
Taxonomic and Phylogenetic Insights into Novel Ascomycota from Forest Woody Litter



1
Center for Mountain Futures, Kunming Institute of Botany, Chinese Academy of Sciences, Honghe 654400, China
2
Department of Economic Plants and Biotechnology, Yunnan Key Laboratory for Wild Plant Resources, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
*
Authors to whom correspondence should be addressed.
Biology 2022, 11(6), 889; https://doi.org/10.3390/biology11060889
Submission received: 17 March 2022
/
Revised: 2 June 2022
/
Accepted: 7 June 2022
/
Published: 9 June 2022
(This article belongs to the Special Issue Studies on Species Diversity and Phylogeny of Ascomycota (Fungi))
Abstract
:Simple Summary
Studies suggest that fungi belonging to Ascomycota are sensitive to environmental changes which are disrupting ecosystems globally, with numerous extinction events, imbuing fungal diversity research with a sense of urgency. Thus, it is crucial we assess the diversity of Ascomycota across different habitats and substrates, and include programs to seek out novel taxa within Ascomycota. Our study provides some insights into the woody-based saprotrophic microfungi in Yunnan, China by introducing two novel Ascomycota species. Furthermore, these species were discovered by a 5-year-old boy, thus highlighting how young enthusiasts can be involved in field studies and make a significant impact.
Abstract
While surveying the mycobiomes of dead woody litter in Yunnan Province, China, numerous isolates with affinity to Pleosporales (Dothideomycetes, Ascomycota) were recovered. The present work characterizes two species associated with dead woody twigs found in terrestrial habitats in the Kunming area of Yunnan. The novel taxa were recognized based on a polyphasic approach, including morphological examination and multiple gene phylogenetic analyses (non-translated loci and protein-coding regions). Neokalmusia jonahhulmei sp. nov. is introduced in Didymosphaeriaceae (Pleosporales) as a woody-based saprobic ascomycete that possesses multiloculate ascostromata immersed under a black clypeus-like structure, and three-septate, brown, fusiform, guttulate ascospores. Thyridaria jonahhulmei (Thyridariaceae) is introduced with teleomorphic and anamorphic (coelomycetous) characteristics. The teleomorph has the following characteristics: globose to subglobose ascomata with an ostiolum, a pruinose layer of yellow to reddish- or orange-brown material appearing around the top of the ostiolar necks, and brown, ellipsoid to fusoid, two-to-three-septate, euseptate, rough-walled ascospores; the anamorph features pycnidial conidiomata, phialidic, ampulliform to doliiform, conidiogenous cells, and brown, guttulate, ellipsoidal, aseptate conidia.
1. Introduction
Forest ecosystems produce a large quantity of litter in various forms, such as leaves, branches, twigs, inflorescence, and other debris. Plant litter maintains pathways involved in nutrient cycling that are crucial to forest productivity [1,2,3,4,5,6]. A great proportion of total forest carbon (C) is contained in these woody debris, providing habitats for thousands of organisms, including fungi. Fungi play a critical role in the decomposition of woody litter as they are capable of actively decomposing lignin and other recalcitrant components found in these materials. However, comprehensive studies of the fungal taxonomic systematics of woody litter are scarce.
Most of the described woody-based fungal species lack biological and ecological information and reliable taxonomic status. Researchers have paid more attention to economic species, such as those producing edible mushrooms, and less attention to the narrowly distributed microfungal species responsible for the decomposition and recycling of woody material. However, in recent years, there have been numerous studies into woody-based microfungal occurrences in Yunnan Province, leading to reports of higher microfungal diversity, especially in Dothideomycetes [7,8,9,10,11,12,13,14,15,16]. At the Centre for Mountain Futures (Kunming Institute of Botany), researchers are investigating microfungal diversity across several substrates in southwest China, including leaf and woody litter, in order to clarify their taxonomy using morphology in conjunction with multigene phylogeny. Accordingly, we have isolated ascomycetes (Pleosporales) from woody litter, collected in a Pinus yunnanensis forest in Yunnan Province (Kunming), China (Figure 1). This study assesses the systematic categorization of two taxonomic novelties in Neokalmusia and Thyridaria, assesses morphological characteristics on natural substrates and in cultures, and conducts phylogenetic analyses.
2. Materials and Methods
2.1. Isolates and Specimens
During fieldwork in the Kunming region, Yunnan, China, characteristic black ascomata/conidiomata on dead woody twigs were collected during the dry season. The local environment features mixed forests (predominantly Fagaceae and Pinus spp., with Pinus yunnanensis being the most abundant tree species) (Figure 1) and a humid temperate climate at elevation 2080 masl. Specimens were stored in Ziploc plastic bags and taken to the laboratory. Isolations were made from single ascospores, according to the methods of Wanasinghe et al. [11]. Voucher specimens were preserved in the herbarium of the Kunming Institute of Botany (KUN-HKAS), and the living cultures were placed in the Culture Collection of Kunming Institute of Botany (KUMCC), Kunming, China. Nomenclatural novelties were deposited in MycoBank as outlined in http://www.MycoBank.org (accessed on 17 March 2022).
2.2. Morphological Observations
Examination of external structures of the fungal specimens were made using a Motic SMZ 168 Series stereo-microscope (Motic Asia, Kowloon, Hong Kong). Micro-morphological characters were examined and evaluated following the protocols provided by Wanasinghe et al. [10]. Macroscopic images of colonies were documented using an iPhone XS Max (Apple Inc., Cupertino, CA, USA) with daylight. Photoplates and images were processed using Adobe Photoshop CS6 (Adobe Systems, San Jose, CA, USA).
2.3. DNA Extraction, PCR Amplifications and Sequencing
The genomic DNA of each isolate was extracted from scraped mycelia following Wanasinghe et al. [10] using the Biospin Fungus Genomic DNA Extraction Kit-BSC14S1 (BioFlux, Shanghai, China). Primers for PCR amplification used were ITS (internal transcribed spacers) = ITS5/ITS4 [17], LSU (partial 28S large subunit rDNA) = LR0R/LR5 [18,19], SSU (partial 18S small subunit rDNA) = NS1/NS4 [17], tef1 (translation elongation factor 1-α) = EF1-983F/EF1-2218R [20,21], and rpb2 (RNA polymerase II second largest subunit) = fRPB2-5f/fRPB2-7cR [22]. The PCR protocols were programmed as described in The PCR protocols of SSU, LSU, ITS, and tef1 and were programmed as described in Wanasinghe et al. [11]. The PCR amplification condition of rpb2 was set as denaturation at 98 °C for 2 min, followed by 35 cycles of denaturation at 98 °C for 10 s, annealing at 52 °C for 10 s, and extension at 72 °C for 20 s, with a final extension step at 72 °C for 5 min. DNA sequencing were performed at a private company for sequencing (BGI, Ltd., Shenzhen, China).
2.4. Molecular Phylogenetic Analyses
2.4.1. Sequencing and Sequence Alignment
Sequences generated from SSU, LSU, ITS, tef1, and rpb2 were first used for BLASTn analyses. Based on BLAST similarities and relevant publications [23,24,25], closely related sequences were downloaded from GenBank (Table 1 and Table 2). Loci were aligned using MAFFT v. 7 [26,27]) under default conditions. Final improvements were made when necessary using BioEdit v.7.0.5.2 software [28].
2.4.2. Phylogenetic Analyses
The single-gene data sets were examined for topological incongruence among loci for members of the analyses. The conflict-free single alignments for each locus were combined into a multi-locus dataset. Concatenated alignment was used to construct maximum-likelihood (ML) and Bayesian (BI) phylogenetic analyses. MrModeltest v. 2.3 [29] was used under the Akaike Information Criterion (AIC) implemented in PAUP v. 4.0b10 to determine the evolutionary models for Bayesian and maximum-likelihood analyses.
The CIPRES webportal [30] was used to execute RAxML [31] and Bayesian analyses [32]. RAxML-HPC2 on XSEDE v. 8.2.10 [30] was used with default parameters and 1000 bootstrap repetitions to construct the ML analysis.
MrBayes analyses were performed setting GTR+I+G as the evolutionary model, with 2 M generations, sampling every 1000 generations, ending the run automatically when standard deviations of split frequencies dropped below 0.01, and with a burnin fraction of 0.25. ML bootstrap values equal or greater than 70% and BYPP greater than 0.95 are given above each node of every tree.
FigTree v1.4.0 program [33] was used to visualize the phylogenetic trees and reorganized in Microsoft Power Point (2007). The finalized datasets and trees were submitted to TreeBASE, submission ID: 29569 (http://purl.org/phylo/treebase/phylows/study/TB2:S29569 (accessed on 17 March 2022)).
3. Results
3.1. Phylogenetic Analyses
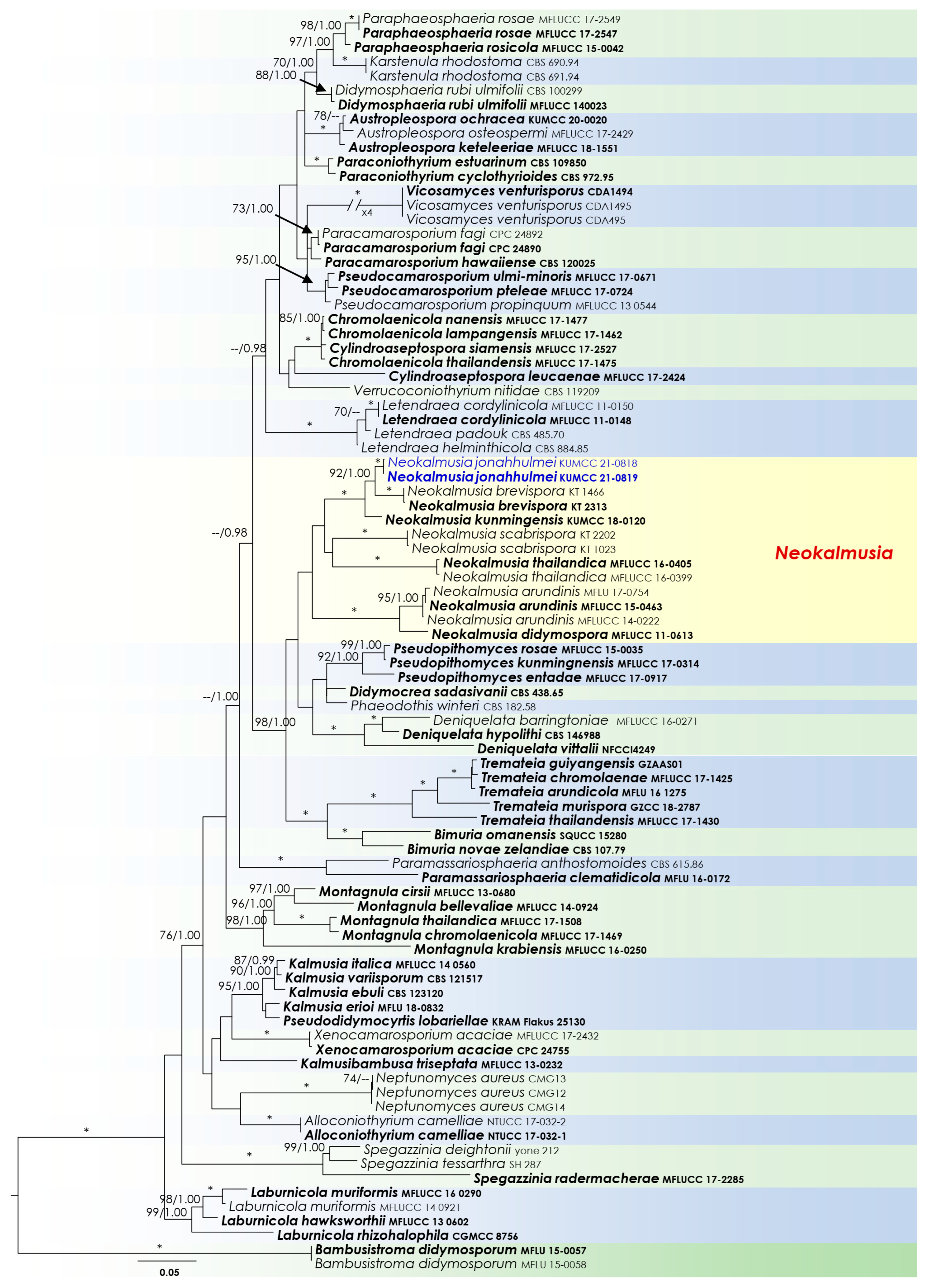
Two analyses were performed in this study. The first is a phylogenetic overview of the genera treated in Didymosphaeriaceae (Figure 2), while the remaining alignment represents the genera in Thyridariaceae (Figure 3). Other details related to both ML and BI analyses from Didymosphaeriaceae and Thyridariaceae datasets are presented in Table 3.
3.2. Taxonomy
Pleosporales Luttr. ex M.E. Barr, Prodromus to class Loculoascomycetes: 67 (1987)
Didymosphaeriaceae Munk, Dansk botanisk Arkiv 15 (2): 128 (1953)
Neokalmusia Ariyaw. & K.D. Hyde, Fungal Diversity 68: 92 (2014)
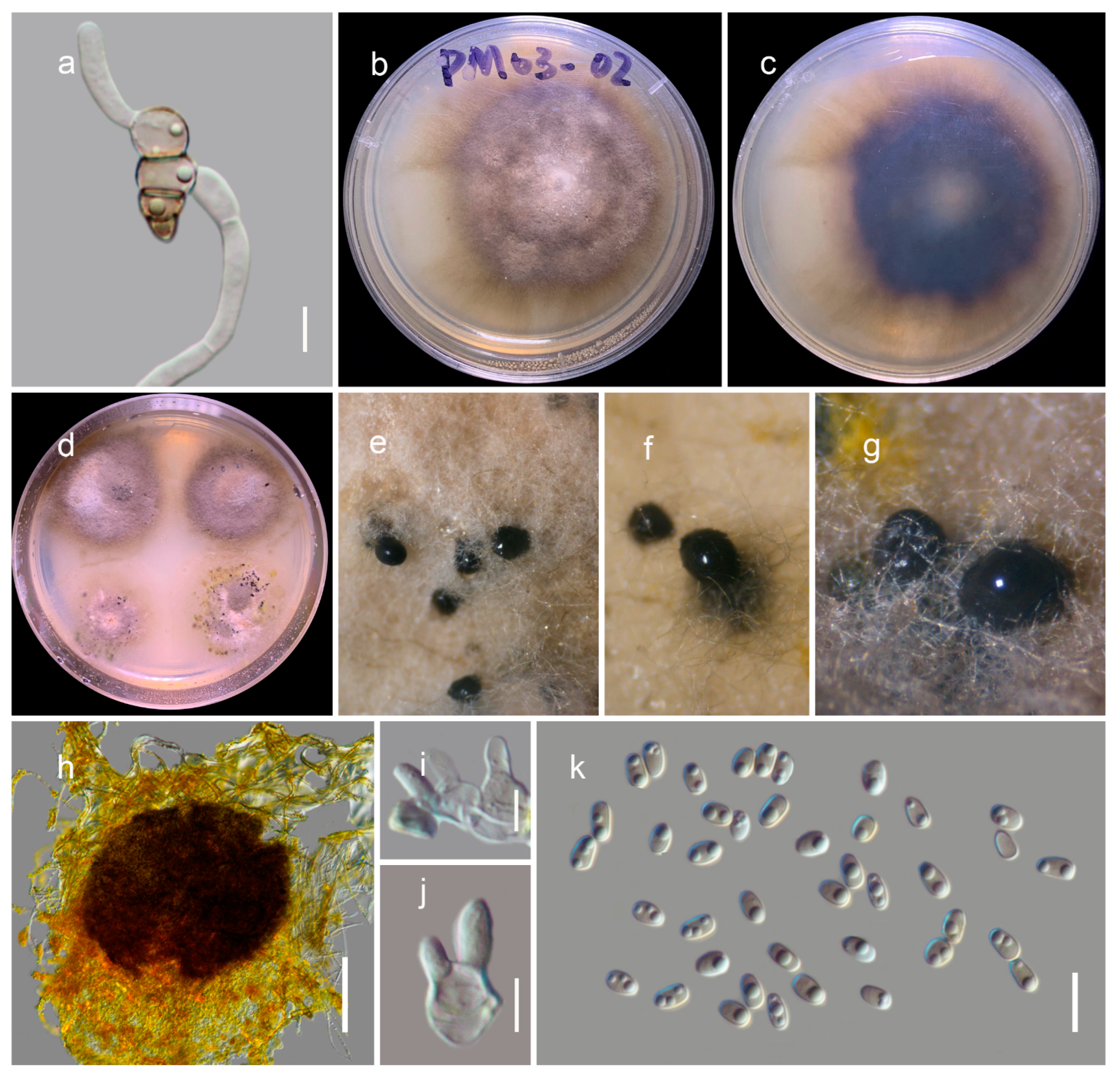
Neokalmusia jonahhulmei Wanas. & Mortimer sp. nov. (Figure 4)
MycoBank: MB843400
Etymology: The epithet is derived from Jonah Hulme Mortimer, who is the collector of this fungus.
Holotype: HKAS122910
Saprobic on dead bamboo culms. Teleomorph: Ascomata 200–300 μm high × 1200–1500 µm diam. (M = 180 × 1400 µm, n = 5), scattered or in groups, immersed under a black clypeus-like structure, composed of host epidermis and fungal mycelium, hemispherical, dark brown to black, multi-loculate. Locules 100–170 μm high, 150–280 μm diam. (M = 137.6 × 217.8 µm, n = 5), immersed within ascostromata, dark brown to black, subglobose to ampulliform, ostiolate. Peridium 10–12 μm wide at the base, 15–30 μm wide at the sides, comprising several layers; outer layers dark brown to brown, with compressed cells of textura angularis; inner layers hyaline, with compressed pseudoparenchymatous cells, arranged in textura angularis. Hamathecium contains 2–3 μm wide, branched, septate, cellular pseudoparaphyses. Asci 65–85 × 10–15 μm (M = 72.8 × 11.8 μm, n = 15), eight-spored, bitunicate, cylindrical to clavate, curved, short pedicel with slightly furcate ends, apically rounded. Ascospores 15–17 × 6–7 μm (M = 15.5 × 6.4 µm, n = 30), bi-seriate, overlapping and are initially hyaline, turning brown at maturity, fusiform, three-transversely septate, slightly curved, constricted at the septa, conically rounded at the ends, and smooth-walled, guttulated, without a distinct mucilaginous sheath. Anamorph: undetermined.
Culture characteristics: colonies reaching 4 cm diameter on PDA after 2 weeks at 20 °C. Mycelium dense, circular, slightly raised, smooth on surface and undulated floccose edge. Colony grey at the centre and coffee brown near margin from the top and dark brown at the bottom. Hyphae septate, branched, hyaline, thin, and smooth-walled.
Known distribution: Yunnan, China, on dead woody litter.
Material examined: China, Yunnan, Kunming, Wuhua, 25.131198 N, 102.590770 E, 2080 m, on dead bamboo culms (Phyllostachys sp.), 20 March 2021, Jonah Hulme Mortimer, PEM03-6-2-1 (HKAS122910, holotype), ex-holotype culture, KUMCC 21-0818. ibid. 25.131178 N, 102.590749 E, PEM03-6-2-4 (HKAS122911), living culture, KUMCC 21-0819.
Notes: The new fungus was collected from dead bamboo culms in Kunming. It is characterized as a typical Neokalmusia taxon based on its immersed, hemispherical multi-loculate ascomata under a black clypeus-like structure, bitunicate, clavate, apically rounded asci and bi-seriate, fusiform, brown ascospores with transverse septa [34]. It has a close phylogenetic affinity to Neokalmusia brevispora (KT 2313, KT 1466) and N. kunmingensis (KUMCC 18-0120), with 100 MLB and 1.00 BYPP support values (Figure 2). Neokalmusia brevispora and N. kunmingensis are reported as saprobes from Japan on the Sasa sp. and on dead bamboo culms in China, respectively [34,35,36]. These three species are morphologically similar with few dimensional differences in asci and ascospores. Neokalmusia brevispora has comparatively larger asci and ascospores (80–118 × 10.5–15 μm; 18–26.5 × 4–7 μm [34]) than N. kunmingensis (63–77 × 9.6–11.4 μm; 13–15 × 4–5 μm [36]) and our new collection (65–85 × 10–15 μm; 15–17 × 6–7 μm, this study). Further comparison of the ITS regions reveals Neokalmusia brevispora and N. kunmingensis feature nucleotide differences from N. jonahhulmei of 66/695 (9.5%) and 55/560 (9.8%), respectively. Comparison of the tef1 nucleotides of Neokalmusia brevispora and N. kunmingensis with our new strains revealed nucleotide differences of 31/917 (3.4%) and 26/904 (2.9%), respectively. The rpb2 region is not available for Neokalmusia brevispora and N. kunmingensis for comparison with our new species. This is the first time rpb2 sequence data for a Neokalmusia species have been provided (Table 1).
Ariyawansa et al. [34] established Neokalmusia to accommodate N. brevispora and N. scabrispora, which are characterized by immersed and subglobose to oblong ascomata with multiple perithecia, thin-walled clypeus-like structure, and verrucose ascospores. These two were found on dead culms of Sasa and Phyllostachys species in Japan, respectively [34,35]. Subsequent studies added four other species viz. N. arundinis (Italy) [37], N. didymospora (Thailand) [38], N kunmingensis (China) [36], and N. thailandica (Thailand) [37]. In this study, we introduce the seventh species in the genus from Phyllostachys species in China. All specimens of these species were obtained from dead culms of Bambusoideae (Poaceae) in China, Italy, Japan, and Thailand; therefore, Neokalmusia appear to be saprobic on bambusicolous hosts in temperate to tropical environments.
Thyridariaceae Q. Tian & K.D. Hyde, Fungal Diversity 63 (1): 254 (2013)
Thyridaria Sacc., Grevillea 4 (29): 21 (1875)
MycoBank: MB843401
Etymology: The epithet is derived from Jonah Hulme Mortimer, who is the collector of this fungus.
Holotype: HKAS122912
Saprobic on dead twigs of Fagaceae sp. Teleomorph: Ascomata 550–750 μm high, 200–300 μm diam. (M = 679.3 × 256.4 µm, n = 5), solitary or gregarious, immersed, coriaceous, heavily pigmented, globose to subglobose, ostiolate, yellow to reddish- or orange-brown pruinose layer forming around the apices of the ostiolar necks. Ostiole 100–150 μm long, 40–70 μm diam. (M = 121.1 × 57.8 µm, n = 5), central papillate, comprising hyaline periphyses. Peridium 15–25 μm wide, wider at the apex (60–70 μm), composed with two layers, with outer stratum comprising pale brown to brown, compressed, thick-walled cells of textura angularis, fused with the host tissues, and inner stratum multi-layered and composed with lightly pigmented to hyaline cells of textura angularis. Hamathecium comprised of 2–2.5 μm wide, branched, septate, cellular pseudoparaphyses, situated between and above the asci, embedded in a gelatinous matrix. Asci 150–200 × 18–22 μm (M = 170.2 × 20.1 μm, n = 25), eight-spored, bitunicate, fissitunicate, cylindrical to cylindric-clavate, long pedicellate (30–60 μm), and apex rounded with an ocular chamber. Ascospores 25–35 × 8–12.5 μm (M = 29.3 × 9.7 µm, n = 30), one-to-two-seriate, overlapping, and pale or yellowish brown when young, turning yellowish brown to brown at maturity, narrowly ellipsoid to fusoid, ends narrowly rounded (sometimes pointed), straight or curved, two-to-three-transversely septate, with median euseptum, slightly constricted at the septa, containing several guttules, with surface finely punctate to verruculose. Anamorph: Coelomycetous. Conidiomata (1–1.5 mm diam.), pycnidial, scattered or grouped, immersed and heavily pigmented. Pycnidial wall comprised with several strata, with brown-walled pseudoparenchymatous cells at outer margin becoming hyaline and thin-walled towards the inner conidiogenous cell-layer. Conidiogenous cells phoma-like, phialidic, ampulliform to doliiform, hyaline, flexuous, and smooth, with a short collarette. Conidia 5.5–8 × 2.5–3.5 μm (M = 6.9 × 3.1 μm, n = 50), first hyaline, turning pale brown, one-celled, straight or curved, ellipsoidal, rounded at both ends, thin and smooth-walled, comprising numerous guttules.
Culture characteristics: Colonies spreading on PDA up-to 4 cm diameter after 2 weeks at 20 °C, circular, whitish at the beginning, and becoming slightly raised and greenish-grey after 4 weeks, reverse dark brown. Hyphae septate, branched, hyaline, thin, smooth-walled, producing conidia after six weeks.
Known distribution: Yunnan, China, on dead woody litter.
Material examined: Material examined: China, Yunnan, Kunming, Wuhua, 25.131178 N, 102.590726 E, 2080 m, on dead twigs of woody litter of Fagaceae sp., 30 January 2021, Jonah Hulme Mortimer, PM03-2-1 (HKAS122912, holotype), ex-holotype culture, KUMCC 21-0816. ibid. 25.131157 N, 102.590708 E, 20 March 2021, PM03-2-3 (HKAS122913), living culture, KUMCC 21-0817.
Notes: Based on multi-gene sequence analyses (SSU, LSU, ITS, tef1 and rpb2), isolates KUMCC 21-0816 and KUMCC 21-0817 cluster with Thyridaria acaciae (CBS 138873), T. aureobrunnea (MFLUCC 21-0090) and T. broussonetiae (TB, TB1a, TB1, TB2) with 100 MLB and 1.00 BYPP bootstrap support values (Figure 3). Thyridaria acaciae is reported from Tanzania (on Acacia tortilis) [39], T. aureobrunnea from decaying wood in Thailand [40], and T. broussonetiae from Croatia (Hippocrepis emerus), Hungary (Amorpha fruticosa), and Italy (Broussonetia papyrifera) [41], whereas our new isolate is from dead woody twigs in Yunnan, China. Morphologically Thyridaria jonahhulmei has a close affinity to T. aureobrunnea and T. broussonetiae in its ascomata, asci, and ascospore characteristics. The asci dimensions of Thyridaria jonahhulmei (150–200 × 18–22 μm) are comparatively larger than T. aureobrunnea (45–61.5 × 7–8.5 µm) and are not significantly different from T. broussonetiae (109–183 × 12–19 µm). Comparison of the 546 ITS (+5.8S) nucleotides reveals 43 (7.9%) nucleotide differences between T. aureobrunnea and T. jonahhulmei. Thyridaria aureobrunnea lacks tef1 and rpb2 gene regions for comparison. Comparison of the 514 ITS (+5.8S) nucleotides of Thyridaria broussonetiae and T. jonahhulmei reveals 25 (4.86%) nucleotide differences, and in tef1 and rpb2 there are 8/712 (1.12%) and 18/1025 (1.75%) nucleotide differences, respectively. Thyridaria acaciae is known only from its asexual morph, and therefore it is not possible to compare their sexual morphologies. However, the asexual morph of Thyridaria jonahhulmei is similar to Thyridaria acacia. They both have ampulliform to doliiform conidiogenous cells and pale brown, aseptate conidia. Thyridaria acacia has subcylindrical conidia that lack guttules, whereas Thyridaria jonahhulmei has ellipsoidal conidia with large guttules. Comparison of the 469 ITS (+5.8S) nucleotides of these two strains reveals 23 (4.9%) nucleotide differences while tef1 and rpb2 gene regions were unavailable for comparison. Therefore, we recognize these isolates belong to two distinct species [42].
Excluding Thyridaria acaciae, T. aureobrunnea and T. broussonetiae, our new species resembles T. eminens (30 × 12 µm [43]), T. koae (14–21 × 6–9 µm [44]), T. minor (15–18 × 5–6.5 µm [45]), T. sambucina (12–15.5 µm [46]), T. subrufa (12–15 × 4–5 µm [46]), and T. triseptata (15–18 µm [47]) with its three-septate pigmented ascospores. Among them, Thyridaria eminens (collected from dead Streblus asper in Philippines [43]) is morphologically closely affiliated with T. jonahhulmei by ascospore dimensions. However, all of these species lack molecular data for further phylogenetic characterization.
4. Discussion
In this study, we describe and illustrate two new species of microfungi on dead woody litter, Neokalmusia jonahhulmei (Didymosphaeriaceae) and Thyridaria jonahhulmei (Thyridariaceae), from Kunming, Yunnan, based on morphological and molecular analyses (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5). Neokalmusia jonahhulmei is introduced with only its sexual characteristics, whereas Thyridaria jonahhulmei is accounted for with both asexual and sexual morphological features.
Didymosphaeriaceae was introduced by Munk [48], and, given that the family is composed of 33 genera, it is considered one of the most specious families in the order Pleosporales [23,25]. Members of Didymosphaeriaceae are known to form numerous different types of life modes, including saprobes, pathogens, or endophytes, and can be found both on land or in water [23,49]. We have presented representative sequence data of all currently available genera listed in Hongsanan et al. [23] and Samarakoon et al. [25] for the phylogenetic analyses (except Barria, Curreya, Julella, and Lineostroma, for which no DNA-based sequence data were available). Additionally, we included the sequences of Pseudodidymocyrtis lobariellae that were introduced by Flakus et al. [50] as a lichenicolous fungus from Bolivia on Lobariella pallida. Multi-gene phylogenetic analyses (Figure 2) revealed that Pseudodidymocyrtis clusters with Kalmusia species, and from the morphological perspective, these two genera appear to have a close resemblance. Therefore, generic delimitation needs defining among these two genera, possibly by using more fresh collections with additional morpho–phylo data.
Even though Thyridaria is one of the oldest genera (introduced in 1875) in Ascomycota, no exact family was available to accommodate this genus [51]. More than 30 species are included in Thyridaria [52]; however, for many of these species, data are lacking illustrations, descriptions, or DNA-based molecular data, leading to ambiguous taxonomic relationships. Therefore, Thyridaria species have suffered from uncertain family placement and have been assigned to different families in Dothideomycetes at various times [41,53,54,55,56,57,58,59,60,61]. Hyde et al. [51] considered its unique morphology and the phylogenetic placement of Thyridaria rubronotata in the Dothideomycetes backbone tree and introduced Thyridariaceae to accommodate this genus. Recently, Jaklitsch and Voglmayr [41] studied several thyridaria-like genera and constructed a multi-gene analysis to clarify intergeneric taxonomic affinities of Thyridaria in the Pleosporales. They found that thyridaria-like genera are distributed among at least nine clades of the Pleosporales. Barr [61] thoroughly reviewed various descriptions of Thyridaria and suggested that a key characteristic of the genus is an ample subiculum surrounding fruiting bodies that are produced under the ascomata wall or in host tissues. Similarly, the intensive study of Jaklitsch and Voglmayr [41] pointed out that this feature is exclusive to delineate Thyridaria species from its phylogenetically closely related affiliates. We also observed that our new Thyridaria species was characterized with a pruinose layer of yellow to reddish- or orange-brown material forming around the apices of the ostiolar necks. The subiculum of Thyridaria species could be an adaptation for its terrestrial-based habitat, and their potential advantages should be further clarified with extensive sampling from both aquatic and non-aquatic habitats.
By contrast, wood-decaying Basidiomycota in tropical China are well studied, which has facilitated a better understanding of global Basidiomycota species diversity and systematics [62,63,64,65,66,67]. Nonetheless, the study of habitat properties and potential lifestyles of woody-based microfungal species is especially important to understanding their functional aspects. The trophic mode of many species can be highly variable, with the ability to switch between mutualistic, pathogenic, and saprotrophic strategies. Pathogenic taxa that cause leaf spots may begin as endophytes, but environmental stressors may necessitate becoming pathogenic or eventually saprotrophic after plant tissue [68,69]. Another example is Scedosporiosis, the human disease caused by Pseudallescheria/Scedosporium complex species (PSC), which can grow saprobically in wood [70,71]. Therefore, owing to their importance in all ecosystems, saprotrophic fungi simply cannot be overlooked in any region.
This study provides some insights into the saprotrophic taxa and contributes knowledge of microfungi associated with woody litter in Yunnan, China by introducing two novel species. Furthermore, these species were discovered by a 5-year-old boy, unassisted by any mycologists at the time of collection, thus highlighting how young enthusiasts can be involved in field studies and make a significant impact.
Author Contributions
Conceptualization, D.N.W.; resources, P.E.M.; writing—original draft preparation, D.N.W.; writing—review and editing, P.E.M.; supervision, P.E.M.; funding acquisition, P.E.M. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was provided by the High-End Foreign Experts in the High-Level Talent 318 Recruitment Plan of Yunnan Province, 2021; the CAS President’s International Fellowship Initiative (number 2021FYB0005); the National Science Foundation of China (NSFC) under the project code 32150410362; and the Postdoctoral Fund from Human Resources and Social Security Bureau of Yunnan Province.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated for this study can be found in the NCBI GenBank, MycoBank, and TreeBASE.
Acknowledgments
Austin G. Smith at World Agroforestry (ICRAF), Kunming Institute of Botany, China, is thanked for English editing. Shaun Pennycook is thanked for nomenclatural advice. Lu Wen Hua and Li Qin Xian are thanked for their invaluable assistance. We acknowledge Kunming Institute of Botany and the Chinese Academy of Sciences for providing the laboratories and instruments for molecular work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bray, J.R.; Gorham, E. Litter production in forests of the world. Adv. Ecol. Res. 1964, 2, 101–157. [Google Scholar] [CrossRef]
- Lowman, M.D. Litter fall and leaf decay in three Australian rainforest formations. J. Ecol. 1988, 76, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Tsukamoto, J.; Tokumoto, Y.; Shuvo, M.A.R. The role of quantitative traits of leaf litter on decomposition and nutrient cycling of the forest ecosystems. J. For. Sci. 2013, 29, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Zhou, G.; Xu, Z. Forest litter fall and its composition: A new data set of observational data from China. Ecology 2016, 97, 1365. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Lang, X.; Du, B.; Zhang, H.; Liu, H.; Zhang, Y.; Shang, L. Litter fall and nutrient return in moist evergreen broad-leaved primary forest and mixed subtropical secondary deciduous broad-leaved forest in China. Eur. J. For. Res. 2016, 135, 77–86. [Google Scholar] [CrossRef]
- Giweta, M. Role of litter production and its decomposition, and factors affecting the processes in a tropical forest ecosystem: A review. J. Ecol. Environ. 2020, 44, 11. [Google Scholar] [CrossRef]
- Bao, D.F.; Luo, Z.L.; Liu, J.K.; Bhat, D.J.; Sarunya, N.; Li, W.L.; Su, H.Y.; Hyde, K.D. Lignicolous freshwater fungi in China III: Three new species and a new record of Kirschsteiniothelia from northwestern Yunnan province. Mycosphere 2018, 9, 755–768. [Google Scholar] [CrossRef]
- Huang, S.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bhat, D.J.; Chomnunti, P.; Hyde, K.D.; Lumyong, S. Morphological and molecular taxonomy of Jahnula dianchia sp. nov. (Jahnulales) from submerged wood in Dianchi Lake, Yunnan China. Mycol. Prog. 2018, 17, 547–555. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Su, H.W.Q. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Wijayawardene, N.N.; Xu, J.; Cheewangkoon, R.; Mortimer, P.E. Taxonomic novelties in Magnolia-associated pleosporalean fungi in the Kunming Botanical Gardens (Yunnan, China). PLoS ONE 2020, 15, e0235855. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Xu, J. Insight into the systematics of microfungi colonizing dead woody twigs of Dodonaea viscosa in Honghe (China). J. Fungi 2021, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, P.E.; Jeewon, R.; Xu, J.C.; Lumyong, S.; Wanasinghe, D.N. Morpho-phylo taxonomy of novel dothideomycetous fungi associated with dead woody twigs in Yunnan Province, China. Front. Microbiol. 2021, 12, 654683. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Hua, Z.; Lv, R.; Yu, Z. Neodactylariales, Neodactylariaceae (Dothideomycetes, Ascomycota): New order and family, with a new species from China. MycoKeys 2020, 73, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Rathnayaka, A.R.; Dayarathne, M.C.; Maharachchikumbura, S.S.N.; Liu, J.K.; Tennakoon, D.S.; Hyde, K.D. Introducing Seriascoma yunnanense sp. nov. (Occultibambusaceae, Pleosporales) based on evidence from morphology and phylogeny. AJOM 2019, 2, 245–253. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Wanasinghe, D.N.; Worthy, F.; Karunarathna, S.C. Addition to Melanommataceae: A new geographical record of Alpinaria rhododendri from Shangri La, China. AJOM 2020, 3, 335–344. [Google Scholar] [CrossRef]
- Yasanthika, E.; Dissanayake, L.S.; Wanasinghe, D.N.; Karunarathna, S.C.; Mortimer, P.E.; Samarakoon, B.C.; Monkai, J.; Hyde, K.D. Lonicericola fuyuanensis (Parabambusicolaceae) a new terrestrial pleosporalean ascomycete from Yunnan Province, China. Phytotaxa 2020, 446, 103–113. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols Appl. 1990, 18, 315–322. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Poli, A.; Bovio, E.; Ranieri, L.; Varese, G.C.; Prigione, V. News from the Sea: A New Genus and Seven New Species in the Pleosporalean Families Roussoellaceae and Thyridariaceae. Diversity 2020, 12, 144. [Google Scholar] [CrossRef] [Green Version]
- Samarakoon, B.C.; Wanasinghe, D.N.; Samarakoon, M.C.; Phookamsak, R.; McKenzie, E.H.C.; Chomnunti, P.; Hyde, K.D.; Lumyong, S.; Karunarathna, S.C. Multi-gene phylogenetic evidence suggests Dictyoarthrinium belongs in Didymosphaeriaceae (Pleosporales, Dothideomycetes) and Dictyoarthrinium musae sp. nov. on Musa from Thailand. MycoKeys 2020, 71, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Kuraku, S.; Zmasek, C.M.; Nishimura, O.; Katoh, K. aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity. Nucleic Acids Res. 2013, 41, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nylander, J.A.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree Version 1.4.0. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 23 January 2022).
- Ariyawansa, H.A.; Tanaka, K.; Thambugala, K.M.; Phookamsak, R.; Tian, Q.; Camporesi, E.; Hongsanan, S.; Monkai, J.; Wanasinghe, D.N.; Mapook, A.; et al. A molecular phylogenetic reappraisal of the Didymosphaeriaceae (=Montagnulaceae). Fungal Divers. 2014, 68, 69–104. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.P.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Thambugala, K.M.; Wanasinghe, D.N.; Phillips, A.J.L.; Camporesi, E.; Bulgakov, T.S.; Phukhamsakda, C.; Ariyawansa, H.A.; Goonasekara, I.D.; Phookamsak, R.; Dissanayake, A.; et al. Mycosphere notes 1-50: Grass (Poaceae) inhabiting Dothideomycetes. Mycosphere 2017, 8, 697–796. [Google Scholar] [CrossRef]
- Dai, D.Q.; Bahkali, A.H.; Ariyawansa, H.A.; Li, W.J.; Bhat, D.J.; Hyde, K.D. Neokalmusia didymospora (Didymosphaeriaceae), a new species from bamboo. Sydowia 2016, 68, 17–25. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Summerell, B.A.; Giraldo, A.; Gené, J.; Guarro, J.; Wanasinghe, D.N.; Hyde, K.D.; Camporesi, E.; et al. Fungal Planet Description Sheets: 281–319. Persoonia 2014, 33, 212–289. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Hidden diversity in Thyridaria and a new circumscription of the Thyridariaceae. Stud. Mycol. 2016, 85, 35–64. [Google Scholar] [CrossRef] [Green Version]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Rehm, H. Ascomycetes philippinenses—VIII. Leafl. Philipp. Bot. 1916, 8, 2945. [Google Scholar]
- Petrak, F. Ein Beitrag zur Pilzflora von Hawai. Sydowia 1952, 6, 363–371. [Google Scholar]
- Saccardo, P.A. Sylloge fungorum XXIV, sect. II. Abellini. Sylloge Fungorum. 1928, 24, 705–1438. [Google Scholar]
- Wehmeyer, L.E. The genus Thyridaria (Pyrenomycetes). Lloydia 1941, 4, 241–262. [Google Scholar]
- Barr, M.E. Melanommatales (Loculoascomycetes). N. Am. Flora Ser. II 1990, 13, 1–129. [Google Scholar]
- Munk, A. The system of the Pyrenomycetes. Dansk botanisk Arkiv. 1953, 15, 1–163. [Google Scholar]
- Gonçalves, M.F.M.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Neptunomyces aureus gen. et sp. nov. (Didymosphaeriaceae, Pleosporales) isolated from algae in Ria de Aveiro, Portugal. MycoKeys 2019, 60, 31–44. [Google Scholar] [CrossRef]
- Flakus, A.; Etayo, J.; Miadlikowska, J.; Lutzoni, F.; Kukwa, M.; Matura, N.; Rodriguez-Flakus, P. Biodiversity assessment of ascomycetes inhabiting Lobariella lichens in Andean cloud forests led to one new family, three new genera and 13 new species of lichenicolous fungi. Plant Fungal Syst. 2019, 64, 283–344. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Gareth, J.E.B.; Liu, J.K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Species Fungorum. Available online: http://www.speciesfungorum.org/Names/Names.asp (accessed on 23 January 2022).
- Wehmeyer, L.E. A revision of Melanconis, Pseudovalsa, Prosthecium and Titania. Univ. Mich. Stud. Sci. Ser. 1941, 14, 1–161. [Google Scholar]
- Wehmeyer, L.E. The Pyrenomycetous Fungi. Mycologia Memoir No. 6. The New York Botanical Garden; J. Cramer Publishing: Lehre, Germany, 1975; pp. 1–250. [Google Scholar]
- Munk, A. Danish pyrenomycetes: A preliminary flora. Dansk Botanisk Arkiv 1957, 17, 491. [Google Scholar]
- Luttrell, E.S. Loculoascomycetes. In The Fungi. An Advanced Treatise, a Taxonomic Review with Keys: Ascomycetes and Fungi Imperfecti; Ainsworth, G.C., Sparrow, F.K., Sussman, A.S., Eds.; Academic Press: New York, NY, USA, 1973; pp. 135–219. [Google Scholar]
- Müller, E.; von Arx, J.A. Pyrenomycetes: Meliolales, Coronophorales, Sphaeriales. New Exot. Fungi. Grevillea 1973, 17, 42–43. [Google Scholar]
- Dennis, R.W.G. British Ascomycetes, 3rd ed.; Cramer, J.: Vaduz, Liechtenstein, 1978; pp. 1–585. [Google Scholar]
- Barr, M.E. A classification of Loculoascomycetes. Mycologia 1979, 71, 935–995. [Google Scholar] [CrossRef]
- Barr, M.E. On the Massariaceae in North America. Mycotaxon 1979, 9, 17–37. [Google Scholar]
- Barr, M.E. The affinities of Thyridaria. Mycotaxon 2003, 88, 271–278. [Google Scholar]
- Dai, Y.C.; Cui, B.K.; Yuan, H.S.; He, S.H.; Wei, Y.L.; Qin, W.M.; Zhou, L.W.; Li, H.J. Wood-inhabiting fungi in southern China 4. Polypores from Hainan province. Ann. Bot. Fenn. 2011, 48, 219–231. [Google Scholar] [CrossRef]
- Dai, Y.; Härkönen, M.; Niemelä, T. Wood-inhabiting fungi in southern China 1. Polypores from Hunan province. Ann. Bot. Fenn. 2003, 40, 381–393. [Google Scholar]
- Dai, Y.; Wei, Y.; Wang, Z. Wood-inhabiting fungi in southern China 2. Polypores from Sichuan province. Ann. Bot. Fenn. 2004, 41, 319–329. [Google Scholar]
- Cui, B.K.; Dai, Y.; Bao, H.Y. Wood-inhabiting fungi in southern China 3. A new species of Phellinus (Hymenochaetales) from tropical China. Mycotaxon 2009, 110, 125–130. [Google Scholar] [CrossRef]
- Wang, B.; Cui, B.K.; Li, H.J.; Du, P.; Jia, B.S. Wood-rotting fungi in eastern China 5. Polypore diversity in Jiangxi Province. Ann. Bot. Fenn. 2011, 48, 237–246. [Google Scholar] [CrossRef]
- Yuan, H.; Dai, Y. Wood-inhabiting fungi in southern China. 6. Polypores from Guangxi autonomous region. Ann. Bot. Fenn. 2012, 49, 341–351. [Google Scholar] [CrossRef]
- Promputtha, I.; Lumyong, S.; Dhanasekaran, V.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. A phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Photita, W.; Lumyong, S.; Lumyong, P.; McKenzie, E.H.C.; Hyde, K.D. Are some endophytes from Musa acuminate latent pathogens? Fungal Divers. 2004, 16, 131–140. [Google Scholar]
- Luplertlop, N. Pseudallescheria/Scedosporium complex species: From saprobic to pathogenic fungus. J. Mycol. Med. 2018, 2, 249–256. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
Figure 1.
Habitat. (a–d) Mixed forest (predominantly Fagaceae and Pinus spp., with Pinus yunnanensis being the most abundant tree species); (e) collector.
Figure 1.
Habitat. (a–d) Mixed forest (predominantly Fagaceae and Pinus spp., with Pinus yunnanensis being the most abundant tree species); (e) collector.

Figure 2.
RAxML phylogram generated from combined dataset of partial SSU, LSU, ITS, tef1, and rpb2 DNA sequence analyses for Didymosphaeriaceae. The tree is rooted to Bambusistroma didymosporum (MFLU 15-0057, MFLU 15-0058). Bootstrap supports ML (MLB) ≥ 70% and Bayesian posterior probabilities (BYPP) ≥ 0.95 are given as MLB/BYPP above the branches. Branches with an asterisk (*) indicate MLB = 100% and BYPP = 1.00. The newly generated isolates are in blue.
Figure 2.
RAxML phylogram generated from combined dataset of partial SSU, LSU, ITS, tef1, and rpb2 DNA sequence analyses for Didymosphaeriaceae. The tree is rooted to Bambusistroma didymosporum (MFLU 15-0057, MFLU 15-0058). Bootstrap supports ML (MLB) ≥ 70% and Bayesian posterior probabilities (BYPP) ≥ 0.95 are given as MLB/BYPP above the branches. Branches with an asterisk (*) indicate MLB = 100% and BYPP = 1.00. The newly generated isolates are in blue.

Figure 3.
Phylogram generated from RAxML based on a concatenated sequence dataset of partial SSU, LSU, ITS, tef1, and rpb2 DNA for Thyridariaceae. Bootstrap values equal to or greater than 70% for ML (MLB) and Bayesian posterior probabilities (BYPP) ≥ 0.95 are shown at each node (as MLB/BYPP). An asterisk (*) represents branches with MLB = 100% and BYPP = 1.00. The new isolates are show in in blue.
Figure 3.
Phylogram generated from RAxML based on a concatenated sequence dataset of partial SSU, LSU, ITS, tef1, and rpb2 DNA for Thyridariaceae. Bootstrap values equal to or greater than 70% for ML (MLB) and Bayesian posterior probabilities (BYPP) ≥ 0.95 are shown at each node (as MLB/BYPP). An asterisk (*) represents branches with MLB = 100% and BYPP = 1.00. The new isolates are show in in blue.

Figure 4.
Neokalmusia jonahhulmei (HKAS122910, holotype). (a,b) Ascostromata on the bamboo culms; (c) horizontal section of ascostromata; (d,e) vertical sections of ascomata; (f) vertical section through two locules; (g) close-up of ostiole; (h) peridium; (i) pseudoparaphyses; (j–l) asci; (m–r) ascospores (r germinated ascospore); (s,t) colonies on PDA after 21 days. Scale bars, (d–f) 100 µm; (g) 50 µm; (h,m–r) 10 µm; (i) 5 µm; (j–l) 20 µm.
Figure 4.
Neokalmusia jonahhulmei (HKAS122910, holotype). (a,b) Ascostromata on the bamboo culms; (c) horizontal section of ascostromata; (d,e) vertical sections of ascomata; (f) vertical section through two locules; (g) close-up of ostiole; (h) peridium; (i) pseudoparaphyses; (j–l) asci; (m–r) ascospores (r germinated ascospore); (s,t) colonies on PDA after 21 days. Scale bars, (d–f) 100 µm; (g) 50 µm; (h,m–r) 10 µm; (i) 5 µm; (j–l) 20 µm.

Figure 5.
Sexual morph of Thyridaria jonahhulmei (HKAS122912, holotype). (a,b) Ascomata on the dead woody twigs; (c,d) cross section of ascomata; (e) close-up of ostiole; (f) peridium; (g) pseudoparaphyses; (h–j) asci; (k–p) ascospores. Scale bars, (d) 200 µm; (e) 50 µm; (f,h–j) 20 µm; (f,k–p) 10 µm.
Figure 5.
Sexual morph of Thyridaria jonahhulmei (HKAS122912, holotype). (a,b) Ascomata on the dead woody twigs; (c,d) cross section of ascomata; (e) close-up of ostiole; (f) peridium; (g) pseudoparaphyses; (h–j) asci; (k–p) ascospores. Scale bars, (d) 200 µm; (e) 50 µm; (f,h–j) 20 µm; (f,k–p) 10 µm.

Figure 6.
Asexual morph of Thyridaria jonahhulmei (KUMCC 21-081, ex-type culture). (a) Germinated ascospore (b–d) colony on PDA (c from the bottom); (e–g) immersed pycnidia in PDA; (h) squashed conidiomata; (i,j) conidiogenous cells; (k) conidia. Scale bars, (h) 100 µm; (i,j) 5 µm; (k) 10 µm.
Figure 6.
Asexual morph of Thyridaria jonahhulmei (KUMCC 21-081, ex-type culture). (a) Germinated ascospore (b–d) colony on PDA (c from the bottom); (e–g) immersed pycnidia in PDA; (h) squashed conidiomata; (i,j) conidiogenous cells; (k) conidia. Scale bars, (h) 100 µm; (i,j) 5 µm; (k) 10 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxa used in the phylogenetic analyses of Didymosphaeriaceae and their corresponding GenBank numbers of partial ITS, LSU, SSU, tef1, and rpb2 sequences. Isolates/sequences in bold were isolated/sequenced in the present study.
Table 1.
Taxa used in the phylogenetic analyses of Didymosphaeriaceae and their corresponding GenBank numbers of partial ITS, LSU, SSU, tef1, and rpb2 sequences. Isolates/sequences in bold were isolated/sequenced in the present study.
| Species | Strain | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | SSU | tef1 | rpb2 | ||
| Alloconiothyrium camelliae | NTUCC 17-032-1 | MT112294 | MT071221 | MT071270 | MT232967 | - |
| Alloconiothyrium camelliae | NTUCC 17-032-2 | MT112295 | MT071222 | MT071271 | MT232965 | - |
| Austropleospora keteleeriae | MFLUCC 18-1551 | NR_163349 | MK347910 | NG_070075 | MK360045 | MK434909 |
| Austropleospora ochracea | KUMCC 20-0020 | MT799859 | MT808321 | MT799860 | MT872714 | - |
| Austropleospora osteospermi | MFLUCC 17-2429 | MK347757 | MK347863 | MK347974 | MK360044 | MK434884 |
| Bambusistroma didymosporum | MFLU 15-0057 | KP761733 | KP761737 | KP761730 | KP761727 | KP761720 |
| Bambusistroma didymosporum | MFLU 15-0058 | KP761734 | KP761738 | KP761731 | KP761728 | KP761721 |
| Bimuria novae-zelandiae | CBS 107.79 | MH861181 | AY016338 | AY016356 | DQ471087 | DQ470917 |
| Bimuria omanensis | SQUCC 15280 | NR_173301 | - | NG_071257 | MT279046 | - |
| Chromolaenicola lampangensis | MFLUCC 17-1462 | MN325016 | MN325010 | MN325004 | MN335649 | MN335654 |
| Chromolaenicola nanensis | MFLUCC 17-1477 | MN325014 | MN325008 | MN325002 | MN335647 | MN335653 |
| Chromolaenicola thailandensis | MFLUCC 17-1475 | MN325019 | MN325013 | MN325007 | MN335652 | MN335656 |
| Cylindroaseptospora leucaenae | MFLUCC 17-2424 | NR_163333 | MK347856 | NG_066310 | MK360047 | - |
| Cylindroaseptospora siamensis | MFLUCC 17-2527 | NR_163337 | MK347866 | NG_066311 | MK360048 | - |
| Deniquelata barringtoniae | MFLUCC 16-0271 | MH275059 | - | MH260291 | MH412766 | MH412753 |
| Deniquelata hypolithi | CBS 146988 | MZ064429 | - | NG_076735 | MZ078250 | MZ078201 |
| Deniquelata vittalii | NFCCI4249 | MF406218 | MF622059 | MF182395 | MF182398 | MF168942 |
| Didymocrea sadasivanii | CBS 438.65 | MH858658 | DQ384066 | DQ384103 | - | - |
| Didymosphaeria rubi ulmifolii | CBS 100299 | MH862698 | AY642523 | JX496124 | - | - |
| Didymosphaeria rubi ulmifolii | MFLUCC 140023 | KJ436586 | NG_063557 | KJ436586 | - | - |
| Kalmusia ebuli | CBS 123120 | KF796674 | JN851818 | JN644073 | - | - |
| Kalmusia erioi | MFLU 18-0832 | MN473058 | MN473046 | MN473052 | MN481599 | - |
| Kalmusia italica | MFLUCC 14 0560 | KP325440 | KP325442 | KP325441 | - | - |
| Kalmusia variisporum | CBS 121517 | NR_145165 | - | JX496143 | - | - |
| Kalmusibambusa triseptata | MFLUCC 13-0232 | KY682697 | KY682696 | KY682695 | - | - |
| Karstenula rhodostoma | CBS 690.94 | - | GU296154 | GU301821 | GU349067 | GU371788 |
| Karstenula rhodostoma | CBS 691.94 | LC014559 | AB797241 | AB807531 | AB808506 | - |
| Laburnicola hawksworthii | MFLUCC 13 0602 | KU743194 | KU743196 | KU743195 | - | - |
| Laburnicola muriformis | MFLUCC 16 0290 | KU743197 | KU743199 | KU743198 | KU743213 | - |
| Laburnicola muriformis | MFLUCC 14 0921 | KU743200 | KU743202 | KU743201 | - | - |
| Laburnicola rhizohalophila | CGMCC 8756 | KJ125522 | - | KJ125523 | KJ125525 | KJ125524 |
| Letendraea cordylinicola | MFLUCC 11 0150 | KM213996 | KM214002 | KM213999 | - | - |
| Letendraea cordylinicola | MFLUCC 11 0148 | NR_154118 | KM214001 | NG_059530 | - | - |
| Letendraea helminthicola | CBS 884.85 | MK404145 | AY016345 | AY016362 | MK404174 | MK404164 |
| Letendraea padouk | CBS 485.70 | - | GU296162 | AY849951 | - | - |
| Montagnula bellevaliae | MFLUCC 14 0924 | NR_155377 | KT443904 | KT443902 | KX949743 | - |
| Montagnula chromolaenicola | MFLUCC 17-1469 | NR_168866 | NG_070157 | NG_070948 | MT235773 | MT235809 |
| Montagnula cirsii | MFLUCC 13 0680 | KX274242 | KX274255 | KX274249 | KX284707 | - |
| Montagnula krabiensis | MFLUCC 16-0250 | MH275070 | MH260343 | MH260303 | MH412776 | - |
| Montagnula thailandica | MFLUCC 17-1508 | MT214352 | NG_070158 | NG_070949 | MT235774 | MT235810 |
| Neokalmusia arundinis | MFLU 17-0754 | MT649882 | MT649880 | MT649878 | MT663766 | - |
| Neokalmusia arundinis | MFLUCC 15-0463 | NR_165852 | NG_068372 | NG_068237 | KY244024 | - |
| Neokalmusia arundinis | MFLUCC 14-0222 | KX965731 | KX986344 | KX954400 | KY271091 | - |
| Neokalmusia brevispora | KT 2313 | LC014574 | AB524460 | AB524601 | AB539113 | - |
| Neokalmusia brevispora | KT 1466 | LC014573 | AB524459 | AB524600 | AB539112 | - |
| Neokalmusia didymospora | MFLUCC 11-0613 | - | KP091435 | KP091434 | - | - |
| Neokalmusia jonahhulmei | KUMCC 21-0818 | ON007043 | ON007039 | ON007048 | ON009133 | ON009137 |
| Neokalmusia jonahhulmei | KUMCC 21-0819 | ON007044 | ON007040 | ON007049 | ON009134 | ON009138 |
| Neokalmusia kunmingensis | KUMCC 18-0120 | MK079886 | MK079887 | MK079889 | MK070172 | - |
| Neokalmusia scabrispora | KT 1023 | LC014575 | AB524452 | AB524593 | AB539106 | - |
| Neokalmusia scabrispora | KT 2202 | LC014576 | AB524453 | AB524594 | AB539107 | - |
| Neokalmusia thailandica | MFLUCC 16-0405 | NR_154255 | KY706137 | NG_059792 | KY706145 | KY706148 |
| Neokalmusia thailandica | MFLUCC 16-0399 | KY706141 | KY706136 | KY706131 | - | - |
| Neptunomyces aureus | CMG12 | MK912121 | - | - | MK948000 | - |
| Neptunomyces aureus | CMG13 | MK912122 | - | - | MK948001 | - |
| Neptunomyces aureus | CMG14 | MK912123 | - | - | MK948002 | - |
| Paracamarosporium fagi | CPC 24890 | NR_154318 | - | NG_070630 | - | - |
| Paracamarosporium fagi | CPC 24892 | KR611887 | - | KR611905 | - | - |
| Paracamarosporium hawaiiense | CBS 120025 | JX496027 | EU295655 | JX496140 | - | - |
| Paraconiothyrium cyclothyrioides | CBS 972.95 | JX496119 | AY642524 | JX496232 | - | - |
| Paraconiothyrium estuarinum | CBS 109850 | JX496016 | AY642522 | JX496129 | - | - |
| Paramassariosphaeria anthostomoides | CBS 615.86 | MH862005 | GU205246 | GU205223 | - | - |
| Paramassariosphaeria clematidicola | MFLU 16-0172 | KU743206 | KU743208 | KU743207 | - | - |
| Paraphaeosphaeria rosae | MFLUCC 17-2547 | MG828935 | MG829150 | MG829044 | MG829222 | - |
| Paraphaeosphaeria rosae | MFLUCC 17-2549 | MG828937 | MG829152 | MG829046 | MG829223 | - |
| Paraphaeosphaeria rosicola | MFLUCC 15-0042 | NR_157528 | MG829153 | MG829047 | - | - |
| Phaeodothis winteri | CBS 182.58 | - | GU296183 | GU301857 | - | - |
| Pseudocamarosporium propinquum | MFLUCC 13 0544 | KJ747049 | KJ819949 | KJ813280 | - | - |
| Pseudocamarosporium pteleae | MFLUCC 17-0724 | NR_157536 | MG829166 | MG829061 | MG829233 | - |
| Pseudocamarosporium ulmi-minoris | MFLUCC 17-0671 | NR_157537 | MG829167 | MG829062 | - | - |
| Pseudodidymocyrtis lobariellae | KRAM Flakus 25130 | NR_169714 | NG_070349 | NG_068933 | - | - |
| Pseudopithomyces entadae | MFLUCC 17-0917 | MK347835 | NG_066305 | MK360083 | MK434899 | |
| Pseudopithomyces kunmingnensis | MFLUCC 17-0314 | MF173607 | MF173606 | MF173605 | - | - |
| Pseudopithomyces rosae | MFLUCC 15-0035 | MG828953 | MG829168 | MG829064 | - | - |
| Spegazzinia deightonii | yone 212 | AB797292 | AB807582 | AB808558 | - | |
| Spegazzinia radermacherae | MFLUCC 17-2285 | MK347740 | MK347848 | MK347957 | MK360088 | MK434893 |
| Spegazzinia tessarthra | SH 287 | JQ673429 | AB797294 | AB807584 | AB808560 | - |
| Tremateia arundicola | MFLU 16 1275 | KX274241 | KX274254 | KX274248 | KX284706 | - |
| Tremateia chromolaenae | MFLUCC 17-1425 | NR_168868 | NG_070160 | NG_068710 | MT235778 | MT235816 |
| Tremateia guiyangensis | GZAAS01 | KX274240 | KX274253 | KX274247 | KX284705 | - |
| Tremateia murispora | GZCC 18-2787 | NR_165916 | MK972750 | MK972751 | MK986482 | - |
| Tremateia thailandensis | MFLUCC 17-1430 | NR_168869 | NG_070161 | NG_068711 | MT235781 | MT235819 |
| Verrucoconiothyrium nitidae | CBS 119209 | EU552112 | - | EU552112 | - | - |
| Vicosamyces venturisporus | CDA1494 | MF802825 | - | MF802828 | - | - |
| Vicosamyces venturisporus | CDA1495 | MF802826 | - | MF802829 | - | - |
| Vicosamyces venturisporus | CDA495 | MF802827 | - | MF802830 | - | - |
| Xenocamarosporium acaciae | CPC 24755 | NR_137982 | - | NG_058163 | - | - |
| Xenocamarosporium acaciae | MFLUCC 17-2432 | MK347766 | MK347873 | MK347983 | MK360093 | - |
Table 2.
Taxa used in the phylogenetic analyses of Thyridariaceae and their corresponding GenBank numbers of partial ITS, LSU, SSU, tef1, and rpb2 sequences. Isolates/sequences in bold were isolated/sequenced in the present study.
Table 2.
Taxa used in the phylogenetic analyses of Thyridariaceae and their corresponding GenBank numbers of partial ITS, LSU, SSU, tef1, and rpb2 sequences. Isolates/sequences in bold were isolated/sequenced in the present study.
| Species | Strain | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | SSU | tef1 | rpb2 | ||
| Arthopyrenia salicis | CBS 368.94 | KF443410 | AY538339 | AY538333 | KF443404 | KF443397 |
| Chromolaenomyces appendiculatus | MFLUCC 17-1455 | NR_168862 | NG_068705 | MT214394 | MT235770 | MT235806 |
| Cycasicola goaensis | MFLUCC 17-0754 | MG828885 | MG829001 | MG829112 | MG829198 | - |
| Cycasicola leucaenae | MFLUCC17-0914 | NR_163322 | MK347942 | MK347833 | MK360046 | MK434900 |
| Liua muriformis | KUMCC 18-0177 | MK433599 | MK433598 | MK433595 | MK426798 | MK426799 |
| Neoroussoella alishanense | AKW 11 FU31018 | MK503818 | MK503824 | MK503830 | MK336182 | MN037757 |
| Neoroussoella alishanense | AKW 03 FU31016 | MK503816 | MK503822 | MK503828 | MK336181 | MN037756 |
| Neoroussoella entadae | MFLUCC 18-0243 | MK347786 | MK348004 | MK347893 | MK360065 | MK434866 |
| Neoroussoella leucaenae | MFLUCC 18-1544 | MK347767 | MK347984 | MK347874 | MK360067 | MK434876 |
| Occultibambusa bambusae | MFLUCC 11-0394 | KU940124 | KU863113 | - | KU940194 | KU940171 |
| Occultibambusa bambusae | MFLUCC 13-0855 | KU940123 | KU863112 | KU872116 | KU940193 | KU940170 |
| Ohleria modesta | MGC | KX650562 | KX650562 | - | KX650533 | KX650582 |
| Ohleria modesta | OM | KX650563 | KX650563 | KX650513 | KX650534 | KX650583 |
| Parathyridaria clematidis | MFLUCC 17-2154 | MT310645 | MT214601 | MT226712 | MT394657 | MT394712 |
| Parathyridaria clematidis | MFLUCC 17-2157 | MT310644 | MT214600 | MT226711 | MT394656 | MT394711 |
| Parathyridaria clematidis | MFLUCC 17-2160 | MT310643 | MT214599 | MT226710 | MT394655 | MT394710 |
| Parathyridaria clematidis | MFLUCC 17-2185 | MT310642 | MT214598 | NG_070668 | MT394654 | MT394709 |
| Parathyridaria ellipsoidea | KNU-JJ-1829 | LC552950 | LC552952 | - | - | - |
| Parathyridaria flabelliae | MUT 4886 | KR014358 | KP671720 | KT587317 | MN605910 | MN605930 |
| Parathyridaria flabelliae | MUT 4859 | KR014355 | KP671716 | KT587315 | MN605909 | MN605929 |
| Parathyridaria percutanea | CBS 128203 | KF322117 | KF366448 | KF366450 | KF407988 | KF366453 |
| Parathyridaria percutanea | CBS 868.95 | KF322118 | KF366449 | KF366451 | KF407987 | KF366452 |
| Parathyridaria philadelphi | CBS 143432 | MH107905 | NG_063958 | - | MH108023 | - |
| Parathyridaria ramulicola | MUT 4397 | KC339235 | KF636775 | MN556311 | MN605913 | MN605933 |
| Parathyridaria ramulicola | CBS 141479 | NR_147657 | KX650565 | KX650514 | KX650536 | KX650584 |
| Parathyridaria robiniae | MUT 2452 | MG813183 | MG816491 | MN556312 | MN605903 | MN605923 |
| Parathyridaria robiniae | MUT 4893 | KM355998 | MN556328 | KM355993 | MN605904 | MN605924 |
| Parathyridaria robiniae | MFLUCC 14-1119 | KY511142 | KY511141 | - | KY549682 | - |
| Parathyridaria rosae | MFLU 17-0623 | NR_157530 | NG_059873 | - | - | - |
| Parathyridaria serratifoliae | MFLUCC 17-2210 | MT310646 | MT214602 | NG_070669 | MT394658 | MT394713 |
| Parathyridaria tyrrhenica | MUT 4966 | KR014366 | KP671740 | KT587309 | MN605911 | MN605931 |
| Parathyridaria tyrrhenica | MUT 5371 | KU314951 | MN556329 | KU314952 | MN605912 | MN605932 |
| Parathyridaria virginianae | MFLUCC 17-2163 | MT310647 | NG_073853 | NG_070670 | MT394659 | MT394714 |
| Parathyridariella dematiacea | MUT 4419 | KC339245 | KF636786 | MN556313 | MN605905 | MN605925 |
| Parathyridariella dematiacea | MUT 5310 | KU255057 | MN556330 | MN556314 | MN605907 | MN605927 |
| Parathyridariella dematiacea | MUT 5381 | KU314959 | MN556331 | KU314960 | MN605908 | MN605928 |
| Parathyridariella dematiacea | MUT 4884 | MN556317 | KP671726 | KT587329 | MN605906 | MN605926 |
| Pseudothyridariella chromolaenae | MFLUCC 17-1472 | NR_168863 | NG_068706 | MT214395 | MT235771 | MT235807 |
| Pseudothyridariella mahakoshae | NFCCI 4215 | MG020435 | MG020438 | MG020441 | MG023140 | MG020446 |
| Roussoella hysterioides | CBS 546.94 | KF443405 | KF443381 | AY642528 | KF443399 | KF443392 |
| Roussoella intermedia | CBS 170.96 | KF443407 | KF443382 | KF443390 | KF443398 | KF443394 |
| Roussoella japanensis | MAFF 239636 | KJ474829 | AB524621 | AB524480 | AB539114 | AB539101 |
| Roussoella margidorensis | MUT 5329 | KU314944 | MN556322 | MN556309 | MN605897 | MN605917 |
| Roussoella mediterranea | MUT 5306 | KU255054 | MN556323 | MN556310 | MN605898 | MN605918 |
| Roussoella padinae | MUT 5341 | KU158153 | MN556325 | KU158176 | MN605900 | MN605920 |
| Roussoella pustulans | KT 1709 | KJ474830 | AB524623 | AB524482 | AB539116 | AB539103 |
| Roussoella siamensis | MFLUCC 11-0149 | KJ474837 | KJ474845 | KU872125 | KJ474854 | KJ474861 |
| Thyridaria acaciae | CBS 138873 | KP004469 | KP004497 | - | - | - |
| Thyridaria aureobrunnea | MFLUCC 21-0090 | MZ538528 | MZ538562 | - | - | - |
| Thyridaria broussonetiae | TB | KX650567 | KX650567 | - | KX650538 | KX650585 |
| Thyridaria broussonetiae | TB1a | KX650569 | KX650569 | - | - | - |
| Thyridaria broussonetiae | TB2 | KX650570 | KX650570 | - | KX650540 | KX650587 |
| Thyridaria broussonetiae | TB1 | KX650568 | KX650568 | KX650515 | KX650539 | KX650586 |
| Thyridaria jonahhulmei | KUMCC 21-0816 | ON007041 | ON007037 | ON007046 | ON009131 | ON009135 |
| Thyridaria jonahhulmei | KUMCC 21-0817 | ON007042 | ON007038 | ON007047 | ON009132 | ON009136 |
| Thyridariella mangrovei | NFCCI 4214 | MG020436 | MG020439 | MG020442 | MG020444 | MG020447 |
| Thyridariella mangrovei | NFCCI 4213 | MG020434 | MG020437 | MG020440 | MG020443 | MG020445 |
| Torula herbarum | CBS 111855 | KF443409 | KF443386 | KF443391 | KF443403 | KF443396 |
| Torula herbarum | CBS 595.96 | KF443408 | KF443385 | KF443387 | KF443402 | KF443395 |
Table 3.
Maximum-likelihood (ML) and Bayesian (BI) analyses results for each sequenced dataset.
| Analyses | Didymosphaeriaceae | Thyridariaceae | |
|---|---|---|---|
| Number of taxa | 88 | 59 | |
| Gene regions | SSU, LSU, ITS, tef1, and rpb2 | SSU, LSU, ITS, tef1, and rpb2 | |
| Number of character positions (including gaps) | 5016 | 4529 | |
| ML optimization likelihood value | −35,672.743881 | −30,606.10565 | |
| Distinct alignment patterns in the matrix | 2249 | 1796 | |
| Number of undetermined characters or gaps (%) | 41.88% | 19.56% | |
| Estimated base frequencies | A | 0.240418 | 0.249274 |
| C | 0.253351 | 0.25578 | |
| G | 0.270784 | 0.267476 | |
| T | 0.235446 | 0.227469 | |
| Substitution rates | AC | 1.561664 | 1.486771 |
| AG | 3.248718 | 3.744601 | |
| AT | 1.433496 | 1.706836 | |
| CG | 1.323566 | 1.014483 | |
| CT | 7.428045 | 7.933665 | |
| GT | 1.0 | 1.0 | |
| Proportion of invariable sites (I) | 0.396829 | 0.505108 | |
| Gamma distribution shape parameter (α) | 0.454368 | 0.442817 | |
| Number of generated trees in BI | 11,301 | 2501 | |
| Number of trees sampled in BI after 25% were discarded as burn-in | 8476 | 1876 | |
| Final split frequency | 0.009959 | 0.009966 | |
| The total of unique site patterns | 2252 | 1798 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wanasinghe, D.N.; Mortimer, P.E. Taxonomic and Phylogenetic Insights into Novel Ascomycota from Forest Woody Litter. Biology 2022, 11, 889. https://doi.org/10.3390/biology11060889
AMA Style
Wanasinghe DN, Mortimer PE. Taxonomic and Phylogenetic Insights into Novel Ascomycota from Forest Woody Litter. Biology. 2022; 11(6):889. https://doi.org/10.3390/biology11060889
Chicago/Turabian StyleWanasinghe, Dhanushka N., and Peter E. Mortimer. 2022. "Taxonomic and Phylogenetic Insights into Novel Ascomycota from Forest Woody Litter" Biology 11, no. 6: 889. https://doi.org/10.3390/biology11060889
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.