
The SLI-SC Mathematical Model of African Swine Fever Transmission among Swine Farms: The Effect of Contaminated Human Vector

 ,
,  , , and
, , and
Abstract
:1. Introduction
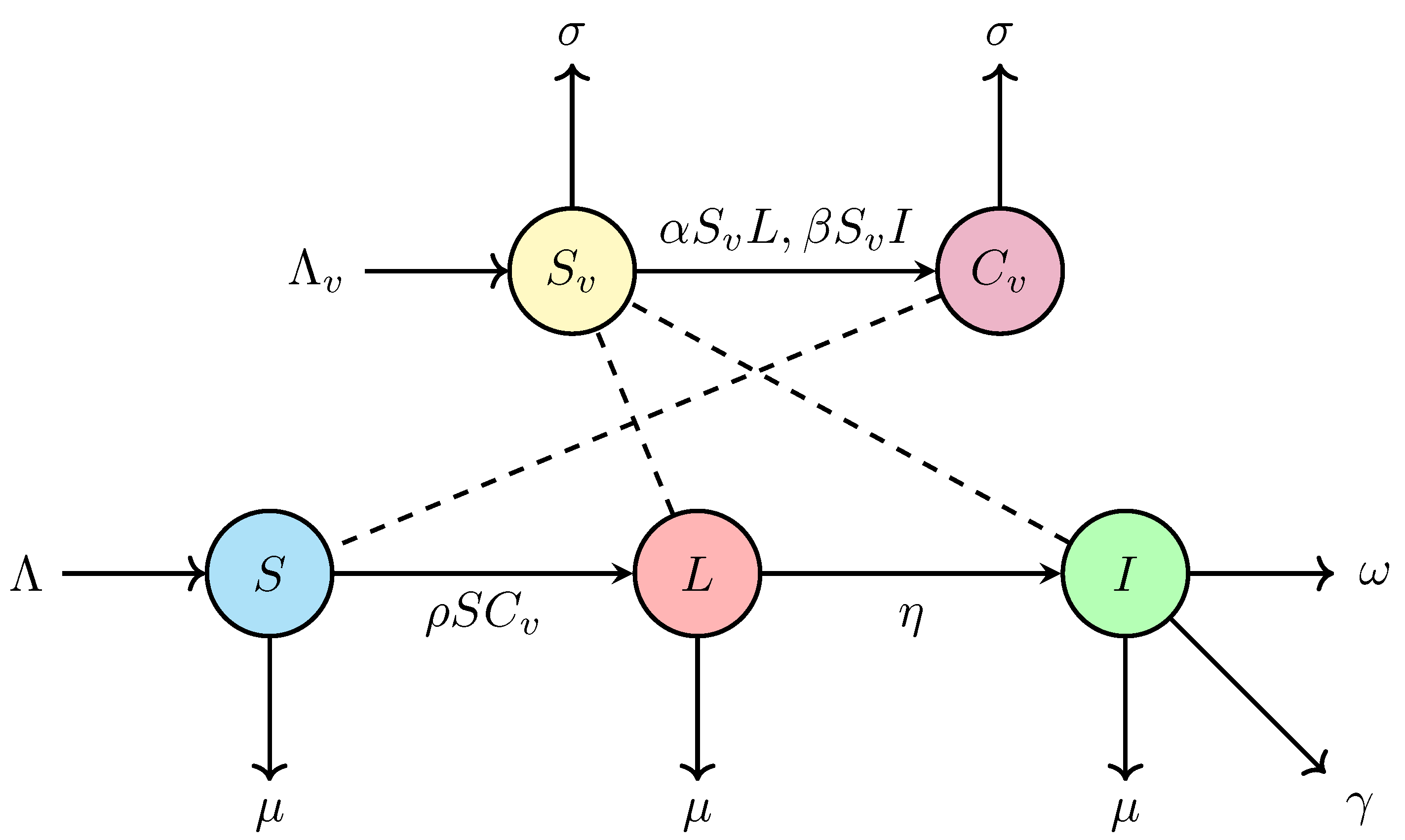
2. Mathematical Modeling
3. Analysis of the Model
3.1. Existence of the Solution
3.2. Positivity of the Solution
3.3. Invariant Region
3.4. Equilibria
- (i)
- The disease-free equilibrium (DFE)
- (ii)
- The endemic equilibriumwherewith , , , .
3.5. The Basic Reproduction Number ()
3.6. The Local Stability of Disease-Free Equilibrium
3.7. The Local Stability of Endemic Equilibrium
3.8. The Global Stability of Disease-Free Equilibrium
3.9. The Global Stability of Endemic Equilibrium
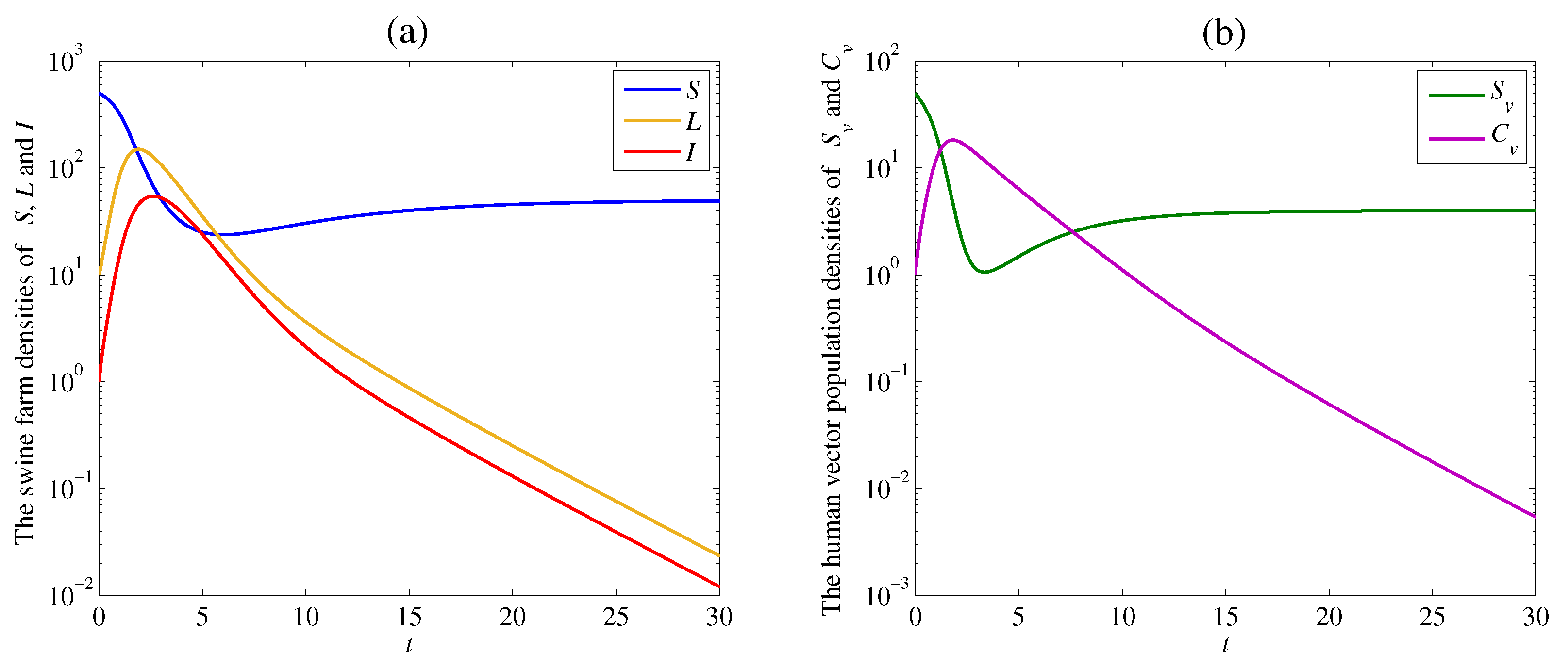
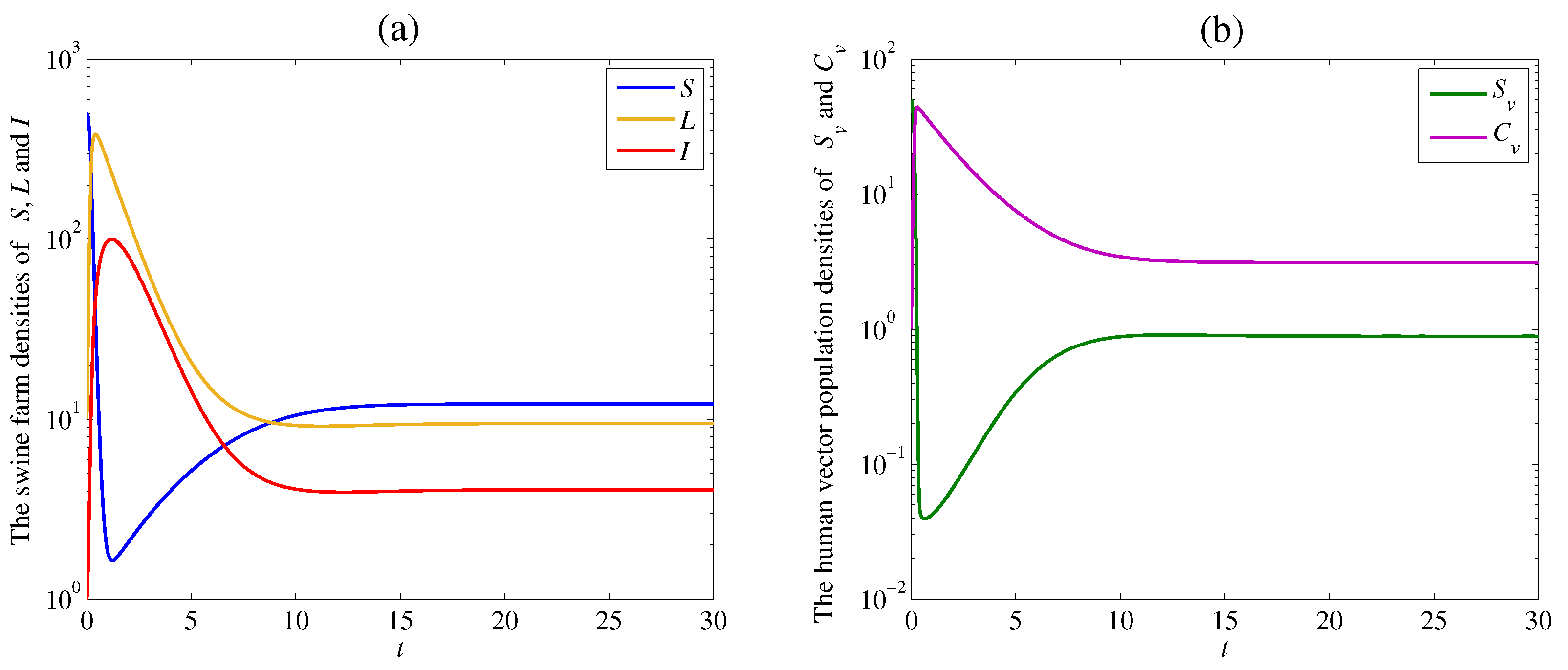
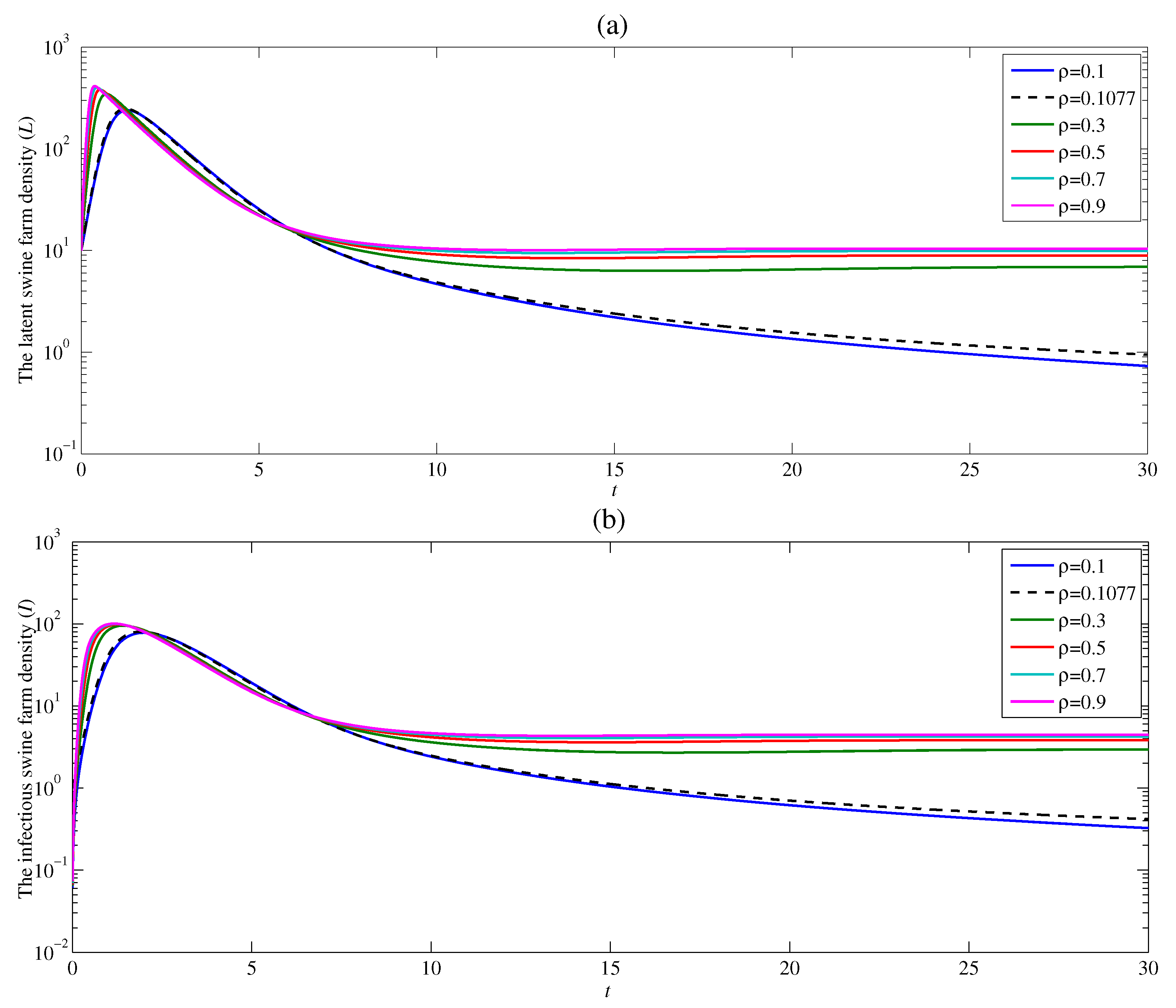
4. Numerical Examples and Discussion
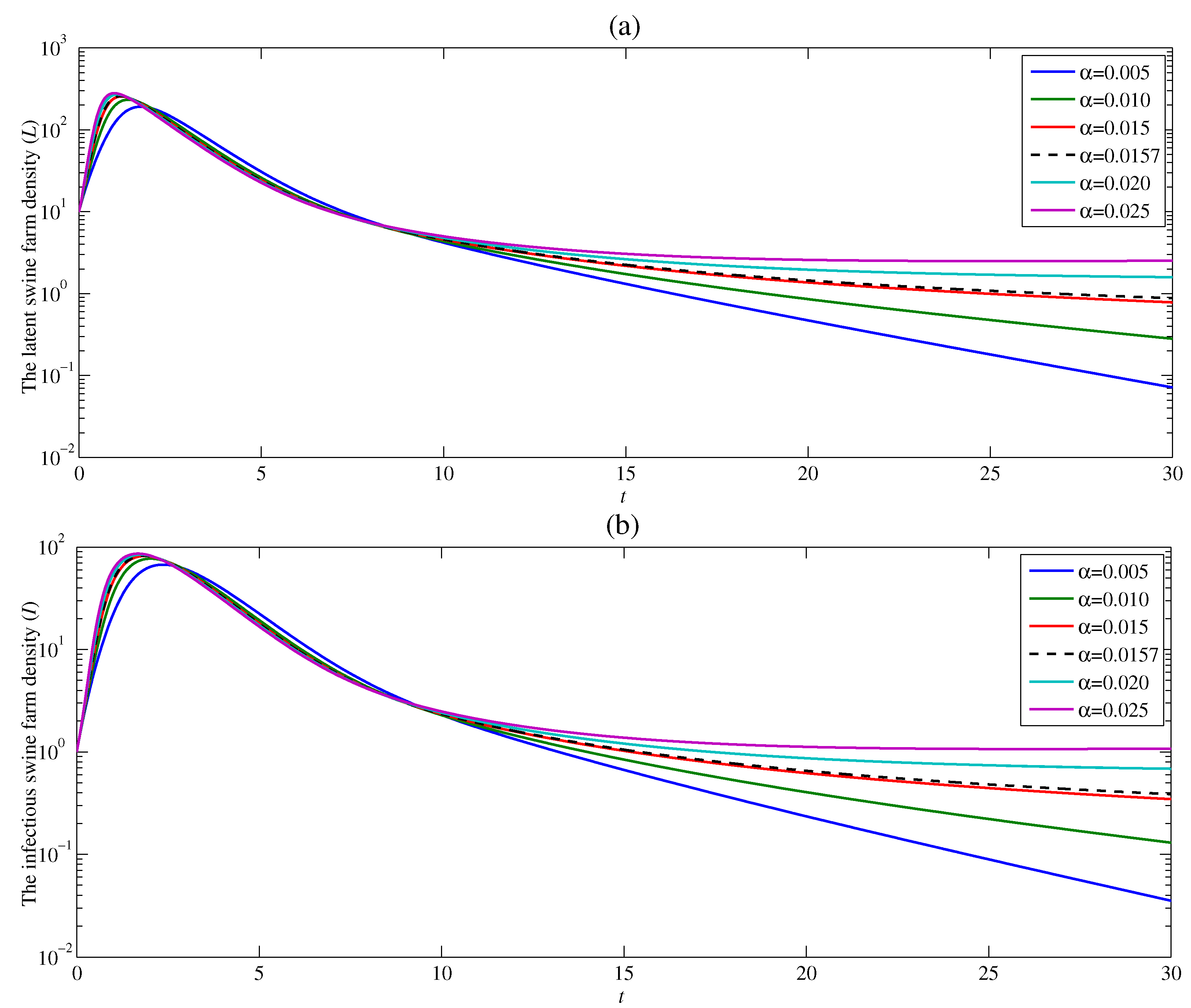
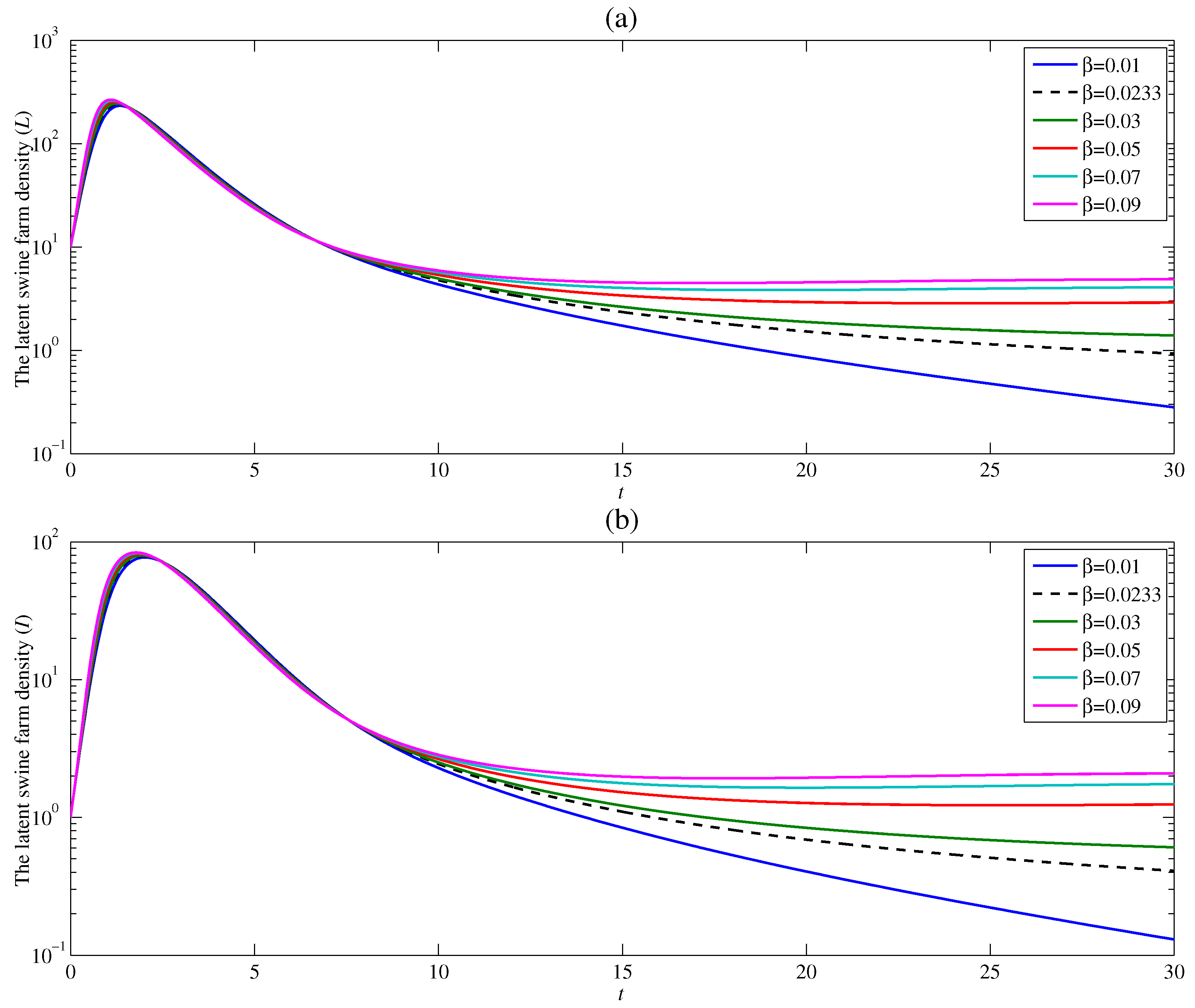
4.1. The Effect of the Contacting Human Vector Rate
4.2. The Effect of the Contacting Human Vector Rate
4.3. The Effect of the Contacting Human Vector Rate
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Nomenclature
| S | density of susceptible swine farm | farm per area unit |
| L | density of latent swine farm | farm per area unit |
| I | density of infectious swine farm | farm per area unit |
| density of susceptible human vector | vector unit per area unit | |
| density of contaminated human vector | vector unit per area unit | |
| recruited rate of swine farm | farm per area unit per unit of time | |
| recruited rate of human vector | vector unit per area unit per unit of time | |
| mortality rate of swine farm by nature | per unit of time | |
| mortality rate of swine farm by ASF | per unit of time | |
| mortality rate of swine farm | per unit of time | |
| by government control policy for ASF | ||
| mortality rate of human vector by nature | per unit of time | |
| transmission rate from L to I | per unit of time | |
| transmission rate from S to L through | area unit per vector unit per unit of time | |
| transmission rate from to through L | area unit per farm per unit of time | |
| transmission rate from to through I | area unit per farm per unit of time |
References
- Dixon, L.K.; Stahl, K.; Jori, F.; Vial, L.; Pfeiffer, D.U. African Swine Fever Epidemiology and Control. Annu. Rev. Anim. Biosci. 2020, 8, 221–246. [Google Scholar] [CrossRef] [Green Version]
- Blome, S.; Franzke, K.; Beer, M. African swine fever—A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Factsheet—African Swine Fever. Available online: https://www.aphis.usda.gov/publications/animal_health/asf.pdf (accessed on 30 April 2022).
- Bonnet, S.I.; Bouhsira, E.; De Regge, N.; Fite, J.; Etoré, F.; Garigliany, M.M.; Jori, F.; Lempereur, L.; Le Potier, M.F.; Quillery, E.; et al. Putative Role of Arthropod Vectors in African Swine Fever Virus Transmission in Relation to Their Bio-Ecological Properties. Viruses 2020, 12, 778. [Google Scholar] [CrossRef]
- Niederwerder, M.C. Risk and Mitigation of African Swine Fever Virus in Feed. Animals 2021, 11, 792. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. What Is African Swine Fever? Available online: https://www.oie.int/en/disease/african-swine-fever/ (accessed on 30 April 2022).
- Chenais, E.; Depner, K.; Guberti, V.; Dietze, K.; Viltrop, A.; Ståhl, K. Epidemiological considerations on African swine fever in Europe 2014–2018. Porc. Health Manag. 2019, 5, 6. [Google Scholar] [CrossRef]
- Wu, K.; Liu, J.; Wang, L.; Fan, S.; Li, Z.; Li, Y.; Yi, L.; Ding, H.; Zhao, M.; Chen, J. Current State of Global African Swine Fever Vaccine Development under the Prevalence and Transmission of ASF in China. Vaccines 2020, 8, 531. [Google Scholar] [CrossRef]
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Agriculture and Consumer Protection Department, Food and Agriculture Organization of the United Nations. ASF Situation in Asia & Pacific Update. Available online: https://www.swineweb.com/asf-situation-in-asia-pacific-update (accessed on 16 May 2022).
- Turlewicz-Podbielska, H.; Kuriga, A.; Niemyjski, R.; Tarasiuk, G.; Pomorska-Mól, M. African Swine Fever Virus as a Difficult Opponent in the Fight for a Vaccine—Current Data. Viruses 2021, 13, 1212. [Google Scholar] [CrossRef]
- Meloni, D.; Franzoni, G.; Oggiano, A. Cell Lines for the Development of African Swine Fever Virus Vaccine Candidates: An Update. Vaccines 2022, 10, 707. [Google Scholar] [CrossRef]
- Nigsch, A.; Costard, S.; Jones, B.A.; Pfeiffer, D.U.; Wieland, B. Stochastic spatio-temporal modelling of African swine fever spread in the European Union during the high risk period. Prev. Vet. Med. 2013, 108, 262–275. [Google Scholar] [CrossRef]
- Lee, H.S.; Thakur, K.K.; Pham-Thanh, L.; Dao, T.D.; Bui, A.N.; Bui, V.N.; Quang, H.N. A stochastic network-based model to simulate farm-level transmission of African swine fever virus in Vietnam. PLoS ONE 2021, 16, e0247770. [Google Scholar] [CrossRef] [PubMed]
- Ndondo, A.; Kasereka, S.; Bisuta, S.; Kyamakya, K.; Doungmo, E.; Ngoie, R.B. Analysis, modeling and optimal control of COVID-19 outbreak with three forms of infection in Democratic Republic of the Congo. Results Phys. 2021, 24, 104096. [Google Scholar] [CrossRef] [PubMed]
- Barongo, M.B.; Bishop, R.P.; Fèvre, E.M.; Knobel, D.L.; Ssematimba, A. A Mathematical Model that Simulates Control Options for African Swine Fever Virus (ASFV). PLoS ONE 2016, 11, e0158658. [Google Scholar] [CrossRef]
- Nielsen, J.P.; Larsen, T.S.; Halasa, T.; Christiansen, L.E. Estimation of the transmission dynamics of African swine fever virus within a swine house. Epidemiol. Infect. 2017, 145, 2787–2796. [Google Scholar] [CrossRef] [Green Version]
- Kouidere, A.; Balatif, O.; Rachik, M. Analysis and optimal control of a mathematical modeling of the spread of African swine fever virus with a case study of South Korea and cost-effectiveness. Chaos Solitons Fractals 2021, 146, 110867. [Google Scholar] [CrossRef]
- Halasa, T.; Boklund, A.; Bøtner, A.; Toft, N.; Thulke, H.H. Simulation of Spread of African Swine Fever, Including the Effects of Residues from Dead Animals. Front. Vet. Sci. 2016, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Mai, T.N.; Sekiguchi, S.; Huynh, T.M.L.; Cao, T.B.P.; Le, V.P.; Dong, V.H.; Vu, V.A.; Wiratsudakul, A. Dynamic Models of Within-Herd Transmission and Recommendation for Vaccination Coverage Requirement in the Case of African Swine Fever in Vietnam. Vet. Sci. 2022, 9, 292. [Google Scholar] [CrossRef]
- Weiss, H.H. The SIR model and the foundations of public health. Mater. Mat. 2013, 2013, 1–17. [Google Scholar]
- Chaiya, I.; Trachoo, K.; Nonlaopon, K.; Prathumwan, D. The Mathematical Model for Streptococcus suis Infection in Pig-Human Population with Humidity Effect. Comput. Mater. Contin. 2022, 71, 2981–2998. [Google Scholar] [CrossRef]
- Acemoglu, D.; Chernozhukov, V.; Werning, I.; Whinston, M.D. Optimal targeted lockdowns in a multigroup SIR model. Am. Econ. Rev. Insights 2021, 3, 487–502. [Google Scholar] [CrossRef]
- Prathumwan, D.; Trachoo, K.; Chaiya, I. Mathematical Modeling for Prediction Dynamics of the Coronavirus Disease 2019 (COVID-19) Pandemic, Quarantine Control Measures. Symmetry 2020, 12, 1404. [Google Scholar] [CrossRef]
- Ji, C.; Jiang, D. Threshold behaviour of a stochastic SIR model. Appl. Math. Model. 2014, 38, 5067–5079. [Google Scholar] [CrossRef]
- Derrick, N.; Grossman, S. Differential Equation with Application; Addision Wesley Publishing Company, Inc.: Reading, MA, USA, 1976. [Google Scholar]
- van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- Diekmann, O.; Heesterbeek, J.A.P.; Roberts, M.G. The construction of next-generation matrices for compartmental epidemic models. J. R. Soc. Interface 2010, 7, 873–885. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| 10 | |
| 0.2 | |
| 0.6 | |
| 0.8 | |
| 0.4 | |
| 2 | |
| 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuchard, P.; Prathumwan, D.; Trachoo, K.; Maiaugree, W.; Chaiya, I. The SLI-SC Mathematical Model of African Swine Fever Transmission among Swine Farms: The Effect of Contaminated Human Vector. Axioms 2022, 11, 329. https://doi.org/10.3390/axioms11070329
Chuchard P, Prathumwan D, Trachoo K, Maiaugree W, Chaiya I. The SLI-SC Mathematical Model of African Swine Fever Transmission among Swine Farms: The Effect of Contaminated Human Vector. Axioms. 2022; 11(7):329. https://doi.org/10.3390/axioms11070329
Chicago/Turabian StyleChuchard, Pearanat, Din Prathumwan, Kamonchat Trachoo, Wasan Maiaugree, and Inthira Chaiya. 2022. "The SLI-SC Mathematical Model of African Swine Fever Transmission among Swine Farms: The Effect of Contaminated Human Vector" Axioms 11, no. 7: 329. https://doi.org/10.3390/axioms11070329