
Cumulus Cell Transcriptome after Cumulus-Oocyte Complex Exposure to Nanomolar Cadmium in an In Vitro Animal Model of Prepubertal and Adult Age
 , ,
, ,  , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
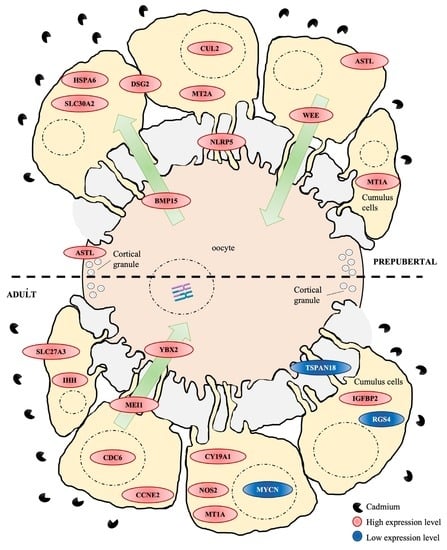
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Collection of Ovaries
2.3. In Vitro Maturation (IVM)
2.4. Cumulus Cells Isolation from Matured Oocytes
2.5. Oocyte Nuclear Chromatin Evaluation by Epifluorescence Microscopy
2.6. RNA-Seq and Data Analysis
2.7. Gene Network Analysis
2.8. RNA-Seq Data Validation by Quantitative RT-PCR
2.9. Statistical Analysis
2.10. Gene Annotation and Literature Search
3. Results
3.1. Nanomolar Cd Does Not Affect Meiotic Progression of Sheep Oocytes
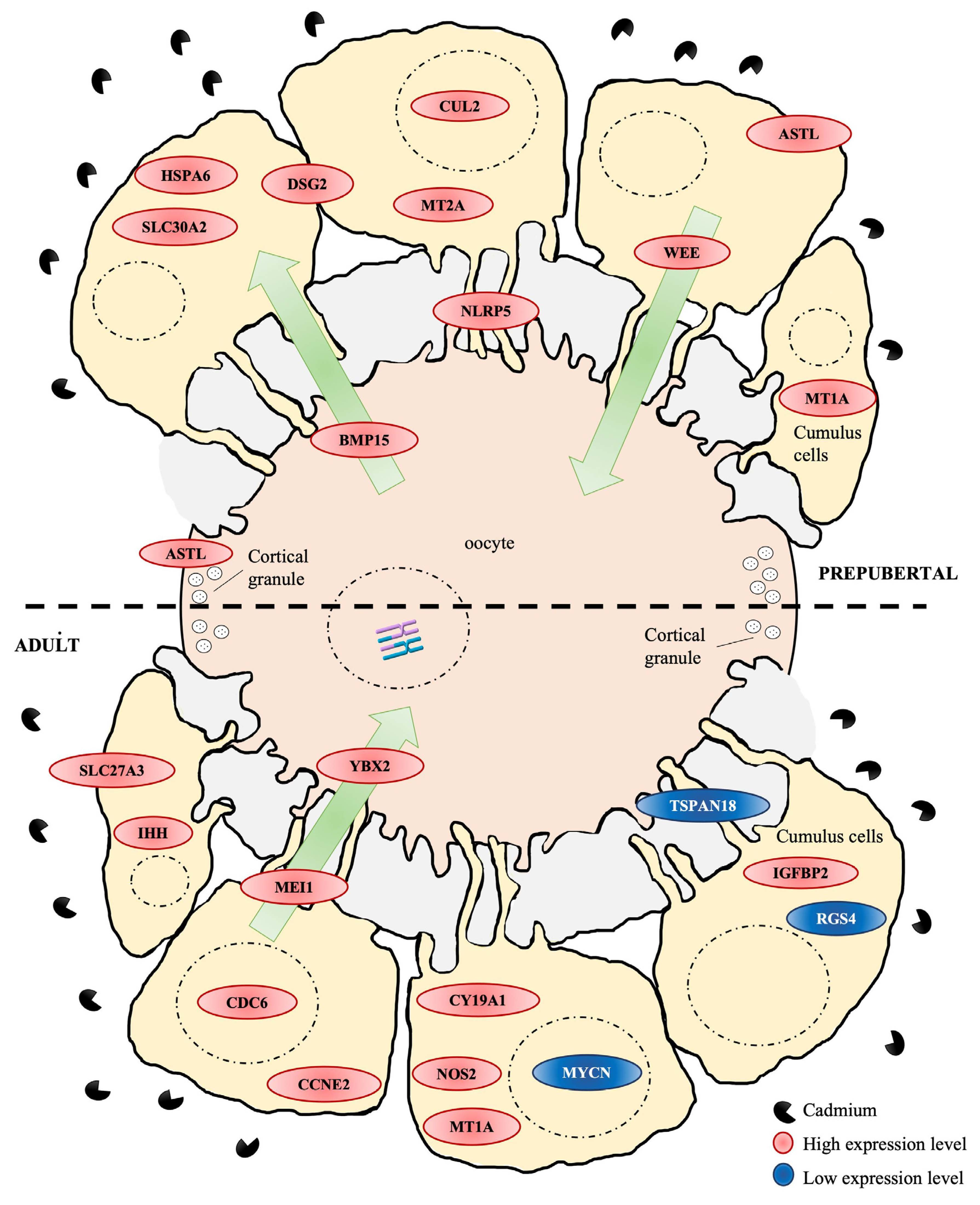
3.2. PCA Shows Relevant Cd-Induced Effects on Transcriptomic Profile of CCs from Adult Sheep
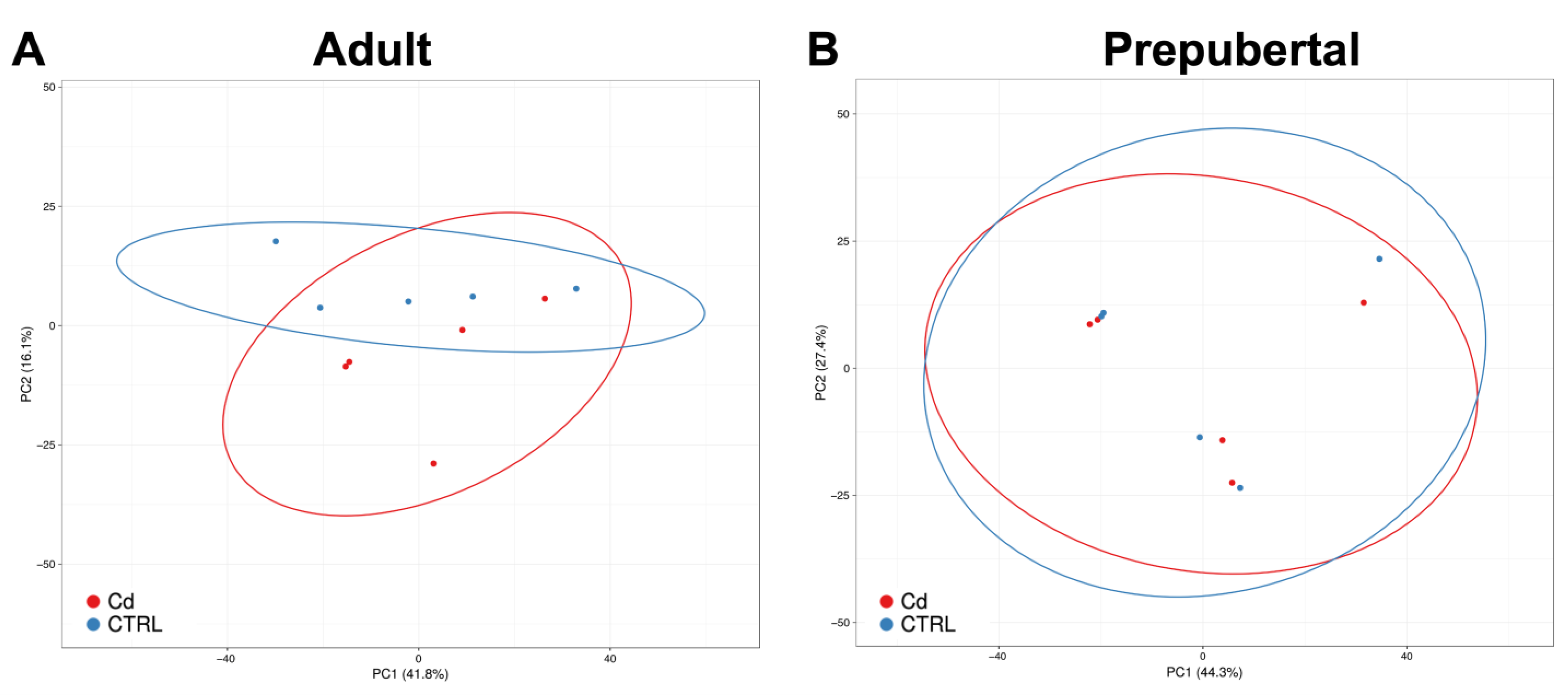
3.3. Cadmium Induces Higher Number of DEGs in CCs from Adult versus Prepubertal Sheep
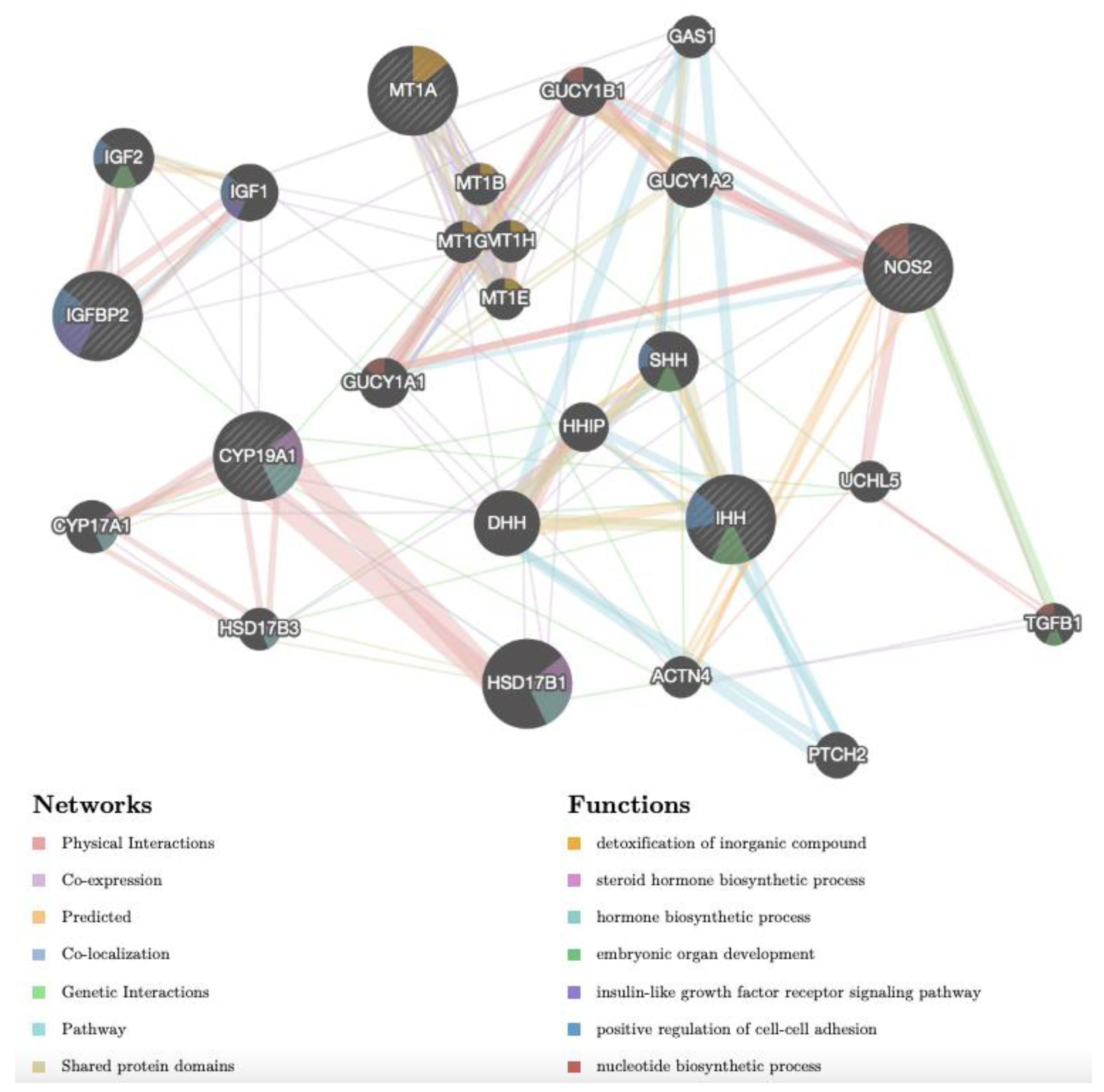
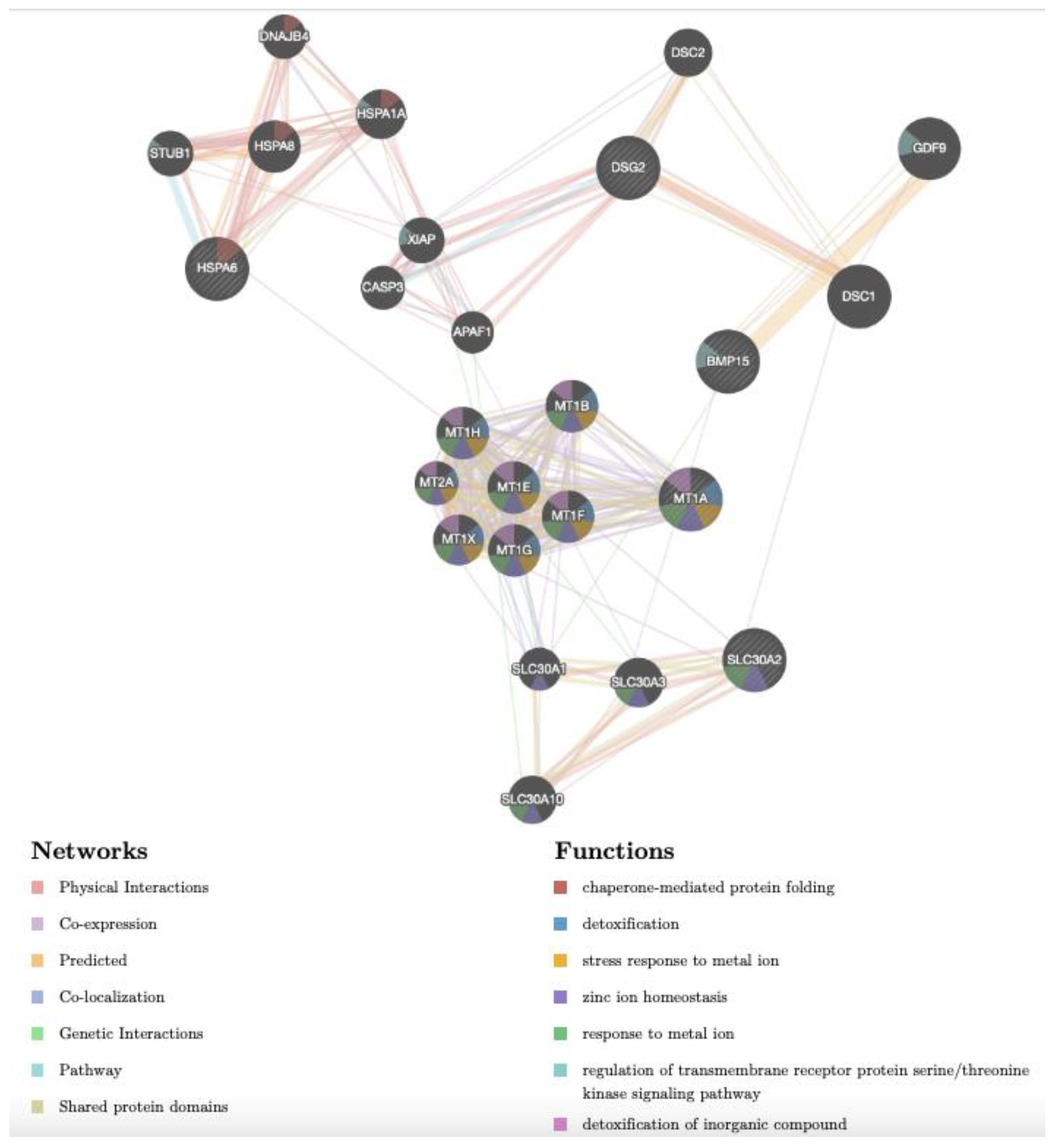
3.4. Gene Networks Identifies Different Cd-Induced Pathways in Adult versus Prepubertal CCs
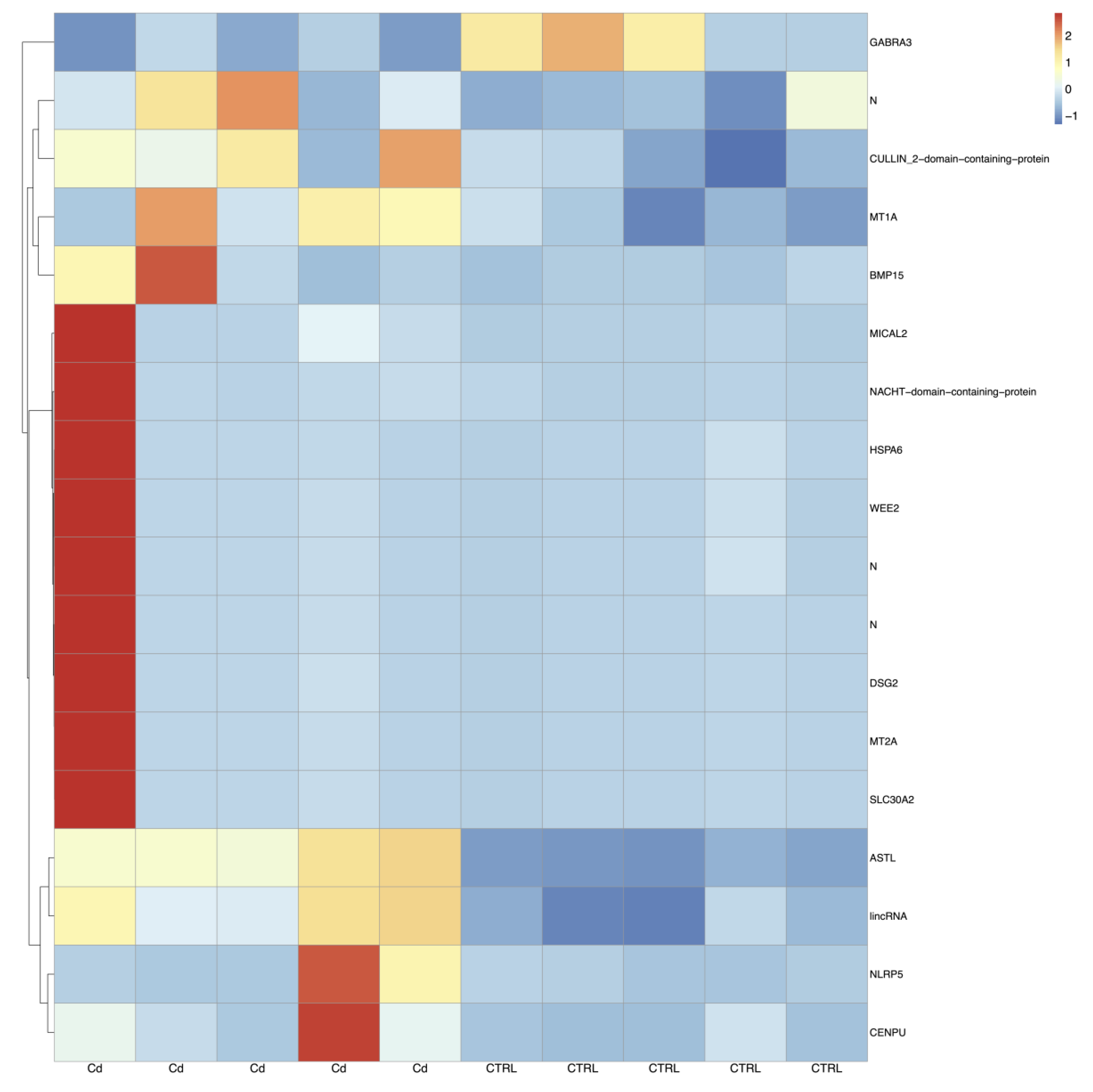
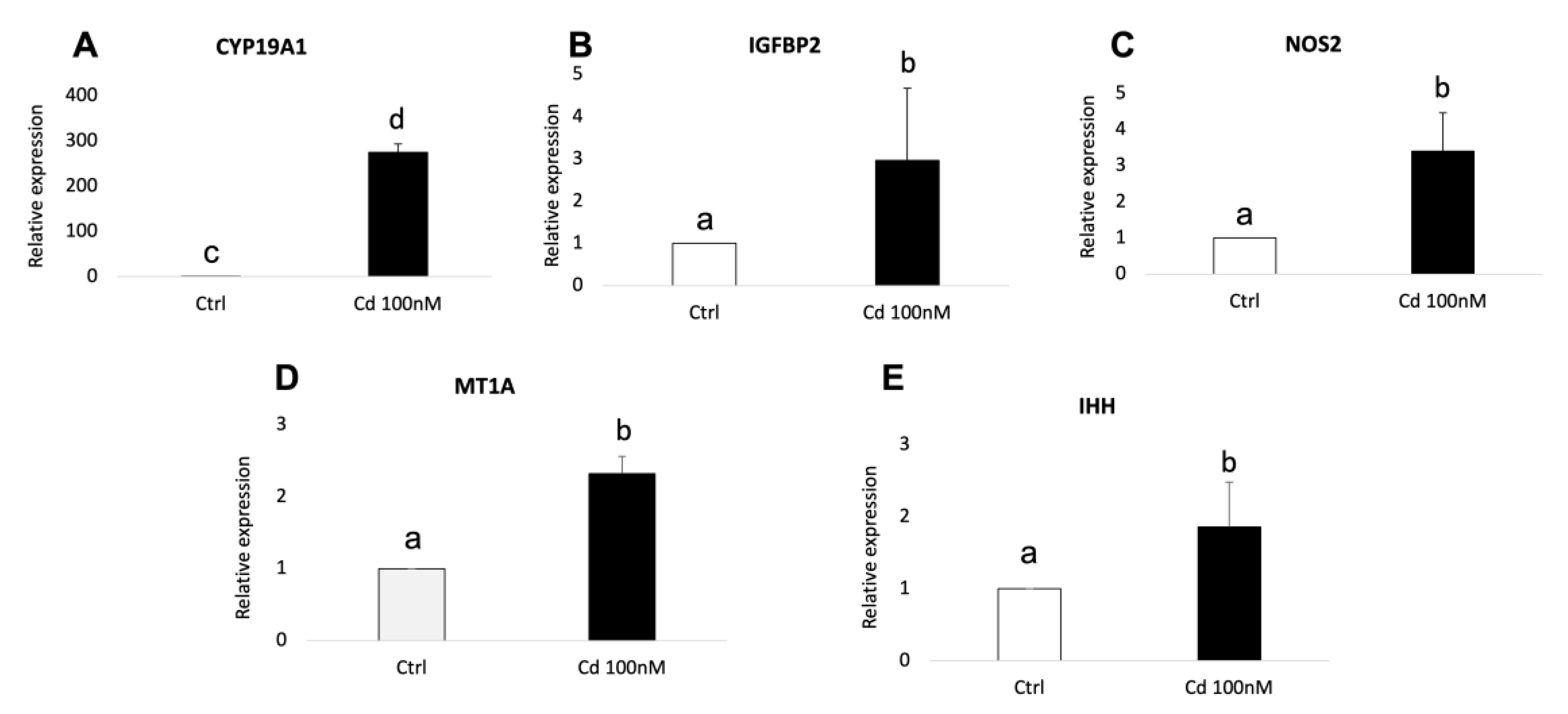
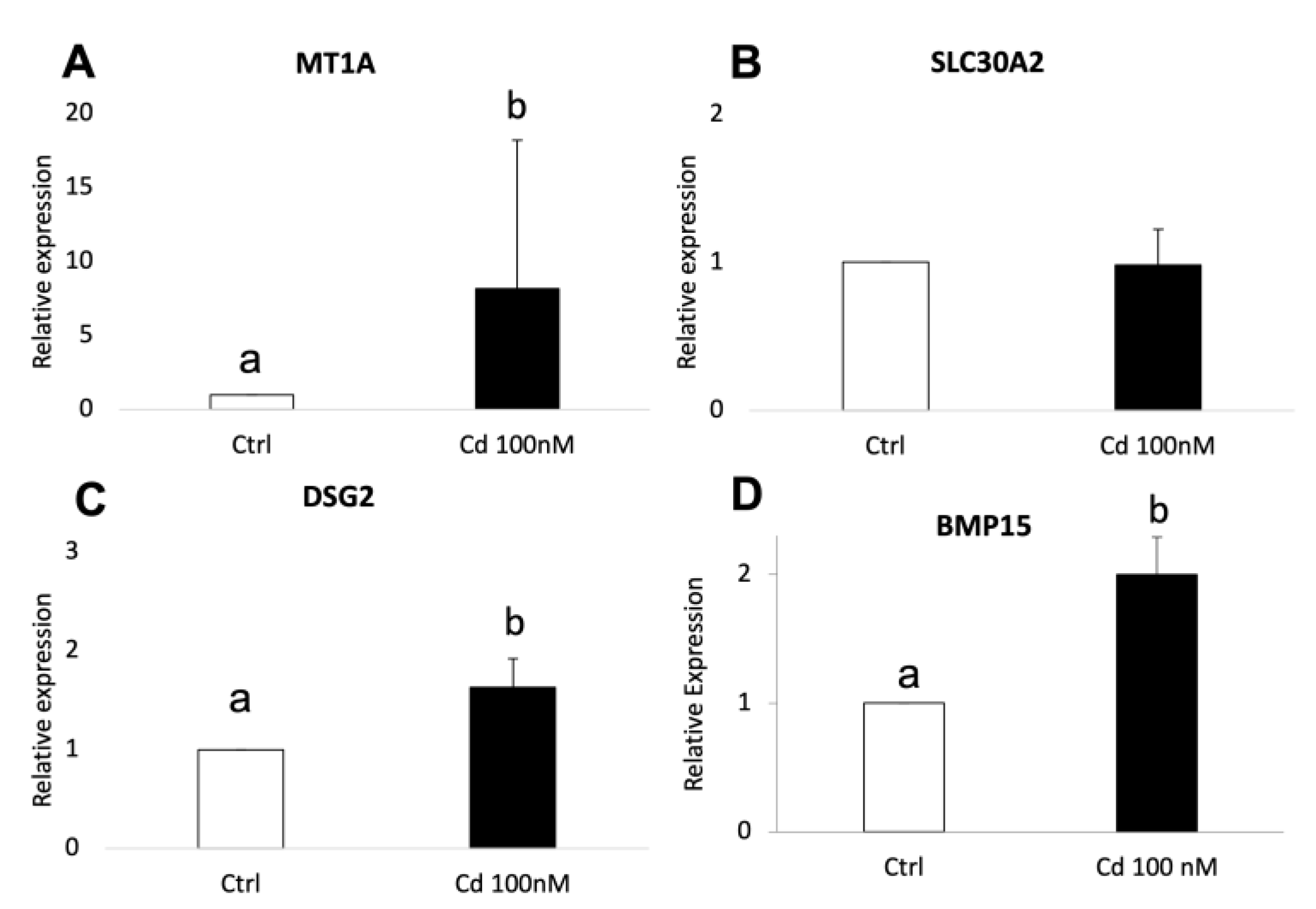
3.5. Cd-Induced CC Expression Pattern Is Consistent with the Sequencing Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Canipari, R.; De Santis, L.; Cecconi, S. Female Fertility and Environmental Pollution. Int. J. Environ. Res. Public Health 2020, 17, 8802. [Google Scholar] [CrossRef] [PubMed]
- Wrzecińska, M.; Kowalczyk, A.; Cwynar, P.; Czerniawska-Piątkowska, E. Disorders of the Reproductive Health of Cattle as a Response to Exposure to Toxic Metals. Biology 2021, 10, 882. [Google Scholar] [CrossRef]
- Rzymski, P.; Tomczyk, K.; Rzymski, P.; Poniedziałek, B.; Opala, T.; Wilczak, M. Impact of Heavy Metals on the Female Reproductive System. Ann. Agric. Environ. Med. 2015, 22, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Bannigan, J. Cadmium: Toxic Effects on the Reproductive System and the Embryo. Reprod. Toxicol. 2008, 25, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.S.; Agrawal, S. Arsenic, Cadmium, and Lead. In Reproductive and Developmental Toxicology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 537–566. [Google Scholar]
- Everson, T.M.; Punshon, T.; Jackson, B.P.; Hao, K.; Lambertini, L.; Chen, J.; Karagas, M.R.; Marsit, C.J. Cadmium-Associated Differential Methylation throughout the Placental Genome: Epigenome-Wide Association Study of Two U.S. Birth Cohorts. Environ. Health Perspect. 2018, 126, 017010. [Google Scholar] [CrossRef]
- Zhu, J.-Q.; Liu, Y.; Zhang, J.-H.; Liu, Y.-F.; Cao, J.-Q.; Huang, Z.-T.; Yuan, Y.; Bian, J.-C.; Liu, Z.-P. Cadmium Exposure of Female Mice Impairs the Meiotic Maturation of Oocytes and Subsequent Embryonic Development. Toxicol. Sci. 2018, 164, 289–299. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, J.; Wu, T.; Jiang, X.; Jia, H.; Qing, S.; An, Q.; Zhang, Y.; Su, J. Reproductive Toxicity of Acute Cd Exposure in Mouse: Resulting in Oocyte Defects and Decreased Female Fertility. Toxicol. Appl. Pharmacol. 2019, 379, 114684. [Google Scholar] [CrossRef]
- Dong, F.; Li, J.; Lei, W.-L.; Wang, F.; Wang, Y.; Ouyang, Y.-C.; Hou, Y.; Wang, Z.-B.; Schatten, H.; Sun, Q.-Y. Chronic Cadmium Exposure Causes Oocyte Meiotic Arrest by Disrupting Spindle Assembly Checkpoint and Maturation Promoting Factor. Reprod. Toxicol. 2020, 96, 141–149. [Google Scholar] [CrossRef]
- Ruslee, S.S.; Zaid, S.S.M.; Bakrin, I.H.; Goh, Y.M.; Mustapha, N.M. Protective Effect of Tualang Honey against Cadmium-Induced Morphological Abnormalities and Oxidative Stress in the Ovary of Rats. BMC Complement. Med. Ther. 2020, 20, 160. [Google Scholar] [CrossRef]
- Tian, J.; Hu, J.; Liu, D.; Yin, J.; Chen, M.; Zhou, L.; Yin, H. Cadmium Chloride-Induced Transgenerational Neurotoxicity in Zebrafish Development. Environ. Toxicol. Pharmacol. 2021, 81, 103545. [Google Scholar] [CrossRef]
- Zenzes, M.T.; Krishnan, S.; Krishnan, B.; Zhang, H.; Casper, R.F. Cadmium Accumulation in Follicular Fluid of Women in in Vitro Fertilization-Embryo Transfer Is Higher in Smokers. Fertil. Steril. 1995, 64, 599–603. [Google Scholar] [CrossRef]
- Piasek, M.; Blanuša, M.; Kostial, K.; Laskey, J.W. Placental Cadmium and Progesterone Concentrations in Cigarette Smokers. Reprod. Toxicol. 2001, 15, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Wdowiak, A.; Wdowiak, E.; Bojar, I. Evaluation of Trace Metals in Follicular Fluid in ICSI-Treated Patients. Ann. Agric. Environ. Med. 2018, 25, 213–218. [Google Scholar] [CrossRef]
- Wu, S.; Wang, M.; Deng, Y.; Qiu, J.; Zhang, X.; Tan, J. Associations of Toxic and Essential Trace Elements in Serum, Follicular Fluid, and Seminal Plasma with In Vitro Fertilization Outcomes. Ecotoxicol. Environ. Saf. 2020, 204, 110965. [Google Scholar] [CrossRef] [PubMed]
- Martino, N.A.; Marzano, G.; Mangiacotti, M.; Miedico, O.; Sardanelli, A.M.; Gnoni, A.; Lacalandra, G.M.; Chiaravalle, A.E.; Ciani, E.; Bogliolo, L.; et al. Exposure to Cadmium during in Vitro Maturation at Environmental Nanomolar Levels Impairs Oocyte Fertilization through Oxidative Damage: A Large Animal Model Study. Reprod. Toxicol. 2017, 69, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Leoni, G.; Bogliolo, L.; Deiana, G.; Berlinguer, F.; Rosati, I.; Pintus, P.P.; Ledda, S.; Naitana, S. Influence of Cadmium Exposure on in Vitro Ovine Gamete Dysfunction. Reprod. Toxicol. 2002, 16, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Mlynarcíková, A.; Scsuková, S.; Vrsanská, S.; Nagyová, E.; Ficková, M.; Kolena, J. Inhibitory Effect of Cadmium and Tobacco Alkaloids on Expansion of Porcine Oocyte-Cumulus Complexes. Cent. Eur. J. Public Health 2004, 12, S62–S64. [Google Scholar] [PubMed]
- Lazzari, G.; Tessaro, I.; Crotti, G.; Galli, C.; Hoffmann, S.; Bremer, S.; Pellizzer, C. Development of an in Vitro Test Battery for Assessing Chemical Effects on Bovine Germ Cells under the ReProTect Umbrella. Toxicol. Appl. Pharmacol. 2008, 233, 360–370. [Google Scholar] [CrossRef]
- Nandi, S.; Gupta, P.S.P.; Selvaraju, S.; Roy, S.C.; Ravindra, J.P. Effects of Exposure to Heavy Metals on Viability, Maturation, Fertilization, and Embryonic Development of Buffalo (Bubalus bubalis) Oocytes In Vitro. Arch. Environ. Contam. Toxicol. 2010, 58, 194–204. [Google Scholar] [CrossRef]
- Tessaro, I.; Modina, S.C.; Crotti, G.; Franciosi, F.; Colleoni, S.; Lodde, V.; Galli, C.; Lazzari, G.; Luciano, A.M. Transferability and Inter-Laboratory Variability Assessment of the in Vitro Bovine Oocyte Fertilization Test. Reprod. Toxicol. 2015, 51, 106–113. [Google Scholar] [CrossRef]
- Akar, Y.; Ahmad, N.; Khalıd, M. The Effect of Cadmium on the Bovine in Vitro Oocyte Maturation and Early Embryo Development. Int. J. Vet. Sci. Med. 2018, 6, S73–S77. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, X.; Chen, Y.; Liu, X.; Sun, Y.; Xiong, B. Glutathione Alleviates the Cadmium Exposure-Caused Porcine Oocyte Meiotic Defects via Eliminating the Excessive ROS. Environ. Pollut. 2019, 255, 113194. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, R.B.; Ritter, L.J.; Armstrong, D.T. Oocyte–Somatic Cell Interactions during Follicle Development in Mammals. Anim. Reprod. Sci. 2004, 82, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Burns, K.H.; Viveiros, M.M.; Eppig, J.J. Intercellular Communication in the Mammalian Ovary: Oocytes Carry the Conversation. Science 2002, 296, 2178–2180. [Google Scholar] [CrossRef] [PubMed]
- Bunel, A.; Jorssen, E.P.; Merckx, E.; Leroy, J.L.; Bols, P.E.; Sirard, M.A. Individual Bovine in Vitro Embryo Production and Cumulus Cell Transcriptomic Analysis to Distinguish Cumulus-Oocyte Complexes with High or Low Developmental Potential. Theriogenology 2015, 83, 228–237. [Google Scholar] [CrossRef]
- Wyse, B.A.; Fuchs Weizman, N.; Kadish, S.; Balakier, H.; Sangaralingam, M.; Librach, C.L. Transcriptomics of Cumulus Cells—A Window into Oocyte Maturation in Humans. J. Ovarian Res. 2020, 13, 93. [Google Scholar] [CrossRef]
- Eppig, J. Oocyte Control of Ovarian Follicular Development and Function in Mammals. Reproduction 2001, 122, 829–838. [Google Scholar] [CrossRef]
- Su, Y.-Q.; Sugiura, K.; Woo, Y.; Wigglesworth, K.; Kamdar, S.; Affourtit, J.; Eppig, J.J. Selective Degradation of Transcripts during Meiotic Maturation of Mouse Oocytes. Dev. Biol. 2007, 302, 104–117. [Google Scholar] [CrossRef]
- Eichenlaub-Ritter, U.; Vogt, E.; Yin, H.; Gosden, R. Spindles, Mitochondria and Redox Potential in Ageing Oocytes. Reprod. Biomed. Online 2004, 8, 45–58. [Google Scholar] [CrossRef]
- Eppig, J.J. Reproduction: Oocytes Call, Granulosa Cells Connect. Curr. Biol. 2018, 28, R354–R356. [Google Scholar] [CrossRef] [Green Version]
- Bachvarova, R. Gene Expression During Oogenesis and Oocyte Development in Mammals. In Oogenesis; Springer: Boston, MA, USA, 1985; pp. 453–524. [Google Scholar]
- Braude, P.; Bolton, V.; Moore, S. Human Gene Expression First Occurs between the Four- and Eight-Cell Stages of Preimplantation Development. Nature 1988, 332, 459–461. [Google Scholar] [CrossRef]
- De La Fuente, R.; Viveiros, M.M.; Burns, K.H.; Adashi, E.Y.; Matzuk, M.M.; Eppig, J.J. Major Chromatin Remodeling in the Germinal Vesicle (GV) of Mammalian Oocytes Is Dispensable for Global Transcriptional Silencing but Required for Centromeric Heterochromatin Function. Dev. Biol. 2004, 275, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Macaulay, A.D.; Gilbert, I.; Caballero, J.; Barreto, R.; Fournier, E.; Tossou, P.; Sirard, M.-A.; Clarke, H.J.; Khandjian, É.W.; Richard, F.J.; et al. The Gametic Synapse: RNA Transfer to the Bovine Oocyte. Biol. Reprod. 2014, 91, 90. [Google Scholar] [CrossRef] [PubMed]
- Macaulay, A.D.; Gilbert, I.; Scantland, S.; Fournier, E.; Ashkar, F.; Bastien, A.; Saadi, H.A.S.; Gagné, D.; Sirard, M.-A.; Khandjian, É.W.; et al. Cumulus Cell Transcripts Transit to the Bovine Oocyte in Preparation for Maturation1. Biol. Reprod. 2016, 94, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Chronowska, E. High-Throughput Analysis of Ovarian Granulosa Cell Transcriptome. Biomed. Res. Int. 2014, 2014, 1–7. [Google Scholar] [CrossRef]
- Hallberg, I.; Persson, S.; Olovsson, M.; Sirard, M.-A.; Damdimopoulou, P.; Rüegg, J.; Sjunnesson, Y.C.B. Perfluorooctane Sulfonate (PFOS) Exposure of Bovine Oocytes Affects Early Embryonic Development at Human-Relevant Levels in an in Vitro Model. Toxicology 2021, 464, 153028. [Google Scholar] [CrossRef]
- Martino, N.A.; Lacalandra, G.M.; Filioli Uranio, M.; Ambruosi, B.; Caira, M.; Silvestre, F.; Pizzi, F.; Desantis, S.; Accogli, G.; Dell’Aquila, M.E. Oocyte Mitochondrial Bioenergy Potential and Oxidative Stress: Within-/between-Subject, inVivo versus in Vitro Maturation, and Age-Related Variations in a Sheep Model. Fertil. Steril. 2012, 97, 720–728. [Google Scholar] [CrossRef]
- Mastrorocco, A.; Martino, N.A.; Marzano, G.; Lacalandra, G.M.; Ciani, E.; Roelen, B.A.J.; Dell’Aquila, M.E.; Minervini, F. The Mycotoxin Beauvericin Induces Oocyte Mitochondrial Dysfunction and Affects Embryo Development in the Juvenile Sheep. Mol. Reprod. Dev. 2019, 86, 1430–1443. [Google Scholar] [CrossRef]
- Dell’Aquila, M.E.; Asif, S.; Temerario, L.; Mastrorocco, A.; Marzano, G.; Martino, N.A.; Lacalandra, G.M.; Roelen, B.A.; Carluccio, A.; Robbe, D.; et al. Ochratoxin A Affects Oocyte Maturation and Subsequent Embryo Developmental Dynamics in the Juvenile Sheep Model. Mycotoxin Res. 2021, 37, 23–37. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Franz, M.; Rodriguez, H.; Lopes, C.; Zuberi, K.; Montojo, J.; Bader, G.D.; Morris, Q. GeneMANIA Update 2018. Nucleic Acids Res. 2018, 46, W60–W64. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Yang, Y.; Zou, J.; Cheng, Y.; Yang, Y.; Wu, J.; Pollak, P.; Yang, Y. Transcriptome Analysis of the Ovary of Beet Armyworm Spodoptera Exigua under Different Exposures of Cadmium Stress. Chemosphere 2020, 251, 126372. [Google Scholar] [CrossRef]
- Liu, P.; Zhao, Y.; Wang, S.; Xing, H.; Dong, W.-F. Effect of Combined Exposure to Silica Nanoparticles and Cadmium Chloride on Female Zebrafish Ovaries. Environ. Toxicol. Pharmacol. 2021, 87, 103720. [Google Scholar] [CrossRef]
- Wang, J.; Peng, X.; Yang, H.; Lv, B.; Wang, Z.; Song, Q. Mul-Tiomics Analysis of Cadmium Stress on the Ovarian Function of the Wolf Spider Pardosa Pseudoannulata. Chemosphere 2020, 248, 125904. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.R.; Ariu, F.; Maltana, A.; Leoni, G.G.; Martino, N.A.; Mastrorocco, A.; Dell’Aquila, M.E.; Bogliolo, L. Protective Effect of Resveratrol against Cadmium-Induced Toxicity on Ovine Oocyte in Vitro Maturation and Fertilization. J. Anim. Sci. Biotechnol. 2022, 13, 83. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, J.; Wen, H.; Feng, Y.; Zhang, X.; Xiang, H.; Cao, Y.; Tong, X.; Ji, Y.; Xue, Z. Analyzing the Transcriptome Profile of Human Cumulus Cells Related to Embryo Quality via RNA Sequencing. Biomed. Res. Int. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Agca, C.; Yakan, A.; Agca, Y. Estrus Synchronization and Ovarian Hyper-Stimulation Treatments Have Negligible Effects on Cumulus Oocyte Complex Gene Expression Whereas Induction of Ovulation Causes Major Expression Changes. Mol. Reprod. Dev. 2013, 80, 102–117. [Google Scholar] [CrossRef]
- Yi, Z.; Meng, T.; Ma, X.; Li, J.; Zhang, C.; Ouyang, Y.; Schatten, H.; Qiao, J.; Sun, Q.; Qian, W. CDC6 Regulates Both G2/M Transition and Metaphase-to-anaphase Transition during the First Meiosis of Mouse Oocytes. J. Cell. Physiol. 2020, 235, 5541–5554. [Google Scholar] [CrossRef]
- Anger, M.; Stein, P.; Schultz, R.M. CDC6 Requirement for Spindle Formation During Maturation of Mouse Oocytes1. Biol. Reprod. 2005, 72, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Cheeseman, I.M.; Hori, T.; Okawa, K.; McLeod, I.X.; Yates, J.R.; Desai, A.; Fukagawa, T. The CENP-H–I Complex Is Required for the Efficient Incorporation of Newly Synthesized CENP-A into Centromeres. Nat. Cell. Biol. 2006, 8, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Li, J.; Wang, Y.; Wang, W.; Zhu, Q.; He, Y.; Lu, Y.; Wu, H.; Li, X.; Zhu, Z.; et al. Novel Role of CXCL14 in Modulating STAR Expression in Luteinized Granulosa Cells: Implication for Progesterone Synthesis in PCOS Patients. Transl. Res. 2021, 230, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Bobe, J.; Montfort, J.; Nguyen, T.; Fostier, A. Identification of New Participants in the Rainbow Trout (Oncorhynchus Mykiss) Oocyte Maturation and Ovulation Processes Using CDNA Microarrays. Reprod. Biol. Endocrinol. 2006, 4, 39. [Google Scholar] [CrossRef]
- Costermans, N.G.J.; Soede, N.M.; Van Tricht, F.; Blokland, M.; Kemp, B.; Keijer, J.; Teerds, K.J. Follicular Fluid Steroid Profile in Sows: Relationship to Follicle Size and Oocyte Quality†. Biol. Reprod. 2020, 102, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Huang, Y.; Lin, H. Estrogen Disorders: Interpreting the Abnormal Regulation of Aromatase in Granulosa Cells (Review). Int. J. Mol. Med. 2021, 47, 73. [Google Scholar] [CrossRef] [PubMed]
- Motahari Rad, H.; Mowla, S.J.; Ramazanali, F.; Rezazadeh Valojerdi, M. Characterization of Altered MicroRNAs Related to Different Phenotypes of Polycystic Ovarian Syndrome (PCOS) in Serum, Follicular Fluid, and Cumulus Cells. Taiwan J. Obstet. Gynecol. 2022, 61, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Heidarzadehpilehrood, R.; Pirhoushiaran, M.; Abdollahzadeh, R.; Binti Osman, M.; Sakinah, M.; Nordin, N.; Abdul Hamid, H. A Review on CYP11A1, CYP17A1, and CYP19A1 Polymorphism Studies: Candidate Susceptibility Genes for Polycystic Ovary Syndrome (PCOS) and Infertility. Genes 2022, 13, 302. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, R.; Prabhu, N.B.; Kabekkodu, S.P.; Rai, P.S. Review on Bisphenol A and the Risk of Polycystic Ovarian Syndrome: An Insight from Endocrine and Gene Expression. Environ. Sci. Pollut. Res. 2022, 29, 32631–32650. [Google Scholar] [CrossRef]
- Guo, J.; Shi, L.; Gong, X.; Jiang, M.; Yin, Y.; Zhang, X.; Yin, H.; Li, H.; Emori, C.; Sugiura, K.; et al. Oocyte-Dependent Activation of MTOR in Cumulus Cells Controls the Development and Survival of Cumulus-Oocyte Complexes. J. Cell. Sci. 2016, 129, 3091–3103. [Google Scholar] [CrossRef]
- Di Stefano, L. E2F7, a Novel E2F Featuring DP-Independent Repression of a Subset of E2F-Regulated Genes. EMBO J. 2003, 22, 6289–6298. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Yoshino, O.; Osuga, Y.; Akiyama, I.; Harada, M.; Koga, K.; Fujimoto, A.; Yano, T.; Taketani, Y. Growth Differentiation Factor 3 Is Induced by Bone Morphogenetic Protein 6 (BMP-6) and BMP-7 and Increases Luteinizing Hormone Receptor Messenger RNA Expression in Human Granulosa Cells. Fertil. Steril. 2012, 97, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Spitschak, M.; Hoeflich, A. Potential Functions of IGFBP-2 for Ovarian Folliculogenesis and Steroidogenesis. Front. Endocrinol. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Maffi, A.S.; Tonellotto Dos Santos, J.; Caetano De Oliveira, F.; Gasperin, B.G.; Schneider, A.; Rincón, J.A.A.; Rabassa, V.R.; Burkert Del Pino, F.A.; Corrêa, M.N.; Brauner, C.C. Insulin Treatment Does Not Affect Follicular Development but Alters Granulosa Cell Gene Expression in Dairy Cows. Theriogenology 2019, 133, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Mazerbourg, S.; Monget, P. Insulin-Like Growth Factor Binding Proteins and IGFBP Proteases: A Dynamic System Regulating the Ovarian Folliculogenesis. Front. Endocrinol. 2018, 9, 134. [Google Scholar] [CrossRef]
- Satrapa, R.A.; Castilho, A.S.; Razza, E.M.; Pegorer, M.F.; Puelker, R.; Barros, C.M. Differential Expression of Members of the IGF System in OPU-Derived Oocytes from Nelore (Bos Indicus) and Holstein (Bos Taurus) Cows. Anim. Reprod. Sci. 2013, 138, 155–158. [Google Scholar] [CrossRef]
- Nuttinck, F.; Charpigny, G.; Mermillod, P.; Loosfelt, H.; Meduri, G.; Freret, S.; Grimard, B.; Heyman, Y. Expression of Components of the Insulin-like Growth Factor System and Gonadotropin Receptors in Bovine Cumulus–Oocyte Complexes during Oocyte Maturation. Domest. Anim. Endocrinol. 2004, 27, 179–195. [Google Scholar] [CrossRef]
- Regassa, A.; Rings, F.; Hoelker, M.; Cinar, U.; Tholen, E.; Looft, C.; Schellander, K.; Tesfaye, D. Transcriptome Dynamics and Molecular Cross-Talk between Bovine Oocyte and Its Companion Cumulus Cells. BMC Genom. 2011, 12, 57. [Google Scholar] [CrossRef]
- Kulus, M.; Kranc, W.; Sujka-Kordowska, P.; Mozdziak, P.; Jankowski, M.; Konwerska, A.; Kulus, J.; Bukowska, D.; Skowroński, M.; Piotrowska-Kempisty, H.; et al. The Processes of Cellular Growth, Aging, and Programmed Cell Death Are Involved in Lifespan of Ovarian Granulosa Cells during Short-Term IVC—Study Based on Animal Model. Theriogenology 2020, 148, 76–88. [Google Scholar] [CrossRef]
- Brązert, M.; Kranc, W.; Nawrocki, M.; Sujka-Kordowska, P.; Konwerska, A.; Jankowski, M.; Kocherova, I.; Celichowski, P.; Jeseta, M.; Ożegowska, K.; et al. New Markers for Regulation of Transcription and Macromolecule Metabolic Process in Porcine Oocytes during in Vitro Maturation. Mol. Med. Rep. 2020, 21, 1537–1551. [Google Scholar] [CrossRef]
- Li, Y.; Xiong, G.; Tan, J.; Wang, S.; Wu, Q.; Wan, L.; Zhang, Z.; Huang, O. Aberrant Activation of the Hedgehog Signaling Pathway in Granulosa Cells from Patients with Polycystic Ovary Syndrome. Bioengineered 2021, 12, 12123–12134. [Google Scholar] [CrossRef]
- Liu, C.; Peng, J.; Matzuk, M.M.; Yao, H.H.-C. Lineage Specification of Ovarian Theca Cells Requires Multicellular Interactions via Oocyte and Granulosa Cells. Nat. Commun. 2015, 6, 6934. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Z.; Wang, Y.; Cai, Q.; Liu, H.; Xu, C.; Zhang, F. IL-15 Participates in the Pathogenesis of Polycystic Ovary Syndrome by Affecting the Activity of Granulosa Cells. Front. Endocrinol. 2022, 13, 787876. [Google Scholar] [CrossRef] [PubMed]
- Machlin, J.H.; Barishansky, S.J.; Kelsh, J.; Larmore, M.J.; Johnson, B.W.; Pritchard, M.T.; Pavone, M.E.; Duncan, F.E. Fibroinflammatory Signatures Increase with Age in the Human Ovary and Follicular Fluid. Int. J. Mol. Sci. 2021, 22, 4902. [Google Scholar] [CrossRef]
- Kaur, S.; Archer, K.J.; Devi, M.G.; Kriplani, A.; Strauss, J.F.; Singh, R. Differential Gene Expression in Granulosa Cells from Polycystic Ovary Syndrome Patients with and without Insulin Resistance: Identification of Susceptibility Gene Sets through Network Analysis. J. Clin. Endocrinol. Metab. 2012, 97, E2016–E2021. [Google Scholar] [CrossRef]
- Hatzirodos, N.; Hummitzsch, K.; Irving-Rodgers, H.F.; Rodgers, R.J. Transcriptome Comparisons Identify New Cell Markers for Theca Interna and Granulosa Cells from Small and Large Antral Ovarian Follicles. PLoS ONE 2015, 10, e0119800. [Google Scholar] [CrossRef] [PubMed]
- Nore, A.; Juarez-Martinez, A.B.; Clément, J.; Brun, C.; Diagouraga, B.; Laroussi, H.; Grey, C.; Bourbon, H.M.; Kadlec, J.; Robert, T.; et al. TOPOVIBL-REC114 Interaction Regulates Meiotic DNA Double-Strand Breaks. Nat. Commun. 2022, 13, 7048. [Google Scholar] [CrossRef]
- Jo, M.; Curry, T.E. Luteinizing Hormone-Induced RUNX1 Regulates the Expression of Genes in Granulosa Cells of Rat Periovulatory Follicles. J. Mol. Endocrinol. 2006, 20, 2156–2172. [Google Scholar] [CrossRef]
- Abe, T.; Lee, A.; Sitharam, R.; Kesner, J.; Rabadan, R.; Shapira, S.D. Germ-Cell-Specific Inflammasome Component NLRP14 Negatively Regulates Cytosolic Nucleic Acid Sensing to Promote Fertilization. Immunity 2017, 46, 621–634. [Google Scholar] [CrossRef]
- Dankert, D.; Demond, H.; Trapphoff, T.; Heiligentag, M.; Rademacher, K.; Eichenlaub-Ritter, U.; Horsthemke, B.; Grümmer, R. Pre- and Postovulatory Aging of Murine Oocytes Affect the Transcript Level and Poly(A) Tail Length of Maternal Effect Genes. PLoS ONE 2014, 9, e108907. [Google Scholar] [CrossRef]
- Molinari, E.; Bar, H.; Pyle, A.M.; Patrizio, P. Transcriptome Analysis of Human Cumulus Cells Reveals Hypoxia as the Main Determinant of Follicular Senescence. Mol. Hum. Reprod. 2016, 22, 866–876. [Google Scholar] [CrossRef] [Green Version]
- Bergandi, L.; Basso, G.; Evangelista, F.; Canosa, S.; Dalmasso, P.; Aldieri, E.; Revelli, A.; Benedetto, C.; Ghigo, D. Inducible Nitric Oxide Synthase and Heme Oxygenase 1 Are Expressed in Human Cumulus Cells and May Be Used as Biomarkers of Oocyte Competence. Reprod. Sci. 2014, 21, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Sammad, A.; Luo, H.; Hu, L.; Zhu, H.; Wang, Y. Transcriptome Reveals Granulosa Cells Coping through Redox, Inflammatory and Metabolic Mechanisms under Acute Heat Stress. Cells 2022, 11, 1443. [Google Scholar] [CrossRef] [PubMed]
- Huo, L.-J.; Liang, C.-G.; Yu, L.-Z.; Zhong, Z.-S.; Yang, Z.-M.; Fan, H.-Y.; Chen, D.-Y.; Sun, Q.-Y. Inducible Nitric Oxide Synthase-Derived Nitric Oxide Regulates Germinal Vesicle Breakdown and First Polar Body Emission in the Mouse Oocyte. Reproduction 2005, 129, 403–409. [Google Scholar] [CrossRef]
- Nath, P.; Maitra, S. Physiological Relevance of Nitric Oxide in Ovarian Functions: An Overview. Gen. Comp. Endocrinol. 2019, 279, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Nanda, N.; Bao, M.; Lin, H.; Clauser, K.; Komuves, L.; Quertermous, T.; Conley, P.B.; Phillips, D.R.; Hart, M.J. Platelet Endothelial Aggregation Receptor 1 (PEAR1), a Novel Epidermal Growth Factor Repeat-Containing Transmembrane Receptor, Participates in Platelet Contact-Induced Activation. J. Biol. Chem. 2005, 280, 24680–24689. [Google Scholar] [CrossRef]
- Chiba, H.; Michibata, H.; Wakimoto, K.; Seishima, M.; Kawasaki, S.; Okubo, K.; Mitsui, H.; Torii, H.; Imai, Y. Cloning of a Gene for a Novel Epithelium-Specific Cytosolic Phospholipase A2, CPLA2δ, Induced in Psoriatic Skin. J. Biol. Chem. 2004, 279, 12890–12897. [Google Scholar] [CrossRef]
- Yang, H.; Lin, S.; Lei, X.; Yuan, C.; Tian, Z.; Yu, Y.; Zhao, Z.; Chen, J. Identification and Profiling of MicroRNAs from Ovary of Estrous Kazakh Sheep Induced by Nutritional Status in the Anestrous Season. Anim. Reprod. Sci. 2016, 175, 18–26. [Google Scholar] [CrossRef]
- Zhou, H.; Xu, Q.-Z.; Zhang, R.; Zhuang, Z.-X.; Ma, Y.-Q.; Wang, W.; Ma, T.-Y.; Sui, Y.; Liu, Y.; Cao, X. Gonadal Transcriptome Analysis of Hybrid Triploid Loaches (Misgurnus anguillicaudatus) and Their Diploid and Tetraploid Parents. PLoS ONE 2018, 13, e0198179. [Google Scholar] [CrossRef]
- Shrestha, K.; Al-Alem, L.; Garcia, P.; Wynn, M.A.A.; Hannon, P.R.; Jo, M.; Drnevich, J.; Duffy, D.M.; Curry, T.E., Jr. Neurotensin Expression, Regulation, and Function during the Ovulatory Period in the Mouse Ovary. Biol. Reprod. 2022, 108, 107–120. [Google Scholar] [CrossRef]
- Cox, L.; Vanderwall, D.K.; Parkinson, K.C.; Sweat, A.; Isom, S.C. Expression Profiles of Select Genes in Cumulus–Oocyte Complexes from Young and Aged Mares. Reprod. Fertil. Dev. 2015, 27, 914. [Google Scholar] [CrossRef]
- Yang, X.; Dunning, K.R.; Wu, L.L.-Y.; Hickey, T.E.; Norman, R.J.; Russell, D.L.; Liang, X.; Robker, R.L. Identification of Perilipin-2 as a Lipid Droplet Protein Regulated in Oocytes during Maturation. Reprod. Fertil. Dev. 2010, 22, 1262. [Google Scholar] [CrossRef]
- Huang, J.Z.; Huang, L.M.; Zeng, Q.J.; Huang, E.F.; Liang, H.P.; Wei, Q.; Xie, X.H.; Ruan, J.M. Distribution and Quantitative Analysis of CIDEa and CIDEc in Broiler Chickens: Accounting for Differential Fat Deposition between Strains. Br. Poult. Sci. 2018, 59, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Congras, A.; Yerle-Bouissou, M.; Pinton, A.; Vignoles, F.; Liaubet, L.; Ferchaud, S.; Acloque, H. Sperm DNA Methylation Analysis in Swine Reveals Conserved and Species-Specific Methylation Patterns and Highlights an Altered Methylation at the GNAS Locus in Infertile Boars1. Biol. Reprod. 2014, 91, 137. [Google Scholar] [CrossRef]
- Lu, X.; Abdalla, I.M.; Nazar, M.; Fan, Y.; Zhang, Z.; Wu, X.; Xu, T.; Yang, Z. Genome-Wide Association Study on Reproduction-Related Body-Shape Traits of Chinese Holstein Cows. Animals 2021, 11, 1927. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.M.; Cánovas, A.; Mantilla-Rojas, C.; Riley, D.G.; Luna-Nevarez, P.; Coleman, S.J.; Speidel, S.E.; Enns, R.M.; Islas-Trejo, A.; Medrano, J.F.; et al. SNP Detection Using RNA-Sequences of Candidate Genes Associated with Puberty in Cattle. Genet. Mol. Res. 2017, 16, 1–17. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; He, B.; Wang, Y.; Yuan, Z.; Yuan, W.; Liao, P.; Deng, Y.; Xiao, J.; Zhu, C.; et al. Cloning and Expression of a Novel Human Gene, Isl-2, Encoded a LIM-Homeodomain Protein. Mol. Biol. Rep. 2007, 34, 19–26. [Google Scholar] [CrossRef]
- Mallya, M.; Campbell, R.D.; Aguado, B. Transcriptional Analysis of a Novel Cluster of LY-6 Family Members in the Human and Mouse Major Histocompatibility Complex: Five Genes with Many Splice Forms. Genomics 2002, 80, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Khole, V. Immune-Mediated Destruction of Ovarian Follicles Associated with the Presence of HSP90 Antibodies. Mol. Reprod. Dev. 2015, 82, 81–89. [Google Scholar] [CrossRef]
- Etchevers, L.; Stassi, A.F.; Belotti, E.M.; Diaz, P.U.; Durante, L.I.; Notaro, U.S.; Chiaraviglio, J.A.; Rey, F.; Salvetti, N.R.; Ortega, H.H.; et al. Exogenous ACTH Stimulus during the Preovulatory Period Alters Patterns of Leukocyte Recruitment in the Ovary of Dairy Cows. Theriogenology 2023, 195, 176–186. [Google Scholar] [CrossRef]
- Atli, M.O.; Mehta, V.; Vezina, C.M.; Wiltbank, M.C. Expression Patterns of Chemokine (C–C Motif) Ligand 2, Prostaglandin F2A Receptor and Immediate Early Genes at MRNA Level in the Bovine Corpus Luteum after Intrauterine Treatment with a Low Dose of Prostaglandin F2A. Theriogenology 2022, 189, 70–76. [Google Scholar] [CrossRef]
- Zhang, W.; Fu, Q.; Yao, K. A Three-mRNA Status Risk Score Has Greater Predictive Ability Compared with a LncRNA-based Risk Score for Predicting Prognosis in Patients with Hepatocellular Carcinoma. Oncol. Lett. 2020, 20, 48. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, X.; Xie, S.; Liu, X.; Fei, C.; Huang, X.; Tang, Y.; Zhou, L. H3K36me2 Methyltransferase NSD2 Orchestrates Epigenetic Reprogramming during Spermatogenesis. Nucleic Acids Res. 2022, 50, 6786–6800. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Xin, X.; Chang, H.-M.; Leung, P.C.K.; Yu, C.; Lian, F.; Wu, H. Expression of Long Noncoding RNAs in the Ovarian Granulosa Cells of Women with Diminished Ovarian Reserve Using High-Throughput Sequencing. J. Ovarian Res. 2022, 15, 119. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.-P.; He, W.-P.; Tan, J.-F.; Yang, Z.-X.; Fan, R.-R.; Ma, N.-F.; Wang, F.-W.; Chen, L.; Li, Y.; Shen, H.-W.; et al. Overexpression of SLC12A5 Is Associated with Tumor Progression and Poor Survival in Ovarian Carcinoma. Int. J. Gynecol. Canc. 2019, 29, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Fábryová, K.; Simon, M. Function of the Cell Surface Molecules (CD Molecules) in the Reproduction Processes. Gen. Physiol. Biophys. 2009, 28, 1–7. [Google Scholar] [CrossRef]
- Mahdipour, M.; Van Tol, H.T.A.; Stout, T.A.E.; Roelen, B.A.J. Validating Reference MicroRNAs for Normalizing QRT-PCR Data in Bovine Oocytes and Preimplantation Embryos. BMC Dev. Biol. 2015, 15, 25. [Google Scholar] [CrossRef]
- Ren, J.; Hao, Y.; Liu, Z.; Li, S.; Wang, C.; Wang, B.; Liu, Y.; Liu, G.; Dai, Y. Effect of Exogenous Glutathione Supplementation on the in Vitro Developmental Competence of Ovine Oocytes. Theriogenology 2021, 173, 144–155. [Google Scholar] [CrossRef]
- Latham, K.E.; Kutyna, K.; Wang, Q. Genetic Variation in Trophectoderm Function in Parthenogenetic Mouse Embryos. Dev. Genet. 1999, 24, 329–335. [Google Scholar] [CrossRef]
- Laitinen, M.; Vuojolainen, K.; Jaatinen, R.; Ketola, I.; Aaltonen, J.; Lehtonen, E.; Heikinheimo, M.; Ritvos, O. A Novel Growth Differentiation Factor-9 (GDF-9) Related Factor Is Co-Expressed with GDF-9 in Mouse Oocytes during Folliculogenesis. Mech. Dev. 1998, 78, 135–140. [Google Scholar] [CrossRef]
- Galloway, S.M.; McNatty, K.P.; Cambridge, L.M.; Laitinen, M.P.E.; Juengel, J.L.; Jokiranta, T.S.; McLaren, R.J.; Luiro, K.; Dodds, K.G.; Montgomery, G.W.; et al. Mutations in an Oocyte-Derived Growth Factor Gene (BMP15) Cause Increased Ovulation Rate and Infertility in a Dosage-Sensitive Manner. Nat. Genet. 2000, 25, 279–283. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Ritter, L.J.; Cranfield, M.; Jeffery, L.A.; Amato, F.; Scott, S.J.; Myllymaa, S.; Kaivo-Oja, N.; Lankinen, H.; Mottershead, D.G.; et al. Immunoneutralization of Growth Differentiation Factor 9 Reveals It Partially Accounts for Mouse Oocyte Mitogenic Activity1. Biol. Reprod. 2004, 71, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Ciccarelli, C.; Barberi, M.; Macchiarelli, G.; Canipari, R. Granulosa Cell-Oocyte Interactions. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 115, S19–S22. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Yan, J.; Duan, R.; Zhu, Y.; Hua, Y.; Liao, Y.; Li, Q.; Li, W.; Ji, S. E3 Ligase Cul2 Mediates Drosophila Early Germ Cell Differentiation through Targeting Bam. Dev. Biol. 2023, 493, 103–108. [Google Scholar] [CrossRef]
- Douville, G.; Sirard, M.-A. Changes in Granulosa Cells Gene Expression Associated with Growth, Plateau and Atretic Phases in Medium Bovine Follicles. J. Ovarian Res. 2014, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Hummitzsch, K.; Bastian, N.A.; Hartanti, M.D.; Wan, Q.; Irving-Rodgers, H.F.; Anderson, R.A.; Rodgers, R.J. Isolation, Culture, and Characterisation of Bovine Ovarian Fetal Fibroblasts and Gonadal Ridge Epithelial-like Cells and Comparison to Their Adult Counterparts. PLoS ONE 2022, 17, e0268467. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Xu, H.; Hou, Y.; Cai, Y.; Rong, Z.; Song, W.; Wang, W.; Li, K. Integrative Prognostic Subtype Discovery in High-grade Serous Ovarian Cancer. J. Cell. Biochem. 2019, 120, 18659–18666. [Google Scholar] [CrossRef]
- Maraldi, T.; Resca, E.; Nicoli, A.; Beretti, F.; Zavatti, M.; Capodanno, F.; Morini, D.; Palomba, S.; la Sala, G.B.; de Pol, A. NADPH Oxidase-4 and MATER Expressions in Granulosa Cells: Relationships with Ovarian Aging. Life Sci. 2016, 162, 108–114. [Google Scholar] [CrossRef]
- Huang, X.; Sun, Q.; Chen, D.; Yang, W.; Zhang, J.; Liu, R.; Zhang, P.; Huang, L.; Zhang, M.; Fu, Q. Nlrp5 and Tle6 Expression Patterns in Buffalo Oocytes and Pre-implantation Embryos. Reprod. Domest. Anim. 2022, 57, 481–488. [Google Scholar] [CrossRef]
- Bebbere, D.; Abazari-Kia, A.; Nieddu, S.; Melis Murgia, B.; Albertini, D.F.; Ledda, S. Subcortical Maternal Complex (SCMC) Expression during Folliculogenesis Is Affected by Oocyte Donor Age in Sheep. J. Assist. Reprod. Genet. 2020, 37, 2259–2271. [Google Scholar] [CrossRef]
- Tong, Z.-B.; Gold, L.; De Pol, A.; Vanevski, K.; Dorward, H.; Sena, P.; Palumbo, C.; Bondy, C.A.; Nelson, L.M. Developmental Expression and Subcellular Localization of Mouse MATER, an Oocyte-Specific Protein Essential for Early Development. Endocrinology 2004, 145, 1427–1434. [Google Scholar] [CrossRef] [Green Version]
- Tong, Z.-B.; Gold, L.; Pfeifer, K.E.; Dorward, H.; Lee, E.; Bondy, C.A.; Dean, J.; Nelson, L.M. Mater, a Maternal Effect Gene Required for Early Embryonic Development in Mice. Nat. Genet. 2000, 26, 267–268. [Google Scholar] [CrossRef]
- Tong, Z.-B.; Nelson, L.M.; Dean, J. Mater Encodes a Maternal Protein in Mice with a Leucine-Rich Repeat Domain Homologous to Porcine Ribonuclease Inhibitor. Mamm. Genome 2000, 11, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Sena, P.; Riccio, M.; Marzona, L.; Nicoli, A.; Marsella, T.; Marmiroli, S.; Bertacchini, J.; Fano, R.A.; La Sala, G.B.; de Pol, A. Human MATER Localization in Specific Cell Domains of Oocytes and Follicular Cells. Reprod. Biomed. Online 2009, 18, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Summers, A.F.; Pohlmeier, W.E.; Sargent, K.M.; Cole, B.D.; Vinton, R.J.; Kurz, S.G.; McFee, R.M.; Cushman, R.A.; Cupp, A.S.; Wood, J.R. Altered Theca and Cumulus Oocyte Complex Gene Expression, Follicular Arrest and Reduced Fertility in Cows with Dominant Follicle Follicular Fluid Androgen Excess. PLoS ONE 2014, 9, e110683. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.M.; Vogt, S.; O’Halloran, T.; Woodruff, T.K. Zinc Availability Regulates Exit from Meiosis in Maturing Mammalian Oocytes. Nat. Chem. Biol. 2010, 6, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, M.L.; Kim, A.M.; O’Halloran, T.; Woodruff, T.K. Zinc Requirement During Meiosis I–Meiosis II Transition in Mouse Oocytes Is Independent of the MOS-MAPK Pathway1. Biol. Reprod. 2011, 84, 526–536. [Google Scholar] [CrossRef]
- Suzuki, T.; Yoshida, N.; Suzuki, E.; Okuda, E.; Perry, A.C.F. Full-Term Mouse Development by Abolishing Zn2+-Dependent Metaphase II Arrest without Ca2+ Release. Development 2010, 137, 2659–2669. [Google Scholar] [CrossRef]
- Kim, A.M.; Bernhardt, M.L.; Kong, B.Y.; Ahn, R.W.; Vogt, S.; Woodruff, T.K.; O’Halloran, T.V. Zinc Sparks Are Triggered by Fertilization and Facilitate Cell Cycle Resumption in Mammalian Eggs. ACS Chem. Biol. 2011, 6, 716–723. [Google Scholar] [CrossRef]
- Lisle, R.S.; Anthony, K.; Randall, M.A.; Diaz, F.J. Oocyte–Cumulus Cell Interactions Regulate Free Intracellular Zinc in Mouse Oocytes. Reproduction 2013, 145, 381–390. [Google Scholar] [CrossRef]
- Oh, J.S.; Han, S.J.; Conti, M. Wee1B, Myt1, and Cdc25 Function in Distinct Compartments of the Mouse Oocyte to Control Meiotic Resumption. J. Cell. Biol. 2010, 188, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Solc, P.; Schultz, R.M.; Motlik, J. Prophase I Arrest and Progression to Metaphase I in Mouse Oocytes: Comparison of Resumption of Meiosis and Recovery from G2-Arrest in Somatic Cells. Mol. Hum. Reprod. 2010, 16, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, A.; Servin, B.; Mulsant, P.; Mandon-Pepin, B. Spatio-Temporal Gene Expression Profiling during In Vivo Early Ovarian Folliculogenesis: Integrated Transcriptomic Study and Molecular Signature of Early Follicular Growth. PLoS ONE 2015, 10, e0141482. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Chen, R.; Paronetto, M.P.; Conti, M. Wee1B Is an Oocyte-Specific Kinase Involved in the Control of Meiotic Arrest in the Mouse. Curr. Biol. 2005, 15, 1670–1676. [Google Scholar] [CrossRef] [PubMed]
- Hanna, C.B.; Yao, S.; Patta, M.C.; Jensen, J.T.; Wu, X. WEE2 Is an Oocyte-Specific Meiosis Inhibitor in Rhesus Macaque Monkeys1. Biol. Reprod. 2010, 82, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mu, J.; Zhao, J.; Zhou, Z.; Chen, B.; Wu, L.; Yan, Z.; Wang, W.; Zhao, L.; Dong, J.; et al. Novel Mutations in WEE2: Expanding the Spectrum of Mutations Responsible for Human Fertilization Failure. Clin. Genet. 2019, 95, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Mitwally, M.F.; Casper, R.F.; Diamond, M.P. The Role of Aromatase Inhibitors in Ameliorating Deleterious Effects of Ovarian Stimulation on Outcome of Infertility Treatment. Reprod. Biol. Endocrinol. 2005, 3, 54. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, T.; Leng, Y.; Chen, S.; Liu, Q.; Feng, J.; Chen, H.; Huang, Y.; Zhang, Q. Hormonal Changes and Folliculogenesis in Female Offspring of Rats Exposed to Cadmium during Gestation and Lactation. Environ. Pollut. 2018, 238, 336–347. [Google Scholar] [CrossRef]
- Das, S.; Mukherjee, D. Effect of Cadmium Chloride on Secretion of 17β-Estradiol by the Ovarian Follicles of Common Carp, Cyprinus Carpio. Gen. Comp. Endocrinol. 2013, 181, 107–114. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.; Penny, D.; Monget, P.; Arraztoa, J.A.; Fogelson, L.J.; Bondy, C.A. Insulin-like Growth Factor Binding Protein 4 Expression Parallels Luteinizing Hormone Receptor Expression and Follicular Luteinization in the Primate Ovary. Biol. Reprod. 2003, 69, 22–29. [Google Scholar] [CrossRef]
- Ramirez, D.C.; Martinez, L.D.; Marchevsky, E.; Gimenez, M.S. Biphasic Effect of Cadmium in Non-Cytotoxic Conditions on the Secretion of Nitric Oxide from Peritoneal Macrophages. Toxicology 1999, 139, 167–177. [Google Scholar] [CrossRef]
- Sangartit, W.; Kukongviriyapan, U.; Donpunha, W.; Pakdeechote, P.; Kukongviriyapan, V.; Surawattanawan, P.; Greenwald, S.E. Tetrahydrocurcumin Protects against Cadmium-Induced Hypertension, Raised Arterial Stiffness and Vascular Remodeling in Mice. PLoS ONE 2014, 9, e114908. [Google Scholar] [CrossRef] [PubMed]
- Refaie, M.M.M.; El-Hussieny, M.; Zenhom, N.M. Protective Role of Nebivolol in Cadmium-Induced Hepatotoxicity via Downregulation of Oxidative Stress, Apoptosis and Inflammatory Pathways. Environ. Toxicol. Pharmacol. 2018, 58, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.A.; Qureshi, H.A.; Al-Sultan, A.I.; Yacoubi, M.T.; Ali, A.A. Protective Effect of Hemin against Cadmium-Induced Testicular Damage in Rats. Toxicology 2009, 257, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Van Voorhis, B.J.; Dunn, M.S.; Snyder, G.D.; Weiner, C.P. Nitric Oxide: An Autocrine Regulator of Human Granulosa-Luteal Cell Steroidogenesis. Endocrinology 1994, 135, 1799–1806. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, C.; Zhang, J.; Lin, J.; Song, D.; Zhang, P.; Liu, Y.; Chen, L.; Zhang, L. Cadmium Impairs Zebrafish Swim Bladder Development via ROS Mediated Inhibition of the Wnt/Hedgehog Pathway. Aquat. Toxicol. 2022, 247, 106180. [Google Scholar] [CrossRef]
- Sabolić, I.; Breljak, D.; Skarica, M.; Herak-Kramberger, C.M. Role of Metallothionein in Cadmium Traffic and Toxicity in Kidneys and Other Mammalian Organs. Biometals 2010, 23, 897–926. [Google Scholar] [CrossRef]
- Liu, Y.P.; Liu, J.; Iszard, M.B.; Andrews, G.K.; Palmiter, R.D.; Klaassen, C.D. Transgenic Mice That Overexpress Metallothionein-I Are Protected from Cadmium Lethality and Hepatotoxicity. Toxicol. Appl. Pharmacol. 1995, 135, 222–228. [Google Scholar] [CrossRef]
- Shen, X.; Liu, W.; Chen, Y.; Guo, Y.; Gao, M.; Chen, W.; Liu, Y.; Liu, S. Diagnostic Significance of Metallothionein Members in Recognizing Cadmium Exposure in Various Organs under Low-Dose Exposure. Chemosphere 2019, 229, 32–40. [Google Scholar] [CrossRef]
- Kluxen, F.M.; Höfer, N.; Kretzschmar, G.; Degen, G.H.; Diel, P. Cadmium Modulates Expression of Aryl Hydrocarbon Receptor-Associated Genes in Rat Uterus by Interaction with the Estrogen Receptor. Arch. Toxicol. 2012, 86, 591–601. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Molecular and Ionic Mimicry and the Transport of Toxic Metals. Toxicol. Appl. Pharmacol. 2005, 204, 274–308. [Google Scholar] [CrossRef] [Green Version]
- Forti, E.; Bulgheroni, A.; Cetin, Y.; Hartung, T.; Jennings, P.; Pfaller, W.; Prieto, P. Characterisation of Cadmium Chloride Induced Molecular and Functional Alterations in Airway Epithelial Cells. Cell Physiol. Biochem. 2010, 25, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Han, S.G.; Castranova, V.; Vallyathan, V. Comparative Cytotoxicity of Cadmium and Mercury in a Human Bronchial Epithelial Cell Line (BEAS-2B) and Its Role in Oxidative Stress and Induction of Heat Shock Protein 70∗. J. Toxicol. Environ. Health A 2007, 70, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, U.; Priem, M.; Bartzsch, C.; Winckler, T.; Feller, K.-H. A Sensitive Sensor Cell Line for the Detection of Oxidative Stress Responses in Cultured Human Keratinocytes. Sensors 2014, 14, 11293–11307. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.-I.; Taniguchi, A.; Okano, T. Highly Sensitive Detection of Cytotoxicity Using a Modified HSP70B′ Promoter. Biotechnol. Bioeng. 2007, 97, 871–876. [Google Scholar] [CrossRef]
- Singh, P.; Chandrasekaran, V.; Hardy, B.; Wilmes, A.; Jennings, P.; Exner, T.E. Temporal Transcriptomic Alterations of Cadmium Exposed Human IPSC-Derived Renal Proximal Tubule-like Cells. Toxicol. Vitr. 2021, 76, 105229. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, G.; Jiang, X.; Wu, G. Resveratrol Ameliorates Toxic Effects of Cadmium on Placental Development in Mouse Placenta and Human Trophoblast Cells. Birth Defects Res. 2021, 113, 1470–1483. [Google Scholar] [CrossRef]
- Park, M.J.; Ahn, J.-W.; Kim, K.H.; Bang, J.; Kim, S.C.; Jeong, J.Y.; Choi, Y.E.; Kim, C.-W.; Joo, B.S. Prediction of Ovarian Aging Using Ovarian Expression of BMP15, GDF9, and C-KIT. Exp. Biol. Med. 2020, 245, 711–719. [Google Scholar] [CrossRef]
- Daneshjou, D.; Soleimani Mehranjani, M.; Zadeh Modarres, S.; Shariatzadeh, M.A. Sitagliptin/Metformin: A New Medical Treatment in Polycystic Ovary Syndrome. Trends Endocrinol. Metab. 2020, 31, 890–892. [Google Scholar] [CrossRef]
- Lu, M.; Fu, D. Structure of the Zinc Transporter YiiP. Science 2007, 317, 1746–1748. [Google Scholar] [CrossRef]
- Chemek, M.; Boughammoura, S.; Ben Mimouna, S.; Chouchene, L.; Banni, M.; Messaoudi, I. Changes of the MRNA Expression Pattern of Zn Transporters: A Probable Mechanism for Cadmium Retention and Zinc Redistribution in the Suckling Rat Tissues. Biol. Trace Elem. Res. 2015, 165, 173–182. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Full Name | Accession Number | Primer (5′→3′) | Ta (°C) | Size bps |
|---|---|---|---|---|---|
| ACTB | Actin beta | NM_001009784 | CCCTGGAGAAGAGCTACGAG TAGTTTCGTGAATGCCGCAG | 59 | 129 |
| MT-1A | Metallothionein-1A | XM_004014996 | CTTGCCACTTGTTCTGGACC AGCTCTTCTTGCAGGAGGG | 59 | 139 |
| DSG2 | Desmoglein 2 | XM_027960751.2 | CCGCCTTTTGGTGTGTTTGT AAGCGTAGCCAGTTAGCAGA | 60 | 106 |
| SLC30A2 | Solute carrier family (zinc transporter), member 2 | XM_012151636.2 | TCATCTGTGGGCTGAGAACG TCCACAACCACCATGTGCTC | 59 | 90 |
| BMP15 | BMP15 bone morphogenetic protein 15 | NM_001114767.2 | TGGTCCTCCTGAGCATCCTT CTCTGAGAGGCCTTGCTACA | 60 | 312 |
| CYP19A1 | Aromatase | NM_001123000.1 | CTCTCCTTCTCAAACCAGACATCTT ATGGCATCTTTCAAGTCCTTGACA | 59 | 88 |
| NOS2 | Nitric Oxide Synthase 2 | XM_012185382.3 | AGAGACGGGGAGATCGGAAA TGGGGATCTCAATGTGGTGC | 59 | 452 |
| IGFBP2 | Insulin-like Growth Factor Binding Protein 2 | NM_001009436.1 | GTGGCAAACATCACCTTGGC CCAGTGTTGGGGTTCACACA | 60 | 259 |
| IHH | Indian hedgehog signaling molecule | XM_027965204.1 | CACGGCCAACAATCACACTG CCCATGCCAAGCTGTGAAAC | 60 | 284 |
| Age Group | Cd Concentration (nM) | N° Evaluated Oocytes | Nuclear Chromatin Configurations (n°, %) | |||
|---|---|---|---|---|---|---|
| GV | MI a TI | MII + PB | Abnormal | |||
| Adult | 0 | 197 | 2 (1.0) | 6 (3.0) | 187 (95.0) | 2 (1.0) |
| 100 | 204 | 6 (3.0) | 8 (3.9) | 181 (88.7) | 9 (4.4) | |
| Prepubertal | 0 | 180 | 4 (2.2) | 10 (5.6) | 161 (89.4) | 5 (2.8) |
| 100 | 161 | 6 (3.7) | 5 (3.1) | 129 (87.6) | 9 (5.6) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martino, N.A.; Picardi, E.; Ciani, E.; D’Erchia, A.M.; Bogliolo, L.; Ariu, F.; Mastrorocco, A.; Temerario, L.; Mansi, L.; Palumbo, V.; et al. Cumulus Cell Transcriptome after Cumulus-Oocyte Complex Exposure to Nanomolar Cadmium in an In Vitro Animal Model of Prepubertal and Adult Age. Biology 2023, 12, 249. https://doi.org/10.3390/biology12020249
Martino NA, Picardi E, Ciani E, D’Erchia AM, Bogliolo L, Ariu F, Mastrorocco A, Temerario L, Mansi L, Palumbo V, et al. Cumulus Cell Transcriptome after Cumulus-Oocyte Complex Exposure to Nanomolar Cadmium in an In Vitro Animal Model of Prepubertal and Adult Age. Biology. 2023; 12(2):249. https://doi.org/10.3390/biology12020249
Chicago/Turabian StyleMartino, Nicola Antonio, Ernesto Picardi, Elena Ciani, Anna Maria D’Erchia, Luisa Bogliolo, Federica Ariu, Antonella Mastrorocco, Letizia Temerario, Luigi Mansi, Valeria Palumbo, and et al. 2023. "Cumulus Cell Transcriptome after Cumulus-Oocyte Complex Exposure to Nanomolar Cadmium in an In Vitro Animal Model of Prepubertal and Adult Age" Biology 12, no. 2: 249. https://doi.org/10.3390/biology12020249