
How Rare Are Argonautoidea Octopuses in the Mediterranean? New Data from Stranding Events, Stomach Contents and Genetics
 , , , , , ,
, , , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
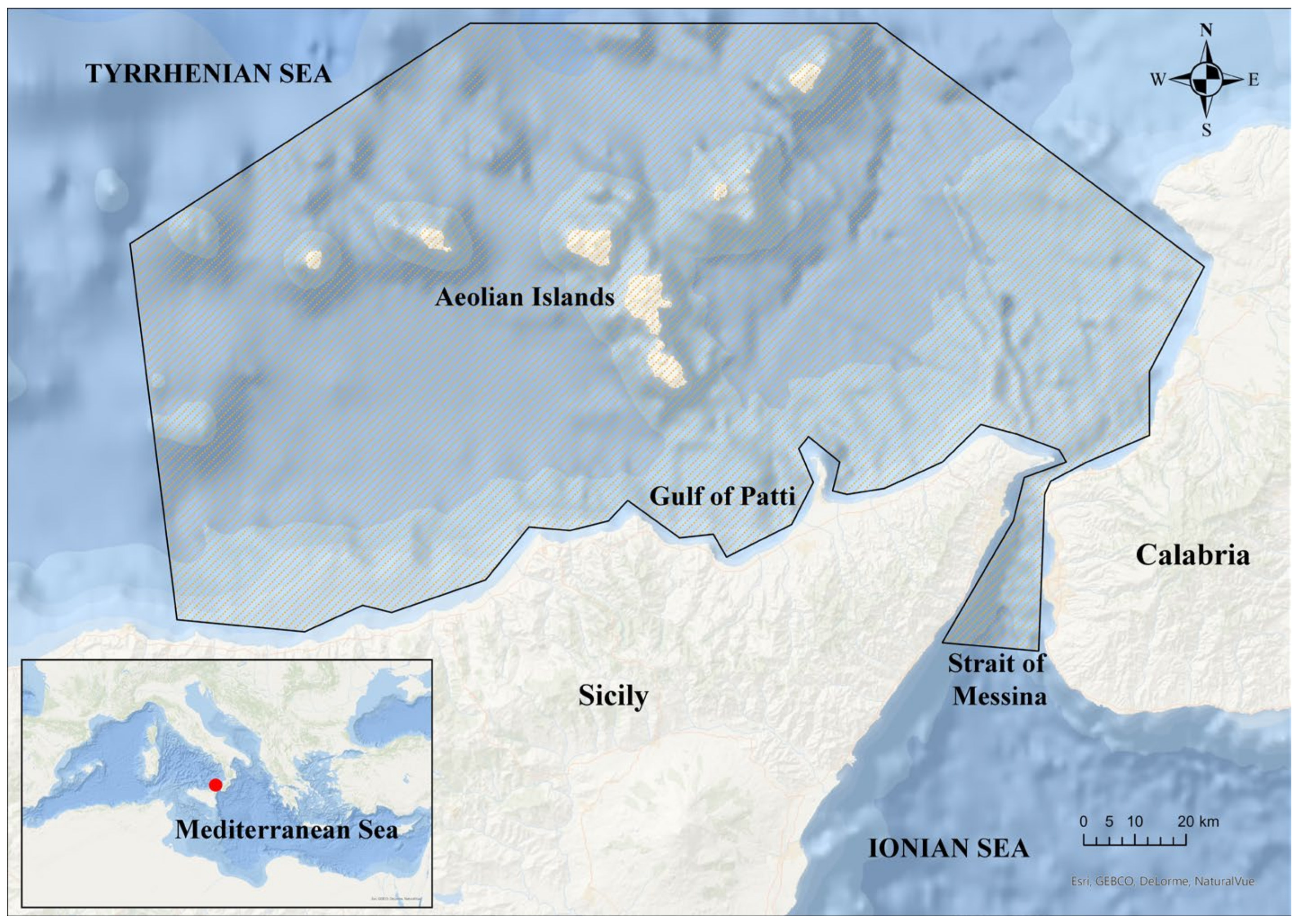
2.1. Sampling
2.1.1. Stranded Individuals
2.1.2. Occasional Catches
2.1.3. Argonautoidea in Stomach Contents of Top Predators
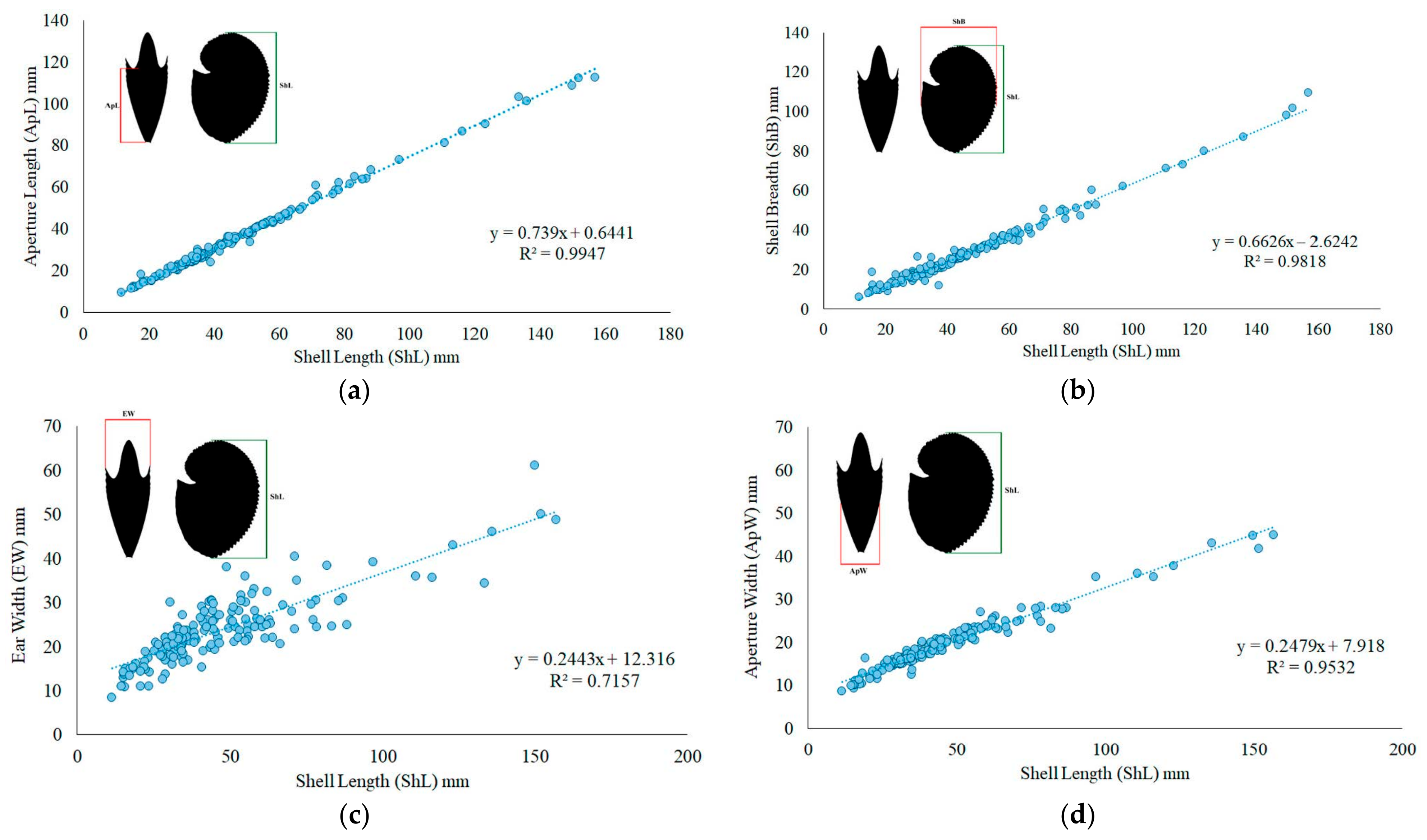
2.2. Morphometric and Biological Analysis
2.3. Occurrence of Holopelagic Octopuses in Predator Diet
2.4. Genetic Analysis
2.4.1. DNA Extraction and Amplification of Mitochondrial Cytochrome c Oxidase Subunit I (COI)
2.4.2. Sequencing and Molecular Analysis
3. Results
3.1. Stranded Individuals and Occasional Catches
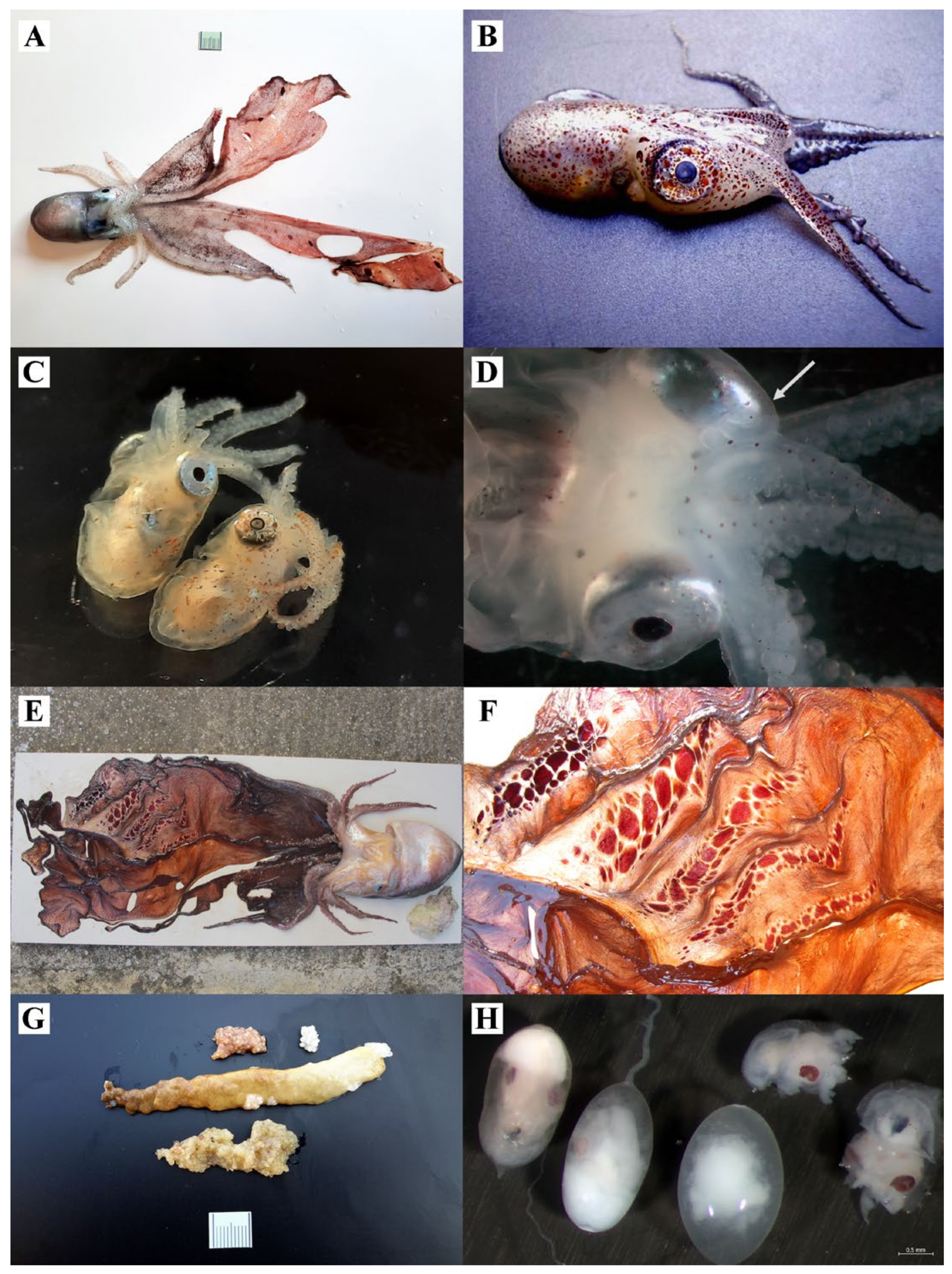
3.1.1. Argonauta argo
3.1.2. Ocythoe tuberculata
3.1.3. Tremoctopus violaceus and Tremoctopus gracilis
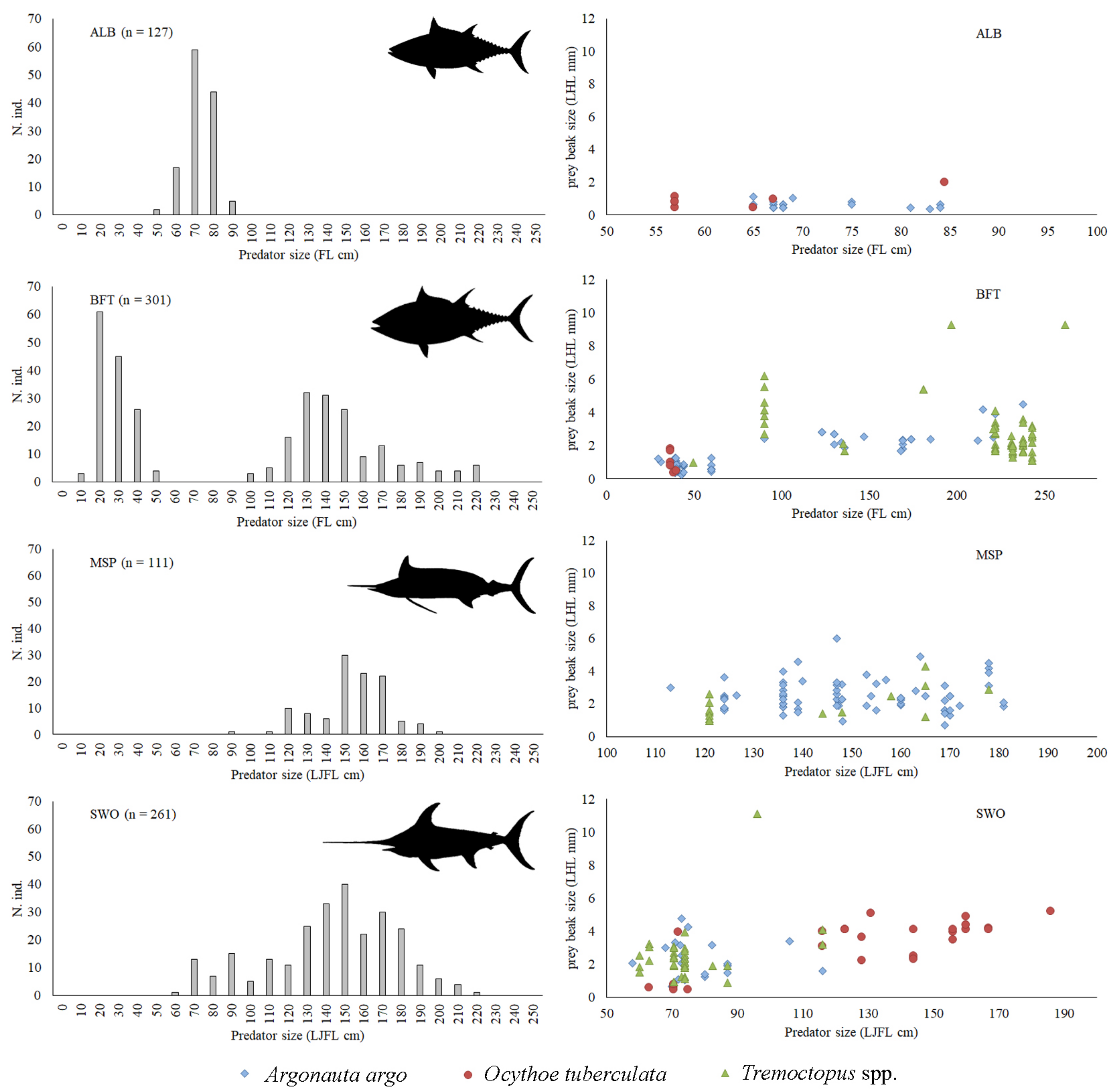
3.2. Occurrence of Holopelagic Octopuses in Predator Diets
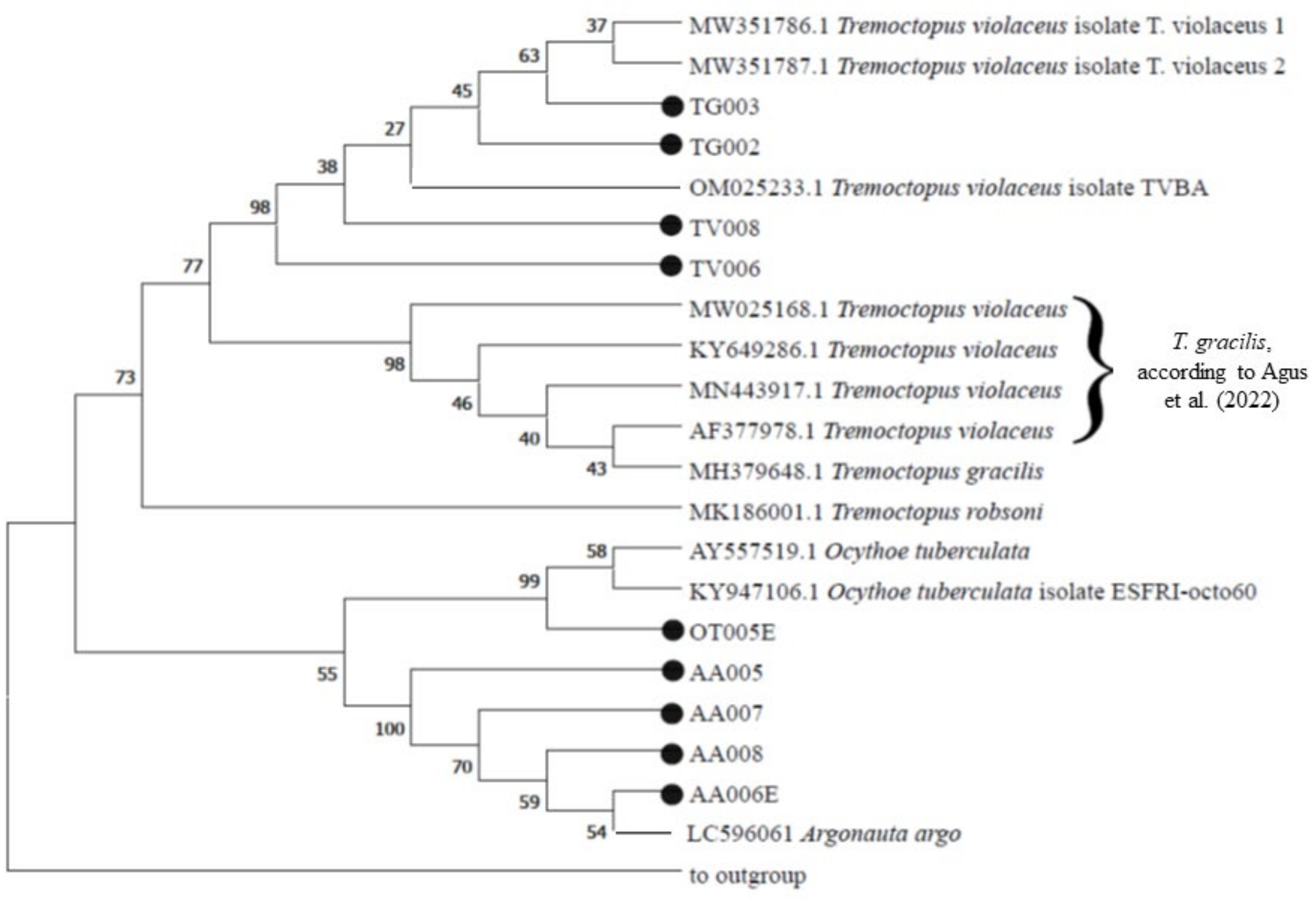
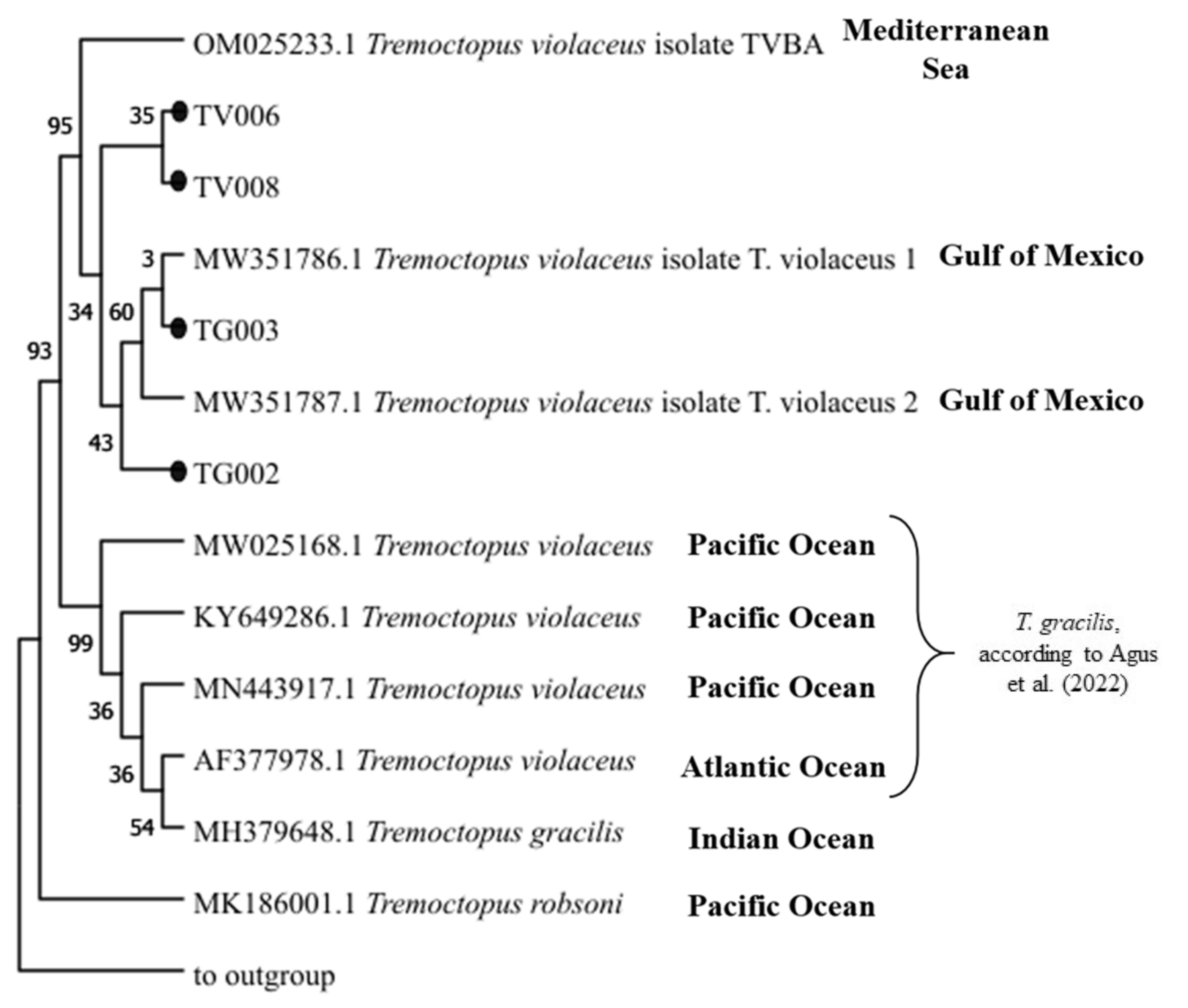
3.3. Genetic Analysis
4. Discussion
- (i)
- A different life span between sexes, since it has been hypothesized that the male may die after detaching the hectocotylized arm and delivering it to the female [68,69,70,71]. Therefore, the male may survive for a short period (just enough time to mature and reproduce); this hypothesis is supported by the continuous reproductive period, which may allow for the turnover of population;
- (ii)
- The female, after receiving the hectocotylus, could devour the male to obtain additional energy to complete egg maturation [14], thus reducing the possibility of sampling males;
- (iii)
- The very small size of males makes them very elusive to sampling.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MolluscaBase. Argonautoidea Cantraine. 1841. Available online: www.marinespecies.org/aphia.php?p=taxdetails&id=14673 (accessed on 26 January 2023).
- Belluscio, A.; Ardizzone, G.D.; Conticelli, M.; Pellicciari, C. Prima documentazione fotografica di una femmina ovigera di Tremoctopus sp. (Octopoda, Tremoctopodidae) nel Mediterraneo. Biol. Mar. Mediterr. 2004, 11, 556–559. [Google Scholar]
- Orsi Relini, L.; Belluscio, A.; Ardizzone, G.D. Tracking the Indopacific pelagic octopus Tremoctopus gracilis in the Mediterranean. Rapp. Comm. Int. Mer Médit. 2004, 37, 415. [Google Scholar]
- Kramer, G. Einige beobachtungen an Tremoctopus violaceus Delle Chiaje. Note Ist. It.-Germ. Biol. Mar. 1937, 25, 3–11. [Google Scholar]
- Bello, G.; Andaloro, F.; Battaglia, P. Non-indigenous cephalopods in the Mediterranean Sea: A review. Acta Adriat. 2020, 61, 113–134. [Google Scholar] [CrossRef]
- Orsi Relini, L. Notes about colour displays observed in female specimens of Tremoctopus (Cephalopoda: Octopoda) and their taxonomic value. Boll. Malacol. 2009, 45, 13–16. [Google Scholar]
- Guerra, A. Mollusca, Cephalopoda. In Fauna Ibérica; Ramos Sanchez, M.A., Tercedor, J.A., Belles i Ros, X., Gosalbez i Noguera, J., Guerra Sierra, A., Macpherson Mayol, E., Piera, F.M., Serrano Marino, J., Templado Gonzalez, J., Eds.; Museo Nacional De Ciencias Naturales, CSIC: Madrid, Spain, 1992; Volume 1. [Google Scholar]
- Bello, G. The biogeography of Mediterranean cephalopods. Biogeographia 2003, 24, 209–226. [Google Scholar] [CrossRef] [Green Version]
- Bello, G. On the purported representation of Haliphron atlanticus (Cephalopoda: Octopoda: Alloposidae) on Mycenaean funerary gold ornaments. Iberus 2020, 38, 213–218. [Google Scholar]
- Clarke, M.R. Beaks, nets and numbers. In The Biology of Cephalopods: Proceedings of the Symposia of the Zoological Society of London; Nixon, M., Messenger, J.B., Eds.; Academic Press: London, UK, 1977; Volume 38, pp. 89–126. [Google Scholar]
- Hoving, H.-J.; Christiansen, S.; Fabrizius, E.; Hauss, H.; Kiko, R.; Linke, P.; Neitzel, P.; Piatkowski, U.; Körtzinger, A. The pelagic in situ observation system (PELAGIOS) to reveal biodiversity, behavior, and ecology of elusive oceanic fauna. Ocean Sci. 2019, 15, 1327–1340. [Google Scholar] [CrossRef] [Green Version]
- Quetglas, A.; Ordines, F.; González, M.; Zaragoza, N.; Mallol, S.; Valls, M.; De Mesa, A. Uncommon pelagic and deep-sea cephalopods in the Mediterranean: New data and literature review. Mediterr. Mar. Sci. 2013, 14, 69–85. [Google Scholar] [CrossRef]
- Finn, J.K. Family Argonautidae. In Cephalopods of the World: An Annotated and Illustrated Catalogue of Cephalopod Species Known to Date; Jereb, P., Roper, C., Norman, M., Finn, J., Eds.; Vol. 3. Octopods and Vampire Squids; FAO Species Catalogue for Fishery Purposes: No. 4; FAO: Rome, Italy, 2014. [Google Scholar]
- Battaglia, P.; Stipa, M.G.; Ammendolia, G.; Pedà, C.; Consoli, P.; Andaloro, F.; Romeo, T. When nature continues to surprise: Observations of the hectocotylus of Argonauta argo, Linnaeus 1758. Eur. Zool. J. 2021, 88, 980–986. [Google Scholar] [CrossRef]
- Bello, G. Teutophagus predators as collectors of oceanic cephalopods: The case of the Adriatic Sea. Bolletino Malacol. 1996, 32, 71–78. [Google Scholar]
- Battaglia, P.; Pedà, C.; Sinopoli, M.; Romeo, T.; Andaloro, F. Cephalopods in the diet of young-of-the-year bluefin tuna (Thunnus thynnus L. 1758, Pisces: Scombridae) from the southern Tyrrhenian Sea (central Mediterranean Sea). Ital. J. Zool. 2013, 80, 560–565. [Google Scholar] [CrossRef] [Green Version]
- Lansdell, M.; Young, J. Pelagic cephalopods from eastern Australia: Species composition, horizontal and vertical distribution determined from the diets of pelagic fishes. Rev. Fish Biol. Fish. 2007, 17, 125–138. [Google Scholar] [CrossRef]
- Romeo, T.; Battaglia, P.; Pedà, C.; Perzia, P.; Consoli, P.; Esposito, V.; Andaloro, F. Pelagic cephalopods of the central Mediterranean Sea determined by the analysis of the stomach content of large fish predators. Helgol. Mar. Res. 2012, 66, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Pedà, C.; Battaglia, P.; Scuderi, A.; Voliani, A.; Mancusi, C.; Andaloro, F.; Romeo, T. Cephalopod prey in the stomach contents of odontocete cetaceans stranded in the western Mediterranean Sea. Mar. Biol. Res. 2015, 11, 593–602. [Google Scholar] [CrossRef]
- Xavier, J.C.; Walker, K.; Elliott, G.; Cherel, Y.; Thompson, D. Cephalopod fauna of south Pacific waters: New information from breeding New Zealand wandering albatrosses. Mar. Ecol. Prog. Ser. 2014, 513, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.M.; Paiva, V.H.; Xavier, J.C. Using seabirds to map the distribution of elusive pelagic cephalopod species. Mar. Ecol. Prog. Ser. 2017, 567, 257–262. [Google Scholar] [CrossRef]
- Battaglia, P.; Ammendolia, G.; Cavallaro, M.; Consoli, P.; Esposito, V.; Malara, D.; Rao, I.; Romeo, T.; Andaloro, F. Influence of lunar phases, winds and seasonality on the stranding of mesopelagic fish in the Strait of Messina (Central Mediterranean Sea). Mar. Ecol. 2017, 38, e12459. [Google Scholar] [CrossRef]
- Battaglia, P.; Stipa, M.G. New record of the football octopus Ocythoe tuberculata Rafinesque 1814, from the southern Tyrrhenian Sea (Italy). In: Tsangarakis, K., et al. New records of rare species in the Mediterranean Sea. Collective Article B. Mediterr. Mar. Sci. 2021, 22, 627–652. [Google Scholar] [CrossRef]
- Thomas, R.F. Systematics, distribution, and biology of cephalopods of the genus Tremoctopus (Octopoda: Tremoctopodidae). Bull. Mar. Sci. 1977, 27, 353–392. [Google Scholar]
- Finn, J.K. Taxonomy and biology of the argonauts (Cephalopoda: Argonautidae) with particular reference to Australian material. Molluscan Res. 2013, 33, 143–222. [Google Scholar] [CrossRef]
- Young, R.E.; Vecchione, M.; Mangold, K.M. Cephalopoda Cuvier 1797. Octopods, Squids, Nautiluses, etc. Version 26 March 2019 (under Construction). Available online: https://tolweb.org/ (accessed on 26 September 2022).
- Clarke, M.R. A Handbook for the Identification of Cephalopod Beaks; Clarendon Press: Oxford, UK, 1986; p. 273. ISBN 019857603X. [Google Scholar]
- Corsini, M.; Lefkaditou, E. Occurrence of Ocythoe tuberculata (Cephalopoda: Ocythoidae) in Greek waters. Boll. Malacol. Pubbl. Mens. Ed. Dalla Unione Malacol. Ital. 1994, 30, 304–310. [Google Scholar]
- Smale, M.J.; Clarke, M.R.; Klages, N.T.W.; Roeleveld, M.A.C. Octopod beak identification—Resolution at a regional level (Cephalopoda, Octopoda: Southern Africa). S. Afr. J. Mar. Sci. 1993, 13, 269–293. [Google Scholar] [CrossRef] [Green Version]
- Laptikhovsky, V.; Salman, A. On reproductive strategies of the epipelagic octopods of the superfamily Argonautoidea (Cephalopoda: Octopoda). Mar. Biol. 2003, 142, 321–326. [Google Scholar] [CrossRef]
- Tutman, P.; Šifner, S.K.; Dulčić, J.; Pallaoro, A.; Gavrilović, A.; Jug-Dujaković, J.; Glamuzina, B. A Note on the distribution and biology of Ocythoe tuberculata (Cephalopoda: Ocythoidae) in the Adriatic Sea. Vie Milieu/Life Environ. 2008, 58, 215–221. [Google Scholar]
- Mereu, M.; Agus, B.; Cau, A.L.; Cuccu, D. On a female of Tremoctopus sp. (Octopoda: Tremoctopodidae) caught in the Sardinian Sea. Biol. Mar. Mediterr. 2012, 19, 214. [Google Scholar]
- Salman, A.; Akalin, M. A rare pelagic cephalopod Ocythoe tuberculata (Octopoda: Argonautoidea): The record fecundity for Octopoda and new data on morphometry. Turk. J. Fish. Aquat. Sci. 2012, 12, 339–344. [Google Scholar] [CrossRef]
- Power, J. Observations Physiques sur le Poulpe de l’Argonauta Argo; Imprimerie Charles de Morgues Freres: Paris, France, 1856; p. 30. [Google Scholar]
- Finn, J.K. Recognising variability in the shells of argonauts (Cephalopoda: Argonautidae): The key to resolving the taxonomy of the family. Mem. Mus. Vic. 2018, 77, 63–104. [Google Scholar] [CrossRef]
- Lu, C.C.; Ickeringill, R. Cephalopod Beak Identification and Biomass Estimation Techniques: Tools for Dietary Studies of Southern Australian Finfishes; Museum Victoria Science Reports; Museum Victoria: Melbourne, Australia, 2002; Volume 6, pp. 1–65. [Google Scholar] [CrossRef] [Green Version]
- Pedà, C.; Battaglia, P.; Romeo, T.; Stipa, M.G.; Longo, F.; Malara, D.; Consoli, P.; Andaloro, F. Photographic Atlas of Cephalopod Beaks from the Mediterranean Sea; Peda, C., Battaglia, P., Romeo, T., Stipa, M.G., Longo, F., Malara, D., Consoli, P., Andaloro, F., Eds.; EBS Print: Lesmo, Italy, 2022; p. 120. ISBN 979-12-5968-638-1. [Google Scholar]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef] [PubMed]
- Agus, B.; Carbonara, P.; Melis, R.; Cannas, R.; Carugati, L.; Cera, J.; Donnaloia, M.; Mulas, A.; Pais, A.; Ruiu, S. First integrative morphological and genetic characterization of Tremoctopus violaceus sensu stricto in the Mediterranean Sea. Animals 2022, 12, 80. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Guerra, A.; González, A.F.; Rocha, F. Appearance of the common paper nautilus Argonauta argo related to the increase of the sea surface temperature in the north-eastern Atlantic. J. Mar. Biol. Assoc. U. K. 2002, 82, 855–858. [Google Scholar] [CrossRef] [Green Version]
- Corsini-Foka, M.; Lorenzini, P.; Tirrò, F.; Sottile, G.; Kondylatos, G. On the recent occurrence of females Argonauta argo (Mollusca: Cephalopoda) in the Adriatic and Aegean Seas with observations on aquarium maintenance. Vie Milieu-Life Environ. 2011, 61, 35–39. [Google Scholar]
- Kim, Y.; Oh, T.Y.; Park, K.J.; An, Y.R.; Kim, H.W.; Kim, D.N.; An, D. Morphological and brooding characteristics of Argonauta argo Linnaeus, 1758 from Korean water. Korean J. Malacol. 2014, 30, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Giacalone, V.M.; Micalizzi, R.; D’Anna, G. A rare occurrence of Argonauta argo (Linnaeus, 1758) (Cephalopoda: Octopoda) off the shores of western Sicily with observations in aquarium. Not. S.I.M. 2015, 33, 11–13. [Google Scholar]
- Cardoso, F.; Paredes, C. La familia Ocythoidae (Cephalopoda: Octopoda) en el Perú. Rev. Peru. Biol. 1998, 5, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Lefkaditou, E.; Kallianiotis, A. New findings of female Ocythoe tuberculata in the northeastern Mediterranean. In Proceedings of the Cephalopod International Advisory Council Symposium 2006 (CIAC ‘06), Hobart, Tasmania, 6–10 February 2006; p. 84. [Google Scholar]
- Berdar, A.; Potoschi, A.; Cavallaro, G.; Cavaliere, A.; Li Greci, F. Su alcuni cefalopodi spiaggiati e pescati nello Stretto di Messina. Mem. Biol. Mar. Oceanogr. 1983, 13, 115–127. [Google Scholar]
- Berdar, A.; Cavallaro, G. Ulteriore contributo alla conoscenza dei cefalopodi spiaggiati lungo la costa siciliana dello Stretto di Messina. Mem. Biol. Mar. Ocean. 1975, 5, 121–138. [Google Scholar]
- Giordano, D.; Busalacchi, B.; Liguori, M.; Perdichizzi, F.; De Domenico, E.; Profeta, A. Occurrence of Ocythoe Tuberculata (Cephalopoda: Ocythoidae) in the Strait of Messina (Central Mediterranean). Rapp. Comm. Int. Mer. Médit. 2010, 39, 530. [Google Scholar]
- Müller, H. Note sur les argonautes male ei les hectocotyles. Ann. Sci. Nat. Zool. 1851, 16, 132–134. [Google Scholar]
- Naef, A. Cephalopoda. In Fauna und Flora des Golfes von Neapel. Monograph, No. 35. English Translation: A. Mercado (1972); IPST Cat. No. 5110/1,2; Israel Program for Scientific Translations Ltd.: Jerusalem, Israel, 1921. [Google Scholar]
- Jatta, G. I Cefalopodi. 23. Monographie. Atlante di 31 Tavole. In Fauna und Flora des Golfes von Neapel und der Angrenzenden Meeres-Abschnitte, Stazione Zoologica di Napoli; R. Friedländer & Sohn: Berlin, Germany, 1896. [Google Scholar]
- Bello, G. Exaptations in argonautoidea (cephalopoda: Coleoidea: Octopoda). Neues Jahrb. Geol. Palaontol.-Abh. 2012, 266, 85–92. [Google Scholar] [CrossRef]
- Kölliker, A., II. Some observations upon the structure of two new species of hectocotyle, parasitic upon Tremoctopus violaceus, D.Ch. and Argonauta argo, Linn.; with an exposition of the hypothesis that these hectocotylæ are the males of the Cephalopoda upon which they are found. Trans. Linn. Soc. London 1846, os-20, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Tsagarakis, K.; Darmanin, S.A.; Al Mabruk, S.A.A.; Auriemma, R.; Azzurro, E.; Badouvas, N.; Bakiu, R.; Bariche, M.; Battaglia, P.; Betti, F. New records of rare species in the Mediterranean Sea (October 2021). Mediterr. Mar. Sci. 2021, 22, 627–652. [Google Scholar] [CrossRef]
- Nishimura, S. Glimpse of the biology of Argonauta argo Linnaeus (Cephalopoda: Octopodida) in the Japanese waters. Publ. Seto Mar. Biol. Lab. 1968, 16, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Rocha, F.; Guerra, Á.; González, Á. A review of reproductive strategies in cephalopods. Biol. Rev. 2001, 76, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Hamabe, M. Egg mass and newborns of Tremoctopus violaceus Delle Chiaje, caught in the harbour of Kasumi, Hyogo Prefecture. Bull. Tokai Reg. Fish. Res. Lab. 1973, 72, 1–5. [Google Scholar]
- Monterosato, T.A. Coquilles marines Marocaines. J. Conchyliol. 1889, 37, 20–40. [Google Scholar]
- Battaglia, P.; Andaloro, F.; Consoli, P.; Esposito, V.; Malara, D.; Musolino, S.; Pedà, C.; Romeo, T. Feeding habits of the Atlantic bluefin tuna, Thunnus thynnus (L. 1758), in the central Mediterranean Sea (Strait of Messina). Helgol. Mar. Res. 2013, 67, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Voss, G.L. A contribution to the life history and biology of the sailfish, Istiophorus americanus Cuv. and Val., in Florida waters. Bull. Mar. Sci. 1953, 3, 206–240. [Google Scholar]
- Bello, G. Tremoctopus violaceus (Cephalopoda: Tremoctopodidae) in the stomach content of a swordfish from the Adriatic Sea. Boll. Malacol. 1993, 29, 45–48. [Google Scholar]
- Romeo, T.; Consoli, P.; Castriota, L.; Andaloro, F. An evaluation of resource partitioning between two billfish, Tetrapturus belone and Xiphias gladius, in the central Mediterranean Sea. J. Mar. Biol. Assoc. U. K. 2009, 89, 849–857. [Google Scholar] [CrossRef]
- Bello, G. How rare is Histioeuthis bonnellii (Cephalopoda: Histioteuthidae) in the eastern Mediterranean Sea? J. Molluscan Stud. 2000, 66, 575–576. [Google Scholar] [CrossRef] [Green Version]
- Nesis, K.N. The biology of paper nautiluses, Argonauta boettgeri and A. hians (Cephalopoda, Octopoda), in the western Pacific and the seas of the east Indian Archipelago. Zool. Zhurnal (Russ. Zool. J.) 1977, 56, 1004–1014. [Google Scholar]
- Iliffe, T.M. Argonaut: Octopus in a parchment shell. Sea Front. 1982, 28, 224–228. [Google Scholar]
- Orenstein, M.; Wood, B. Paper nautilus (Argonauta argo) Marine Invertebrates of Bermuda. Available online: http://www.thecephalopodpage.org/MarineInvertebrateZoology/Argonautaargo.html (accessed on 18 January 2021).
- Norman, M. Cephalopods—A World Guide: Octopuses, Argonauts, Cuttlefish, Squid, Nautilus; Conch Books: Harxheim, Germany, 2003; p. 320. ISBN 3-92591-32-5. [Google Scholar]
- Melis, R.; Vacca, L.; Cuccu, D.; Mereu, M.; Cau, A.; Follesa, M.C.; Cannas, R. Genetic population structure and phylogeny of the common octopus Octopus vulgaris Cuvier, 1797 in the western Mediterranean Sea through nuclear and mitochondrial markers. Hydrobiologia 2018, 807, 277–296. [Google Scholar] [CrossRef]
- Jiménez-Badillo, M.d.L.; Meiners-Mandujano, C.; Galindo-Cortes, G.; Morillo-Velarde, P.S.; González-Gómez, R.; de los Angeles Barriga-Sosa, I.; Pliego-Cárdenas, R. The first record of Tremoctopus violaceus sensu stricto Delle Chiaje, 1830 in southwestern Gulf of Mexico gives a hint of the taxonomic status of Tremoctopus gracilis. Zookeys 2021, 1012, 55. [Google Scholar] [CrossRef]
- Dai, L.; Zheng, X.; Kong, L.; Li, Q.I. DNA barcoding analysis of Coleoidea (Mollusca: Cephalopoda) from Chinese waters. Mol. Ecol. Resour. 2012, 12, 437–447. [Google Scholar] [CrossRef]
- Hirota, K.; Yoshida, M.; Itoh, T.; Toyoda, A.; Setiamarga, D.H.E. The full mitochondrial genome sequence of the greater argonaut Argonauta argo (Cephalopoda, Argonautoidea) and its phylogenetic position in Octopodiformes. Mitochondrial DNA Part B 2021, 6, 1451–1453. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.A.; Hirota, K.; Imoto, J.; Okuno, M.; Tanaka, H.; Kajitani, R.; Toyoda, A.; Itoh, T.; Ikeo, K.; Sasaki, T. Gene recruitments and dismissals in the argonaut genome provide insights into pelagic lifestyle adaptation and shell-like eggcase reacquisition. Genome Biol. Evol. 2022, 14, evac140. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.W.; Chang, C.W.; Shen, K.N.; Ju, Y.M.; Lin, H.D. Complete mitochondrial genome and the phylogenetic position of the pelagic octopus Tremoctopus violaceus (Mollusca: Tremoctopodidae). Mitochondrial DNA Part B 2018, 3, 1248–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AA.VV. Valutazione dello stato e della Distribuzione dell’aguglia Imperiale (Tetrapturus belone). Relazione Finale, Misura a Titolarità 1.40 Lettere c) e i) del PO FEAMP 2014/2020. 2021. Available online: https://www.regione.sicilia.it/sites/default/files/2022-03/Elenco%20misura%201.40%20al%2031.12.2021%20-%20anno%202021.pdf (accessed on 15 October 2022).
- Carlini, D.B.; Young, R.E.; Vecchione, M. A molecular phylogeny of the Octopoda (Mollusca: Cephalopoda) evaluated in light of morphological evidence. Mol. Phylogenet. Evol. 2001, 21, 388–397. [Google Scholar] [CrossRef]
- Carrasco, S.A.; Varela, A.I.; Ibáñez, C.M.; Sellanes, J.; Thiel, M. Paralarval and juvenile stages as a proxy for cephalopod diversity in the Juan Fernández and Desventuradas ecoregion, southeast Paific Ocean. Bull. Mar. Sci. 2020, 96, 263–280. [Google Scholar] [CrossRef]
- Braid, H.E.; Bolstad, K.S. Cephalopod biodiversity of the Kermadec Islands: Implications for conservation and some future taxonomic priorities. Invertebr. Syst. 2019, 33, 402–425. [Google Scholar]
- Lindgren, A.R.; Giribet, G.; Nishiguchi, M.K. A combined approach to the phylogeny of Cephalopoda (Mollusca). Cladistics 2004, 20, 454–486. [Google Scholar] [CrossRef]
- Kim, J.B.; Yang, J.H.; Kim, Y.; Lee, S.J. First record of a rare pelagic octopod, Ocythoe tuberculata (Cephalopoda: Ocythoidae), in Korea. Anim. Cells Syst. 2017, 21, 419–423. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | AN | Taxonomy | Sim | Taxonomy | Sim |
|---|---|---|---|---|---|
| BLASTn Search Implemented in GenBank | (%) | BOLD-IDS | (%) | ||
| Search | |||||
| AA005 | - | nd | nd | nd | nd |
| AA015 | - | nd | nd | nd | nd |
| AA015E | - | nd | nd | nd | nd |
| AA006 | - | nd | nd | nd | nd |
| AA006E | - | nd | nd | nd | nd |
| AA006U | - | nd | nd | nd | nd |
| AA007 | - | nd | nd | nd | nd |
| AA008 | - | nd | nd | nd | nd |
| OT005 | - | nd | nd | nd | nd |
| OT005E | OP132818 | AY557519, Ocythoe tuberculata | 100 | Ocythoe tuberculata, KY947106 | 100 |
| TG002 | OP132821 | OM025233, Tremoctopus violaceus isolate TVBA | 98 | Tremoctopus | 97 |
| violaceus, private sequence | |||||
| TG002U | - | nd | nd | nd | nd |
| TG003 | - | nd | nd | nd | nd |
| TV006 | OP132819 | OM025233, Tremoctopus violaceus isolate TVBA | 98 | Tremoctopus | 85 |
| violaceus, private sequence | |||||
| TV008 | - | nd | nd | nd | nd |
| TV011 | OP132820 | OM025233, Tremoctopus violaceus isolate TVBA | 99 | Tremoctopus | 99 |
| violaceus, private sequence |
| Sample | AN | Taxonomy | Sim | Taxonomy | Sim |
|---|---|---|---|---|---|
| BLASTn Search Implemented in GenBank | (%) | BOLD-IDS | (%) | ||
| Search | |||||
| AA005 | OP132805 | LC596061, Argonauta argo UMUT:RM33391 | 100 | BOLD:AAJ8256, Argonauta argo | 99.8 |
| AA015 | OP132806 | LC596061, Argonauta argo UMUT:RM33391 | 100 | Argonauta | 100 |
| hians, private sequence | |||||
| AA015E | - | nd | nd | nd | nd |
| AA006 | OP132807 | AB191273, Argonauta argo mitochondrial COI gene | 100 | BOLD:AAJ8256, Argonauta argo | 89 |
| AA006E | OP132808 | LC596061, Argonauta argo UMUT:RM33391 | 100 | Argonauta | 100 |
| hians, private sequence | |||||
| AA006U | - | nd | |||
| AA007 | OP132809 | LC596061, Argonauta argo UMUT:RM33391 | 100 | BOLD:AAJ8256, Argonauta argo | 99.8 |
| AA008 | OP132810 | LC596061, Argonauta argo UMUT:RM33391 | 100 | Argonauta | 99.8 |
| hians, private sequence | |||||
| OT005 | OP132811 | KY947106, Ocythoe tuberculata isolate ESFRI-octo60 | 99 | nd | nd |
| OT005E | OP132812 | AY557519, Ocythoe tuberculata cytochrome c oxidase subunit I gene | 100 | BOLD:ADR2058, Ocythoe tuberculata | 100 |
| TG002 | OP132816 | MW351787, Tremoctopus violaceus isolate T. violaceus 2 | 99 | Tremoctopus violaceus, private sequence | 100 |
| TG002U | - | nd | nd | nd | nd |
| TG003 | OP132817 | MW351787, Tremoctopus violaceus isolate T. violaceus 2 | 100 | nd | nd |
| TV006 | OP132813 | MW351787, Tremoctopus violaceus isolate T. violaceus 2 | 98 | nd | nd |
| TV008 | OP132814 | OM025233, Tremoctopus violaceus isolate TVBA | 100 | Tremoctopus violaceus, private sequence | 97 |
| TV011 | OP132815 | MW351787, Tremoctopus violaceus isolate T. violaceus 2 | 99 | nd | nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Battaglia, P.; Pedà, C.; Rizzo, C.; Stipa, M.G.; Arcadi, E.; Longo, F.; Ammendolia, G.; Cavallaro, M.; Rao, I.; Villari, A.; et al. How Rare Are Argonautoidea Octopuses in the Mediterranean? New Data from Stranding Events, Stomach Contents and Genetics. Biology 2023, 12, 420. https://doi.org/10.3390/biology12030420
Battaglia P, Pedà C, Rizzo C, Stipa MG, Arcadi E, Longo F, Ammendolia G, Cavallaro M, Rao I, Villari A, et al. How Rare Are Argonautoidea Octopuses in the Mediterranean? New Data from Stranding Events, Stomach Contents and Genetics. Biology. 2023; 12(3):420. https://doi.org/10.3390/biology12030420
Chicago/Turabian StyleBattaglia, Pietro, Cristina Pedà, Carmen Rizzo, Maria Giulia Stipa, Erika Arcadi, Francesco Longo, Giovanni Ammendolia, Mauro Cavallaro, Ignazio Rao, Alberto Villari, and et al. 2023. "How Rare Are Argonautoidea Octopuses in the Mediterranean? New Data from Stranding Events, Stomach Contents and Genetics" Biology 12, no. 3: 420. https://doi.org/10.3390/biology12030420