
Programmable Polyproteams of Tyrosine Ammonia Lyases as Cross-Linked Enzymes for Synthesizing p-Coumaric Acid

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
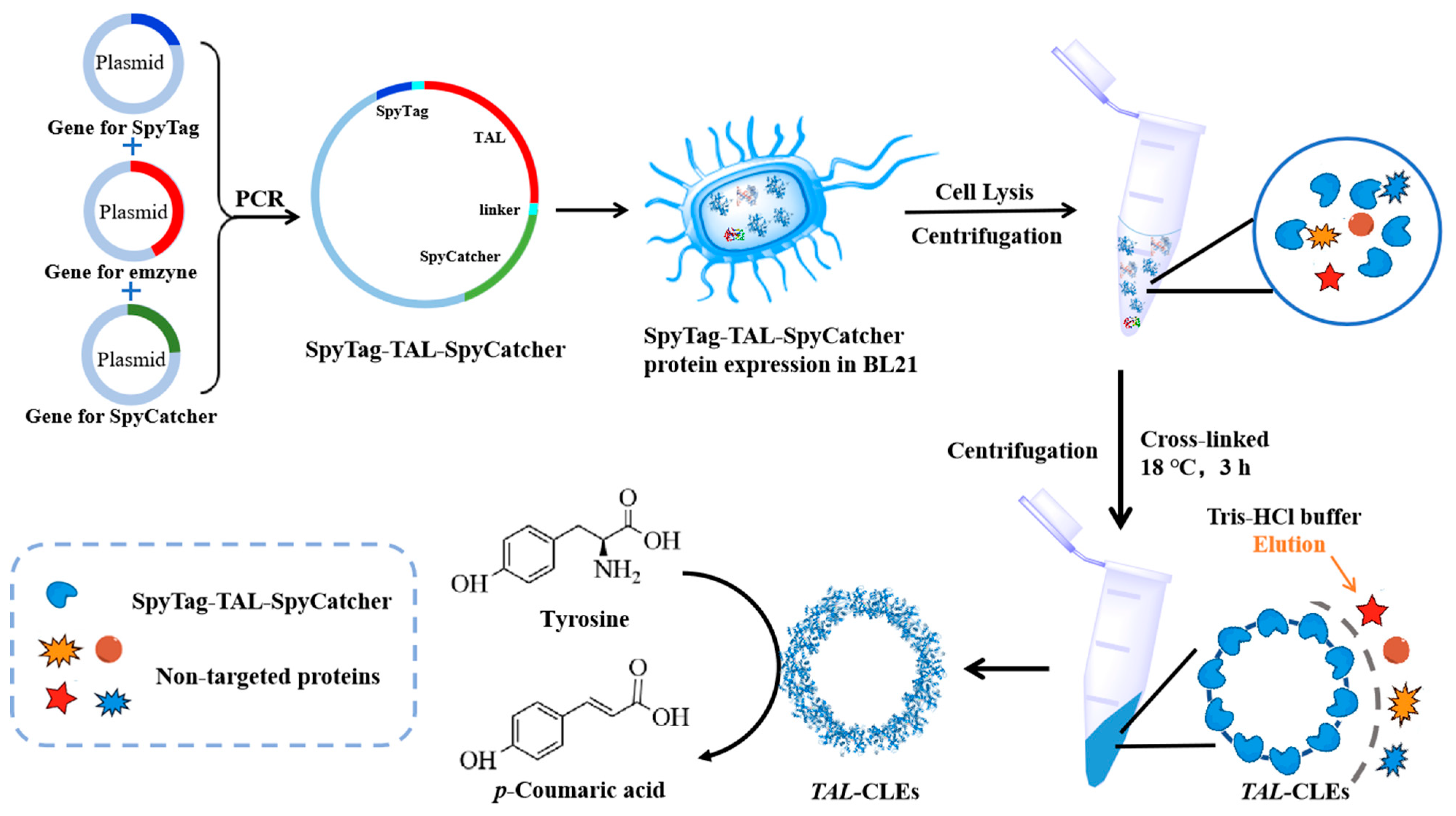
2.2. Plasmid Construction, Protein Expression, and Characterization
2.2.1. Plasmid Construction
2.2.2. Protein Expression and Purification
2.2.3. Protein Characterization
2.3. Preparation and Characterization of CLEs from the Cell Lysate
2.3.1. Preparation of TAL-CLEs and CLEs-GA from the Cell Lysate
2.3.2. Enzymatic Activity Assay
2.3.3. Characterization of TAL-CLEs
2.3.4. Thermal Stability of the TAL-CLEs and WtTAL
2.3.5. Enzymatic Synthesis of p-Coumaric Acid Using TAL-CLEs
3. Results and Discussion
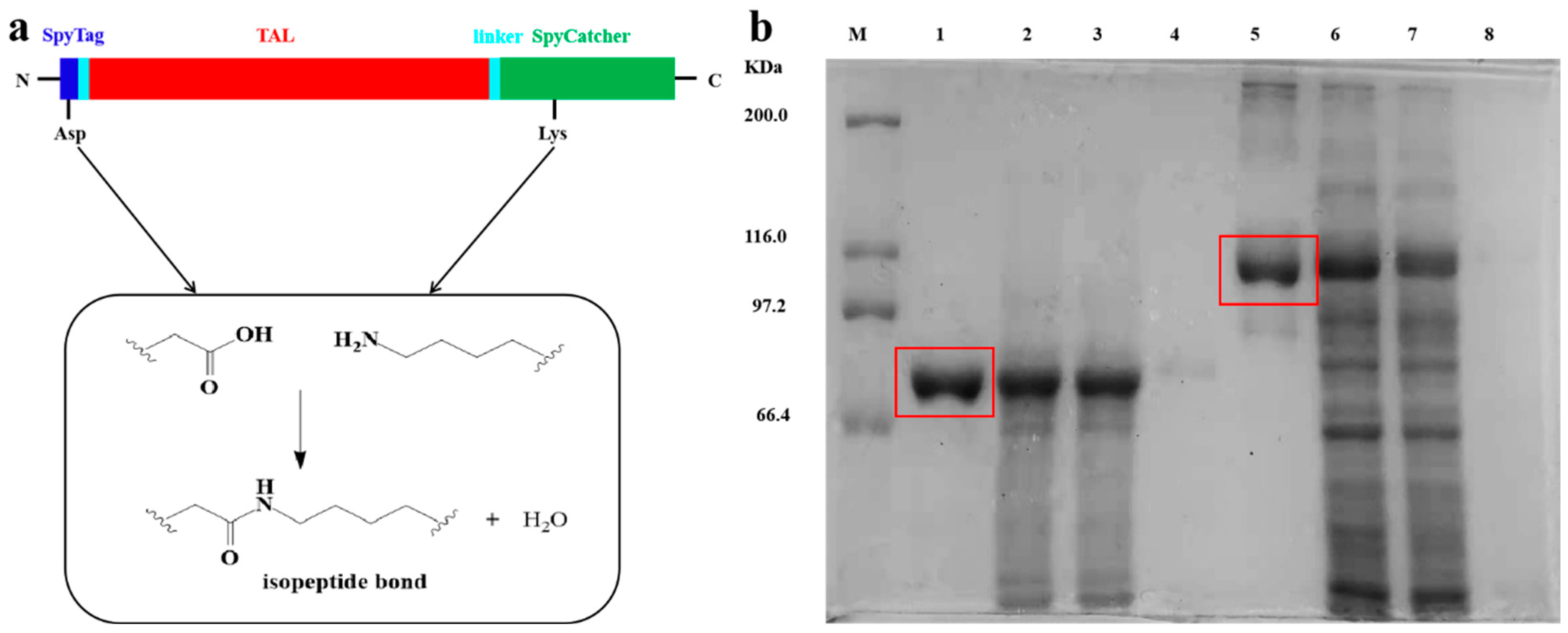
3.1. Expression and Characterization of TAL and SpyTag-TAL-SpyCatcher
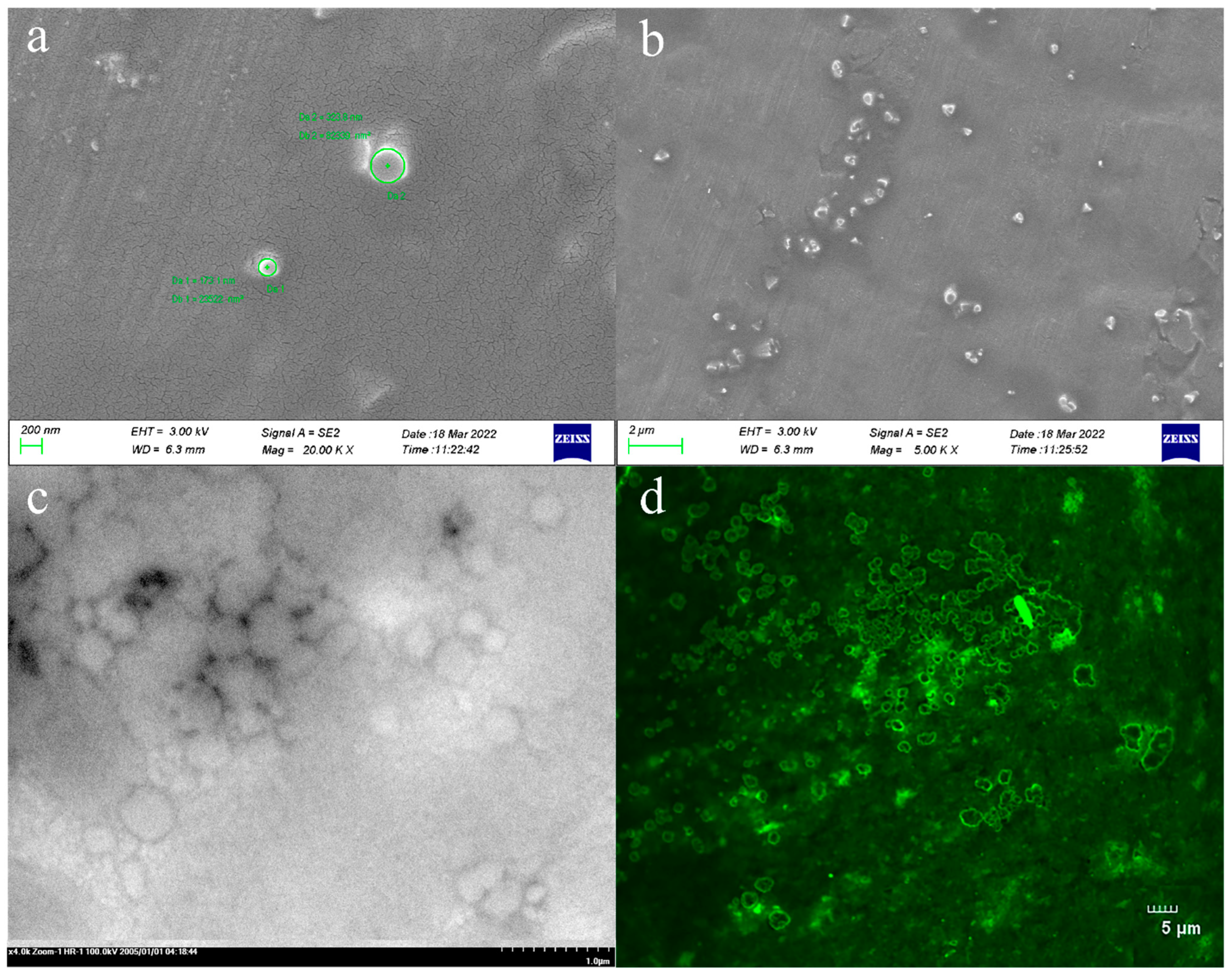
3.2. Morphological Characterization of TAL-CLEs
3.2.1. SEM, TEM, and CLSM Characterization
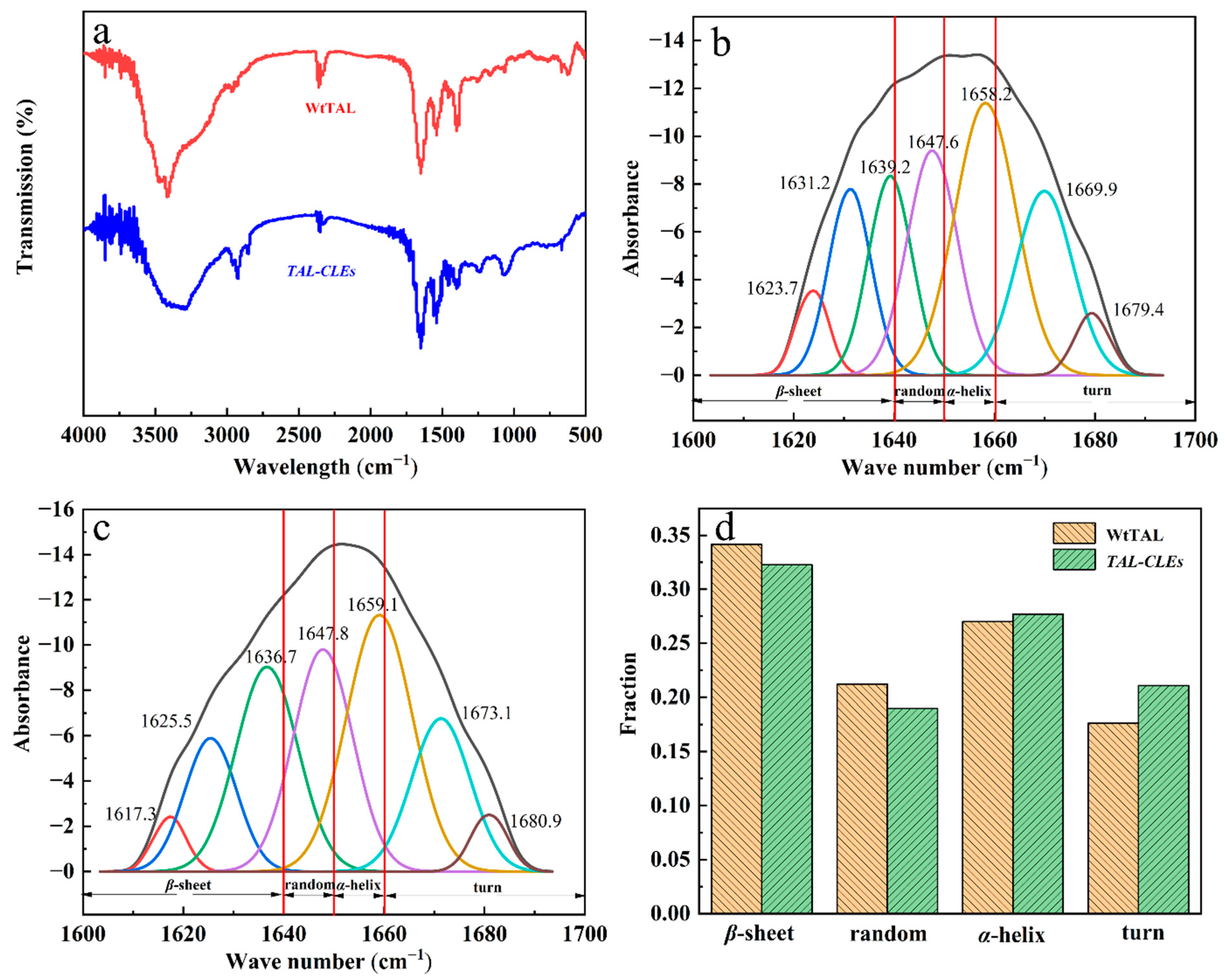
3.2.2. FT-IR Analysis of the Secondary Structure of the TAL-CLEs
3.2.3. Enzymatic Activity Assay
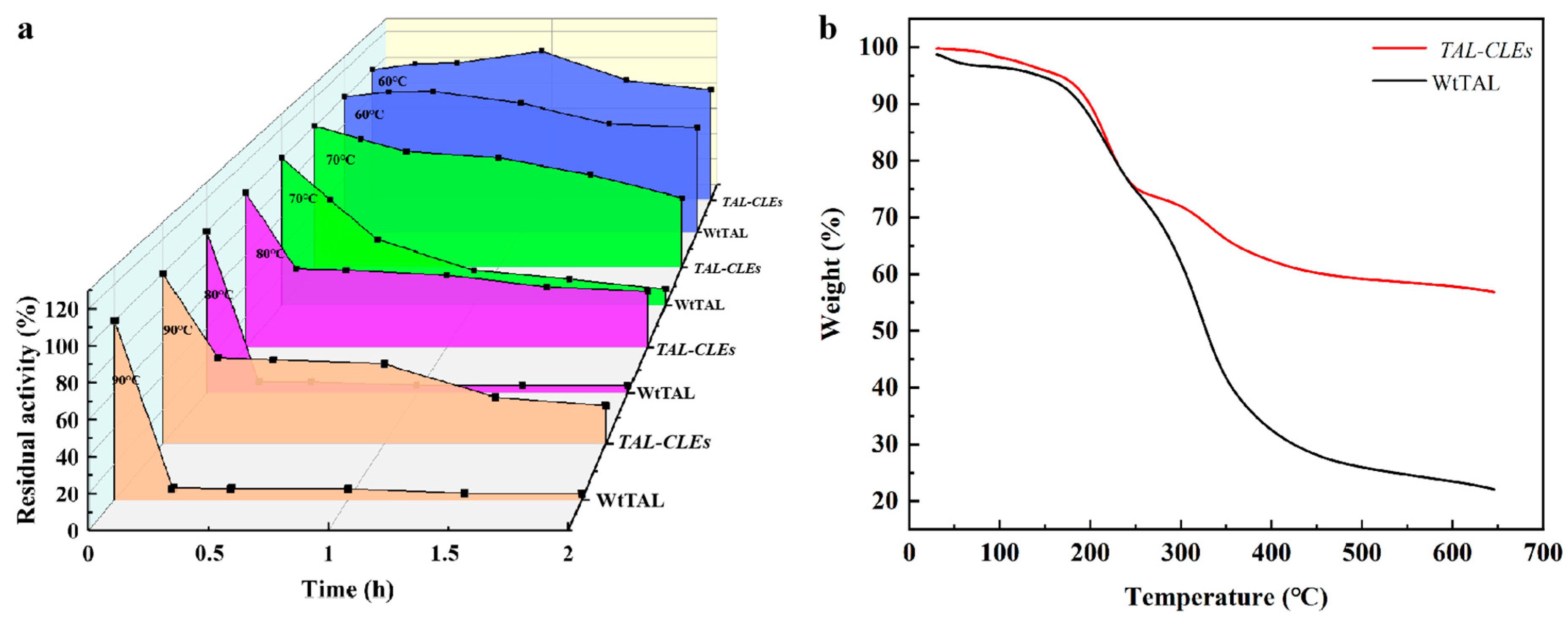
3.2.4. Thermal Stability of the TAL-CLEs
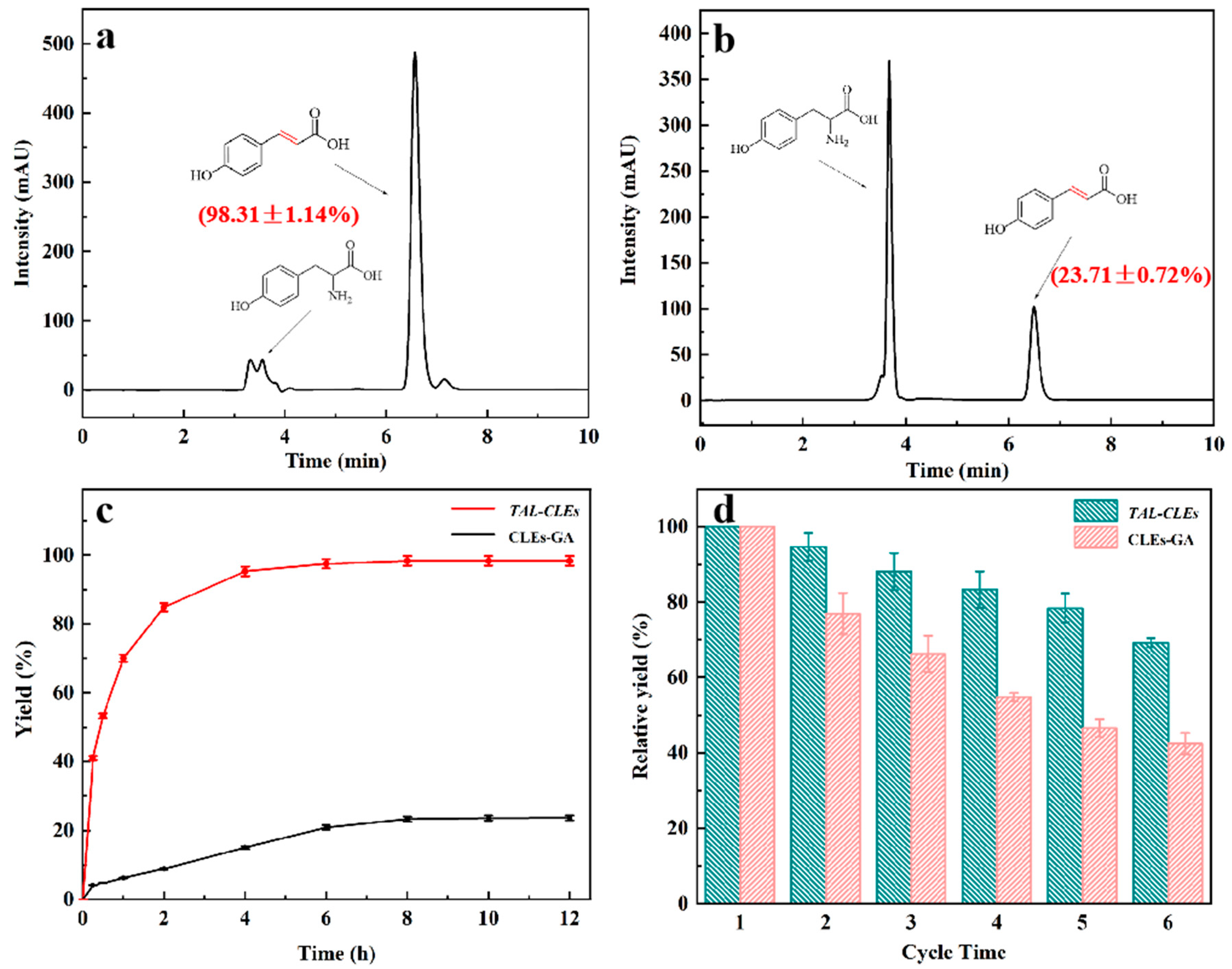
3.3. Enzymatic Synthesis of p-Coumaric Acid Using CLEs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, D.M.; Chen, J.; Shi, Y.P. Advances on methods and easy separated support materials for enzymes immobilization. Trends Anal. Chem. 2018, 102, 332–342. [Google Scholar] [CrossRef]
- Arana Pena, S.; Carballares, D.; Morellon Sterlling, R.; Berenguer Murcia, A.; Alcantara, A.R.; Rodrigues, R.C.; Fernandez Lafuente, R. Enzyme co-immobilization: Always the biocatalyst designers’ choice…or not? Biotechnol. Adv. 2021, 51, 107584. [Google Scholar] [CrossRef] [PubMed]
- Pinney, M.M.; Mokhtari, D.A.; Akiva, E.; Yabukarski, F.; Sanchez, D.M.; Liang, R.B.; Doukov, T.; Martinez, T.J.; Babbitt, P.C.; Herschlag, D. Parallel molecular mechanisms for enzyme temperature adaptation. Science 2021, 371, eaay2784. [Google Scholar] [CrossRef] [PubMed]
- Zahirinejad, S.; Hemmati, R.; Homaei, A.; Dinari, A.; Hosseinkhani, S.; Mohammadi, S.; Vianello, F. Nano-organic supports for enzyme immobilization: Scopes and perspectives. Colloids Surf. B 2021, 204, 111774. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.; Ermler, U.; Shima, S. The methanogenic CO2 reducing-and-fixing enzyme is bifunctional and contains 46 4Fe-4S clusters. Science 2016, 354, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.M.; Griffin, E.G. Adsorption of invertase. J. Am. Chem. Soc. 1916, 38, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.Q.; Johnston, T.; Quin, M.B.; Schmidt-Dannert, C. Developing a protein scaffoldingsystem for rapid enzyme immobilization and optimization of enzyme functions for biocatalysis. ACS Synth. Biol. 2019, 8, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Gennari, A.; Simon, R.; Sperotto, N.D.D.M.; Bizarro, C.V.; Basso, L.A.; Machado, P.; Benvenutti, E.V.; Renard, G.; Chies, J.M.; Volpato, G.; et al. Application of cellulosic materials as supports for single-step purification and immobilization of a recombinant beta-galactosidase via cellulose-binding domain. Int. J. Biol. Macromol. 2022, 199, 307–317. [Google Scholar] [CrossRef]
- Morsi, R.; Al-Maqdi, K.A.; Bilal, M.; Iqbal, H.M.N.; Khaleel, A.; Shah, I.L.; Ashraf, S.S. Immobilized soybean peroxidase hybrid biocatalysts for efficient degradation of various emerging pollutants. Biomolecules 2021, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; Jiang, W.; Fu, X.l.; Tian, L.b.; Zhou, S.F. 3D-Printed Aldo-keto reductase within biocompatible polymers as catalyst for chiral drug intermediate. Chem. Eng. J. 2022, 429, 132293. [Google Scholar] [CrossRef]
- Shakeri, F.; Ariaeenejad, S.; Ghollasi, M.; Motamedi, E. Synthesis of two novel bio-based hydrogels using sodium alginate and chitosan and their proficiency in physical immobilization of enzymes. Sci. Rep. 2022, 12, 2072. [Google Scholar] [CrossRef]
- Liu, F.; Shi, Z.; Su, W.; Wu, J. State of the art and applications in nanostructured biocatalysis. Biotechnol. Biotechnol. Equip. 2022, 36, 117–133. [Google Scholar] [CrossRef]
- Parthasarathy, R.V.; Martin, C.R. Synthesis of polymeric microcapsule arrays and their use for enzyme immobilization. Nature 1994, 369, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Rivero, C.W.; Garcia, N.S.; Fernandez-Lucas, J.; Betancor, L.; Romanelli, G.P.; Trelles, J.A. Green production of cladribine by using immobilized 2′-deoxyribosyltransferase from Lactobacillus delbrueckii stabilized through a double covalent/entrapment technology. Biomolecules 2021, 11, 657. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, J.; Luo, Z.; Xie, T.; Xiao, Q.; Pei, X.; Wang, A. Controllably crosslinked dual enzymes enabled by genetic-encoded non-standard amino acid for efficiently enantioselective hydrogenation. Int. J. Biol. Macromol. 2022, 205, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Pereira Goncalves, M.C.; Kieckbusch, T.G.; Perna, R.F.; Fujimoto, J.T.; Morales, S.A.V.; Romanelli, J.P. Trends on enzyme immobilization researches based on bibliometric analysis. Process Biochem. 2019, 76, 95–110. [Google Scholar] [CrossRef]
- Li, H.; Wang, R.; Wang, A.; Zhang, J.; Yin, Y.; Pei, X.; Zhang, P. Rapidly and precisely cross-linked enzymes using bio-orthogonal chemistry from cell lysate for the synthesis of (s)-1-(2,6-dichloro-3-fluorophenyl) ethanol. ACS Sustain. Chem. Eng. 2020, 8, 6466–6478. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, R.; Zhang, J.; Luo, Z.; Xiao, Q.; Xie, T.; Pei, X.; Gao, P.; Wang, A. Efficiently enantioselective hydrogenation photosynthesis of (R)-1-3,5-bis(trifluoromethyl)phenyl ethanol over a CLEs-TiO2 bioinorganic hybrid materials. ACS Appl. Mater. Interfaces 2021, 13, 41454–41463. [Google Scholar] [CrossRef]
- Chauhan, V.; Kaushal, D.; Dhiman, V.K.; Kanwar, S.S.; Singh, D.; Dhiman, V.K.; Pandey, H. An insight in developing carrier-free immobilized enzymes. Front. Bioeng. Biotechnol. 2022, 10, 794411. [Google Scholar] [CrossRef]
- Wilson, L.; Illanes, A.; Romero, O.; Vergara, J.; Mateo, C. Carrier-bound and carrier-free penicillin acylase biocatalysts for the thermodynamically controlled synthesis of beta-lactam compounds in organic medium. Enzym. Microb. Technol. 2008, 43, 442–447. [Google Scholar] [CrossRef]
- Sheldon, R.A. Cross-linked enzyme aggregates as industrial biocatalysts. Org. Process Res. Dev. 2011, 15, 213–223. [Google Scholar] [CrossRef]
- Naseer, S.; Jie, O.; Chen, X.; Pu, S.; Guo, Y.; Zhang, X.; Li, D.; Yang, C. Immobilization of beta-glucosidase by self-catalysis and compared to crosslinking with glutaraldehyde. Int. J. Biol. Macromol. 2020, 154, 1490–1495. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Han, N.; Wu, Y.; Wei, D. Oriented covalent immobilization of recombinant protein A on the glutaraldehyde activated agarose support. Int. J. Biol. Macromol. 2018, 120, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Neumann, H.; Wang, K.H.; Davis, L.; Garcia-Alai, M.; Chin, J.W. Encoding multiple unnatural amino acids via evolution of a quadruplet-decoding ribosome. Nature 2010, 464, 441–444. [Google Scholar] [CrossRef] [Green Version]
- Smolskaya, S.; Andreev, Y.A. Site-specific incorporation of unnatural amino acids into escherichia coli recombinant protein: Methodology development and recent achievement. Biomolecules 2019, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, R.A.; Basso, A.; Brady, D. New frontiers in enzyme immobilisation: Robust biocatalysts for a circular bio-based economy. Chem. Soc. Rev. 2021, 50, 5850–5862. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, R.; Luo, Z.Y.; Jia, D.M.; Chen, H.M.; Xiao, Q.J.; Zhang, P.F.; Pei, X.L.; Wang, A.M. Controlled chemical assembly of enzymes in cell lysate enabled by genetic-encoded nonstandard amino acids. Mater. Chem. Front. 2022, 6, 182–193. [Google Scholar] [CrossRef]
- Sun, X.B.; Cao, J.W.; Wang, J.K.; Lin, H.Z.; Gao, D.Y.; Qian, G.Y.; Park, Y.D.; Chen, Z.F.; Wang, Q. SpyTag/SpyCatcher molecular cyclization confers protein stability and resilience to aggregation. New Biotechnol. 2019, 49, 28–36. [Google Scholar] [CrossRef]
- Dong, W.; Sun, H.; Chen, Q.; Hou, L.; Chang, Y.; Luo, H. SpyTag/Catcher chemistry induces the formation of active inclusion bodies in E. coli. Int. J. Biol. Macromol. 2022, 199, 358–371. [Google Scholar] [CrossRef]
- Qu, J.L.; Cao, S.; Wei, Q.X.; Zhang, H.W.; Wang, R.; Kang, W.; Ma, T.; Zhang, L.; Liu, T.G.; Au, S.W.N.; et al. Synthetic multienzyme complexes, catalytic nanomachineries for cascade biosynthesis in vivo. Acs Nano 2019, 13, 9895–9906. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Y.; Zhou, X.; Liu, N.; Ming, D.; Zhu, L.; Jiang, L. Improving the thermostability of trehalose synthase from Thermomonospora curvata by covalent cyclization using peptide tags and investigation of the underlying molecular mechanism. Int. J. Biol. Macromol. 2021, 168, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Veggiani, G.; Nakamura, T.; Brenner, M.D.; Gayet, R.V.; Yan, J.; Robinson, C.V.; Howarth, M. Programmable polyproteams built using twin peptide superglues. Proc. Natl. Acad. Sci. USA 2016, 113, 1202–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.L.; Hoon, S.S.; Wong, F.T. Kinetic controlled Tag-Catcher interactions for directed covalent protein assembly. PLoS ONE 2016, 11, e0165074. [Google Scholar] [CrossRef]
- Keeble, A.H.; Yadav, V.K.; Ferla, M.P.; Bauer, C.C.; Chuntharpursat-Bon, E.; Huang, J.; Bon, R.S.; Howarth, M. Resource DogCatcher allows loop-friendly protein-protein ligation. Cell Chem. Biol. 2022, 29, 339. [Google Scholar] [CrossRef]
- Proeschel, M.; Kraner, M.E.; Horn, A.H.C.; Schaefer, L.; Sonnewald, U.; Sticht, H. Probing the potential of CnaB-type domains for the design of tag/catcher systems. PLoS ONE 2017, 12, e0179740. [Google Scholar] [CrossRef]
- Hentrich, C.; Kellmann, S.J.; Putyrski, M.; Cavada, M.; Hanuschka, H.; Knappik, A.; Ylera, F. Periplasmic expression of SpyTagged antibody fragments enables rapid modular antibody assembly. Cell Chem. Biol. 2021, 28, 813. [Google Scholar] [CrossRef]
- Schoene, C.; Fierer, J.O.; Bennett, S.P.; Howarth, M. SpyTag/SpyCatcher cyclization confers resilience to boiling on a mesophilic enzyme. Angew. Chem. Int. Ed. 2014, 53, 6101–6104. [Google Scholar] [CrossRef] [Green Version]
- Schoene, C.; Bennett, S.P.; Howarth, M. SpyRing interrogation: Analyzing how enzyme resilience can be achieved with phytase and distinct cyclization chemistries. Sci. Rep. 2016, 6, 21151. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Wang, X.; Zhang, D.; Wu, S.; Zhang, G. Enhanced thermal stability of lichenase from Bacillus subtilis 168 by SpyTag/SpyCatcher-mediated spontaneous cyclization. Biotechnol. Biofuels 2016, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Fok, H.K.F.; Luo, J.; Yang, Y.; Wang, R.; Huang, X.; Sun, F. B-12-induced reassembly of split photoreceptor protein enables photoresponsive hydrogels with tunable mechanics. Sci. Adv. 2022, 8, eabm5482. [Google Scholar] [CrossRef]
- Freitas, A.I.; Domingues, L.; Aguiar, T.Q. Tag-mediated single-step purification and immobilization of recombinant proteins toward protein-engineered advanced materials. J. Adv. Res. 2022, 36, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Brune, K.D.; Leneghan, D.B.; Brian, I.J.; Ishizuka, A.S.; Bachmann, M.F.; Draper, S.J.; Biswas, S.; Howarth, M. Plug-and-Display: Decoration of Virus-Like Particles via isopeptide bonds for modular immunization. Sci. Rep. 2016, 6, 19234. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, J.; Cartannaz, J.; Tourcier, G.; Contreras-Martel, C.; Kleman, J.P.; Morlot, C.; Vernet, T.; Di Guilmi, A.M. Autocatalytic association of proteins by covalent bond formation: A bio molecular welding toolbox derived from a bacterial adhesin. Sci. Rep. 2017, 7, 43564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chang, Y.; Jia, R.; Sun, H.; Tian, J.; Luo, H.; Yu, H.; Shen, Z. SpyTag/SpyCatcher cyclization and covalent immobilization in enhancing cephalosporin C acylase stability. Process Biochem. 2020, 95, 260–268. [Google Scholar] [CrossRef]
- Xue, Z.; McCluskey, M.; Cantera, K.; Ben-Bassat, A.; Sariaslani, R.S.; Huang, L. Improved production of p-hydroxycinnamic acid from tyrosine using a novel thermostable phenylalanine/tyrosine ammonia lyase enzyme. Enzym. Microb. Technol. 2007, 42, 58–64. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, B.H.; Liu, Z.N.; Qiao, J.J.; Zhao, G.R. Combinatorial optimization of resveratrol production in engineered E. coli. J. Agric. Food Chem. 2018, 66, 13444–13453. [Google Scholar] [CrossRef]
- Xue, Z.; McCluskey, M.; Cantera, K.; Sariaslani, F.S.; Huang, L. Identification, characterization and functional expression of a tyrosine ammonia-lyase and its mutants from the photosynthetic bacterium Rhodobacter sphaeroides. J. Ind. Microbiol. Biotechnol. 2007, 34, 599–604. [Google Scholar] [CrossRef]
- van Pelt, S.; Quignard, S.; Kubac, D.; Sorokin, D.Y.; van Rantwijk, F.; Sheldon, R.A. Nitrile hydratase CLEAs: The immobilization and stabilization of an industrially important enzyme. Green Chem. 2008, 10, 395–400. [Google Scholar] [CrossRef]
- Wang, A.; Zhang, F.; Chen, F.; Wang, M.; Li, H.; Zeng, Z.; Xie, T.; Chen, Z. A facile technique to prepare cross-linked enzyme aggregates using p-benzoquinone as cross-linking agent. Korean J. Chem. Eng. 2011, 28, 1090–1095. [Google Scholar] [CrossRef]
- Terao, Y.; Kawabata, S.; Nakata, M.; Nakagawa, I.; Hamada, S. Molecular characterization of a novel fibronectin-binding protein of Streptococcus pyogenes strains isolated from toxic shock-like syndrome patients. J. Biol. Chem. 2002, 277, 47428–47435. [Google Scholar] [CrossRef] [Green Version]
- Zakeri, B.; Fierer, J.O.; Celik, E.; Chittock, E.C.; Schwarz-Linek, U.; Moy, V.T.; Howarth, M. Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proc. Natl. Acad. Sci. USA 2012, 109, E690–E697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, M.; Xu, Q.; Jiang, L.; Huang, H. SpyTag/SpyCatcher cyclization enhances the thermostability of firefly luciferase. PLoS ONE 2016, 11, e0162318. [Google Scholar] [CrossRef] [PubMed]
- da Silva-Buzanello, R.A.; Schuch, A.F.; Gasparin, A.W.; Torquato, A.S.; Scremin, F.R.; Canan, C.; Soares, A.L. Quality parameters of chicken breast meat affected by carcass scalding conditions. Asian-Australas. J. Anim. Sci. 2019, 32, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Miao, L.; Li, J.; Fu, S.; An, G.; Si, C.; Dong, Z.; Luo, Q.; Yu, S.; Xu, J.; et al. Self-assembly of cricoid proteins induced by “soft nanoparticles”: An approach to design multienzyme-cooperative antioxidative systems. Acs Nano 2015, 9, 5461–5469. [Google Scholar] [CrossRef]
- Li, L.; Fierer, J.O.; Rapoport, T.A.; Howarth, M. Structural analysis and optimization of the covalent association between SpyCatcher and a peptide Tag. J. Mol. Biol. 2014, 426, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Jin, W.; Cai, L.; Liu, X.; Qiu, Y.; Zhang, G. Green preparation of covalently co-immobilized multienzymes on silica nanoparticles for clean production of reducing sugar from lignocellulosic biomass. J. Clean. Prod. 2021, 314, 127994. [Google Scholar] [CrossRef]
- Vicente, F.A.; Plazl, I.; Ventura, S.P.M.; Znidarsic-Plazl, P. Separation and purification of biomacromolecules based on microfluidics. Green Chem. 2020, 22, 4391–4410. [Google Scholar] [CrossRef]
- Trotman, R.J.; Camp, C.E.; Ben-Bassat, A.; DiCosimo, R.; Huang, L.; Crum, G.A.; Sariaslani, F.S.; Haynie, S.L. Calcium alginate bead immobilization of cells containing tyrosine ammonia lyase activity for use in the production of p-hydroxycinnamic acid. Biotechnol. Prog. 2008, 23, 638–644. [Google Scholar] [CrossRef]
- Nadar, S.S.; Rathod, V.K. Magnetic macromolecular cross linked enzyme aggregates (CLEAs) of glucoamylase. Enzym. Microb. Technol. 2016, 83, 78–87. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Enzyme | Enzyme Activity (U·mg−1) |
|---|---|---|
| 1 | WtTAL | 0.43 ± 0.02 |
| 2 | SpyTag-TAL-SpyCatcher | 0.35 ± 0.02 |
| 3 | TAL-CLEs | 1.82 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, M.; Luo, Z.; Chen, H.; Ma, B.; Qiao, L.; Xiao, Q.; Zhang, P.; Wang, A. Programmable Polyproteams of Tyrosine Ammonia Lyases as Cross-Linked Enzymes for Synthesizing p-Coumaric Acid. Biomolecules 2022, 12, 997. https://doi.org/10.3390/biom12070997
Jia M, Luo Z, Chen H, Ma B, Qiao L, Xiao Q, Zhang P, Wang A. Programmable Polyproteams of Tyrosine Ammonia Lyases as Cross-Linked Enzymes for Synthesizing p-Coumaric Acid. Biomolecules. 2022; 12(7):997. https://doi.org/10.3390/biom12070997
Chicago/Turabian StyleJia, Mingyu, Zhiyuan Luo, Haomin Chen, Bianqin Ma, Li Qiao, Qinjie Xiao, Pengfei Zhang, and Anming Wang. 2022. "Programmable Polyproteams of Tyrosine Ammonia Lyases as Cross-Linked Enzymes for Synthesizing p-Coumaric Acid" Biomolecules 12, no. 7: 997. https://doi.org/10.3390/biom12070997