
Testing the Resilience, Physiological Plasticity and Mechanisms Underlying Upper Temperature Limits of Antarctic Marine Ectotherms
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collections and Animal Husbandry
2.2. Identification of Anemone Species
2.3. Acclimation Experiments: Temperature Incubations
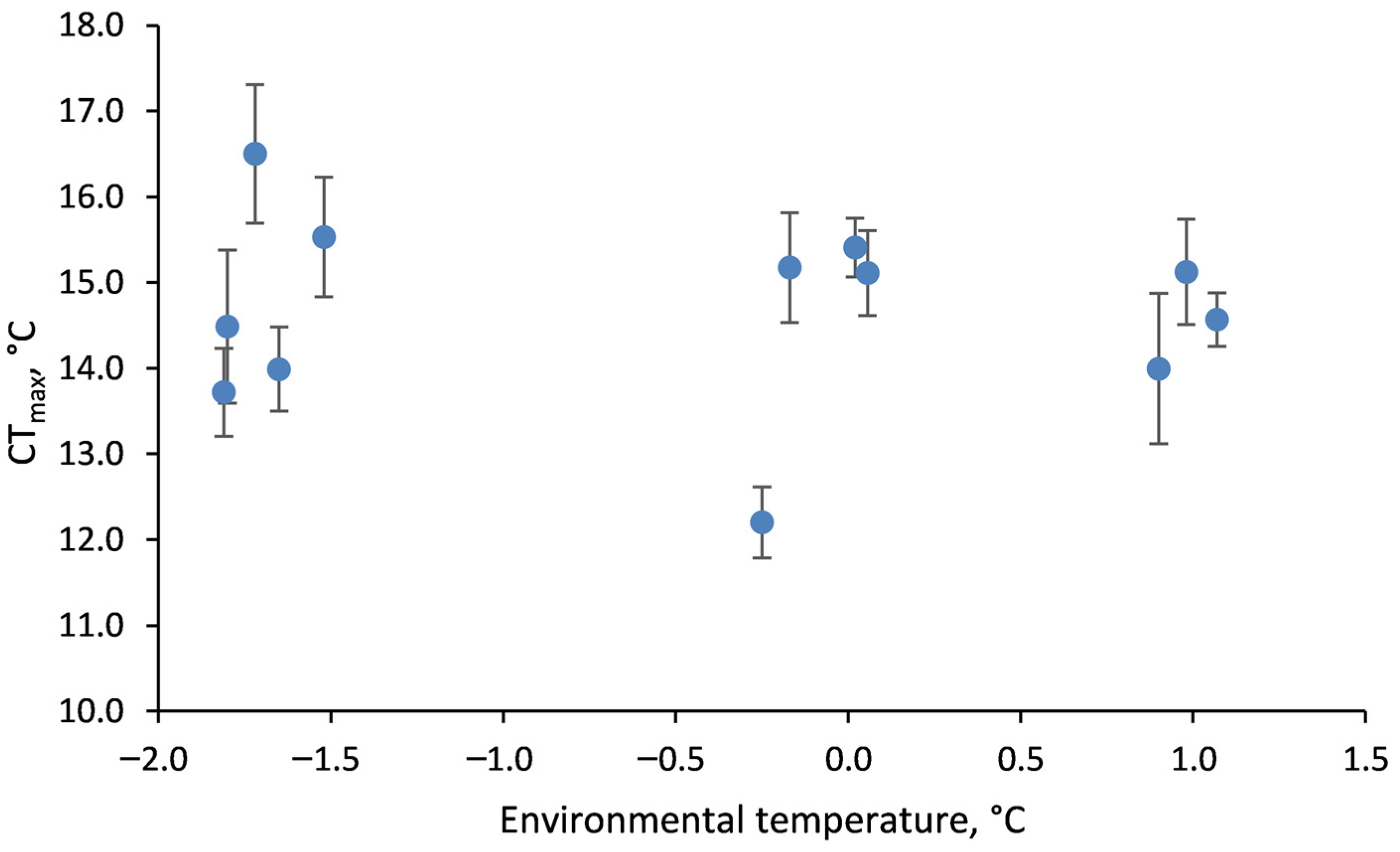
2.4. Field Comparisons
2.5. Temperature–Oxygen Incubations
2.6. Statistical Analysis
3. Results
3.1. Identification of Anemone Species
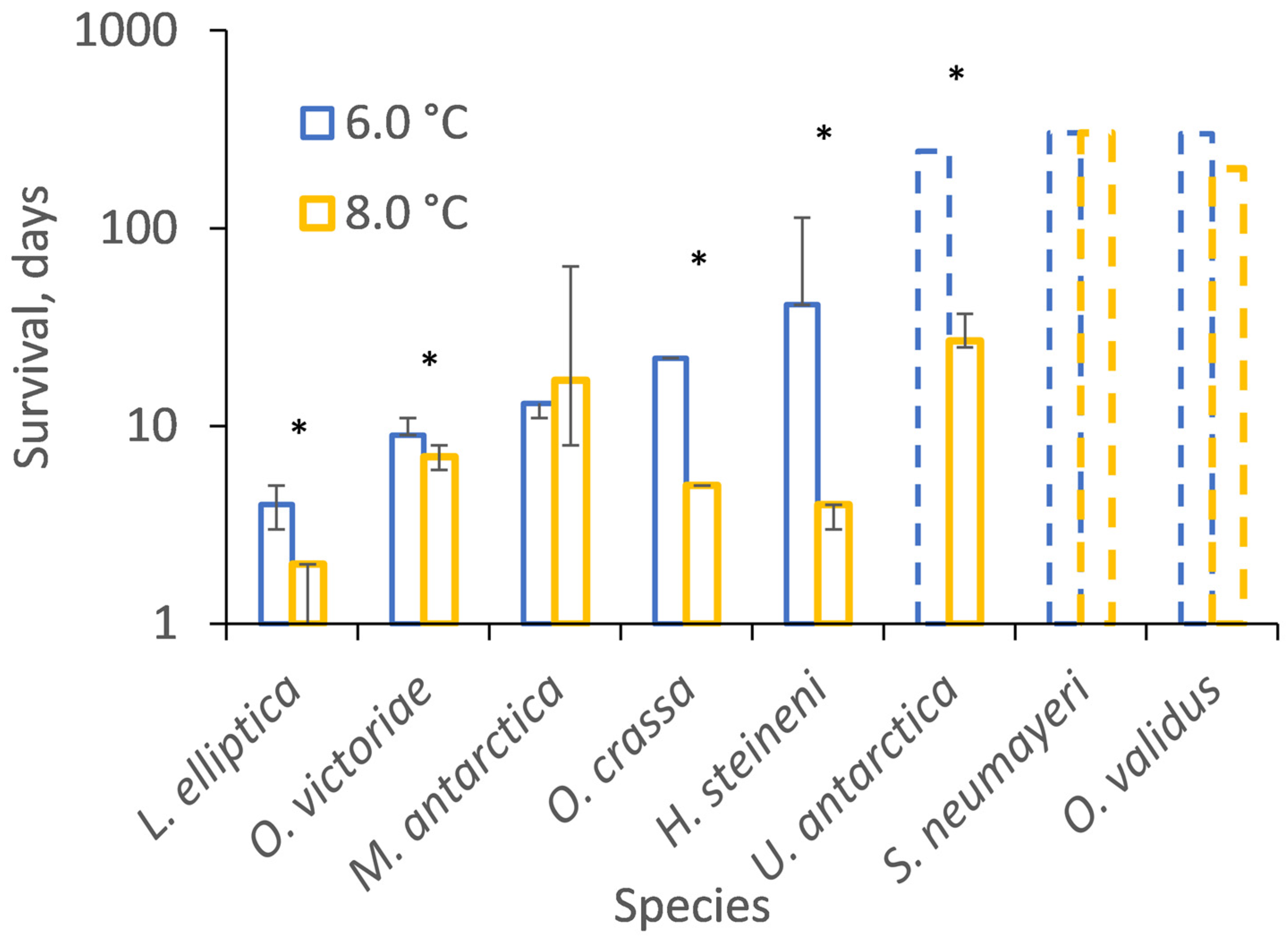
3.2. Temperature Incubations
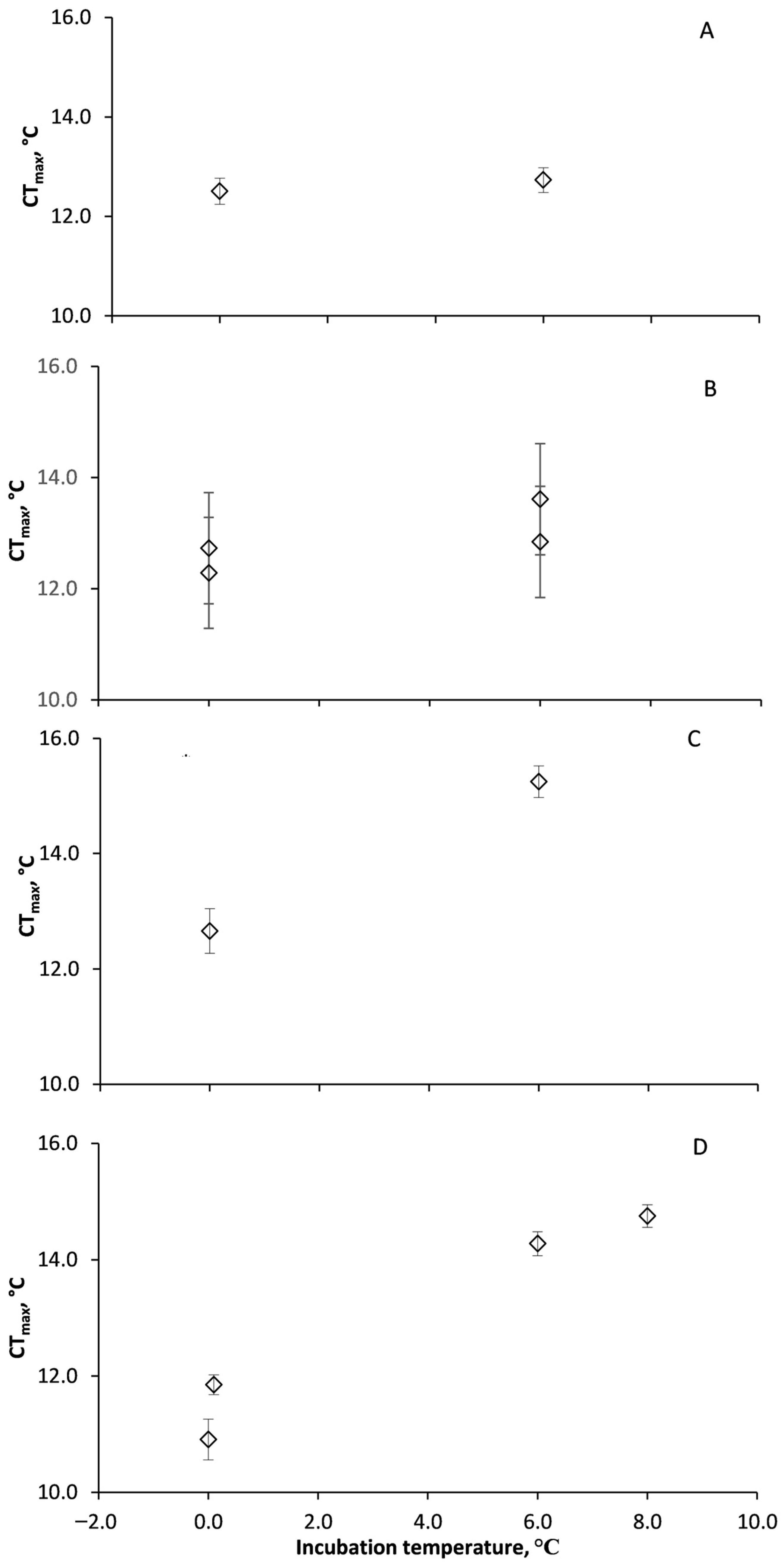
3.3. Acclimation of Acute CTmax
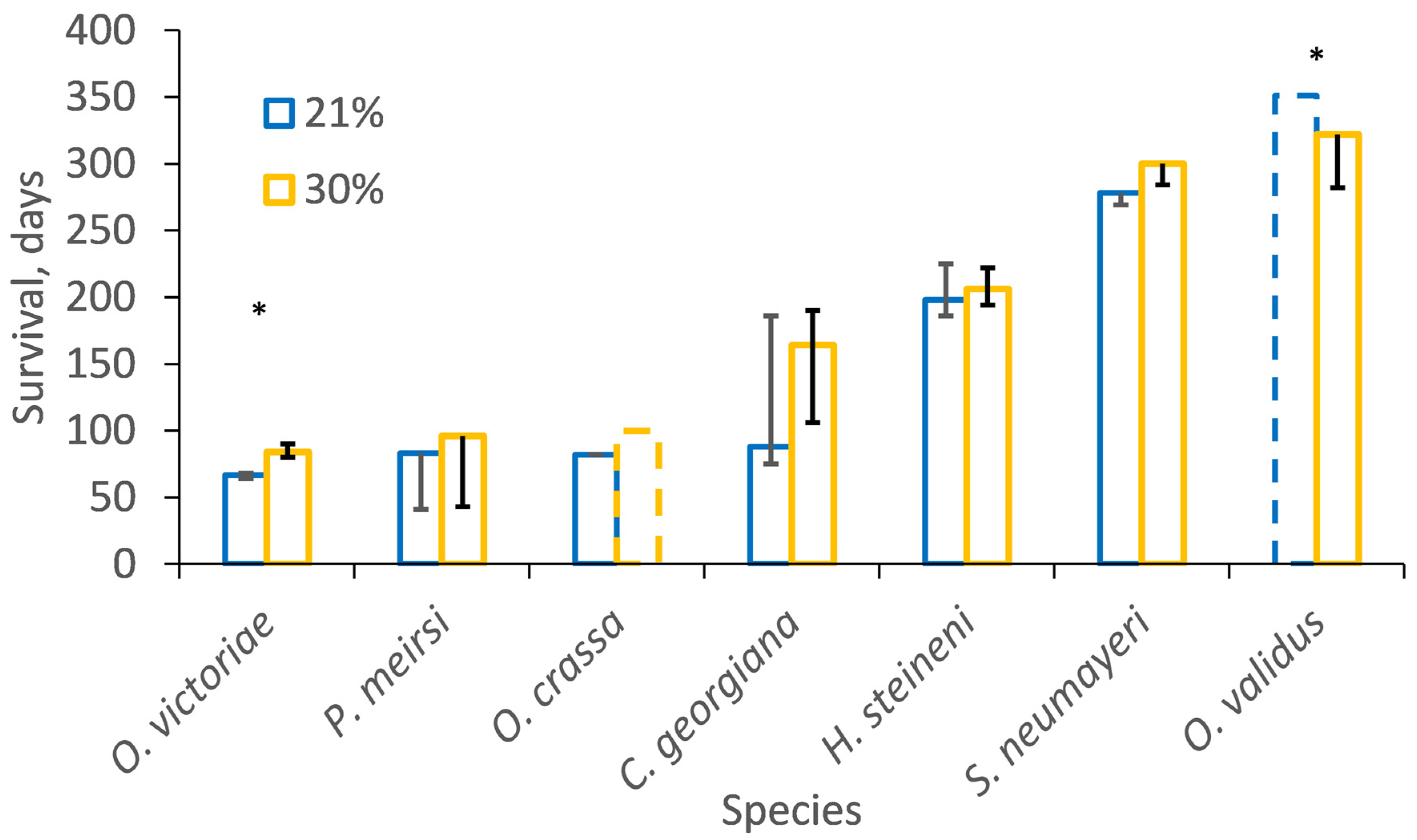
3.4. Temperature–Oxygen Incubations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J.; Chown, S.L.; Calosi, P.; Bernardo, J.; Bilton, D.T.; Clarke, A.; Clusella-Trullas, S.; Ghalambor, C.K.; Konarzewski, M.; Peck, L.S.; et al. Macrophysiology: A conceptual reunification. Am. Nat. 2009, 174, 595–612. [Google Scholar] [CrossRef]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. A 2002, 132, 739–761. [Google Scholar] [CrossRef]
- Lindquist, S. The heat shock response. Annu. Rev. Biochem. 1986, 55, 1151–1159. [Google Scholar] [CrossRef]
- Tomanek, L. Environmental proteomics: Changes in the proteome of marine organisms in response to environmental stress, pollutants, infection, symbiosis, and development. Ann. Rev. Mar. Sci. 2011, 3, 373–399. [Google Scholar] [CrossRef]
- Collier, R.J.; Baumgard, L.H.; Zimbelman, R.B.; Xiao, Y. Heat stress: Physiology of acclimation and adaptation. Anim. Front. 2019, 9, 12–19. [Google Scholar] [CrossRef]
- Hobday, A.J.; Alexander, L.V.; Perkins, S.E.; Smale, D.A.; Straub, S.C.; Oliver, E.C.J.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Feng, M.; et al. A hierarchical approach to defining marine heatwaves. Prog. Oceanogr. 2016, 141, 227–238. [Google Scholar] [CrossRef]
- Stillman, J. Acclimation underlies susceptibility to climate change. Science 2003, 301, 65. [Google Scholar] [CrossRef]
- Leroi, A.M.; Bennett, A.F.; Lenski, R.E. Temperature acclimation and competitive fitness: An experimental test of the Beneficial Acclimation Hypothesis. Proc. Natl. Acad. Sci. USA 1994, 91, 1917–1921. [Google Scholar] [CrossRef]
- Wilson, R.S.; Franklin, C.E. Testing the beneficial acclimation hypothesis. Trends Ecol. Evol. 2002, 17, 66–70. [Google Scholar] [CrossRef]
- Gunderson, A.R.; Stillman, J.H. Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc. R. Soc. B 2015, 282, 20150401. [Google Scholar] [CrossRef] [PubMed]
- Morley, S.A.; Peck, L.S.; Heiser, S.; Sunday, J.; Bates, A.E. Physiological acclimation extends predicted persistence of species across the globe. Glob. Ecol. Biodivers. 2019, 28, 1018–1037. [Google Scholar] [CrossRef]
- Peck, L.S.; Clark, M.S.; Morley, S.A.; Massey, A.; Rossetti, H. Animal temperature limits and ecological relevance: Effects of size, activity and rates of change. Funct. Ecol. 2009, 23, 248–256. [Google Scholar] [CrossRef]
- Molina, A.N.; Pulgar, J.M.; Rezende, E.L.; Carter, M.J. Heat tolerance of marine ectotherms in a warming Antarctica. Glob. Chang. Biol. 2023, 29, 179–188. [Google Scholar] [CrossRef]
- Clark, M.S.; Peck, L.S. Hsp70 heat shock proteins and environmental stress in Antarctic marine organisms: A mini-review. Mar. Genom. 2009, 2, 11–18. [Google Scholar] [CrossRef]
- Bates, A.E.; Lee, R.W.; Tunnicliffe, V.; Lamare, M.D. Deep-sea hydrothermal vent animals seek cool fluids in a highly variable thermal environment. Nat. Commun. 2010, 1, 14. [Google Scholar] [CrossRef]
- Morley, S.A.; Chu, J.W.F.; Peck, L.S.; Bates, A.E. Temperatures leading to heat escape responses in Antarctic marine ectotherms match acute thermal limits. Front. Physiol. 2022, 13, 1077376. [Google Scholar] [CrossRef]
- Clark, M.S.; Villota Nieva, L.; Hoffman, J.I.; Davies, A.J.; Trivedi, U.H.; Turner, F.; Ashton, G.; Peck, L.S. Lack of long-term acclimation in Antarctic encrusting species suggests vulnerability to warming. Nat. Commun. 2019, 10, 3383. [Google Scholar] [CrossRef]
- Collins, M.; Peck, L.S.; Clark, M.S. Large within, and between, species differences in marine cellular responses: Unpredictability in a changing environment. Sci. Total Environ. 2021, 794, 148594. [Google Scholar] [CrossRef]
- Clark, M.S.; Sommer, U.; Sihra, J.K.; Thorne, A.S.; Morley, S.A.; King, M.; Viant, M.R.; Peck, L.S. Biodiversity in marine invertebrate responses to acute warming revealed by a comparative multi-omics approach. Glob. Chang Biol. 2017, 23, 318–330. [Google Scholar] [CrossRef]
- Clark, T.D.; Sandblom, E.; Jutfelt, F. Aerobic scope measurements of fishes in an era of climate change: Respirometry, relevance and recommendations. J. Exp. Biol. 2013, 216, 2771–2782. [Google Scholar] [CrossRef]
- Verberk, W.C.; Overgaard, J.; Ern, R.; Bayley, M.; Wang, T.; Boardman, L.; Terblanche, J.S. Does oxygen limit thermal tolerance in arthropods? A critical review of current evidence. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 192, 64–78. [Google Scholar] [CrossRef]
- Richard, J.; Morley, S.A.; Thorne, M.A.; Peck, L.S. Estimating long term survival temperatures at the assemblage level in the marine environment: Towards macrophysiology. PLoS ONE 2012, 7, e34655. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Venables, H.; Meredith, M.P.; Hendry, K.R.; ten Hoopen, P.; Peat, H.; Chapman, A.; Beaumont, J.; Piper, R.; Miller, A.J.; Mann, P.; et al. Sustained, year-round oceanographic measurements from Rothera Research Station, Antarctica, 1997–2017. Sci. Data 2023, 10, 265. [Google Scholar] [CrossRef]
- Allignol, A.; Latouche, A. CRAN Task View: Survival Analysis, Version 2023-09-10. 2023. Available online: https://CRAN.R-project.org/view=Survival (accessed on 1 January 2023).
- De Leij, R.; Grange, L.J.; Peck, L.S. Functional thermal limits are determined by rate of warming during simulated marine heatwaves. Mar. Ecol. Prog. Ser. 2022, 685, 183–196. [Google Scholar] [CrossRef]
- Carrasco, J.F.; Bozkurt, D.; Cordero, R.R. A review of the observed air temperature in the Antarctic Peninsula. did the warming trend come back after the early 21st hiatus? Polar Sci. 2021, 28, 100653. [Google Scholar] [CrossRef]
- Peck, L.S.; Webb, K.E.; Bailey, D. Extreme sensitivity of biological function to temperature in Antarctic marine species. Funct. Ecol. 2004, 18, 625–630. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Bock, C.; Mark, F.C. Oxygen- and capacity-limited thermal tolerance: Bridging ecology and physiology. J. Exp. Biol. 2017, 220, 2685–2696. [Google Scholar] [CrossRef]
- McArley, T.J.; Sandblom, E.; Herbert, N.A. Fish and hyperoxia—From cardiorespiratory and biochemical adjustments to aquaculture and ecophysiology implications. Fish Fish. 2020, 22, 324–355. [Google Scholar] [CrossRef]
- Abele, D.; Puntarulo, S. Formation of reactive species and induction of antioxidant defence systems in polar and temperate marine invertebrates and fish. Comp. Biochem. Physiol. A 2004, 138, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Franklin, C.E.; Davison, W.; Kraft, P. Stenotherms at subzero temperatures: Thermal dependence of swimming performance in Antarctic fish. J. Comp. Physiol. 2001, 171, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Seebacher, F.; Davison, W.; Lowe, C.J.; Franklin, C.E. A falsification of the thermal specialization paradigm: Compensation for elevated temperatures in Antarctic fishes. Biol. Lett. 2005, 1, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Franklin, C.E.; Davison, W.; Seebacher, F. Antarctic fish can compensate for rising temperatures: Thermal acclimation of cardiac performance in Pagothenia borchgrevinki. J. Exp. Biol. 2007, 210, 3068–3074. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.; Davison, W. The Antarctic notothenioid fish Pagothenia borchgrevinki is thermally flexible: Acclimation changes oxygen consumption. Polar Biol. 2008, 31, 317–326. [Google Scholar] [CrossRef]
- Bilyk, K.T.; Devries, A.L. Heat tolerance and its plasticity in Antarctic fishes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 158, 382–390. [Google Scholar]
- Strobel, A.; Bennecke, S.; Leo, E.; Mintenbeck, K.; Pörtner, H.O.; Mark, F.C. Metabolic shifts in the Antarctic fish Notothenia rossii in response to rising temperature and PCO2. Front. Zool. 2012, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Gao, L.; Luo, Y.; Li, X.; Zheng, X.; Zhang, X.; Cheng, X.; Jia, F.; Purich, A.; Santoso, A.; et al. Southern Ocean warming and its climatic impacts. Sci. Bull. 2023, 68, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Brokordt, K.B.; Himmelman, J.H.; Guderley, H.E. Effect of reproduction on escape responses and muscle metabolic capacities in the scallop Chlamys islandica Müller. J. Exp. Mar. Biol. Ecol. 2000, 251, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Obermüller, B.O.; Morley, S.A.; Clark, M.S.; Barnes, D.K.A.; Peck, L.S. Antarctic intertidal limpet ecophysiology: A winter-summer comparison. J. Exp. Mar. Biol. Ecol. 2011, 403, 39–45. [Google Scholar] [CrossRef]
- Clark, M.S.; Geissler, P.; Waller, C.; Fraser, K.P.P.; Barnes, D.K.A.; Peck, L.S. Low heat shock thresholds in wild Antarctic inter-tidal limpets (Nacella concinna). Cell Stress Chaperones 2008, 13, 51–58. [Google Scholar] [CrossRef]
- Brown, J.H.; Gilooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Towards a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Duration (d) | Tank | Treatment | Temperature (°C) | Oxygen (% O2) | Oxygen (mg/L) * |
|---|---|---|---|---|---|---|
| (A) | ||||||
| 2010/11 | 100 | 1 | Control | 0.9 ± 0.05 | 21.0 ± 0.1 | 13.9 |
| 2 | Normoxic | 3.0 ± 0.01 | 21.0 ± 0.1 | 13.2 | ||
| 3 | Hyperoxic | 3.0 ± 0.02 | 29.3 ± 0.5 | 18.3 | ||
| (B) | ||||||
| 2014/15 | 351 | 1 | Control | 0.1 ± 0.01 | 21.0 ± 0.1 | 14.2 |
| 2 | Normoxic | 8.6 ± 0.5 (10.0 ± 0.01) | 21.0 ± 0.3 | 11.4 | ||
| 3 | Normoxic | 8.1 ± 0.2 (9.9 ± 0.01) | 21.5 ± 0.5 | 11.8 | ||
| 4 | Hyperoxic | 8.1 ± 0.2 (10.0 ± 0.01) | 30.5 ± 0.6 | 16.8 | ||
| 5 | Hyperoxic | 8.0 ± 0.2 (10.0 ± 0.01) | 31.3 ± 0.7 | 17.2 |
| Species | Incubation | Mortalities | Survivors | % Survival |
|---|---|---|---|---|
| S. neumayeri | 6 °C | 10 | 21 | 68 |
| 8 °C | 2 | 10 | 83 | |
| O. validus | 6 °C | 1 | 19 | 95 |
| 8 °C | Technical failure | |||
| O. victoriae | 6 °C | 27 | 0 | 0 |
| 8 °C | 23 | 0 | 0 | |
| O. crassa | 6 °C | 24 | 0 | 0 |
| 8 °C | 25 | 0 | 0 | |
| M. antarctica | 6 °C | 13 | 0 | 0 |
| 8 °C | 19 | 0 | 0 | |
| U. antarctica | 6 °C | 1 | 13 | 93 |
| 8 °C | 16 | 0 | 0 | |
| L. elliptica | 6 °C | 19 | 0 | 0 |
| 8 °C | 15 | 0 | 0 | |
| H. steineni | 6 °C | 21 | 4 | 16 |
| 8 °C | 25 | 0 | 0 | |
| O. validus | 21% | 8 | 11 | 58 |
| 30% | 12 | 8 | 40 | |
| O. crassa | 21% | 26 | 4 | 13 |
| 30% | 4 | 26 | 87 | |
| P. miersi | 21% | 15 | 15 | 50 |
| 30% | 15 | 15 | 50 | |
| O. victoriae | 21% | 30 | 0 | 0 |
| 30% | 30 | 0 | 0 | |
| C. georgiana | 21% | 20 | 0 | 0 |
| 30% | 20 | 0 | 0 | |
| S. neumayeri | 21% | 18 | 3 | 14 |
| 30% | 19 | 0 | 0 | |
| H. steineni | 21% | 20 | 0 | 0 |
| 30% | 19 | 1 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morley, S.A.; Bates, A.E.; Clark, M.S.; Fitzcharles, E.; Smith, R.; Stainthorp, R.E.; Peck, L.S. Testing the Resilience, Physiological Plasticity and Mechanisms Underlying Upper Temperature Limits of Antarctic Marine Ectotherms. Biology 2024, 13, 224. https://doi.org/10.3390/biology13040224
Morley SA, Bates AE, Clark MS, Fitzcharles E, Smith R, Stainthorp RE, Peck LS. Testing the Resilience, Physiological Plasticity and Mechanisms Underlying Upper Temperature Limits of Antarctic Marine Ectotherms. Biology. 2024; 13(4):224. https://doi.org/10.3390/biology13040224
Chicago/Turabian StyleMorley, Simon A., Amanda E. Bates, Melody S. Clark, Elaine Fitzcharles, Rebecca Smith, Rose E. Stainthorp, and Lloyd S. Peck. 2024. "Testing the Resilience, Physiological Plasticity and Mechanisms Underlying Upper Temperature Limits of Antarctic Marine Ectotherms" Biology 13, no. 4: 224. https://doi.org/10.3390/biology13040224