
Systemic Copper Disorders Influence the Olfactory Function in Adult Rats: Roles of Altered Adult Neurogenesis and Neurochemical Imbalance


Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
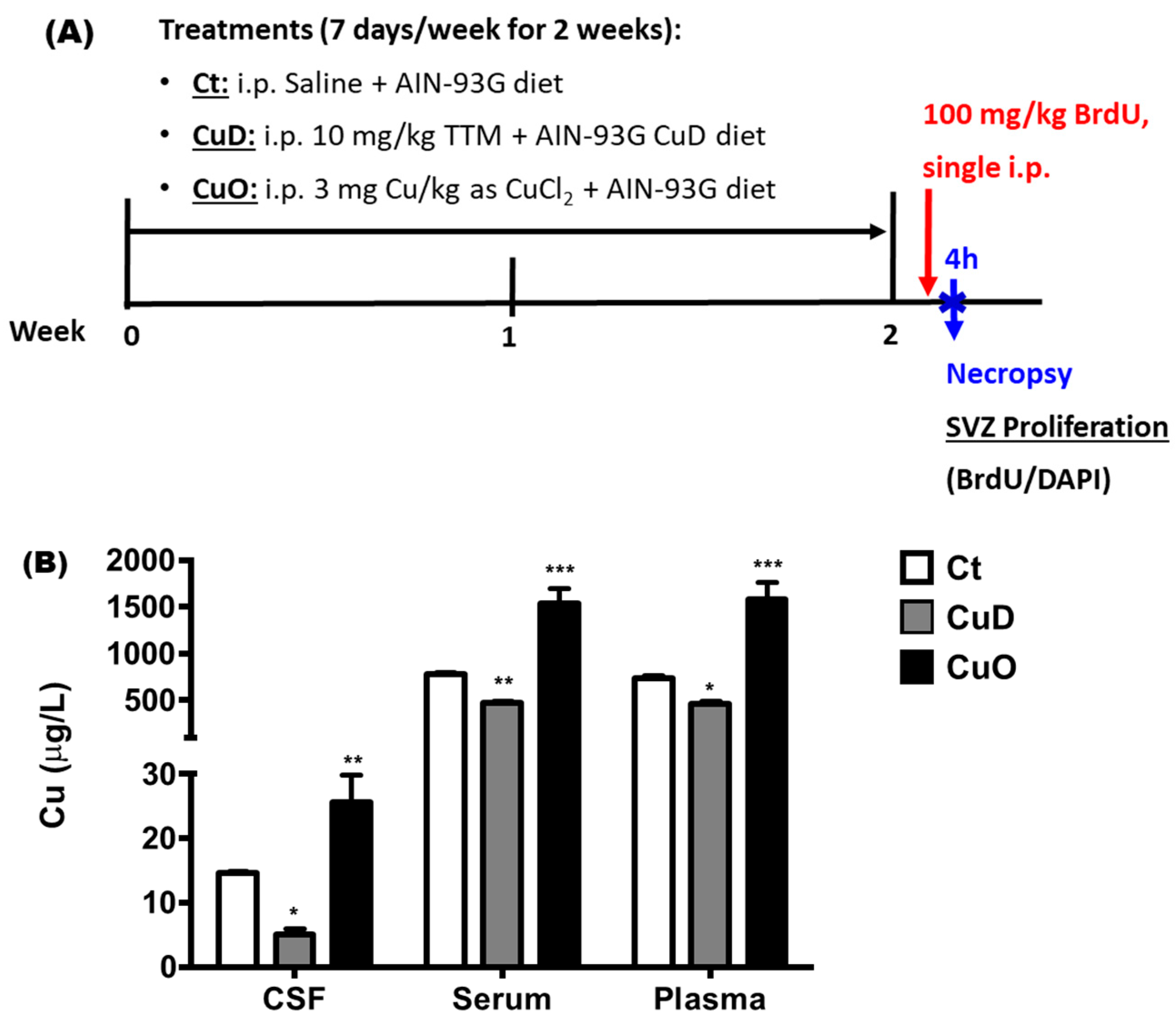
2.2. Animals and Treatment
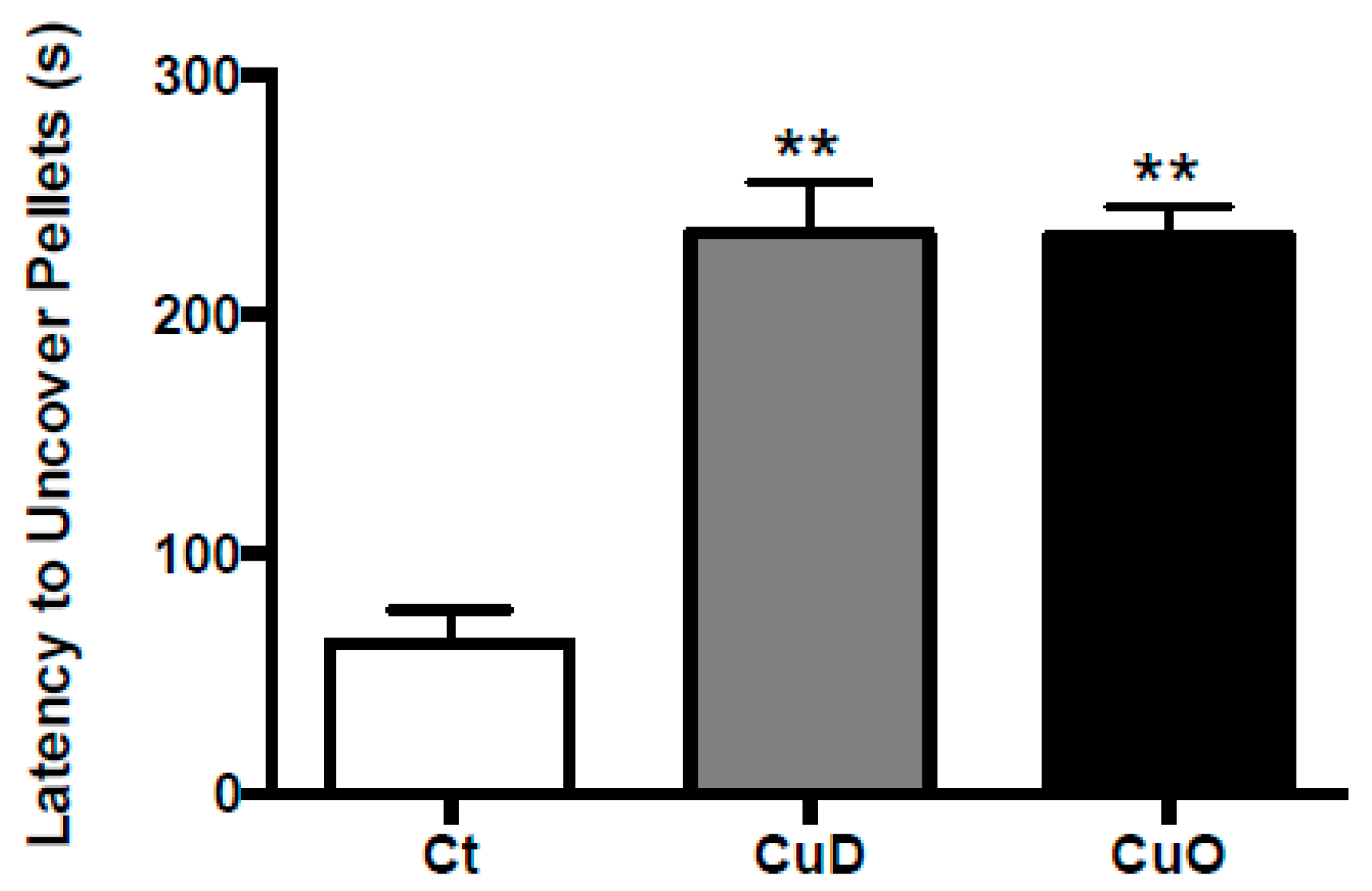
2.3. Buried Pellet Test
2.4. Brain Tissue Collection
2.5. Determination of Cu Concentration by Atomic Absorption Spectroscopy (AAS)
2.6. Immunohistochemistry (IHC) Staining
2.7. Confocal Imaging and Cell Counting
2.8. High-Performance Liquid Chromatography (HPLC)
2.9. Quantitative Real-Time RT-PCR
2.10. Statistical Analysis
3. Results
3.1. Systemic Cu Dyshomeostasis and Impaired Olfactory Function Following CuD or CuO Treatment
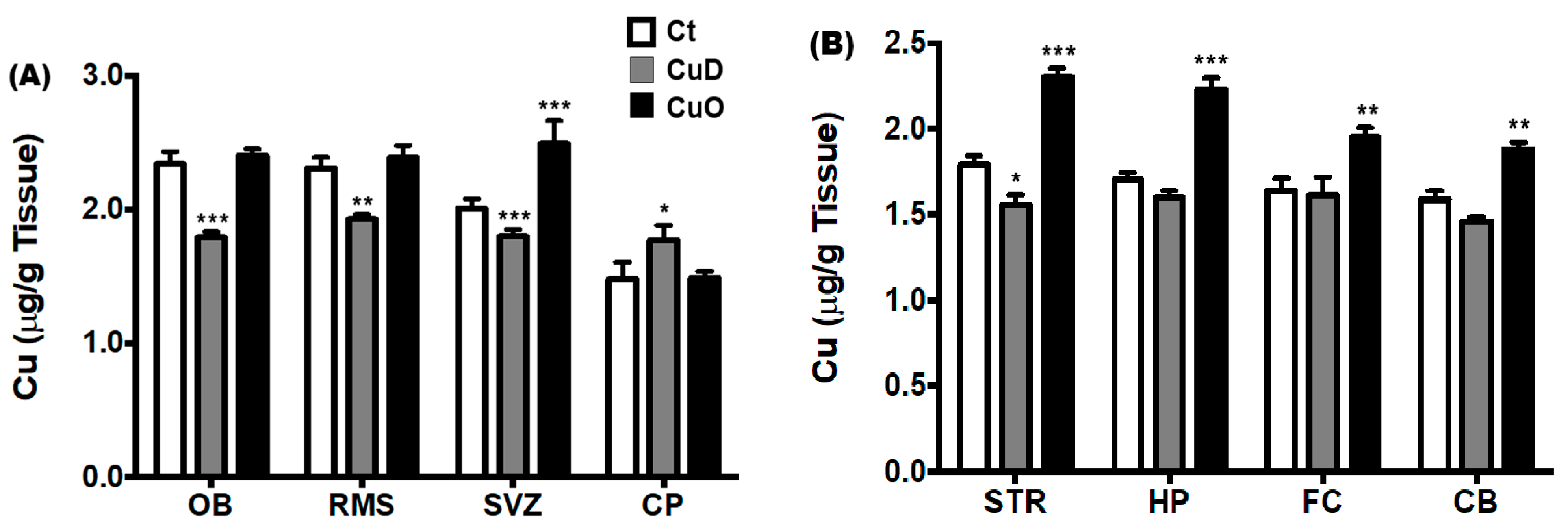
3.2. Altered Brain Regional Cu Levels Following CuD or CuO Treatment
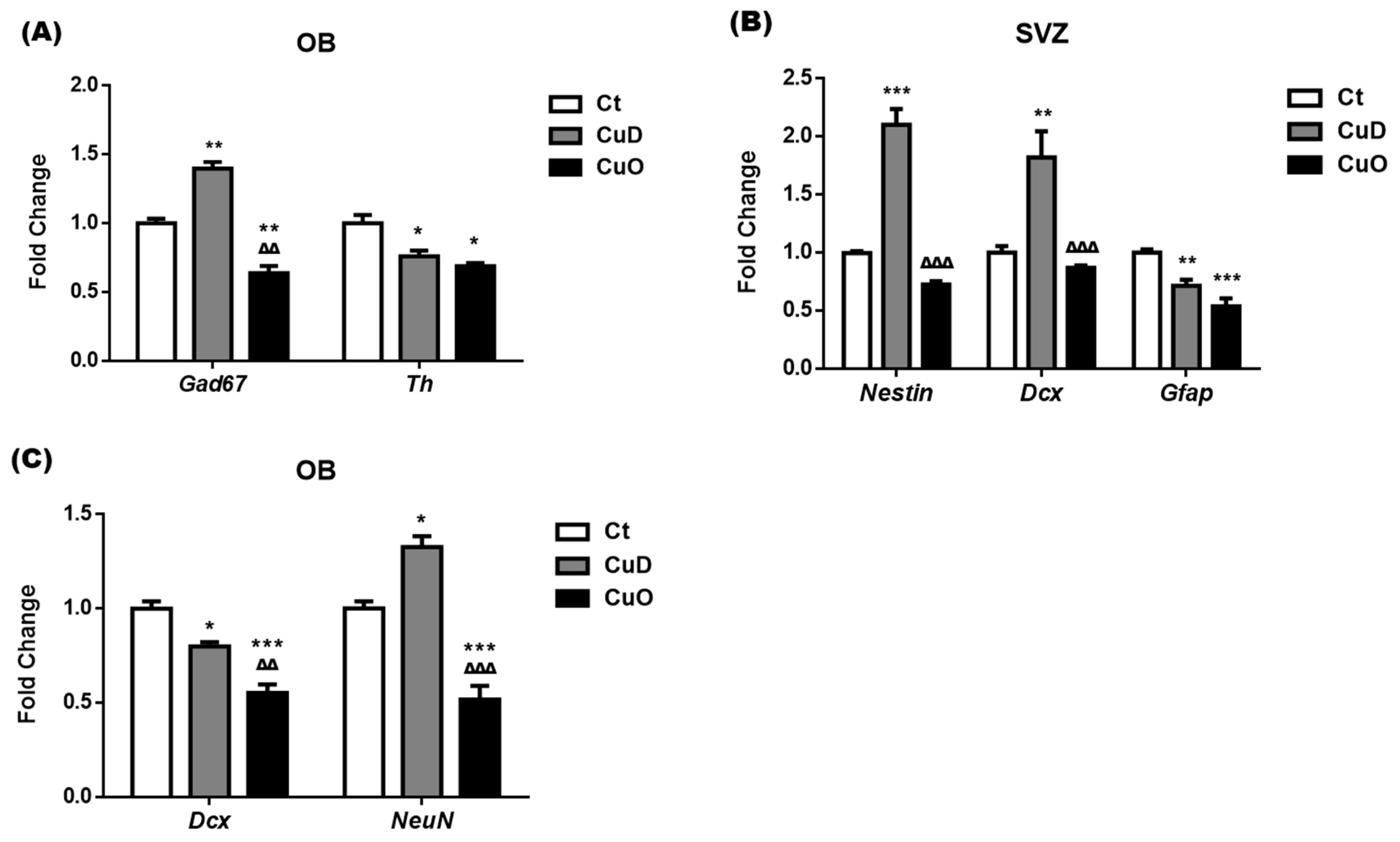
3.3. Altered Expressions of Neuronal Markers Following Systemic CuD or CuO Treatment
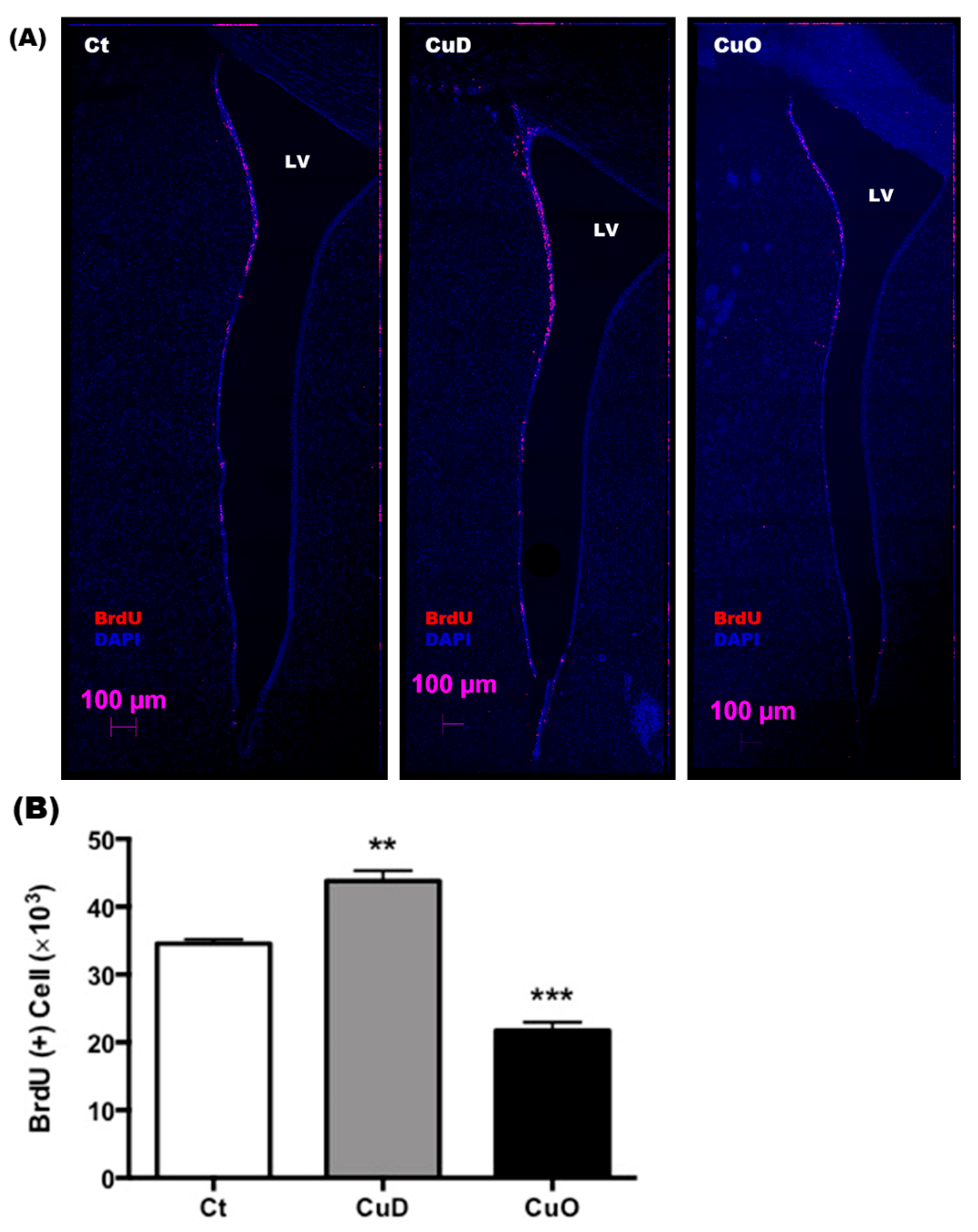
3.4. Altered NSPC Proliferation in Adult SVZ Following Systemic CuD or CuO Treatment
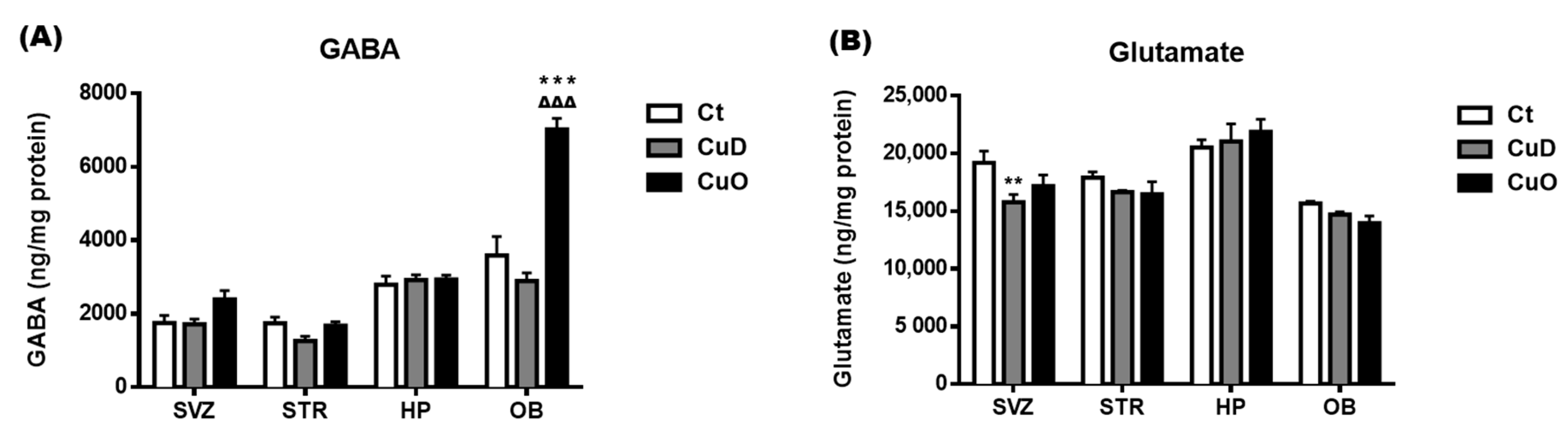
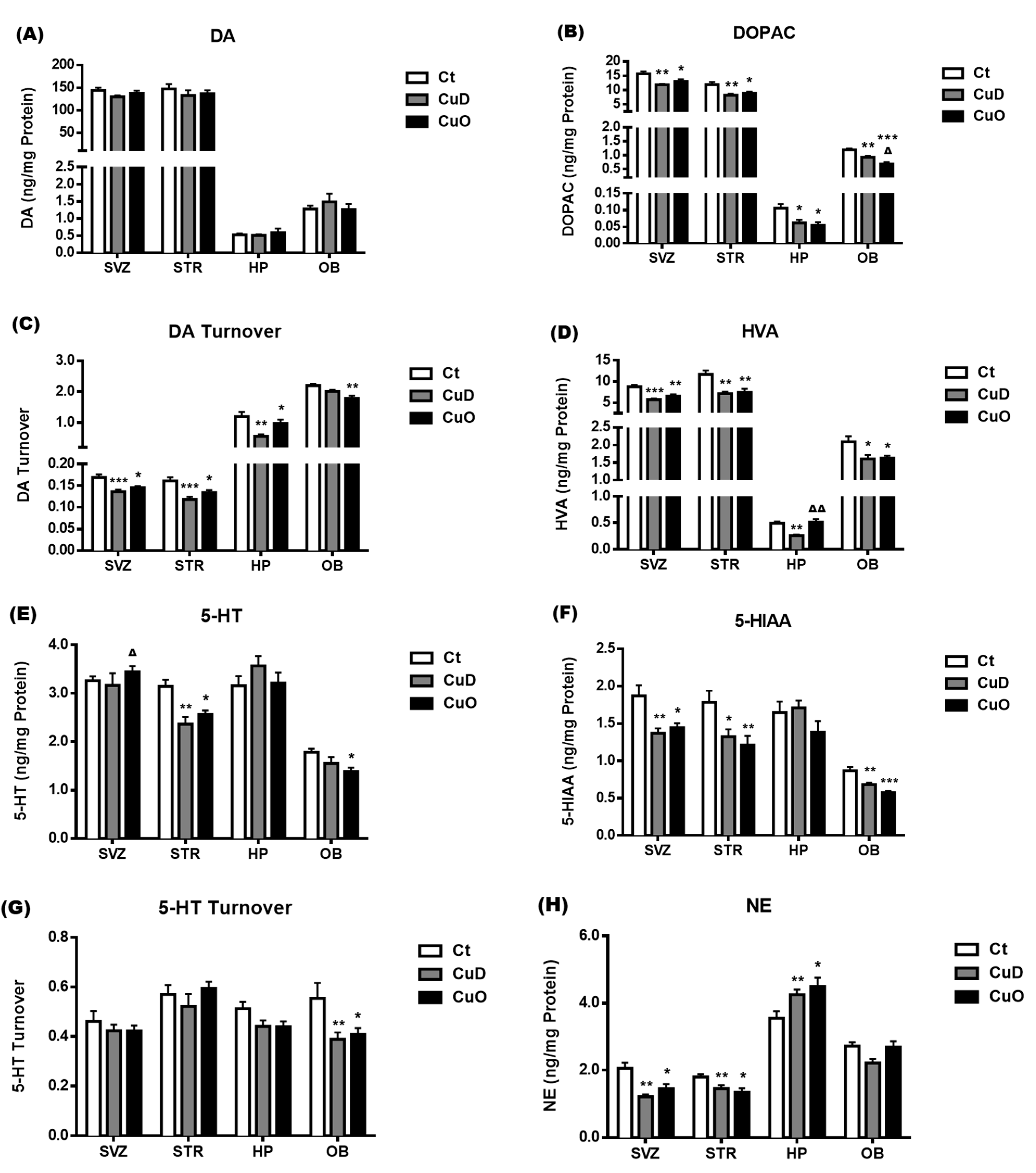
3.5. Neurochemical Imbalance Following Systemic CuD or CuO Treatment
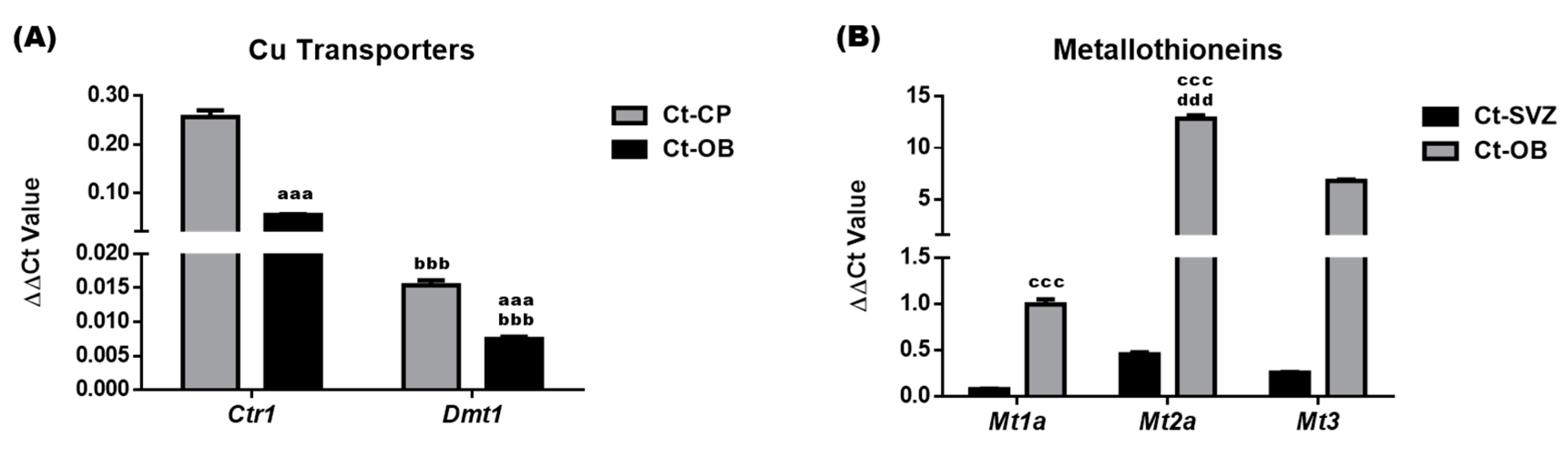
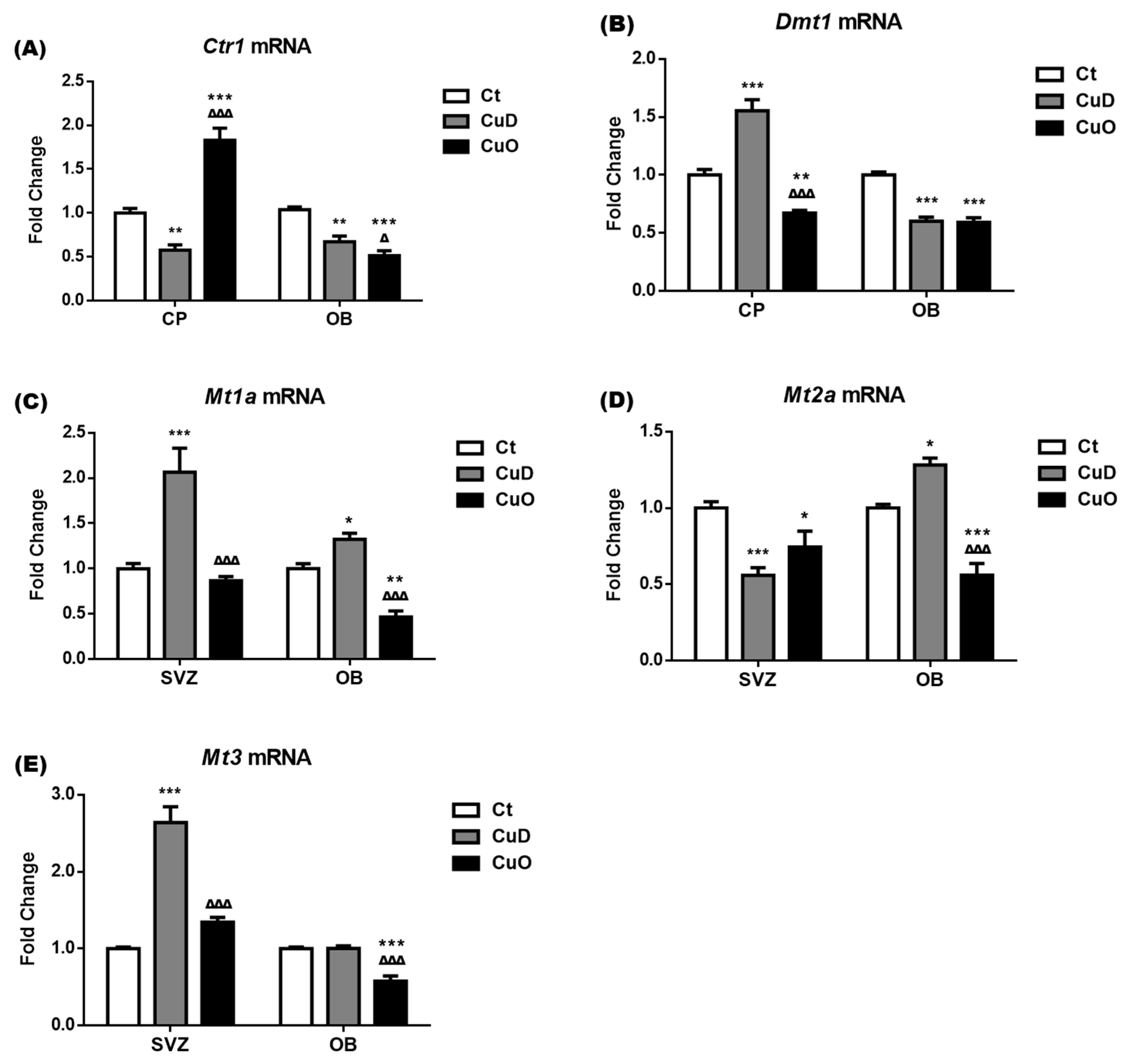
3.6. Altered Expression of Cu Transport-Associated mRNA Following Systemic CuD or CuO Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lorraine, G.; Christine, K.; Harry, J.M. Iron and copper in fetal development. Semin. Cell Dev. Biol. 2011, 22, 637–644. [Google Scholar] [CrossRef]
- Turski, M.L.; Thiele, D.J. New roles for copper metabolism in cell proliferation, signaling, and disease. J. Biol. Chem. 2009, 284, 717–721. [Google Scholar] [CrossRef] [Green Version]
- Uriu-Adams, J.; Scherr, R.; Lanoue, L.; Keen, C.L. Influence of copper on early development: Prenatal and postnatal considerations. Biofactors 2010, 36, 136–152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xu, L.; Wu, J.J.; Wang, W.M.; Mei, J.; Ma, X.F.; Liu, J.X. Transcriptional responses and mechanisms of copper-induced dysfunctional locomotor behavior in zebrafish embryos. Toxicol. Sci. 2015, 148, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Joseph, R.P.; Bruce, B. Dietary copper deficiency alters protein levels of rat dopamine β-monooxygenase and tyrosine monooxygenase. Exp. Biol. Med. 2001, 226, 199–207. [Google Scholar] [CrossRef]
- Scheiber, I.F.; Mercer, J.F.B.; Dringen, R. Metabolism and functions of copper in brain. Prog. Neurobiol. 2014, 116, 33–57. [Google Scholar] [CrossRef] [PubMed]
- Skjorringe, T.; Moller, L.; Moos, T. Impairment of interrelated iron- and copper homeostatic mechanisms in brain contributes to the pathogenesis of neurodegenerative disorders. Front. Pharmacol. 2012, 3, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Kako, K.; Kashiwabara, S.; Takehara, A.; Inada, A.; Arai, H.; Nakada, K.; Kodama, H.; Hayashi, J.; Baba, T.; et al. Mammalian copper chaperone Cox17p has an essential role in activation of cytochrome c oxidase and embryonic development. Mol. Cell Biol. 2002, 22, 7614–7621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danks, D.M. Disorders of Copper Transport. In The Metabolic and Molecular Bases of Inherited Disease; Scriver, C.L., Beaudet, A.L., Sly, W.S., Valle, D., Eds.; McGraw-Hill: New York, NY, USA, 1995; pp. 2211–2235. [Google Scholar]
- Telianidis, J.; Hung, Y.H.; Materia, S.; Fontaine, S.L. Role of the P-Type ATPases, ATP7A and ATP7B in brain copper homeostasis. Front. Aging Neurosci. 2013, 5, 44. [Google Scholar] [CrossRef] [Green Version]
- Gaggelli, E.; Kozlowski, H.; Valensin, D.; Valensin, G. Copper homeostasis and neurodegenerative disorders (Alzheimer’s, prion, and Parkinson’s diseases and amyotrophic lateral sclerosis). Chem. Rev. 2006, 106, 1995–2044. [Google Scholar] [CrossRef] [PubMed]
- Greenough, M.A.; Camakaris, J.; Bush, A.I. Metal dyshomeostasis and oxidative stress in Alzheimer’s disease. Neurochem. Int. 2013, 62, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.M.; Mercer, J.F.B.; Chen, N.; Double, K.L. Copper dyshomeostasis in Parkinson’s disease: Implications for pathogenesis and indications for novel therapeutics. Clin. Sci. 2016, 130, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Liddell, J.R.; Bush, A.I.; White, A.R. Copper in brain and neurodegeneration. In Encyclopedia of Inorganic and Bioinorganic Chemistry; Cuotta, V., Scott, R.A., Eds.; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar] [CrossRef]
- Rivera-Mancia, S.; Perez-Neri, I.; Rios, C.; Tristan-Lopez, L.; Rivera-Espinosa, L.; Montes, S. The transition metals copper and iron in neurodegenerative diseases. Chem. Biol. Interact. 2010, 186, 184–199. [Google Scholar] [CrossRef]
- Jiang, Y.M.; Zheng, W.; Long, L.L.; Zhao, W.J.; Li, X.R.; Mo, X.A.; Lu, J.P.; Fu, X.; Li, W.M.; Liu, S.T.; et al. Brain magnetic resonance imaging and manganese concentrations in red blood cells of smelting workers: Search for biomarkers of manganese exposure. Neurotoxicology 2007, 28, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Li, G.J.; Zhang, L.L.; Lu, L.; Wu, P.; Zheng, W. Occupational exposure to welding fume among welders: Alterations of manganese, iron, zinc, copper, and lead in body fluids and the oxidative stress status. J. Occup. Environ. Med. 2004, 46, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.X.; Du, X.Q.; Zheng, W. Alteration of saliva and serum concentrations of manganese, copper, zinc, cadmium and lead among career welders. Toxicol. Lett. 2008, 176, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Jiang, W.; Zheng, W. Age-dependent increase of brain copper levels and expressions of copper regulatory proteins in the subventricular zone and choroid plexus. Front. Mol. Neurosci. 2015, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Guilarte, T.R.; McGlothan, J.L.; Degaonkar, M.; Chen, M.K.; Barker, P.B.; Syversen, T.; Schneider, J.S. Evidence for cortical dysfunction and widespread manganese accumulation in the nonhuman primate brain following chronic manganese exposure: A 1H-MRS and MRI study. Toxicol. Sci. 2006, 94, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.C.; Minski, M.J.; Chan, A.W.; Leung, T.K.; Lim, L. Manganese mineral interactions in brain. NeuroToxicology 1999, 20, 433–444. [Google Scholar]
- Zheng, W.; Jiang, Y.M.; Zhang, Y.S.; Jiang, W.; Wang, X.Q.; Cowan, D.M. Chelation therapy of manganese intoxication with para-aminosalicylic acid (PAS) in Sprague-Dawley rats. Neurotoxicology 2009, 30, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Pushkar, Y.; Robison, G.A.; Sullivan, G.; Fu, X.; Kohne, M.; Jiang, W.; Rohr, S.; Lai, B.; Marcus, M.A.; Zakharova, T.; et al. Aging results in copper accumulations in subventricular astrocytes. Aging Cell 2013, 12, 823–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.; O’Neal, S.; Hong, L.; Jiang, W.; Zheng, W. Elevated adult neurogenesis in brain subventricular zone following in vivo manganese exposure: Roles of copper and DMT1. Toxicol. Sci. 2015, 143, 482–498. [Google Scholar] [CrossRef] [Green Version]
- El Meskini, R.; Crabtree, K.L.; Cline, L.B.; Mains, R.E.; Eipper, B.A.; Ronnett, G.V. ATP7A (Menkes protein) functions in axonal targeting and synaptogenesis. Mol. Cell Neurosci. 2007, 34, 409–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haremaki, T.; Fraser, S.T.; Kuo, Y.M.; Baron, M.H.; Weinstein, D.C. Vertebrate Ctr1 coordinates morphogenesis and progenitor cell fate and regulates embryonic stem cell differentiation. Proc. Natl. Acad. Sci. USA 2007, 104, 12029–12034. [Google Scholar] [CrossRef] [Green Version]
- Niciu, M.J.; Ma, X.M.; El Meskini, R.; Pachter, J.S.; Mains, R.E.; Eipper, B.A. Altered ATP7A expression and other compensatory responses in a murine model of Menkes disease. Neurobiol. Dis. 2007, 27, 278–291. [Google Scholar] [CrossRef] [Green Version]
- Lledo, P.M.; Alonso, M.; Grubb, M.S. Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci. 2006, 7, 179–193. [Google Scholar] [CrossRef]
- Maki, T.; Liang, A.C.; Miyamoto, N.; Lo, E.H.; Arai, K. Mechanisms of oligodendrocyte regeneration from ventricular-subventricular zone-derived progenitor cells in white matter diseases. Front. Cell Neurosci. 2013, 7, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lois, C.; García-Verdugo, J.M.; Alvarez-Buylla, A. Chain migration of neuronal precursors. Science 1996, 271, 978–981. [Google Scholar] [CrossRef]
- Martino, G.; Pluchino, S. The therapeutic potential of neural stem cells. Nat. Rev. Neurosci. 2006, 7, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Berendse, H.; Booij, J.; Francot, C.M.; Bergmans, P.L.; Hijman, R.; Stoof, J.C.; Wolters, J.C. Subclinical dopaminergic dysfunction in asymptomatic Parkinson’s disease patients’ relatives with a decreased sense of smell. Ann. Neurol. 2001, 50, 34–41. [Google Scholar] [CrossRef]
- Langston, J. The Parkinson’s complex: Parkinsonism is just the tip of the iceberg. Ann. Neurol. 2006, 59, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, E.; Baker, K.; Lyons, K.; Koller, W. Abnormal performance on the PD test battery by asymptomatic first-degree relatives. Neurology 1999, 52, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Ponsen, M.; Stoffers, D.; Booij, J.; van Eck-Smit, B.; Wolters, E.; Berendse, H. Idiopathic hyposmia as a preclinical sign of Parkinson’s disease. Ann. Neurol. 2004, 56, 173–181. [Google Scholar] [CrossRef]
- Ross, G.W.; Petrovitch, H.; Abbott, R.D.; Tanner, C.M.; Popper, J.; Masaki, K.; Launer, L.; White, L.R. Association of olfactory dysfunction with risk for future Parkinson’s disease. Ann. Neurol. 2008, 3, 167–173. [Google Scholar] [CrossRef]
- Chen LWang, X.; Doty, R.L.; Cao, S.; Yang, J.; Sun, F.; Yan, X. Olfactory impairment in Wilson’s disease. Brain Behav. 2021, 11, e02022. [Google Scholar] [CrossRef]
- Mueller, A.; Reuner, U.; Landis, B.; Kitzler, H.; Reichmann, H.; Hummel, T. Extrapyramidal symptoms in Wilson’s disease are associated with olfactory dysfunction. Mov. Disord. 2006, 21, 1311–1316. [Google Scholar] [CrossRef]
- Zheng, W.; Monnot, A.D. Regulation of brain iron and copper homeostasis by brain barrier systems: Implication in neurodegenerative diseases. Pharmacol. Ther. 2012, 133, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Chen, J.Y.; Zheng, W. Relative contribution of CTR1 and DMT1 in copper transport by the blood-CSF barrier: Implication in manganese-induced neurotoxicity. Toxicol. Appl. Pharmacol. 2012, 260, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Zhang, Y.S.; Jiang, W.; Monnot, A.D.; Bates, C.A.; Zheng, W. Regulation of copper transport crossing brain barrier systems by Cu-ATPases: Effect of manganese exposure. Toxicol. Sci. 2014, 139, 432–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, J.; Aschner, M.; Zatta, P.; Vasak, M. Roles of the metallothionein family of proteins in the central nervous system. Brain Res. Bull. 2001, 55, 133–145. [Google Scholar] [CrossRef]
- Kuo, Y.M.; Gybina, A.A.; Pyatskowit, J.W.; Gitschier, J.; Prohaska, J.R. Copper transporter protein (Ctr1) levels in mice are tissue specific and dependent on copper status. J. Nutr. 2006, 136, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Gybina, A.A.; Tkac, I.; Prohaska, J.R. Copper deficiency alters the neurochemical profile of developing rat brain. Nutr. Neurosci. 2009, 12, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Liao, Z.; Medeiros, D.M.; McCune, S.A.; Prochaska, L.J. Cardiac levels of fibronectin, laminin, isomyosins, and cytochrome c oxidase of weanling rats are more vulnerable to copper deficiency than those of postweanling rats. J. Nutr. Biochem. 1995, 6, 385–391. [Google Scholar] [CrossRef]
- Mizuno, S.; Yasuo, M.; Bogaard, H.J.; Kraskauskas, D.; Alhussaini, A.; Gomez-Arroyo, J.; Farkas, D.; Farkas, L.; Voelkel, N.F. Copper deficiency induced emphysema is associated with focal adhesion kinase inactivation. PLoS ONE 2012, 7, e30678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, K.T.; Proshaska, J.R. Copper deficiency in rodents alters dopamine beta-mono-oxygenase activity, mRNA and protein level. Br. J. Nutr. 2009, 102, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, C.A., Jr.; Hazelrig, J.B. Copper deficiency and copper toxicity in the rat. Am. J. Physiol. 1968, 215, 334–338. [Google Scholar] [CrossRef]
- Pal, A.; Vasishta, R.K.; Prasad, R. Hepatic and hippocampus iron status is not altered in response to increased serum ceruloplasmin and serum “free” copper in Wistar rat model for non-Wilsonian brain copper toxicosis. Biol. Trace Elem. Res. 2013, 154, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Badyal, R.K.; Vasishta, R.K.; Attri, S.V.; Thapa, B.R.; Prasad, R. Biochemical, histological, and memory impairment effects of chronic copper toxicity: A model for non-Wilsonian brain copper toxicosis in Wistar rat. Biol. Trace Elem. Res. 2013, 153, 257–268. [Google Scholar] [CrossRef]
- Pal, A.; Prasad, R. Regional distribution of copper, zinc and iron in brain of Wistar rat model for non-Wilsonian brain copper toxicosis. Indian J. Clin. Biochem. 2016, 31, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Zhou, Z.; Chen, T.; Zhang, J.; McCalin, C.J. Copper deficiency exacerbates bile duct ligation-induced liver injury and fibrosis in rats. J. Pharmacol. Exp. Ther. 2011, 339, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Choi, Y.; Ahn, M.; Jung, K.; Shin, T. Olfactory dysfunction in autoimmune central nervous system neuroinflammation. Mol. Neurobiol. 2018, 55, 8499–8508. [Google Scholar] [CrossRef]
- Lehmkuhl, A.M.; Dirr, E.R.; Fleming, S.M. Olfactory assays for mouse models of neurodegenerative disease. J. Vis. Exp. 2014, 25, e51804. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Crawley, J.N. Simple behavioral assessment of mouse olfaction. Curr. Protoc. Neurosci. 2009, 8, 24. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.S.; Zheng, W. Copper transport to the brain by the blood-brain barrier and blood-CSF barrier. Brain Res. 2009, 1248, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coggeshall, R.E.; Lekan, H.A. Methods for determining numbers of cells and synpses: A case for more uniform standards of review. J. Comp. Neurol. 1996, 364, 6–15. [Google Scholar] [CrossRef]
- Adamson, S.F.; Shen, S.; Jiang, W.; Lai, V.; Wang, X.; Shannahan, J.H.; Cannon, J.; Chen, J.H.; Zheng, W. Subchronic manganese exposure impairs neurogenesis in the adult rat hippocampus. Toxicol. Sci. 2018, 163, 592–608. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, S.L.; Hong, L.; Fu, S.; Jiang, W.; Jones, A.; Nie, L.H.; Zheng, W. Manganese accumulation in bone following chronic exposure in rats: Steady-state concentration and half-life in bone. Toxicol. Lett. 2014, 229, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Agim, Z.S.; Cannon, J.R. Alteration in the nigrostriatal dopamine system after acute systemic PhIP exposure. Toxicol. Lett. 2018, 287, 31–41. [Google Scholar] [CrossRef]
- Fu, S.; Jiang, W.; Gao, X.; Zeng, A.; Cholger, D.; Cannon, J.; Chen, J.; Zheng, W. Aberrant adult neurogenesis in the subventricular zone-rostral migratory stream-olfactory bulb system following subchronic manganese exposure. Toxicol. Sci. 2016, 150, 347–368. [Google Scholar] [CrossRef] [Green Version]
- Nagayama, S.; Homma, R.; Imamura, F. Neuronal organization of olfactory bulb circuits. Front. Neural Circuits 2014, 8, 98. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Buylla, A.; Lim, D.A. For the long run: Maintaining germinal niches in the adult brain. Neuron 2004, 41, 683–686. [Google Scholar] [CrossRef] [Green Version]
- Ghashghaei, H.T.; Lai, C.; Anton, E.S. Neuronal migration in the adult brain: Are we there yet? Nat. Rev. Neurosci. 2007, 8, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Lie, D.C.; Song, H.; Colamarino, S.A.; Ming, G.L.; Gage, F.H. Neurogenesis in the adult brain: New strategies for central nervous system diseases. Ann. Rev. Pharmacol. Toxicol. 2004, 44, 399–421. [Google Scholar] [CrossRef] [Green Version]
- Kawano, T.; Margolis, F.L. Transsynaptic regulation of olfactory bulb catecholamines in mice and rats. J. Neurochem. 1982, 39, 342–348. [Google Scholar] [CrossRef]
- Huisman, E.; Uylings, H.B.; Hoogland, P.V. A 100% increase of dopaminergic cells in the olfactory bulb may explain hyposmia in Parkinson’s disease. Mov. Disord. 2004, 19, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Marin, C.; Vilas, D.; Langdon, C.; Alobid, I.; López-Chacón, M.; Haehner, A.; Hummel, T.; Mullol, J. Olfactory dysfunction in neurodegenerative disease. Curr. Allergy Asthma Rep. 2018, 18, 42. [Google Scholar] [CrossRef]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; Flores-Cuadrado, A.; Rioja-Corroto, E.; Gonzalez-Rodriguez, M.; Villar-Conde, S.; Astillero-Lopez, V.; Cabello-de la Rosa, J.P.; Gallardo-Alcañiz, M.J.; Vaamonde-Gamo, J.; et al. The human olfactory system in two proteinopathies: Alzheimer’s and Parkinson’s diseases. Transl. Neurodegener. 2020, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Fullard, M.E.; Morley, J.F.; Duda, J.E. Olfactory dysfunction as an early biomarker in Parkinson’s disease. Neurosci. Bull. 2017, 33, 515–525. [Google Scholar] [CrossRef]
- Baldwin, D.H.; Sandahl, J.F.; Labenia, J.S.; Scholz, N.L. Sublethal effects of copper on coho salmon: Impacts on nonoverlapping receptor pathways in the peripheral olfactory nervous system. Environ. Toxicol. Chem. 2003, 22, 2266–2274. [Google Scholar] [CrossRef]
- Julliard, A.K.; Saucier, D.; Astic, L. Time-course of apoptosis in the olfactory epithelium of rainbow trout exposed to low copper level. Tissue Cell 1996, 28, 367–377. [Google Scholar] [CrossRef]
- Mclntyre, J.K.; Baldwin, D.H.; Beauchamp, D.A.; Scholz, N.L. Low-level copper exposures increase visibility and vulnerability of juvenile coho salmon to cutthroat trout predators. Ecol. Appl. 2012, 22, 1460–1471. [Google Scholar] [CrossRef] [PubMed]
- Dew, W.A.; Wood, C.M.; Pyle, G.G. Effects of continuous copper exposure and calcium on the olfactory response of fathead minnows. Environ. Sci. Technol. 2012, 46, 9019–9026. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J.; Pierre, T.S.; Minnich, J.L.; Barbeau, A. Determination of Na+, K+, Mg2+, Cu2+, Zn2+, and Mn2+ in rat brain regions. Can. J. Biochem. 1973, 51, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Cherian, M.G. Regional distribution of metallothionein, zinc, and copper in the brain of different strains of rats. Biol Trace Elem. Res. 1999, 69, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Friedman, B.; Price, J.L. Fiber systems in the olfactory bulb and cortex: A study in adult and developing rats, using the timm method with the light and electron microscope. J. Comp. Neurol. 1984, 223, 88–109. [Google Scholar] [CrossRef]
- Jo, S.M.; Won, M.H.; Cole, T.B.; Jensen, M.S.; Palmiter, R.D.; Danscher, G. Zinc-enriched (ZEN) terminals in mouse olfactory bulb. Brain Res. 2000, 26, 227–236. [Google Scholar] [CrossRef]
- Pérez-Clausell, J.; Danscher, G. Intravesicular localization of zinc in rat telencephalic boutons. A histochemical study. Brain Res. 1985, 24, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Vergara, C.; Wolff, D. Divalent cations as modulators of neuronal excitability: Emphasis on copper and zinc. Biol. Res. 2006, 39, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Horning, M.S.; Trombley, P.Q. Zinc and copper influence excitability of rat olfactory bulb neurons by multiple mechanisms. J. Neurophysiol. 2001, 86, 1652–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trombley, P.Q.; Horning, M.S.; Blakemore, L.J. Interactions between carnosine and zinc and copper: Implications for neuromodulation and neuroprotection. Biochemistry 2000, 65, 807–816. [Google Scholar]
- Trombley, P.Q.; Shepherd, G.M. Differential modulation by zinc and copper of amino acid receptors from rat olfactory bulb neurons. J. Neurophysiol. 1996, 76, 2536–2546. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.Y.; Heffern, K.; Cheresh, J.; Gallagher, E.P. Differential copper-induced death and regeneration of olfactory sensory neuron populations and neurobehavioral function in larval zebrafish. Neurotoxicology 2018, 59, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Saari, J.T. Increased type I collagen content and DNA binding activity of a single-stranded, cytosine-rich sequence in the high-salt buffer protein extract of the copper-deficient rat heart. J. Nutr. Biochem. 2004, 15, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Prohaska, J.R.; Bailey, W.R. Regional specificity in alterations of rat brain copper and catecholamines following perinatal copper deficiency. J. Neurochem. 1994, 63, 1551–1557. [Google Scholar] [CrossRef]
- D’Ambrosi, N.; Rossi, L. Copper at synapse: Release, binding and modulating of neurotransmission. Neurochem. Int. 2015, 90, 36–45. [Google Scholar] [CrossRef]
- Gaier, E.D.; Eipper, B.A.; Mains, R.E. Copper signaling in the mammalian nervous system: Synaptic effects. J. Neurosci. Res. 2013, 91, 2–19. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.S.; Zoriy, M.V.; Pickhardt, C.; Palomero-Gallagher, N.; Zilles, K. Imaging of copper, zinc, and other elements in thin section of human brain samples (hippocampus) by laser ablation inductively coupled plasma mass spectrometry. Anal. Chem. 2005, 77, 3208–3216. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.M.; Hare, D.J.; Cottam, V.; Chen, N.; Hilgers, L.; Halliday, G.; Mercer, J.F.; Double, K.L. Localization of copper and copper transporters in the human brain. Metallomics 2013, 5, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Dobrowolska, J.; Dehnhardt, M.; Matusch, A.; Zoriy, M.; Palomero-Gallagher, N.; Koscielniak, P.; Zilles, K.; Becker, J.S. Quantitative imaging of zinc, copper and lead in three distinct regions of the human brain by laser ablation inductively coupled plasma mass spectrometry. Talanta 2008, 74, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Lech, T.; Sadlik, J.K. Copper concentration in body tissues and fluids in normal subjects of southern Poland. Biol. Trace Elem. Res. 2007, 118, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Ramos, P.; Santos, A.; Pinto, N.R.; Mendes, R.; Magalhães, T.; Almeida, A. Anatomical region differences and age-related changes in copper, zinc, and manganese levels in the human brain. Biol. Trace Elem. Res. 2014, 161, 190–201. [Google Scholar] [CrossRef]
- Jackson, B.; Harper, S.; Smith, L.; Flinn, J. Elemental mapping and quantitative analysis of Cu, Zn, and Fe in rat brain sections by laser ablation ICP-MS. Anal. Bioanal. Chem. 2006, 384, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Olusola, A.O.; Obodozie, O.O.; Nssien, M.; Adaramoye, A.; Adesanoye, O.; Odama, L.E.; Emerole, G.O. Concentrations of copper, iron, and zinc in the major organs of the wistar albino and wild black rats: A comparative study. Biol. Trace Elem. Res. 2004, 98, 265–274. [Google Scholar] [CrossRef]
- Waggoner, D.J.; Drisaldi, B.; Bartnikas, T.B.; Casareno, R.L.; Prohaska, J.R.; Gitlin, J.D.; Harris, D.A. Brain copper content and cuproenzyme activity do not vary with prion protein expression level. J. Biol. Chem. 2000, 275, 7455–7458. [Google Scholar] [CrossRef] [Green Version]
- Palm, R.; Wahlström, G.; Hallmans, G. Age related changes in weight and the concentrations of zinc and copper in the brain of the adult rat. Lab. Anim. 1990, 24, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Serpa, R.F.; de Jesus, E.F.; Anjos, M.J.; de Oliverira, L.F.; Marins, L.A.; do Carmo, M.G.; Correa, J.D.; Rocha, M.S.; Lopes, R.T.; Martinez, A.M. Topographic trace-elemental analysis in the brain of Wistar rats by X-ray microfluorescence with synchrontron radiation. Anal. Sci. 2008, 24, 839–842. [Google Scholar] [CrossRef] [Green Version]
- Tarohda, T.; Yamamoto, M.; Amamo, R. Regional distribution of manganese, iron, copper, and zine in the rat brain during development. Anal. Bioanal. Chem. 2004, 380, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Becker, J.S.; Wu, Q.; Oliverira, M.F.; Bozza, F.A.; Schwager, A.L.; Hoffman, J.M.; Morton, K.A. Bioimaging of copper alterations in the aging mouse brain by autoradiography, laser ablation inductively coupled plasma mass spectrometry and immunohistochemistry. Metallomics 2010, 2, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, W.J.; Allen, N. Determination of Cu, Mn, Fe, and Ca in six regions of normal human brain, by atomic absorption spectroscopy. Clin. Chem. 1981, 27, 562–564. [Google Scholar] [CrossRef]
- Propescu, B.F.; George, M.J.; Bergmann, U.; Garachtchenko, A.V.; Kelly, M.E.; McCrea, R.P.; Luning, K.; Devon, R.M.; George, G.N.; Hanson, A.D.; et al. Mapping metals in Parkinson’s and normal brain using rapid-scanning x-ray fluorescence. Phys. Med. Biol. 2009, 54, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Warren, P.J.; Earl, C.J.; Thompson, R.H. The distribution of copper in human brain. Brain 1960, 83, 709–717. [Google Scholar] [CrossRef]
- Sullivan, B.; Robison, G.; Osborn, J.; Kay, M.; Thompson, P.; Davis, K.; Zakharova, T.; Antipova, O.; Pushkar, Y. On the nature of the Cu-rich aggregates in brain astrocytes. Redox Biol. 2017, 11, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, B.; Robison, G.; Pushkar, Y.; Young, J.K.; Manaye, K.F. Copper accumulation in rodent brain astrocytes: A species difference. J. Trace Elem. Med. Biol. 2017, 39, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, Y.; Evans, P.; Dodd, G.H. Characterization of the total lipid and fatty acid composition of rat olfactory mucosa. J. Lipid Res. 1989, 30, 877–884. [Google Scholar] [CrossRef]
- Sebio, R.M.; Ferrarotti, N.; Lairion, F.; Magriñá, C.S.; Fuda, J.; Torti, H.; Boveris, A.; Repetto, M.G. Brain oxidative stress in rat with chronic iron or copper overload. J. Inorg. Biochem. 2019, 199, 110799. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Gene | Primer Sequence |
|---|---|---|
| Reference Gene | Actb | Forward: 5′-AGCCATGTACGTAGCCATCC-3′ |
| Reverse: 5′-CTCTCAGCTGTGGTGGTGAA-3′ | ||
| Cu Transporter | Ctr1 | Forward: 5′-TCGGCCTCACACTCCCACGA-3′ |
| Reverse: 5′-CGAAGCAGACCCTCTCGGGC-3′ | ||
| Dmt1 | Forward: 5′-TCGCAGGCGGCATCTTGGTC-3′ | |
| Reverse: 5′-TACCGAGCGCCCACAGTCCA-3′ | ||
| Cu Binding Proteins | Mt1a | Forward: 5′-GCCTTCTTGTCGCTTACACC-3′ |
| Reverse: 5′-AGGAGCAGCAGCTCTTCTTG-3′ | ||
| Mt2a | Forward: 5′-ACAGATGGATCCTGCTCCTG-3′ | |
| Reverse: 5′-GAGAACCGGTCAGGGTTGTA-3′ | ||
| Mt3 | Forward: 5′-CCCTGCAGGATGTGAGAAGT-3′ | |
| Reverse: 5′-TTTGCTGTGCATGGGATTTA-3′ | ||
| Neuronal Markers | Gfap | Forward: 5′-TAGCATAAGTGGAGAGGGAA-3′ |
| Reverse: 5′-GGATTCAGAGCCAAGTGTAA-3′ | ||
| Nestin | Forward: 5′-ATGAGGGGCAAATCTGGGAA-3′ | |
| Reverse: 5′-CCAGGTGGCCTTCTGTAGAA-3′ | ||
| Dcx | Forward: 5′-ACTGAATGCTTAGGGGCCTT-3′ | |
| Reverse: 5′-CTGACTTGCCACTCTCCTGA-3′ | ||
| NeuN | Forward: 5′-TTCCCACCACTCTCTTGTCC-3′ | |
| Reverse: 5′-GCAGCCGCATAGACTCTACC-3′ | ||
| Th | Forward: 5′-CAGGGCTGCTGTCTTCCTAC-3′ | |
| Reverse: 5′-GGGCTGTCCAGTACGTCAAT-3′ | ||
| Gad67 | Forward: 5′-CACAAACTCAGCGGCATAGA-3′ | |
| Reverse: 5′-CTGGAAGAGGTAGCCTGCAC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamson, S.X.-F.; Zheng, W.; Agim, Z.S.; Du, S.; Fleming, S.; Shannahan, J.; Cannon, J. Systemic Copper Disorders Influence the Olfactory Function in Adult Rats: Roles of Altered Adult Neurogenesis and Neurochemical Imbalance. Biomolecules 2021, 11, 1315. https://doi.org/10.3390/biom11091315
Adamson SX-F, Zheng W, Agim ZS, Du S, Fleming S, Shannahan J, Cannon J. Systemic Copper Disorders Influence the Olfactory Function in Adult Rats: Roles of Altered Adult Neurogenesis and Neurochemical Imbalance. Biomolecules. 2021; 11(9):1315. https://doi.org/10.3390/biom11091315
Chicago/Turabian StyleAdamson, Sherleen Xue-Fu, Wei Zheng, Zeynep Sena Agim, Sarah Du, Sheila Fleming, Jonathan Shannahan, and Jason Cannon. 2021. "Systemic Copper Disorders Influence the Olfactory Function in Adult Rats: Roles of Altered Adult Neurogenesis and Neurochemical Imbalance" Biomolecules 11, no. 9: 1315. https://doi.org/10.3390/biom11091315