
Non-Selective PDE4 Inhibition Induces a Rapid and Transient Decrease of Serum Potassium in Mice
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs
2.2. Animals
2.3. Measurement of Serum Electrolytes
2.4. Measurement of Core Body Temperature
2.5. Data and Statistical Analysis
3. Results
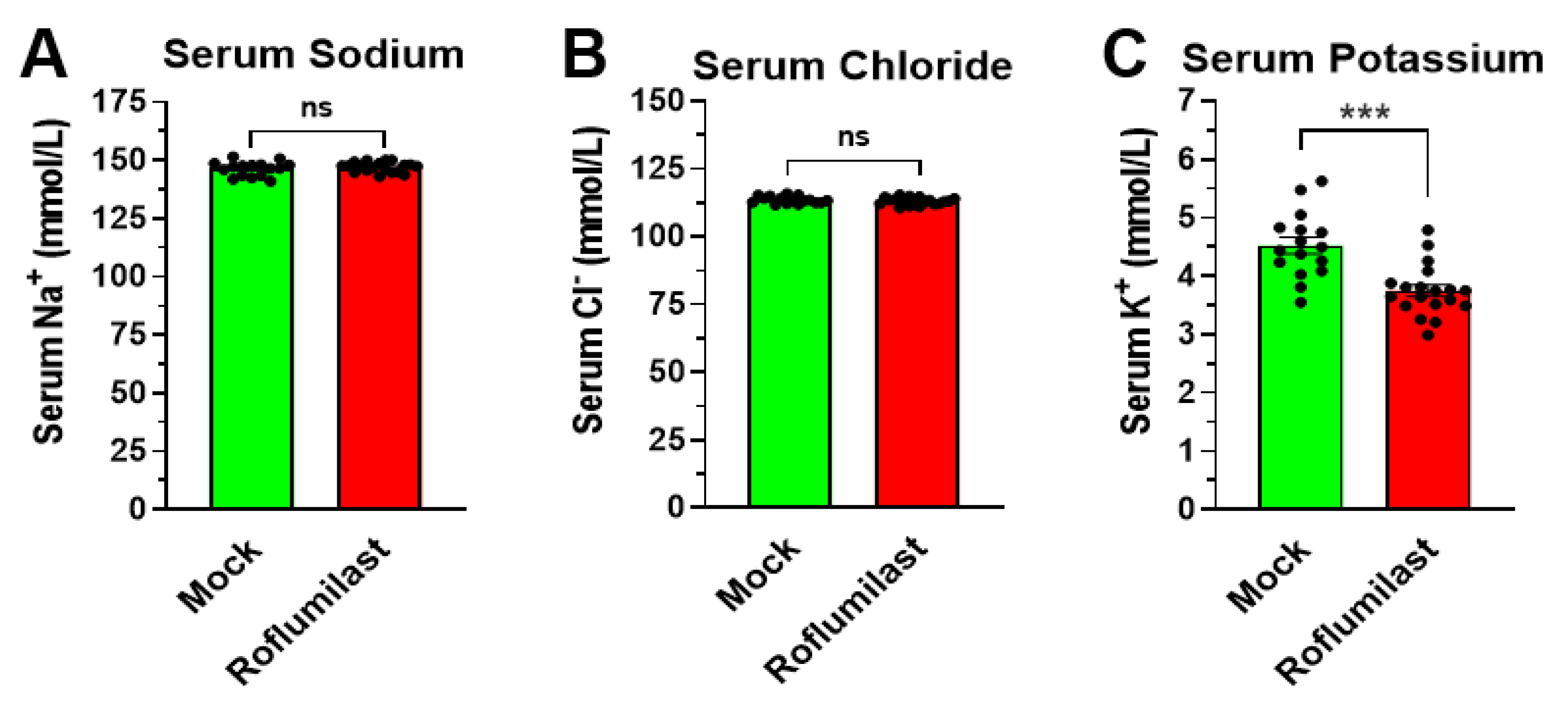
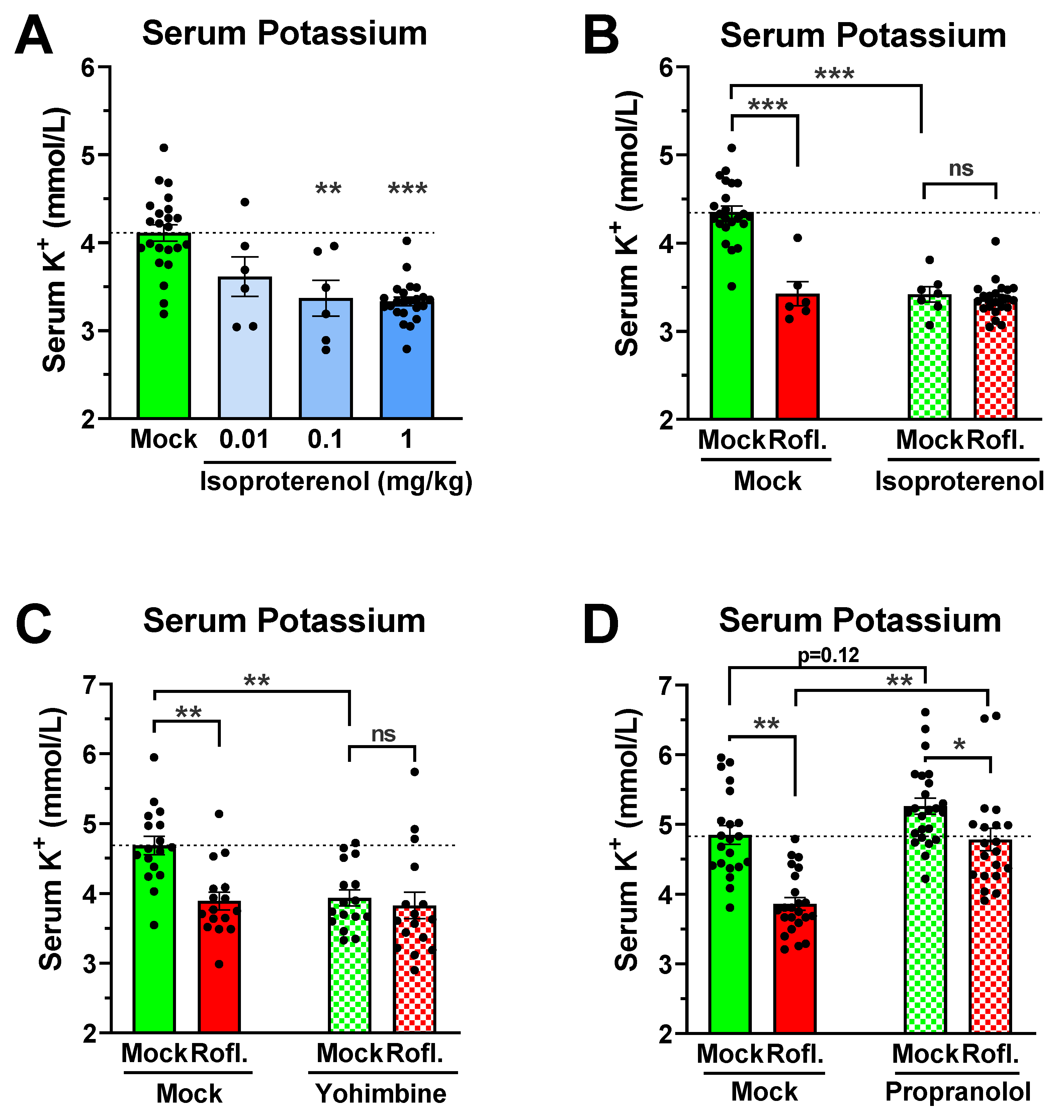
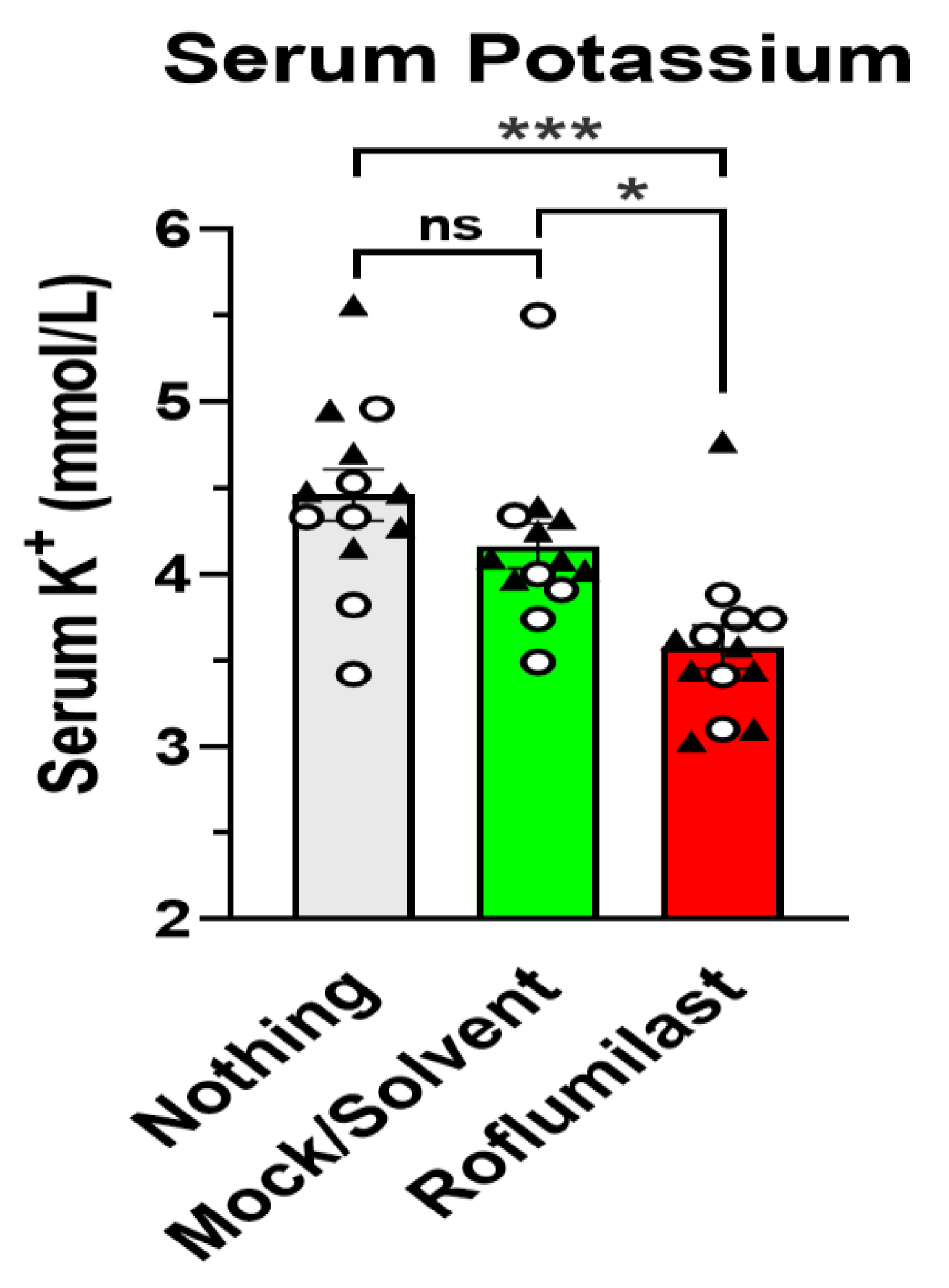
3.1. Treatment with the PAN-PDE4 Inhibitor Roflumilast Reduces Serum Potassium, but Not Serum Sodium or Chloride Levels
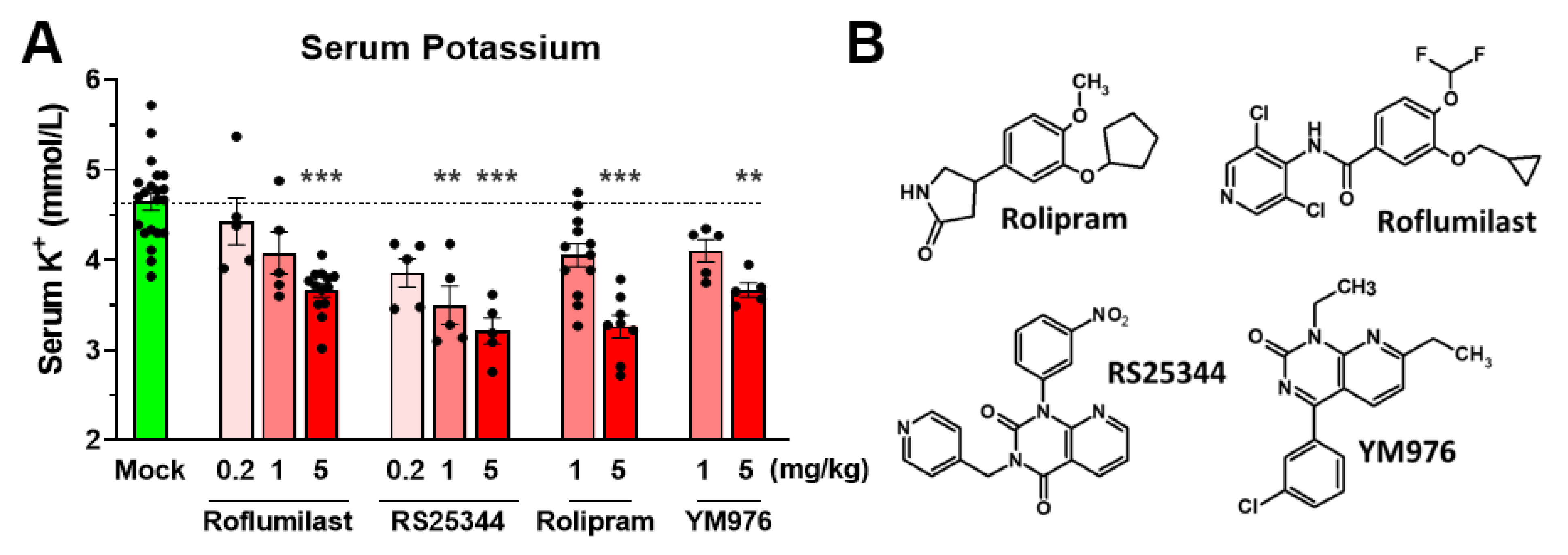
3.2. A Reduction in Serum Potassium Levels Is a Class Effect of PAN-Selective PDE4 Inhibitors
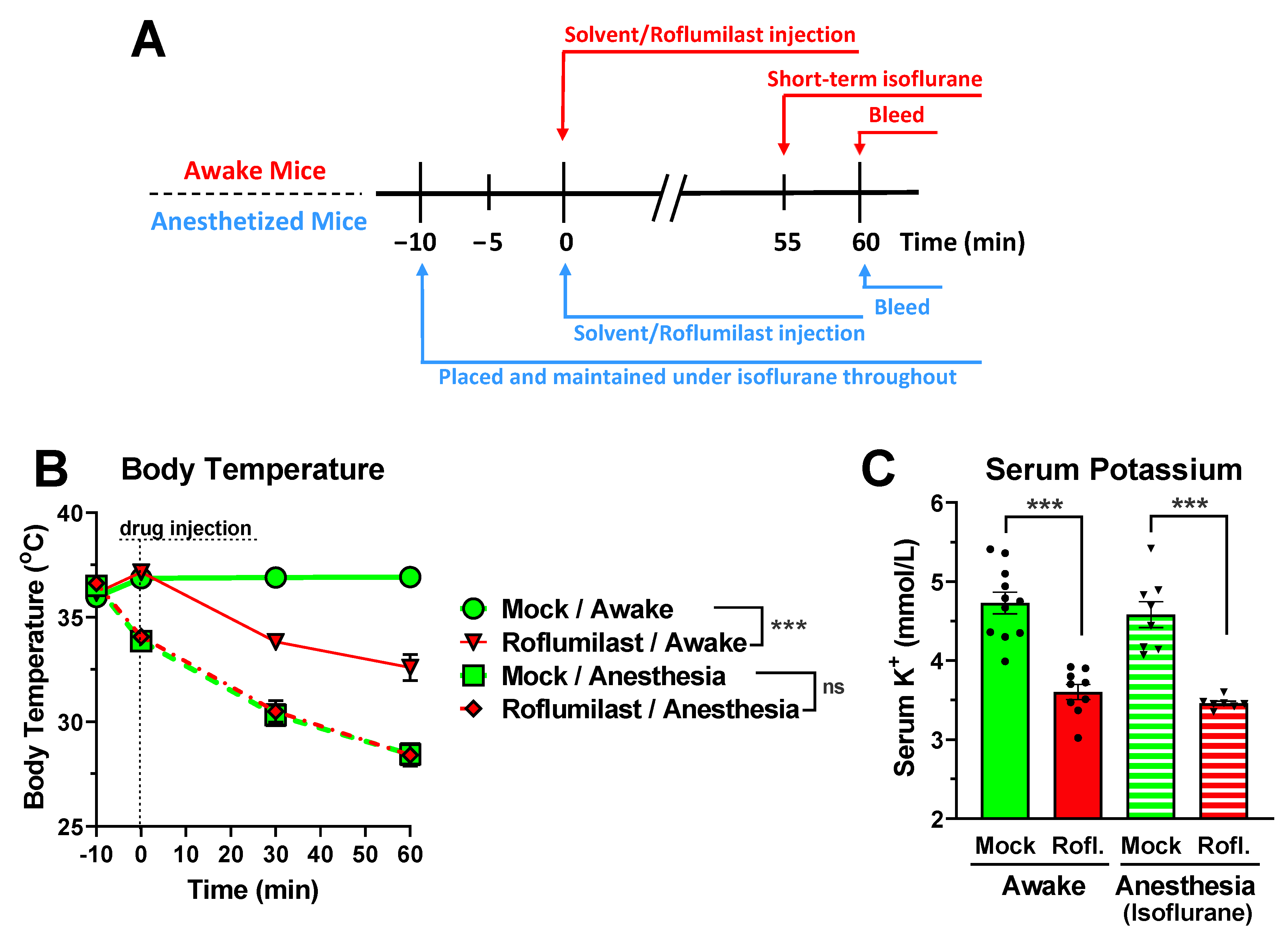
3.3. Reduced Serum Potassium Levels Are Not Dependent on PDE4 Inhibitor-Induced Hypothermia or Hypokinesia
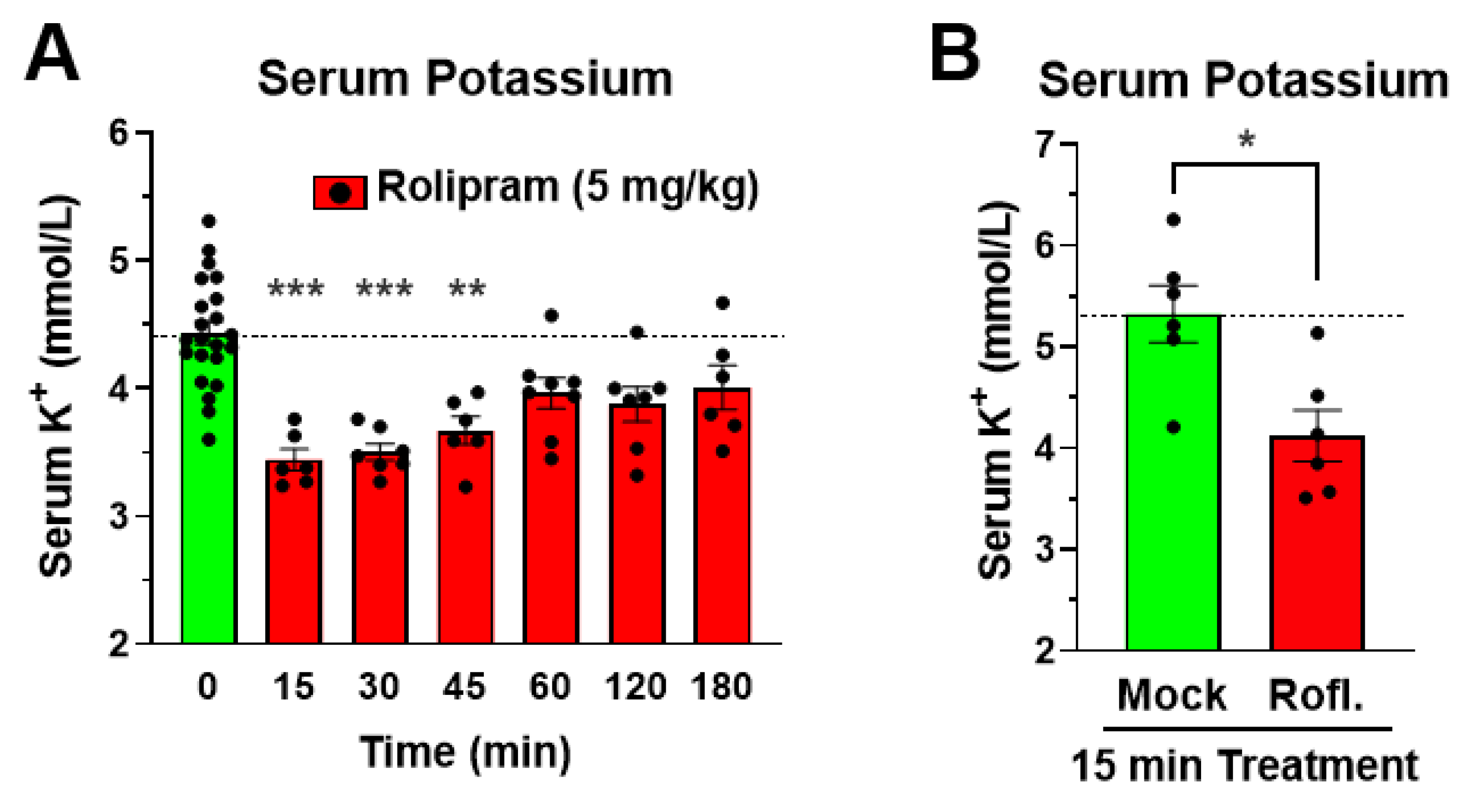
3.4. The Reduction in Serum Potassium Following PDE4 Inhibitor Treatment Is Rapid in Onset
3.5. Role of Adrenergic Signaling in Mediating PDE4 Inhibitor-Induced Hypokalemia
4. Discussion
4.1. A Reduction in Serum Potassium Levels Is a Class Effect of PAN-PDE4 Inhibitors in Mice
4.2. A cAMP-Mediated Transcellular Shift of Serum Potassium as a Potential Mechanism of PDE4 Inhibitor-Induced Hypokalemia
4.3. Potential Physiologic Relevance of PDE4 Inhibitor-Induced Hypokalemia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
Appendix A

References
- Clausen, T. Hormonal pharmacological modification of plasma potassium homeostasis. Fundam. Clin. Pharmacol. 2010, 24, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.F.; Clegg, D.J. Physiology and pathophysiology of potassium homeostasis. Adv. Physiol. Educ. 2016, 40, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Palmer, B.F. Regulation of Potassium Homeostasis. Clin. J. Am. Soc. Nephrol. 2015, 10, 1050–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLafferty, E.; Johnstone, C.; Hendry, C.; Farley, A. Fluid and electrolyte balance. Nurs. Stand. 2014, 28, 42–49. (In English) [Google Scholar] [CrossRef] [PubMed]
- Nielsen, O.B.; Clausen, T. The Na+/K(+)-pump protects muscle excitability and contractility during exercise. Exerc. Sport Sci. Rev. 2000, 28, 159–164. [Google Scholar]
- Hertz, L.; Gerkau, N.J.; Xu, J.; Durry, S.; Song, D.; Rose, C.R.; Peng, L. Roles of astrocytic Na(+),K(+)-ATPase and glycogenolysis for K(+) homeostasis in mammalian brain. J. Neurosci. Res. 2015, 93, 1019–1030. (In English) [Google Scholar] [CrossRef]
- Clifford, P.S. Skeletal muscle vasodilatation at the onset of exercise. J. Physiol. 2007, 583, 825–833. [Google Scholar] [CrossRef]
- McKenna, M.J.; Bangsbo, J.; Renaud, J.M. Muscle K+, Na+, and Cl disturbances and Na+-K+ pump inactivation: Implications for fatigue. J. Appl. Physiol. 2008, 104, 288–295. (In English) [Google Scholar] [CrossRef] [PubMed]
- Ho, K. A critically swift response: Insulin-stimulated potassium and glucose transport in skeletal muscle. Clin. J. Am. Soc. Nephrol. 2011, 6, 1513–1516. (In English) [Google Scholar] [CrossRef] [Green Version]
- Pirkmajer, S.; Chibalin, A.V. Na,K-ATPase regulation in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E1–E31. (In English) [Google Scholar] [CrossRef] [Green Version]
- Allon, M.; Copkney, C. Albuterol and insulin for treatment of hyperkalemia in hemodialysis patients. Kidney Int. 1990, 38, 869–872. [Google Scholar] [CrossRef]
- Deenstra, M.; Haalboom, J.R.; Struyvenberg, A. Decrease of plasma potassium due to inhalation of beta-2-agonists: Absence of an additional effect of intravenous theophylline. Eur. J. Clin. Investig. 1988, 18, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Montoliu, J.; Lens, X.M.; Revert, L. Potassium-lowering effect of albuterol for hyperkalemia in renal failure. Arch. Intern. Med. 1987, 147, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Han, S.W. Therapeutic approach to hyperkalemia. Nephron 2002, 92 (Suppl. S1), 33–40. (In English) [Google Scholar] [CrossRef]
- Baillie, G.S.; Tejeda, G.S.; Kelly, M.P. Therapeutic targeting of 3′,5′-cyclic nucleotide phosphodiesterases: Inhibition and beyond. Nat. Rev. Drug Dis. 2019, 18, 770–796. (In English) [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Beavo, J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: Essential components in cyclic nucleotide signaling. Annu. Rev. Biochem. 2007, 76, 481–511. (In English) [Google Scholar] [CrossRef]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharmacol. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef]
- Conti, M.; Richter, W.; Mehats, C.; Livera, G.; Park, J.Y.; Jin, C. Cyclic AMP-specific PDE4 phosphodiesterases as critical components of cyclic AMP signaling. J. Biol. Chem. 2003, 278, 5493–5496. (In English) [Google Scholar] [CrossRef] [Green Version]
- Paes, D.; Schepers, M.; Rombaut, B.; van den Hove, D.; Vanmierlo, T.; Prickaerts, J. The Molecular Biology of Phosphodiesterase 4 Enzymes as Pharmacological Targets: An Interplay of Isoforms, Conformational States, and Inhibitors. Pharmacol. Rev. 2021, 73, 1016–1049. [Google Scholar] [CrossRef]
- Bolger, G.B.; Conti, M.; Houslay, M.D. Cellular Functions of PDE4 Enzymes. In Cyclic Nucleotide Phosphodiesterases in Health and Disease; Beavo, J.A., Francis, S.H., Houslay, M.D., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 99–130. [Google Scholar]
- Houslay, M.D.; Schafer, P.; Zhang, K.Y. Keynote review: Phosphodiesterase-4 as a therapeutic target. Drug Discov. Today 2005, 10, 1503–1519. (In English) [Google Scholar] [CrossRef]
- Abou Saleh, L.; Boyd, A.; Aragon, I.V.; Koloteva, A.; Spadafora, D.; Mneimneh, W.; Barrington, R.A.; Richter, W. Ablation of PDE4B protects from Pseudomonas aeruginosa-induced acute lung injury in mice by ameliorating the cytostorm and associated hypothermia. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2021, 35, e21797. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.; Conti, M. Induction of the cyclic nucleotide phosphodiesterase PDE4B is essential for LPS-activated TNF-alpha responses. Proc. Natl. Acad. Sci. USA 2002, 99, 7628–7633. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.L.; Ding, S.L.; Lin, S.C. Phosphodiesterase 4 and its inhibitors in inflammatory diseases. Chang. Gung. Med. J. 2012, 35, 197–210. (In English) [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, W.E.; Wahlang, B.; Wang, Y.; Zhang, J.; Vadhanam, M.V.; Joshi-Barve, S.; Bauer, P.; Cannon, R.; Ahmadi, A.R.; Sun, Z.; et al. Phosphodiesterase 4 Inhibition as a Therapeutic Target for Alcoholic Liver Disease: From Bedside to Bench. Hepatology 2019, 70, 1958–1971. [Google Scholar] [CrossRef]
- Richter, W.; Menniti, F.S.; Zhang, H.T.; Conti, M. PDE4 as a target for cognition enhancement. Expert Opin. Targets 2013, 17, 1011–1027. (In English) [Google Scholar] [CrossRef] [Green Version]
- Reneerkens, O.A.; Rutten, K.; Steinbusch, H.W.; Blokland, A.; Prickaerts, J. Selective phosphodiesterase inhibitors: A promising target for cognition enhancement. Psychopharmacology 2009, 202, 419–443. [Google Scholar] [CrossRef] [Green Version]
- Kamel, R.; Leroy, J.; Vandecasteele, G.; Fischmeister, R. Cyclic nucleotide phosphodiesterases as therapeutic targets in cardiac hypertrophy and heart failure. Nat. Rev. Cardiol. 2022. (In English) [Google Scholar] [CrossRef]
- Houslay, M.D.; Baillie, G.S.; Maurice, D.H. cAMP-Specific phosphodiesterase-4 enzymes in the cardiovascular system: A molecular toolbox for generating compartmentalized cAMP signaling. Circ. Res. 2007, 100, 950–966. (In English) [Google Scholar] [CrossRef]
- Wu, C.; Rajagopalan, S. Phosphodiesterase-4 inhibition as a therapeutic strategy for metabolic disorders. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2016, 17, 429–441. [Google Scholar] [CrossRef]
- Boyd, A.; Aragon, I.V.; Abou Saleh, L.; Southers, D.; Richter, W. The cAMP-phosphodiesterase 4 (PDE4) controls β-adrenoceptor- and CFTR-dependent saliva secretion in mice. Biochem. J. 2021, 478, 1891–1906. [Google Scholar] [CrossRef]
- Boyd, A.; Aragon, I.V.; Rich, J.; McDonough, W.; Oditt, M.; Irelan, D.; Fiedler, E.; Abou Saleh, L.; Richter, W. Assessment of PDE4 Inhibitor-Induced Hypothermia as a Correlate of Nausea in Mice. Biology 2021, 10, 1355. [Google Scholar] [CrossRef] [PubMed]
- McDonough, W.; Aragon, I.V.; Rich, J.; Murphy, J.M.; Abou Saleh, L.; Boyd, A.; Koloteva, A.; Richter, W. PAN-selective inhibition of cAMP-phosphodiesterase 4 (PDE4) induces gastroparesis in mice. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 12533–12548. [Google Scholar] [CrossRef] [PubMed]
- McDonough, W.; Rich, J.; Aragon, I.V.; Abou Saleh, L.; Boyd, A.; Richter, A.; Koloteva, A.; Richter, W. Inhibition of type 4 cAMP-phosphodiesterases (PDE4s) in mice induces hypothermia via effects on behavioral and central autonomous thermoregulation. Biochem. Pharmacol. 2020, 180, 114158. [Google Scholar] [CrossRef]
- Thompson, W.J. Cyclic nucleotide phosphodiesterases: Pharmacology, biochemistry and function. Pharmacol. Ther. 1991, 51, 13–33. [Google Scholar] [CrossRef]
- Giembycz, M.A.; Field, S.K. Roflumilast: First phosphodiesterase 4 inhibitor approved for treatment of COPD. Drug Des. Devel. 2010, 4, 147–158. (In English) [Google Scholar]
- Hatzelmann, A.; Morcillo, E.J.; Lungarella, G.; Adnot, S.; Sanjar, S.; Beume, R.; Schudt, C.; Tenor, H. The preclinical pharmacology of roflumilast—A selective, oral phosphodiesterase 4 inhibitor in development for chronic obstructive pulmonary disease. Pulm. Pharmacol. Ther. 2010, 23, 235–256. [Google Scholar] [CrossRef]
- Aoki, M.; Fukunaga, M.; Sugimoto, T.; Hirano, Y.; Kobayashi, M.; Honda, K.; Yamada, T. Studies on mechanisms of low emetogenicity of YM976, a novel phosphodiesterase type 4 inhibitor. J. Pharm. Exp. 2001, 298, 1142–1149. (In English) [Google Scholar]
- Moriuchi, H.; Nakahara, T.; Maruko, T.; Sakamoto, K.; Ishii, K. Relaxant effect of YM976, a novel phosphodiesterase 4 inhibitor, on bovine tracheal smooth muscle. Eur. J. Pharm. 2003, 470, 57–64. (In English) [Google Scholar] [CrossRef]
- Aragon, I.V.; Boyd, A.; Abou Saleh, L.; Rich, J.; McDonough, W.; Koloteva, A.; Richter, W. Inhibition of cAMP-phosphodiesterase 4 (PDE4) potentiates the anesthetic effects of Isoflurane in mice. Biochem. Pharmacol. 2021, 186, 114477. [Google Scholar] [CrossRef]
- Buse, S.; Blancher, M.; Viglino, D.; Pasquier, M.; Maignan, M.; Bouzat, P.; Annecke, T.; Debaty, G. The impact of hypothermia on serum potassium concentration: A systematic review. Resuscitation 2017, 118, 35–42. [Google Scholar] [CrossRef]
- Koht, A.; Cane, R.; Cerullo, L.J. Serum potassium levels during prolonged hypothermia. Intensive Care Med. 1983, 9, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Sprung, J.; Cheng, E.Y.; Gamulin, S.; Kampine, J.P.; Bosnjak, Z.J. Effects of acute hypothermia and beta-adrenergic receptor blockade on serum potassium concentration in rats. Crit. Care Med. 1991, 19, 1545–1551. (In English) [Google Scholar] [CrossRef]
- Sprung, J.; Gamulin, S.; Bosnjak, Z.J.; Kampine, J.P. Potassium correction of hypothermic hypokalemia induces hyperkalemia after rewarming. Can. J. Anaesth. 1990, 37, S69. (In English) [Google Scholar] [PubMed]
- Zorbas, Y.G.; Kakuris, K.K.; Charapakhin, K.P.; Afoninos, A.B. Potassium Deposition during and after Hypokinesia in Potassium Supplemented and Unsupplemented Rats. Int. J. Med. Sci. 2005, 2, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federenko, Y.F.; Deogenov, V.A.; Kakuris, K.K.; Yerullis, K.B. Muscle Potassium and Potassium Losses during Hypokinesia in Healthy Subjects. Biol. Trace Elem. Res. 2011, 143, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Johansen, F.F.; Jørgensen, H.S.; Reith, J. Prolonged drug-induced hypothermia in experimental stroke. J. Stroke Cereb. Dis. 2003, 12, 97–102. (In English) [Google Scholar] [CrossRef]
- Huss, M.K.; Chum, H.H.; Chang, A.G.; Jampachairsi, K.; Pacharinsak, C. The Physiologic Effects of Isoflurane, Sevoflurane, and Hypothermia Used for Anesthesia in Neonatal Rats (Rattus norvegicus). J. Am. Assoc. Lab. Anim. Sci. JAALAS 2016, 55, 83–88. [Google Scholar]
- Kuroki, C.; Takahashi, Y.; Ootsuka, Y.; Kanmura, Y.; Kuwaki, T. The impact of hypothermia on emergence from isoflurane anesthesia in orexin neuron-ablated mice. Anesth. Analg. 2013, 116, 1001–1005. [Google Scholar] [CrossRef]
- Liu, M.; Hu, X.; Liu, J. The effect of hypothermia on isoflurane MAC in children. Anesthesiology 2001, 94, 429–432. [Google Scholar] [CrossRef]
- Shimaoka, H.; Shiina, T.; Suzuki, H.; Horii, Y.; Horii, K.; Shimizu, Y. Successful induction of deep hypothermia by isoflurane anesthesia and cooling in a non-hibernator, the rat. J. Physiol. Sci. JPS 2021, 71, 10. [Google Scholar] [CrossRef]
- Atanasovska, T.; Smith, R.; Graff, C.; Tran, C.T.; Melgaard, J.; Kanters, J.K.; Petersen, A.C.; Tobin, A.; Kjeldsen, K.P.; McKenna, M.J. Protection against severe hypokalemia but impaired cardiac repolarization after intense rowing exercise in healthy humans receiving salbutamol. J. Appl. Physiol. 2018, 125, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Moratinos, J.; Reverte, M. Effects of catecholamines on plasma potassium: The role of alpha- and beta-adrenoceptors. Fundam. Clin. Pharm. 1993, 7, 143–153. (In English) [Google Scholar] [CrossRef]
- Brown, M.J.; Brown, D.C.; Murphy, M.B. Hypokalemia from beta2-receptor stimulation by circulating epinephrine. N. Engl. J. Med. 1983, 309, 1414–1419. (In English) [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.C.; Richter, W.; Conti, M. Insights into the physiological functions of PDE4 from knockout mice. In Cyclic Nucleotide Phosphodiesterases in Health and Disease; Beavo, J., Francis, S., Houslay, M., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 323–346. [Google Scholar]
- Huston, E.; Lynch, M.J.; Mohamed, A.; Collins, D.M.; Hill, E.V.; MacLeod, R.; Krause, E.; Baillie, G.S.; Houslay, M.D. EPAC and PKA allow cAMP dual control over DNA-PK nuclear translocation. Proc. Natl. Acad. Sci. USA 2008, 105, 12791–12796. (In English) [Google Scholar] [CrossRef] [PubMed]
- Marti, G.; Schwarz, C.; Leichtle, A.B.; Fiedler, G.M.; Arampatzis, S.; Exadaktylos, A.K.; Lindner, G. Etiology and symptoms of severe hypokalemia in emergency department patients. Eur. J. Emerg. Med. Off. J. Eur. Soc. Emerg. Med. 2014, 21, 46–51. [Google Scholar] [CrossRef]
- Viera, A.J.; Wouk, N. Potassium Disorders: Hypokalemia and Hyperkalemia. Am. Fam. Physician 2015, 92, 487–495. [Google Scholar]
- Kardalas, E.; Paschou, S.A.; Anagnostis, P.; Muscogiuri, G.; Siasos, G.; Vryonidou, A. Hypokalemia: A clinical update. Endocr. Connect. 2018, 7, R135–R146. [Google Scholar] [CrossRef] [Green Version]
- McDonough, A.A.; Youn, J.H. Role of muscle in regulating extracellular [K+]. Semin. Nephrol. 2005, 25, 335–342. (In English) [Google Scholar] [CrossRef]
- Struthers, A.D.; Davies, D.L.; Harland, D.; Price, J.S.; Brown, R.A.; Quigley, C.; Brown, M.J. Adrenaline causes potassium influx in skeletal muscle and potassium efflux in cardiac muscle in rats: The role of Na/K ATPase. Life Sci. 1987, 40, 101–108. (In English) [Google Scholar] [CrossRef]
- Hayoz, S.; Cubano, L.; Maldonado, H.; Bychkov, R. Protein kinase A and C regulate leak potassium currents in freshly isolated vascular myocytes from the aorta. PLoS ONE 2013, 8, e75077. [Google Scholar] [CrossRef] [Green Version]
- Paris, S.; Rozengurt, E. Cyclic AMP stimulation of Na-K pump activity in quiescent swiss 3T3 cells. J. Cell. Physiol. 1982, 112, 273–280. (In English) [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Bossuyt, J.; Martin, J.L.; Despa, S.; Bers, D.M. Role of phospholemman phosphorylation sites in mediating kinase-dependent regulation of the Na+-K+-ATPase. Am. J. Physiol. Cell Physiol. 2010, 299, C1363–C1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geering, K. FXYD proteins: New regulators of Na-K-ATPase. Am. J. Physiol. Ren. Physiol. 2006, 290, F241–F250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boguslavskyi, A.; Tokar, S.; Prysyazhna, O.; Rudyk, O.; Sanchez-Tatay, D.; Lemmey, H.A.L.; Dora, K.A.; Garland, C.J.; Warren, H.R.; Doney, A.; et al. Phospholemman Phosphorylation Regulates Vascular Tone, Blood Pressure, and Hypertension in Mice and Humans. Circulation 2021, 143, 1123–1138. [Google Scholar] [CrossRef] [PubMed]
- Connors, E.C.; Ballif, B.A.; Morielli, A.D. Homeostatic regulation of Kv1.2 potassium channel trafficking by cyclic AMP. J. Biol. Chem. 2008, 283, 3445–3453. [Google Scholar] [CrossRef]
- Cui, J.; Melman, Y.; Palma, E.; Fishman, G.I.; McDonald, T.V. Cyclic AMP regulates the HERG K(+) channel by dual pathways. Curr. Biol. 2000, 10, 671–674. [Google Scholar] [CrossRef] [Green Version]
- Bibert, S.; Roy, S.; Schaer, D.; Horisberger, J.D.; Geering, K. Phosphorylation of phospholemman (FXYD1) by protein kinases A and C modulates distinct Na,K-ATPase isozymes. J. Biol. Chem. 2008, 283, 476–486. (In English) [Google Scholar] [CrossRef] [Green Version]
- Boguslavskyi, A.; Pavlovic, D.; Aughton, K.; Clark, J.E.; Howie, J.; Fuller, W.; Shattock, M.J. Cardiac hypertrophy in mice expressing unphosphorylatable phospholemman. Cardiovasc. Res. 2014, 104, 72–82. (In English) [Google Scholar] [CrossRef]
- Pavlovic, D.; Fuller, W.; Shattock, M.J. Novel regulation of cardiac Na pump via phospholemman. J. Mol. Cell. Cardiol. 2013, 61, 83–93. (In English) [Google Scholar] [CrossRef] [Green Version]
- Fuller, W.; Howie, J.; McLatchie, L.M.; Weber, R.J.; Hastie, C.J.; Burness, K.; Pavlovic, D.; Shattock, M.J. FXYD1 phosphorylation in vitro and in adult rat cardiac myocytes: Threonine 69 is a novel substrate for protein kinase C. Am. J. Physiol. Cell. Physiol. 2009, 296, C1346–C1355. (In English) [Google Scholar] [CrossRef] [Green Version]
- Reverte, M.; García-Barrado, M.J.; Moratinos, J. Role of alpha-adrenoceptors in control of plasma potassium in conscious rabbits. J. Auton. Pharm. 1991, 11, 305–313. (In English) [Google Scholar] [CrossRef]
- Brown, M.J. Hypokalemia from beta 2-receptor stimulation by circulating epinephrine. Am. J. Cardiol. 1985, 56, 3D–9D. (In English) [Google Scholar] [CrossRef]
- Li, H.; Zuo, J.; Tang, W. Phosphodiesterase-4 Inhibitors for the Treatment of Inflammatory Diseases. Front. Pharm. 2018, 9, 1048. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobin, P.; Belacel-Ouari, M.; Bedioune, I.; Zhang, L.; Leroy, J.; Leblais, V.; Fischmeister, R.; Vandecasteele, G. Cyclic nucleotide phosphodiesterases in heart and vessels: A therapeutic perspective. Arch. Cardiovasc. Dis. 2016, 109, 431–443. (In English) [Google Scholar] [CrossRef] [PubMed]
- Siuciak, J.A. The role of phosphodiesterases in schizophrenia: Therapeutic implications. CNS Drugs 2008, 22, 983–993. (In English) [Google Scholar] [CrossRef] [PubMed]
- Wahlang, B.; Mcclain, C.; Barve, S.; Gobejishvili, L. Role of cAMP and phosphodiesterase signaling in liver health and disease. Cell. Signal. 2018, 49, 105–115. [Google Scholar] [CrossRef]
- Hertz, L.; Xu, J.; Song, D.; Yan, E.; Gu, L.; Peng, L. Astrocytic and neuronal accumulation of elevated extracellular K(+) with a 2/3 K(+)/Na(+) flux ratio-consequences for energy metabolism, osmolarity and higher brain function. Front. Comput. Neurosci. 2013, 7, 114. (In English) [Google Scholar] [CrossRef] [Green Version]
- Espay, A.J. Neurologic complications of electrolyte disturbances and acid-base balance. Handb. Clin. Neurol. 2014, 119, 365–382. (In English) [Google Scholar] [CrossRef]
- Peng, T.; Qi, B.; He, J.; Ke, H.; Shi, J. Advances in the Development of Phosphodiesterase-4 Inhibitors. J. Med. Chem. 2020, 63, 10594–10617. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boyd, A.; Lochmaier, P.; Irelan, D.; Fiedler, E.; Lee, J.Y.; Fouty, B.; Abou Saleh, L.; Richter, W. Non-Selective PDE4 Inhibition Induces a Rapid and Transient Decrease of Serum Potassium in Mice. Biology 2022, 11, 1582. https://doi.org/10.3390/biology11111582
Boyd A, Lochmaier P, Irelan D, Fiedler E, Lee JY, Fouty B, Abou Saleh L, Richter W. Non-Selective PDE4 Inhibition Induces a Rapid and Transient Decrease of Serum Potassium in Mice. Biology. 2022; 11(11):1582. https://doi.org/10.3390/biology11111582
Chicago/Turabian StyleBoyd, Abigail, Peter Lochmaier, Daniel Irelan, Edward Fiedler, Ji Young Lee, Brian Fouty, Lina Abou Saleh, and Wito Richter. 2022. "Non-Selective PDE4 Inhibition Induces a Rapid and Transient Decrease of Serum Potassium in Mice" Biology 11, no. 11: 1582. https://doi.org/10.3390/biology11111582