
CSN5A Subunit of COP9 Signalosome Is Required for Resetting Transcriptional Stress Memory after Recurrent Heat Stress in Arabidopsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Heat Treatments
2.3. Chromatin Immunoprecipitation
2.4. qRT-PCR Analysis
3. Results
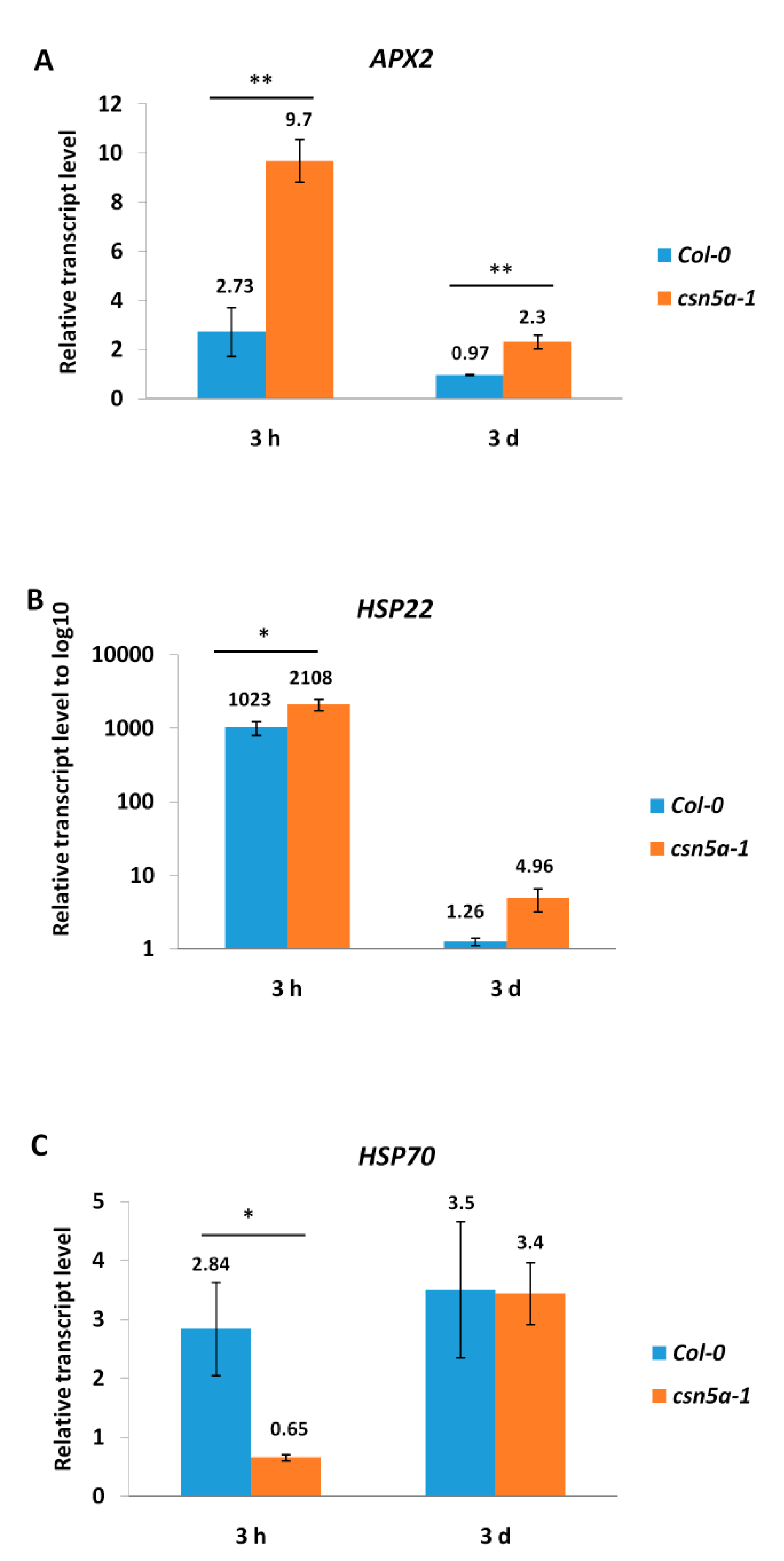
3.1. CSN5A Regulates the Expression of Heat Stress Memory Genes APX2 and HSP22 But Not HSP70 after Recurrent Heat Stress
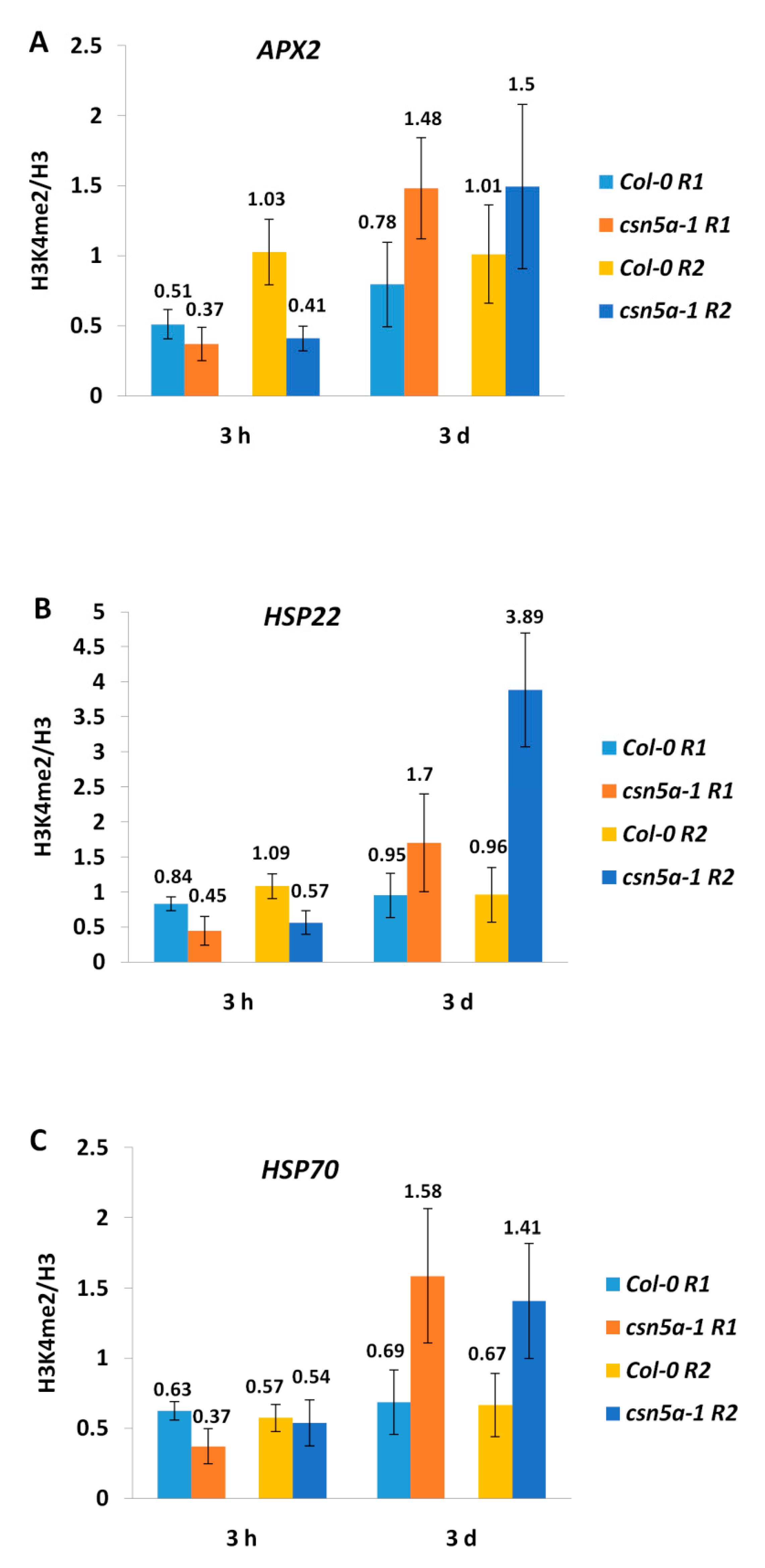
3.2. Expression of Heat Stress Memory Genes Is Not Associated with H3K4 Methylation
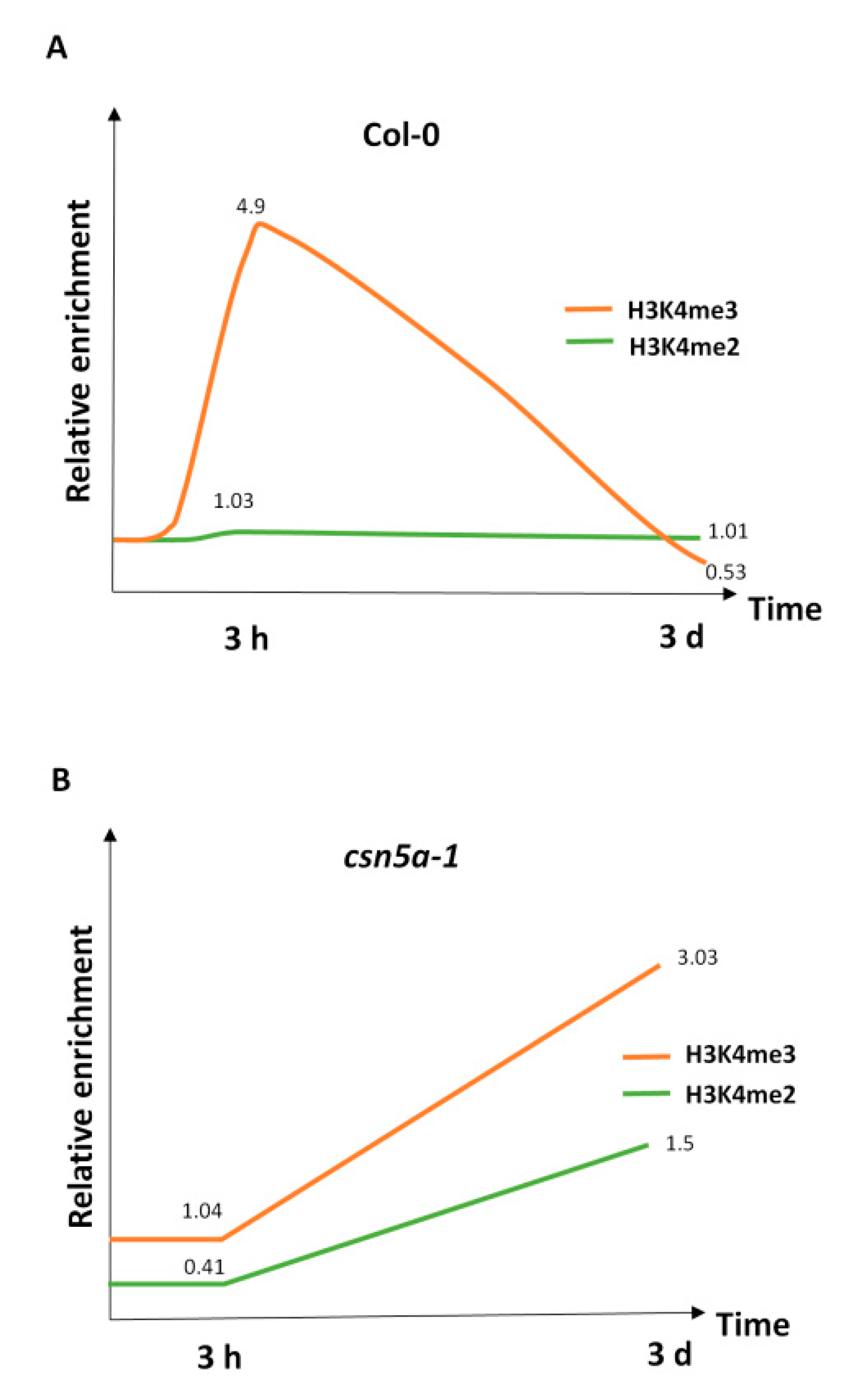
3.3. CSN5A Is Required for Deposition as Well as Restoration of the H3K4me3 Level 3 h and 3 d after Recurrent Heat Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deng, X.W.; Dubiel, W.G.; Wei, N.; Hofmann, K.; Mundt, K.; Colicelli, L.; Kato, J.; Naumann, M.; Segal, D.; Seeger, M. Unified nomenclature for the COP9 signalosome and its subunits: An essential regulator of development. Trends Genet. 2000, 16, 202–203. [Google Scholar] [CrossRef]
- Chamovitz, D.A.; Wei, N.; Osterlund, M.T.; von Arnim, A.G.; Staub, J.M.; Matsui, M.; Deng, X.W. The COP9 complex, a novel multisubunit nuclear regulator involved in light control of a plant developmental switch. Cell 1996, 86, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Wei, N.; Chamovitz, D.A.; Deng, X.W. Arabidopsis Cop9 Is a Component of a Novel Signaling Complex Mediating Light Control of Development. Cell 1994, 78, 117–124. [Google Scholar] [CrossRef]
- Cope, G.A.; Suh, G.S.B.; Aravind, L.; Schwarz, S.E.; Zipursky, S.L.; Koonin, E.V.; Deshaies, R.J. Role of predicted metalloprotease motif of Jab1/Csn5 in cleavage of Nedd8 from Cul1. Science 2002, 298, 608–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Chamovitz, D.A. Role of Cop9 Signalosome Subunits in the Environmental and Hormonal Balance of Plant. Biomolecules 2019, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Chamovitz, D.A. Revisiting the COP9 signalosome as a transcriptional regulator. EMBO Rep. 2009, 10, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Emberley, E.D.; Mosadeghi, R.; Deshaies, R.J. Deconjugation of Nedd8 from Cul1 is directly regulated by Skp1-F-box and substrate, and the COP9 signalosome inhibits deneddylated SCF by a noncatalyticmechanism. J. Biol. Chem. 2012, 287, 29679–29689. [Google Scholar] [CrossRef] [Green Version]
- Fischer, E.S.; Scrima, A.; Böhm, K.; Matsumoto, S.; Lingaraju, G.M.; Faty, M.; Yasuda, T.; Cavadini, S.; Wakasugi, M.; Hanaoka, F.; et al. The molecular basis of CRL4DDB2/CSA ubiquitin ligase architecture, targeting, and activation. Cell 2011, 147, 1024–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuller, T.; Diament, A.; Yahalom, A.; Zemach, A.; Atar, S.; Chamovitz, D.A. The COP9 signalosome influences the epigenetic landscape of Arabidopsis thaliana. Bioinformatics 2019, 35, 2718–2723. [Google Scholar] [CrossRef]
- Yahalom, A.; Kim, T.H.; Roy, B.; Singer, R.; Von Arnim, A.G.; Chamovitz, D.A. Arabidopsis eIF3e is regulated by the COP9 signalosome and has an impact on development and protein translation. Plant J. 2008, 53, 300–311. [Google Scholar] [CrossRef]
- Dohmann, E.M.N.; Levesque, M.P.; De Veylder, L.; Reichardt, I.; Juergens, G.; Schmid, M.; Schwechheimer, C. The Arabidopsis COP9 signalosome is essential for G2 phase progression and genomic stability. Development 2008, 135, 2013–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oron, E.; Tuller, T.; Li, L.; Rozovsky, N.; Yekutieli, D.; Rencus-Lazar, S.; Segal, D.; Chor, B.; Edgar, B.A.; Chamovitz, D.A. Genomic analysis of COP9 signalosome function in Drosophila melanogaster reveals a role in temporal regulation of gene expression. Mol. Syst. Biol. 2007, 3, 108. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.; Atar, S.; Atias, O.; Oron, E.; Segal, D.; Hirsch, J.A.; Tuller, T.; Orian, A.; Chamovitz, D.A. Drosophila COP9 signalosome subunit 7 interacts with multiple genomic loci to regulate development. Nucleic Acids Res. 2014, 42, 9761–9770. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Feng, S.; Nakayama, N.; Crosby, W.L.; Irish, V.; Deng, X.W.; Wei, N. The COP9 Signalosome Interacts with SCFUFO and Participates in Arabidopsis Flower Development. Plant Cell 2003, 15, 1071–1082. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, X.; Giroux, M.J.; Huang, L. A wheat COP9 subunit 5-like gene is negatively involved in host response to leaf rust. Mol. Plant Pathol. 2017, 18, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Jaskiewicz, M.; Conrath, U.; Peterhänsel, C. Chromatin modification acts as a memory for systemic acquired resistance in the plant stress response. EMBO Rep. 2011, 12, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Sani, E.; Herzyk, P.; Perrella, G.; Colot, V.; Amtmann, A. Hyperosmotic priming of Arabidopsis seedlings establishes a long-term somatic memory accompanied by specific changes of the epigenome. Genome Biol. 2013, 14, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Bernatavichute, Y.V.; Cokus, S.; Pellegrini, M.; Jacobsen, S.E. Genome-wide analysis of mono-, di- and trimethylation of histone H3 lysine 4 in Arabidopsis thaliana. Genome Biol. 2009, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Ndamukong, I.; Xu, Z.; Lapko, H.; Fromm, M.; Avramova, Z. ATX1-generated H3K4me3 is required for efficient elongation of transcription, not initiation, at ATX1-regulated genes. PLoS Genet. 2012, 8, e1003111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J. 2016, 35, 162–175.21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.R.; Bäurle, I. Arabidopsis miR156 regulates tolerance to recurring environmental stress through SPL transcription factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef] [Green Version]
- Brzezinka, K.; Altmann, S.; Bäurle, I. BRUSHY1/TONSOKU/MGOUN3 is required for heat stress memory. Plant Cell Environ. 2019, 42, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Yadav, B.S.; Dhanapal, S.; Berliner, M.; Finkelshtein, A.; Chamovitz, D.A. CSN5A Subunit of COP9 Signalosome Temporally Buffers Response to Heat in Arabidopsis. Biomolecules 2019, 9, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohmann, E.M.N.; Kuhnle, C.; Schwechheimer, C. Loss of the constitutive photomorphogenic9 signalosome subunit 5 is sufficient to cause the cop/det/fus mutant phenotype in Arabidopsis. Plant Cell 2005, 17, 1967–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, A.; Alvarez-Venegas, R.; Avramova, Z. An efficient chromatin immunoprecipitation (ChIP) protocol for studying histone modifications in Arabidopsis plants. Nat. Protoc. 2008, 3, 1018. [Google Scholar] [CrossRef] [PubMed]
- Avramova, Z. Transcriptional “memory” of a stress: Transient chromatin and memory (epigenetic) marks at stress-response genes. Plant J. 2015, 83, 149–159. [Google Scholar] [CrossRef] [PubMed]
- D’Urso, A.; Brickner, J.H. Mechanisms of epigenetic memory. Trends Genet. 2014, 30, 230–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzezinka, K.; Altmann, S.; Czesnick, H.; Nicolas, P.; Gorka, M.; Benke, E.; Kabelitz, T.; Jähne, F.; Graf, A.; Kappel, C.; et al. Arabidopsis FORGETTER1 mediates stress-induced chromatin memory through nucleosome remodeling. elife 2016, 5, e17061. [Google Scholar] [CrossRef]
- Liu, H.C.; Lämke, J.; Lin, S.Y.; Hung, M.J.; Liu, K.M.; Charng, Y.Y.; Bäurle, I. Distinct heat shock factors and chromatin modifications mediate the organ-autonomous transcriptional memory of heat stress. Plant J. 2018, 95, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Yekondi, S.; Chen, P.W.; Tsai, C.H.; Yu, C.W.; Wu, K.; Zimmerli, L. Environmental history modulates arabidopsis pattern-triggered immunity in a HISTONE ACETYLTRANSFERASE1-dependent manner. Plant Cell 2014, 26, 2676–2688. [Google Scholar] [CrossRef] [Green Version]
- Zong, W.; Zhong, X.; You, J.; Xiong, L. Genome-wide profiling of histone H3K4-trimethylation and gene expression in rice under drought stress. Plant Mol. Biol. 2013, 81, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Howe, F.S.; Fischl, H.; Murray, S.C.; Mellor, J. Is H3K4me3 instructive for transcription activation? Bioessays 2017, 39, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Hofmann, K.; Von Arnim, A.G.; Chamovitz, D.A. PCI complexes: Pretty complex interactions in diverse signaling pathways. Trends Plant Sci. 2001, 6, 379–386. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.K.; Dhanapal, S.; Finkelshtein, A.; Chamovitz, D.A. CSN5A Subunit of COP9 Signalosome Is Required for Resetting Transcriptional Stress Memory after Recurrent Heat Stress in Arabidopsis. Biomolecules 2021, 11, 668. https://doi.org/10.3390/biom11050668
Singh AK, Dhanapal S, Finkelshtein A, Chamovitz DA. CSN5A Subunit of COP9 Signalosome Is Required for Resetting Transcriptional Stress Memory after Recurrent Heat Stress in Arabidopsis. Biomolecules. 2021; 11(5):668. https://doi.org/10.3390/biom11050668
Chicago/Turabian StyleSingh, Amit Kumar, Shanmuhapreya Dhanapal, Alin Finkelshtein, and Daniel A. Chamovitz. 2021. "CSN5A Subunit of COP9 Signalosome Is Required for Resetting Transcriptional Stress Memory after Recurrent Heat Stress in Arabidopsis" Biomolecules 11, no. 5: 668. https://doi.org/10.3390/biom11050668