
The Parasporal Body of Bacillus thuringiensis subsp. israelensis: A Unique Phage Capsid-Associated Prokaryotic Insecticidal Organelle
 , , and
, , and 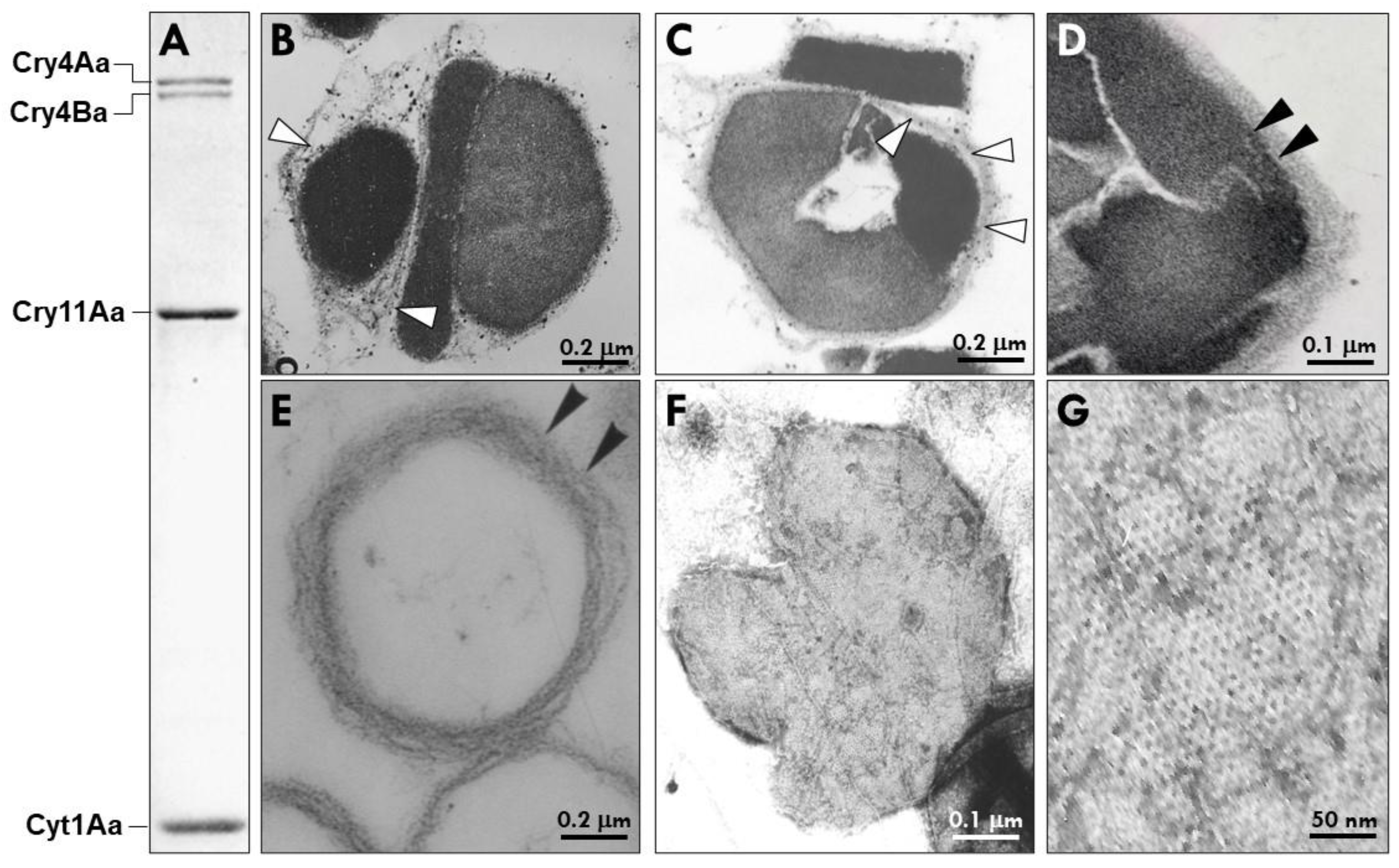{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
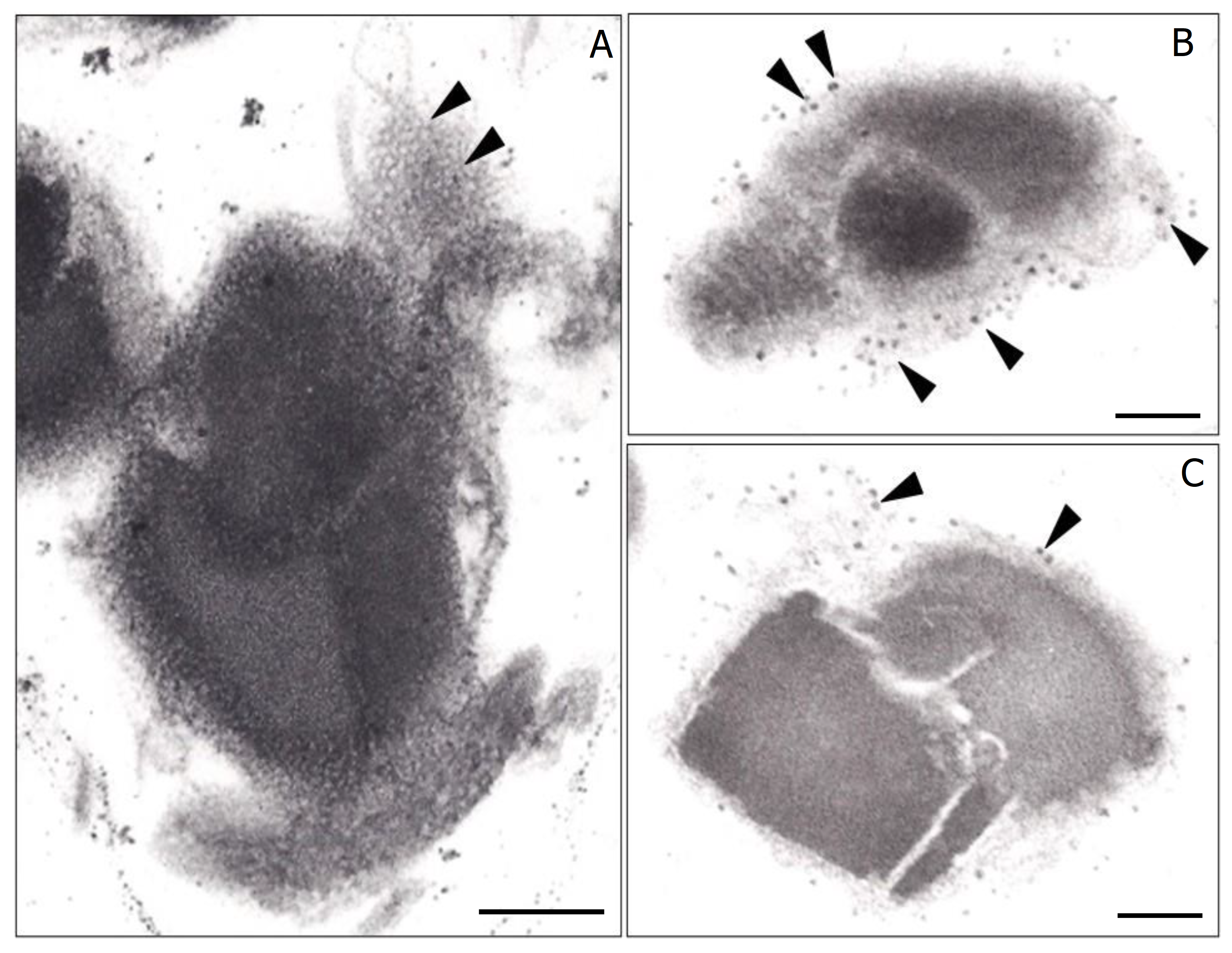{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Studies Supporting Classification of Bti’s PB as a Prokaryotic Insecticidal Organelle
2.1. Bti’s PB Is a Complex of Heterogenous Microcompartments Enveloped by a Well-Defined Multilamellar Fibrous Matrix (MFM)
2.2. Bti’s MFM Contains a Unique Collection of Proteins
2.3. Bt073, Bt113 and Bt148
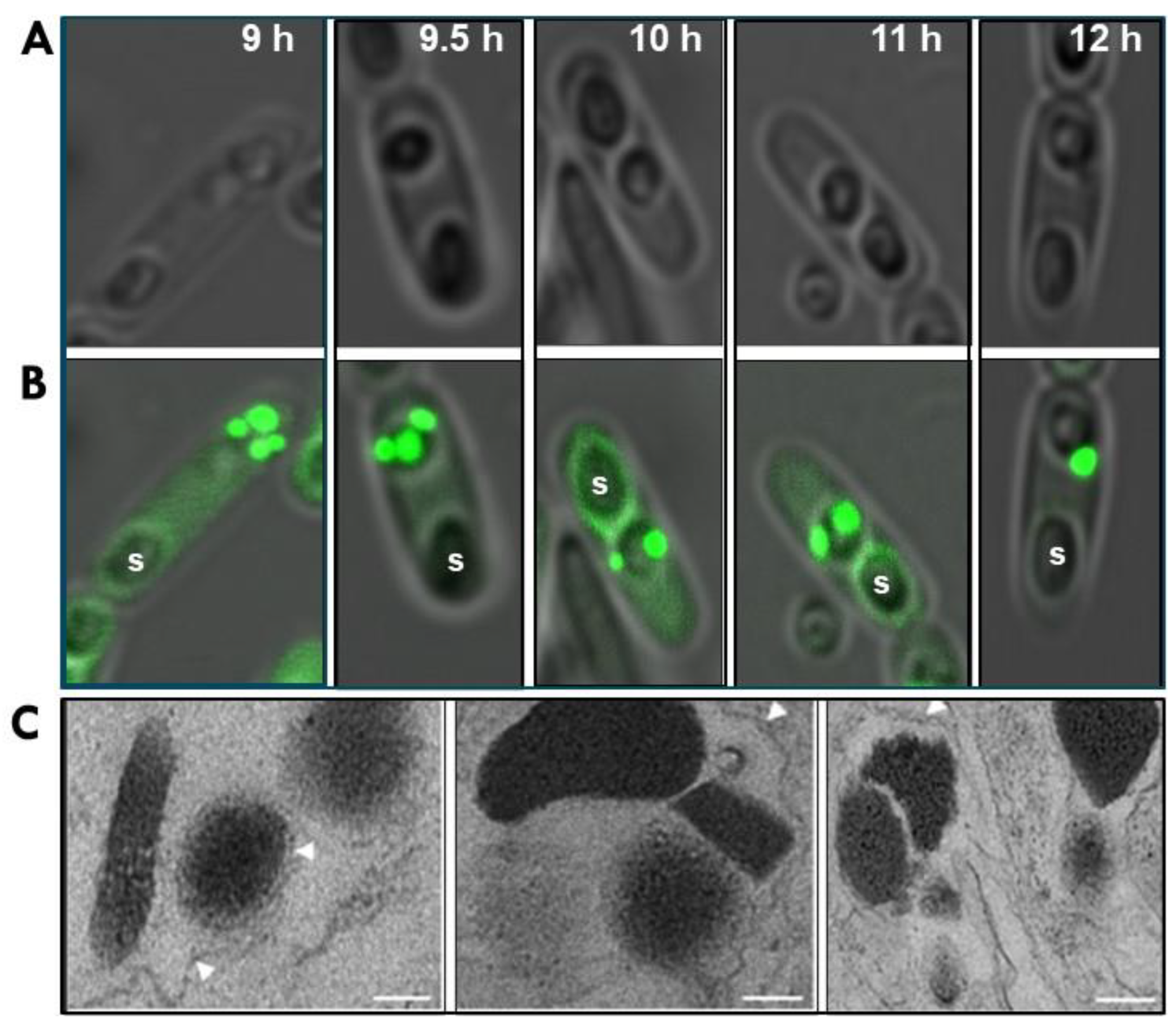
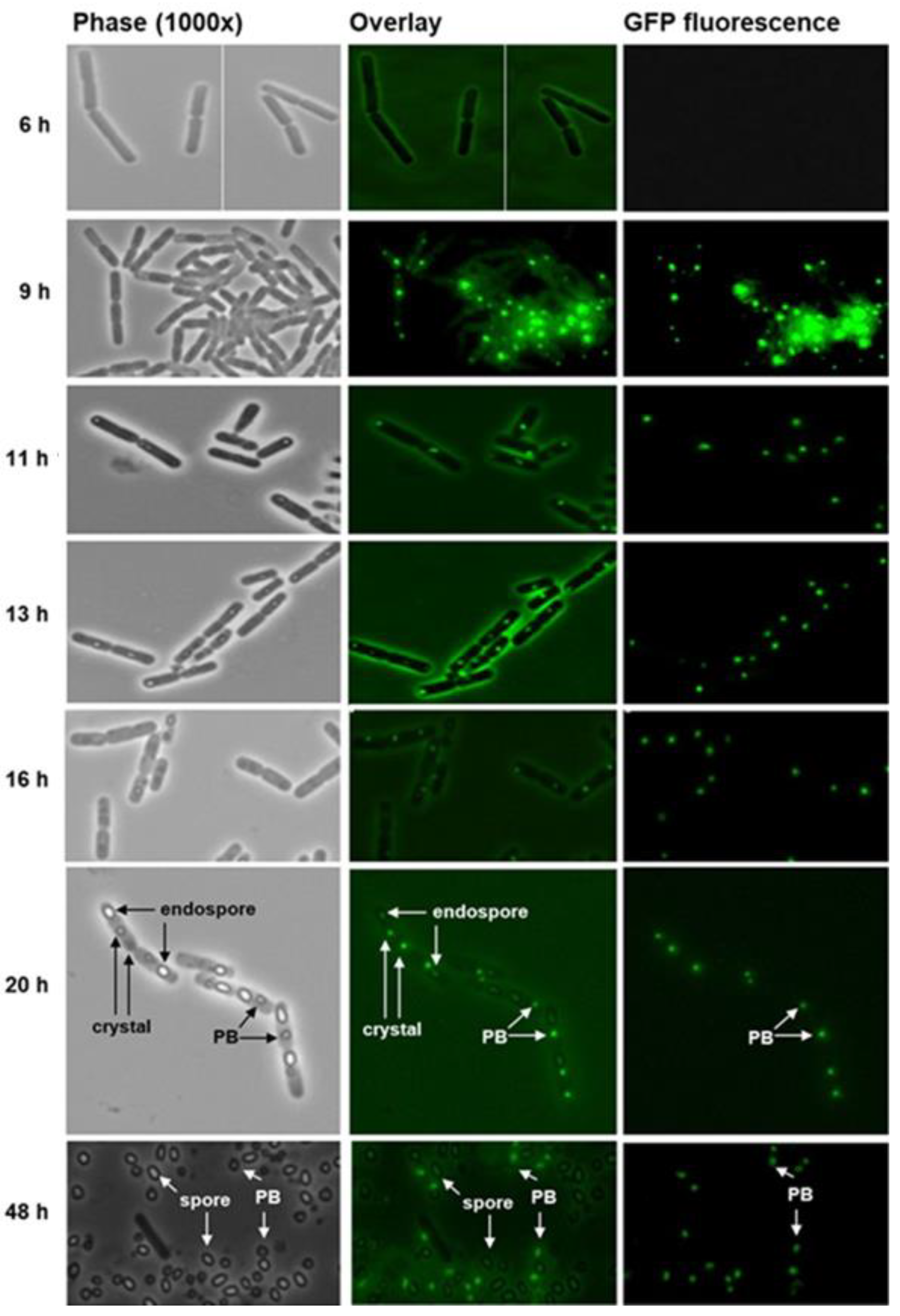
2.4. Bt152 Tracks the Fusion of PB Compartments into a Single Parasporal Body
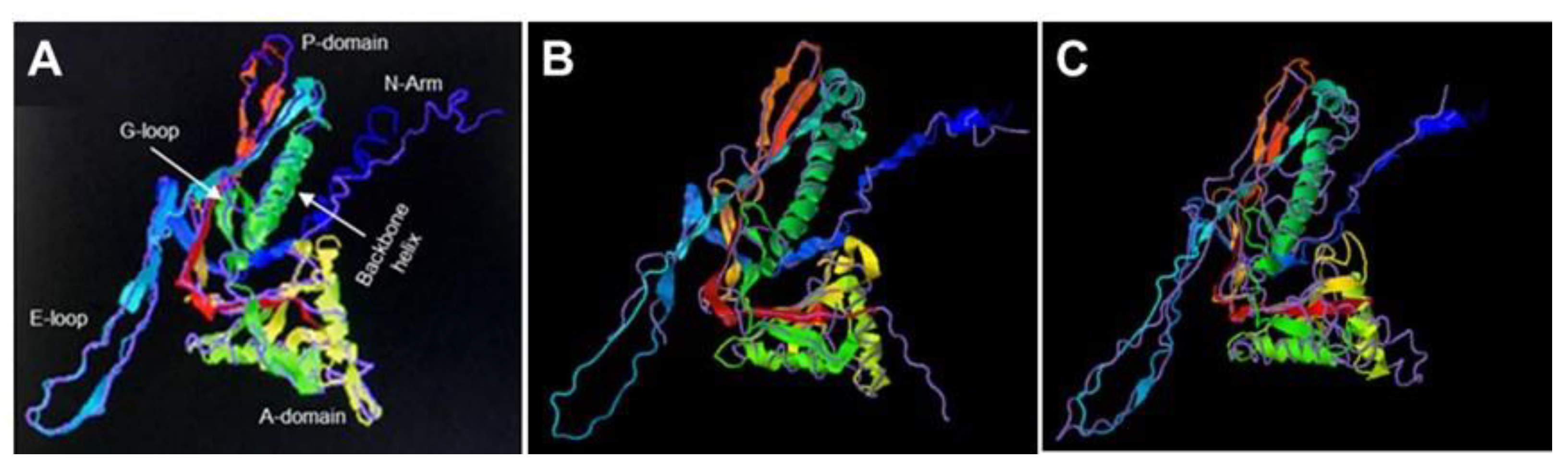
2.5. Bt075 Is Structurally Similar to Encapsulin Shell Proteins
2.6. Signature Sequence for the Bti’s Insecticidal Proteins
3. Is Bti’s PB a Novel Prokaryotic Organelle to Store Proteins?
4. Conclusions
5. Materials and Methods
5.1. Bacterial Strains and Propagation
5.2. In Vivo Localization of Bt075
5.3. Sequence Analyses, Molecular Modeling and Phylogenetic Trees
5.4. Confocal Microscopy
5.5. Electron Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Greening, C.; Lithgow, T. Formation and function of bacterial organelles. Nat. Rev. Microbiol. 2021, 18, 677–689. [Google Scholar] [CrossRef]
- Lacey, L.A. Bacillus thuringiensis serovariety israelensis and Bacillus sphaericus for mosquito control. J. Am. Mosq. Control. Assoc. 2007, 23, 133–163. [Google Scholar] [CrossRef]
- Federici, B.A.; Park, H.W.; Sakano, Y. Insecticidal Proteins Crystals of Bacillus thuringiensis; Shively, J.M., Ed.; Inclusions in Prokaryotes, Microbiol Monogr (1); Springer: Berlin/Heidelberg, Germany, 2006; pp. 196–232. [Google Scholar]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and its dipteran-specific toxins. Toxins 2014, 6, 1222–1243. [Google Scholar] [CrossRef]
- Bruh, C.A.; Despres, L.; Fror, O.; Patil, C.A.; Poulin, B.; Tetreau, G.; Allgeier, S. Environmental and socioeconomic effects of mosquito control in Europe using the biocide Bacillus thuringiensis subsp. israelensis (Bti). Sci. Total. Environ. 2020, 724, 137800. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2021, 186, 107438. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberon, M. Bacillus thuringiensis: A story of a successful biopesticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Grassmann, A.J.; Reisig, D.D. Management of insect pest with Bt crops in the United States. Annu. Rev. Entomol. 2023, 68, 31–49. [Google Scholar] [CrossRef]
- Berry, C.; O’Neil, S.; Ben-Dov, E.; Jones, A.F.; Murphy, L.; Quail, M.A.; Holden, M.T.; Harris, D.; Zaritsky, A.; Parkhill, J. Complete sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 2002, 68, 5082–5095. [Google Scholar] [CrossRef]
- Palma, L.; Munoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: And overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef]
- Silva-Filha, M.H.N.L.; Romão, T.P.; Rezende, T.M.T.; Carvalho, K.d.S.; Gouveia de Menezes, H.S.; Alexandre do Nascimento, N.; Soberón, M.; Bravo, A. Bacterial Toxins Active against Mosquitoes: Mode of Action and Resistance. Toxins 2021, 13, 523. [Google Scholar] [CrossRef]
- Heckel, D.G. How do toxins from Bacillus thuringiensis kill insects? And evolutionary perspective. Arch. Insect Biochem. Physiol. 2022, 4, e2167. [Google Scholar] [CrossRef]
- Guerchicoff, A.; Delecluse, A.; Rubinstein, C.P. The Bacillus thuringiensis cyt genes for hemolytic endotoxins constitute a gene family. Appl. Environ. Microbiol. 2001, 67, 1090–1096. [Google Scholar] [CrossRef]
- Anaya, P.; Onofre, J.; Torres-Quintero, M.C.; Sánchez, J.; Gill, S.S.; Bravo, A.; Soberón, M. Oligomerization is a key step for Bacillus thuringiensis Cyt1Aa1 insecticidal activity but not for toxicity against red blood cells. Insect Biochem. Mol. Biol. 2020, 119, 103317. [Google Scholar] [CrossRef] [PubMed]
- Tetreau, G.; Banneville, A.S.; Andreeva, E.A.; Brewster, A.S.; Hunter, M.S.; Sierra, R.G.; Teulon, J.M.; Young, I.D.; Burke, N.; Grünewald, T.A.; et al. Serial femtosecond crystallography on in vivo-grown crystals drives elucidation of mosquitocidal Cyt1Aa1 bioactivation cascade. Nat. Commun. 2020, 11, 1153. [Google Scholar] [CrossRef] [PubMed]
- Bideshi, D.K.; Park, H.-W.; Hice, R.D.; Wirth, M.C.; Federici, B.A. Highly effective broad spectrum chimeric larvicide that targets vector mosquitoes using a lipophilic protein. Sci. Rep. 2017, 7, 11282. [Google Scholar] [CrossRef]
- Lai, L.; Villanueva, M.; Muruzabal-Galarza, A.; Beatriz Fernández, A.; Unzue, A.; Toledo-Arana, A.; Caballero, A.; Caballero, C.J. Bacillus thuringiensis Cyt Proteins as Enablers of Activity of Cry and Tpp Toxins against Aedes albopictus. Toxins 2023, 15, 211. [Google Scholar] [CrossRef]
- Diaz-Mendoza, M.; Bideshi, D.K.; Federici, B.A. A 54 -kilodalton proteins encoded by pBtoxis is required for parasporal body structural integrity in Bacillus thuringiensis subsp. israelensis. J. Bacteriol. 2012, 194, 1562–1571. [Google Scholar] [CrossRef]
- Ibarra, J.E.; Federici, B.A. Parasporal bodies of Bacillus thuringiensis subsp. morrisoni (PG14) and Bacillus thuringiensis subsp. israelensis are similar in protein composition and toxicity. FEMS Microbiol. Lett. 1989, 34, 79–84. [Google Scholar] [CrossRef]
- Galjart, N.J.; Sivasubramanian, N.; Federici, B.A. Plasmid location, cloning, and sequence analysis of the gene encoding a 27.3-kilodalton cytolytic protein from Bacullus thuringiensis subsp. morrisoni (PG14). Curr. Microbiol. 1987, 16, 171–177. [Google Scholar] [CrossRef]
- Gabashvili, A.N.; Chemelyuk, N.S.; Efremova, M.V.; Malinoskaya, J.A.; Semkina, A.S.; Abakumov, M.A. Encapsulins-Bacterial protein nanocompartments: Structure, properties, and applications. Biomolecules 2020, 10, 966. [Google Scholar] [CrossRef] [PubMed]
- Giessen, T.W. Encapsulins: Microbial nanocompartments with application in biomedicine, nanobiotechnology and materials science. Curr. Opin. Chem. Biol. 2016, 34, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nichols, R.J.; Cassidy-Amstutz, C.; Chaijarasphong, T.; Savage, D.F. Encapsulins: Molecular biology of the shell. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 583–594. [Google Scholar] [CrossRef]
- Lau, Y.H.; Giessen, T.W.; Altenburg, W.J.; Silver, P.A. Prokaryotic nanocompartments form synthetic organelles in a eukaryote. Nat. Commun. 2018, 9, 1311. [Google Scholar] [CrossRef]
- Duda, R.L.; Teschke, C.M. The amazing HK97 fold: Versatile results of modest differences. Curr. Opin. Virol. 2019, 36, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.V.; Carvalhi, A.J.; Pereira, A.S. Encapsulin nanocages: Protein encapsulation and iron sequestration. Coord. Chem. Rev. 2021, 448, 214188. [Google Scholar] [CrossRef]
- Giessen, T.W. Encapsulins. Annu. Rev. Biochem. 2022, 19, 353–380. [Google Scholar] [CrossRef]
- Chmelyuk, N.S.; Oda, V.V.; Gabashvili, A.N.; Abakumov, M.A. Encapsiluns: Structure, properties, and biotechnological applications. Biochemistry 2023, 88, 35–49. [Google Scholar] [PubMed]
- McHugh, C.A.; Fontana, J.; Nemecek, D.; Cheng, N.; Aksyuk, A.A.; Heymann, J.B.; Winkler, D.C.; Lam, A.S.; Wall, J.S.; Steven, A.C.; et al. A virus capsid-like nanocompartment that store iron and protect bacteria from oxidative stress. EMBO J. 2014, 33, 1896–1911. [Google Scholar] [CrossRef]
- Lien, K.A.; Dinshaw, K.; Nichols, R.J.; Cassidy-Amstutz, C.; Knight, M.; Singh, R.; Eltis, L.D.; Savage, D.F.; Stanley, S.A. A nanocompartment system contributes of defense against oxidative stress in Mycobacterium tuberculosis. Elife 2021, 10, e74358. [Google Scholar] [CrossRef]
- Wu, D.; Federici, B.A. A 20-kilodalton protein preserves cell viability and promotes CytA crystal formation in Bacillus thuringiensis. J. Bacteriol. 1993, 175, 5276–5280. [Google Scholar] [CrossRef]
- Manasherob, R.; Zaritsk, A.; Ben-Dov, E.; Saxena, D.; Barak, Z.; Einav, M. Effect of accessory proteins P19 and P20 on cytolytic activity of Cyt1Aa1 from Bacillus thuringiensis subsp. israelensis in Escherichia coli. Curr. Microbiol. 2001, 43, 355–364. [Google Scholar] [CrossRef]
- Cohen, S.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A.; Dym, O. Cyt1Aa1 toxin: Crystal structure reveals implications for its membrane-perforating function. J. Mol. Biol. 2011, 413, 804–814. [Google Scholar] [CrossRef]
- Norris, V.; Mileykovskaya Eugenia Matsumoto, K. Expanding the transertion hypothesis. Biochem. Anal. Biochem. 2015, 4, 1000234. [Google Scholar]
- Kurland, C.G. Molecular characterization of ribonucleic acids from Escherichia coli ribosomes. J. Mol. Biol. 1960, 2, 83–91. [Google Scholar] [CrossRef]
- Watson, Z.L.; Ward, F.R.; Méheust, R.; Ad, O.; Schepartz, A.; Banfield, J.F.; Cate, J.H. Structure of the bacterial ribosome at 2.0 A resolution. eLife 2020, 9, e60482. [Google Scholar] [CrossRef]
- Kennedy, N.W.; Mills, C.E.; Nichols, T.M.; Abrahamson, C.H.; Tullman-Ercek, D. Bacterial microcompartmens: Tiny organelles with big potential. Curr. Opin. Microbiol. 2021, 63, 36–42. [Google Scholar] [CrossRef]
- Demchuk, A.M.; Patel, T.R. The biomedical and bioengineering of protein microcompartments. Biotechnol. Adv. 2020, 41, 107547. [Google Scholar] [CrossRef]
- McDowell, H.B.; Hoiczyk, E. Bacterial nanocompartments: Structure, functions and applications. J. Bacteriol. 2022, 15, e0034621. [Google Scholar] [CrossRef]
- Stewart, A.M.; Steward, K.L.; Yeates, T.O.; Bobik, T.A. Advances in the world of bacterial microcompartments. Trends Biochem. Sci. 2021, 46, 406–416. [Google Scholar] [CrossRef]
- Planamente, S.; Frank, S. Bioengineering of bacterial microcompartments. Biochem. Soc. Trans. 2019, 47, 765–777. [Google Scholar] [CrossRef]
- Kumar, G.; Sinha, S. Biophysical approaches to understand and re-purposebacterial microcompartments. Curr. Opin. Microbiol. 2021, 63, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Benisch, R.; Giessen, T.W. Encapsulin cargo loading: Progress and potential. J. Material. Chem. B 2023, 11, 4377–4388. [Google Scholar] [CrossRef] [PubMed]
- Rose, S.M.; Radhakrishnan, A.; Sinha, S. Innate and engineered attributes of bacterial microcompartments for applications in bio-materials science. J. Mater. Chem. B 2023, 11, 4842–4854. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y. New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, C.; Li, Y.; Pearce, R.; Bell, E.W.; Zhang, Y. Folding non-homology proteins by compiling deep-learning contact maps with I-TASSER assembly simulations. Cell Rep. Methods 2021, 1, 100014. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudd, S.R.; Miranda, L.S.; Curtis, H.R.; Bigot, Y.; Diaz-Mendoza, M.; Hice, R.; Nizet, V.; Park, H.-W.; Blaha, G.; Federici, B.A.; et al. The Parasporal Body of Bacillus thuringiensis subsp. israelensis: A Unique Phage Capsid-Associated Prokaryotic Insecticidal Organelle. Biology 2023, 12, 1421. https://doi.org/10.3390/biology12111421
Rudd SR, Miranda LS, Curtis HR, Bigot Y, Diaz-Mendoza M, Hice R, Nizet V, Park H-W, Blaha G, Federici BA, et al. The Parasporal Body of Bacillus thuringiensis subsp. israelensis: A Unique Phage Capsid-Associated Prokaryotic Insecticidal Organelle. Biology. 2023; 12(11):1421. https://doi.org/10.3390/biology12111421
Chicago/Turabian StyleRudd, Sarah R., Leticia Silva Miranda, Hannah R. Curtis, Yves Bigot, Mercedes Diaz-Mendoza, Robert Hice, Victor Nizet, Hyun-Woo Park, Gregor Blaha, Brian A. Federici, and et al. 2023. "The Parasporal Body of Bacillus thuringiensis subsp. israelensis: A Unique Phage Capsid-Associated Prokaryotic Insecticidal Organelle" Biology 12, no. 11: 1421. https://doi.org/10.3390/biology12111421