Azambuja, J.H.; Mancuso, R.I.; Via, F.I.D.; Torello, C.O.; Saad, S.T.O. Protective effect of green tea and epigallocatechin-3-gallate in a LPS-induced systemic inflammation model. J. Nutr. Biochem. 2022, 101, 108920. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, G.; Tang, M.; Fang, J.; Jiang, H. Epigallocatechin Gallate Can Protect Mice From Acute Stress Induced by LPS While Stabilizing Gut Microbes and Serum Metabolites Levels. Front. Immunol. 2021, 12, 640305. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Ichinose, T.; Yoshida, S.; Ito, T.; He, C.; Yoshida, Y.; Arashidani, K.; Takano, H.; Sun, G.; Shibamoto, T. PM2.5-induced lung inflammation in mice: Differences of inflammatory response in macrophages and type II alveolar cells. J. Appl. Toxicol. 2017, 37, 1203–1218. [Google Scholar] [CrossRef]
- Sarma, J.V.; Ward, P.A. Oxidants and redox signaling in acute lung injury. Compr. Physiol. 2011, 1, 1365–1381. [Google Scholar] [CrossRef] [PubMed]
- Tasaka, S.; Amaya, F.; Hashimoto, S.; Ishizaka, A. Roles of oxidants and redox signaling in the pathogenesis of acute respiratory distress syndrome. Antioxid. Redox Signal. 2008, 10, 739–753. [Google Scholar] [CrossRef]
- Zhang, H.; Xue, L.; Li, B.; Tian, H.; Zhang, Z.; Tao, S. Therapeutic potential of bixin in PM2.5 particles-induced lung injury in an Nrf2-dependent manner. Free Radic. Biol. Med. 2018, 126, 166–176. [Google Scholar] [CrossRef]
- Li, R.; Kou, X.; Xie, L.; Cheng, F.; Geng, H. Effects of ambient PM2.5 on pathological injury, inflammation, oxidative stress, metabolic enzyme activity, and expression of c-fos and c-jun in lungs of rats. Environ. Sci. Pollut. Res. Int. 2015, 22, 20167–20176. [Google Scholar] [CrossRef]
- Stoiber, W.; Obermayer, A.; Steinbacher, P.; Krautgartner, W.D. The Role of Reactive Oxygen Species (ROS) in the Formation of Extracellular Traps (ETs) in Humans. Biomolecules 2015, 5, 702–723. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, L.; Guo, F.; Cao, Y.; Hu, W.; Shi, Y.; Lin, X.; Hou, J.; Li, L.; Ding, X.; et al. Effects of epigallocatechin-3-gallate on the HMGB1/RAGE pathway in PM2.5-exposed asthmatic rats. Biochem. Biophys. Res. Commun. 2019, 513, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, H.; Choi, D.H.; Park, K.; Kim, H.J.; Park, M. Application of green tea catechins, polysaccharides, and flavonol prevent fine dust induced bronchial damage by modulating inflammation and airway cilia. Sci. Rep. 2021, 11, 2232. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lee, W.; Cui, Y.R.; Ahn, G.; Jeon, Y.J. Protective effect of green tea catechin against urban fine dust particle-induced skin aging by regulation of NF-kappaB, AP-1, and MAPKs signaling pathways. Environ. Pollut. 2019, 252, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Z.; Wang, Z.J.; Bai, F.; Qin, X.J.; Cao, J.; Lv, J.Y.; Zhang, M.S. Epigallocatechin-3-gallate protects HUVECs from PM2.5-induced oxidative stress injury by activating critical antioxidant pathways. Molecules 2015, 20, 6626–6639. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, Q.; Liang, Y.; Pan, W.; Bei, Y.; Zhang, Y.; Wang, J.; Jiao, Z. miR-486 inhibits PM2.5-induced apoptosis and oxidative stress in human lung alveolar epithelial A549 cells. Ann. Transl. Med. 2018, 6, 209. [Google Scholar] [CrossRef]
- Porto, B.N.; Stein, R.T. Neutrophil Extracellular Traps in Pulmonary Diseases: Too Much of a Good Thing? Front. Immunol. 2016, 7, 311. [Google Scholar] [CrossRef]
- Yang, H.; Biermann, M.H.; Brauner, J.M.; Liu, Y.; Zhao, Y.; Herrmann, M. New Insights into Neutrophil Extracellular Traps: Mechanisms of Formation and Role in Inflammation. Front. Immunol. 2016, 7, 302. [Google Scholar] [CrossRef]
- Simpson, J.L.; Grissell, T.V.; Douwes, J.; Scott, R.J.; Boyle, M.J.; Gibson, P.G. Innate immune activation in neutrophilic asthma and bronchiectasis. Thorax 2007, 62, 211–218. [Google Scholar] [CrossRef]
- Li, H.; Zhou, X.; Tan, H.; Hu, Y.; Zhang, L.; Liu, S.; Dai, M.; Li, Y.; Li, Q.; Mao, Z.; et al. Neutrophil extracellular traps contribute to the pathogenesis of acid-aspiration-induced ALI/ARDS. Oncotarget 2018, 9, 1772–1784. [Google Scholar] [CrossRef]
- Grabcanovic-Musija, F.; Obermayer, A.; Stoiber, W.; Krautgartner, W.D.; Steinbacher, P.; Winterberg, N.; Bathke, A.C.; Klappacher, M.; Studnicka, M. Neutrophil extracellular trap (NET) formation characterises stable and exacerbated COPD and correlates with airflow limitation. Respir. Res. 2015, 16, 59. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, T.; Hermann, E.; Moller, S.; Klinger, M.; Solbach, W.; Laskay, T.; Behnen, M. Flavonoids and 5-aminosalicylic acid inhibit the formation of neutrophil extracellular traps. Mediat. Inflamm. 2013, 2013, 710239. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, S.; Tang, R.; Qiu, H.; Huang, Q.; Mason, T.G.; Tian, L. Major air pollutants and risk of COPD exacerbations: A systematic review and meta-analysis. Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 3079–3091. [Google Scholar] [CrossRef] [PubMed]
- Scibor, M.; Balcerzak, B.; Galbarczyk, A.; Jasienska, G. Associations between Daily Ambient Air Pollution and Pulmonary Function, Asthma Symptom Occurrence, and Quick-Relief Inhaler Use among Asthma Patients. Int. J. Environ. Res. Public Health 2022, 19, 4852. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, K.-I.; Nakaguchi, S.; Shiota, S.; Nakada, Y.; Oyama, K.; Sakakibara, O.; Shimoda, M.; Sugimoto, A.; Ichitani, M.; Takihara, T.; et al. Preventive Effect of Epigallocatechin Gallate, the Main Component of Green Tea, on Acute Lung Injury Caused by Air Pollutants. Biomolecules 2022, 12, 1196. https://doi.org/10.3390/biom12091196
Tanaka K-I, Nakaguchi S, Shiota S, Nakada Y, Oyama K, Sakakibara O, Shimoda M, Sugimoto A, Ichitani M, Takihara T, et al. Preventive Effect of Epigallocatechin Gallate, the Main Component of Green Tea, on Acute Lung Injury Caused by Air Pollutants. Biomolecules. 2022; 12(9):1196. https://doi.org/10.3390/biom12091196
Chicago/Turabian StyleTanaka, Ken-Ichiro, Shunsuke Nakaguchi, Sachie Shiota, Yuka Nakada, Kaho Oyama, Okina Sakakibara, Mikako Shimoda, Akio Sugimoto, Masaki Ichitani, Takanobu Takihara, and et al. 2022. "Preventive Effect of Epigallocatechin Gallate, the Main Component of Green Tea, on Acute Lung Injury Caused by Air Pollutants" Biomolecules 12, no. 9: 1196. https://doi.org/10.3390/biom12091196