
L-Tryptophan Mitigates Cannibalism and Improves Growth of Asian seabass, Lates calcarifer Reared in a RAS System
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Experimental Design
2.3. Experimental Feeds
2.4. Feeding Trial
2.5. Estimation of Growth Parameters and Cannibalism
2.6. Biochemical Analysis of Fish Samples
2.7. Statistical Analysis
3. Results
3.1. Growth Parameters
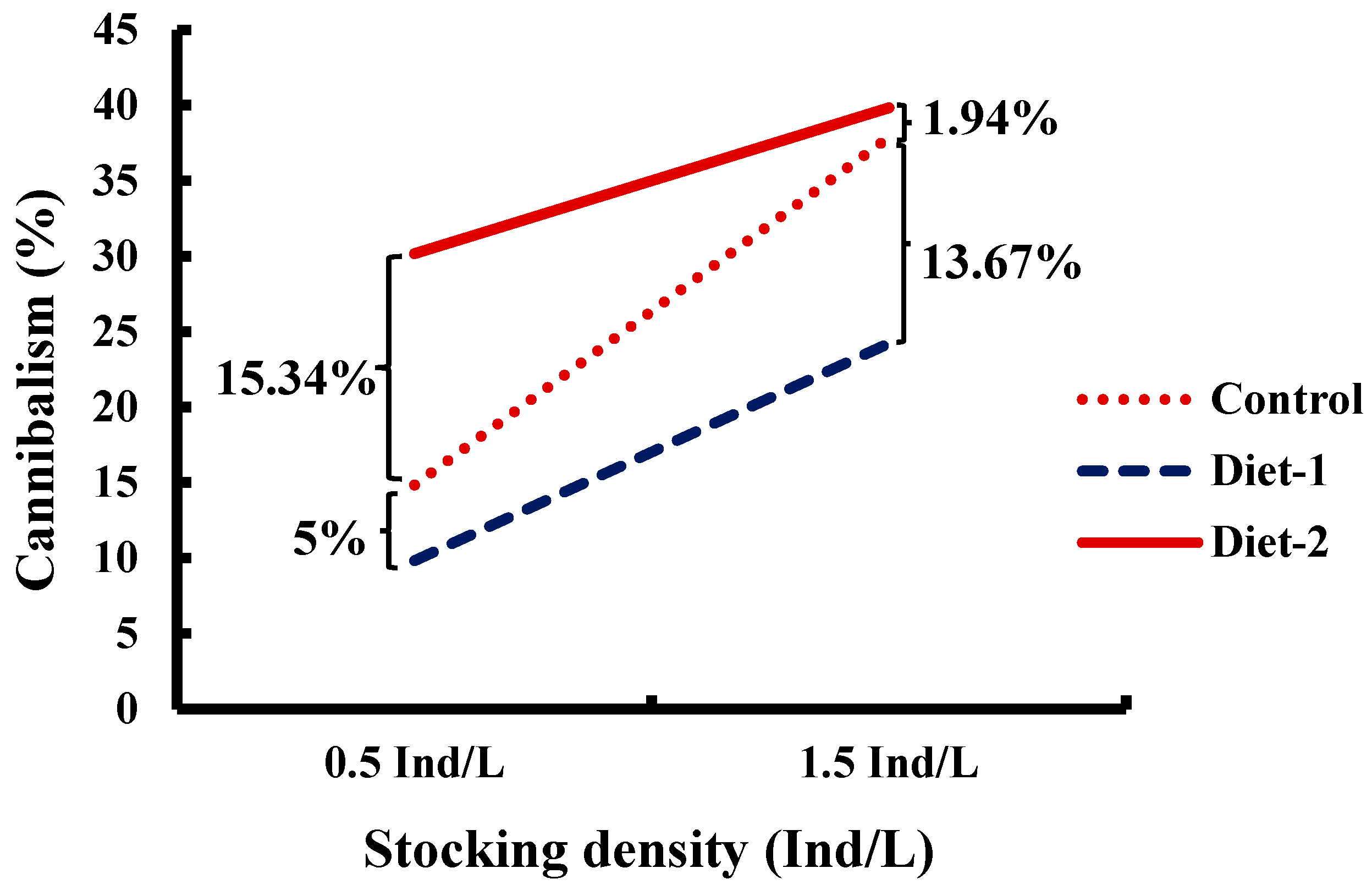
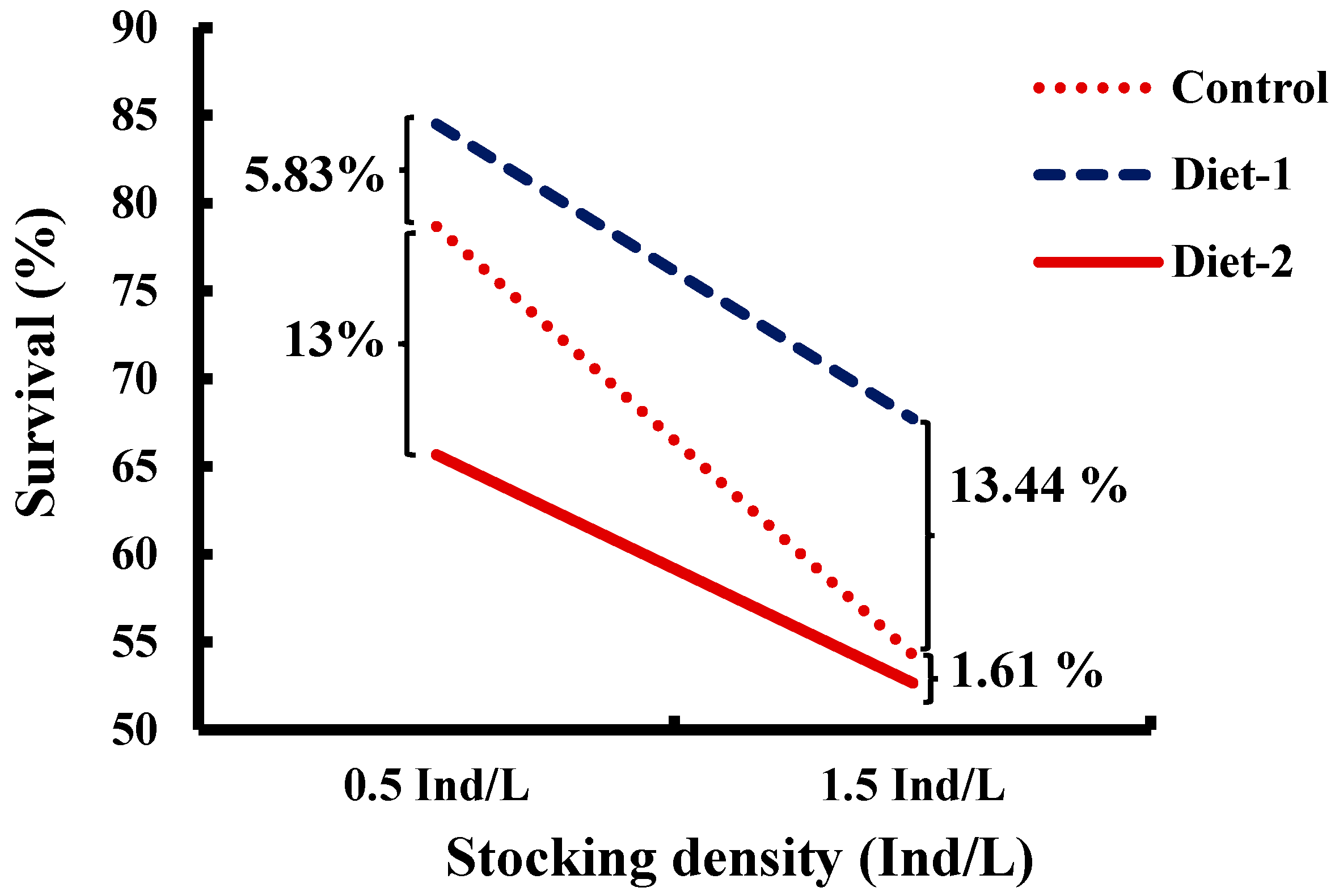
3.2. Cannibalism, Mortality, and Survival
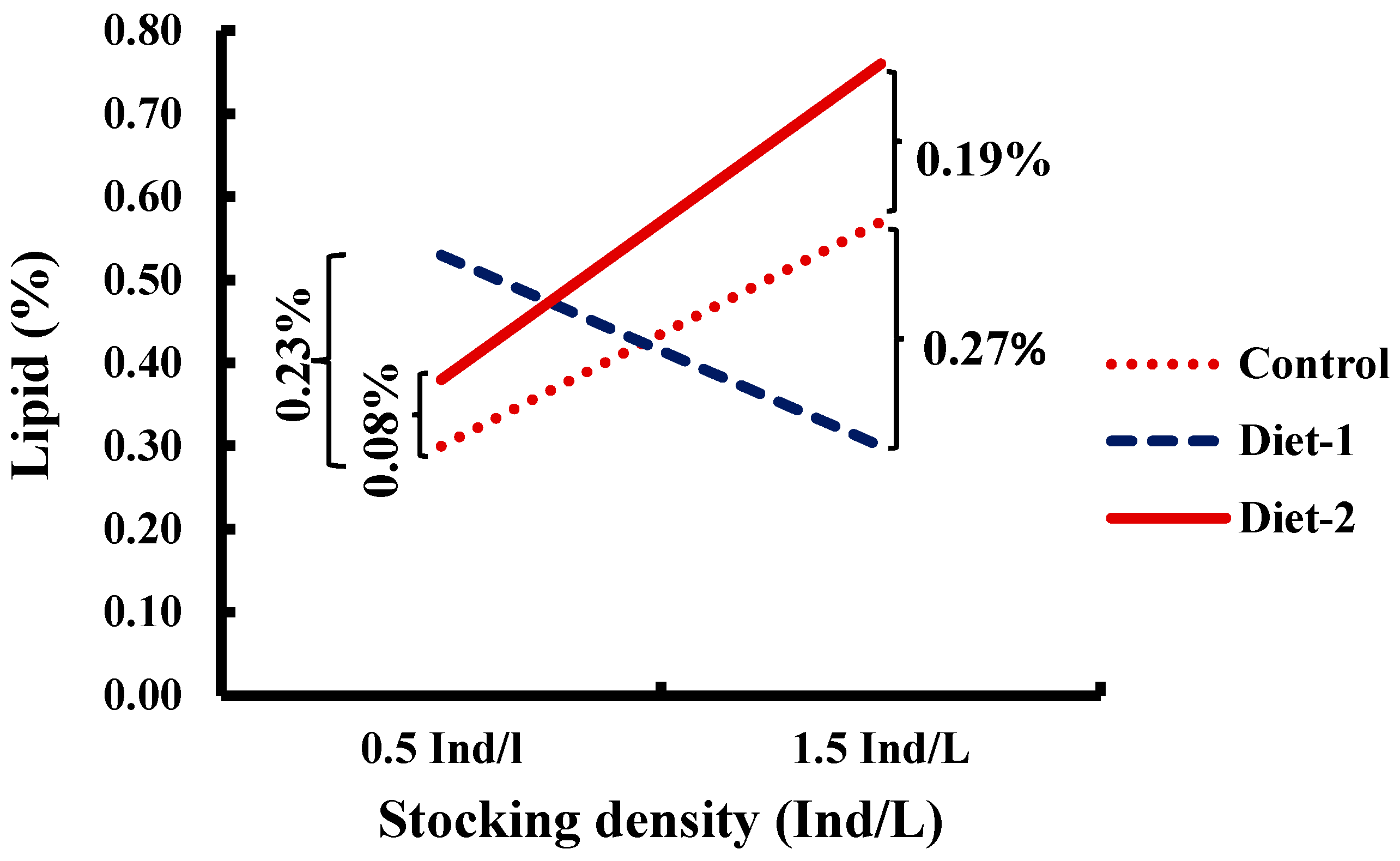
3.3. Biochemical Composition of Fish
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rimmer, M.A.; Russell, D.J. Aspects of the biology and culture of Lates calcarifer. In Tropical Mariculture; De Silava, S.S., Ed.; Academic Press: New York, NY, USA, 1998; pp. 449–476. [Google Scholar]
- de Jesus-Ayson, E.G.; Ayson, F.G. Reproductive biology of the Asian seabass, Lates calcarifer. In Biology and Culture of Asian Seabass Lates Calcarifer; CRC Press: Boca Raton, FL, USA, 2014; pp. 67–76. [Google Scholar]
- Greenwood, P.H. A review of the family centropomidae (Pisces: Perciformes). In Bulletin of the British museum (Natural History). Zoology; British Museum (Natural History): London, UK, 1976; Volume 29, pp. 1–81. [Google Scholar]
- FAO. Policy Measures for Managing Quality and Reducing Postharvest Losses in Fresh Produce Supply Chains in South Asian Countries. Rome. 2017. Available online: http://www.fao.org/3/i7954e/i7954e.pdf (accessed on 14 October 2022).
- Venkatachalam, S.; Kandasamy, K.; Krishnamoorthy, I.; Narayanasamy, R. Survival and growth of fish (Lates calcarifer) under integrated mangrove-aquaculture and open aquaculture systems. Aquac. Rep. 2018, 9, 18–24. [Google Scholar] [CrossRef]
- Arasu, A.R.T.; Kailasam, M.; Sundaray, J.K.; Subburai, R.; Thiagarajan, G.; Karaiyan, K. Improved Hatchery Technology for Asian Seabass Lates calcarifer (Bloch); Central Institute of Brackish Water Aquaculture, Indian Council of Agricultural Research: Tamil Nadu, India, 2008; Volume 34, pp. 1–38. [Google Scholar]
- Kestemont, P.; Jourdan, S.; Houbart, M.; Me’lard, C.; Paspatis, M.; Fontaine, P.; Cuvier, A.; Kentouri, M.; Baras, E. Size heterogeneity, cannibalism and competition in cultured predatory fish larvae: Biotic and abiotic influences. Aquaculture 2003, 227, 333–356. [Google Scholar] [CrossRef]
- Hecht, T.; Appelbaum, S. Observations on intraspecific aggression and coeval sibling cannibalism by larval and juvenile Clarias gariepinus (Clariidae: Pisces) under controlled conditions. J. Zool. 1988, 214, 21–44. [Google Scholar] [CrossRef]
- Hecht, T.; Pienaar, G.A. A review of cannibalism and its implications in fish larviculture. J. World Aquac. Soc. 1993, 24, 246–261. [Google Scholar] [CrossRef]
- Baras, E.; Jobling, M. Dynamics of intracohort cannibalism in cultured fish. Aquac. Res. 2002, 33, 461–479. [Google Scholar] [CrossRef]
- Baras, E. Cannibalism in fish larvae: What have we learned? In Larval Fish Aquaculture; Qin, J.G., Ed.; Nova Science Publishers: New York, NY, USA, 2013. [Google Scholar]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Bolasina, S.; Tagawa, M.; Yamashita, Y.; Tanaka, M. Effect of stocking density on growth, digestive enzyme activity and cortisol level in larvae and juveniles of Japanese flounder, Paralichthys olivaceus. Aquaculture 2006, 259, 432–443. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M.S.; Tort, L.; Robaina, L.; Vergara, J.M. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Winberg, S.; Nilsson, G.E. Roles of brain monoamine neurotransmitters in agonistic behaviour and stress reactions, with particular reference to fish. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1993, 106, 597–614. [Google Scholar] [CrossRef]
- Winberg, S.; Øverli, Ø.; Lepage, O. Suppression of aggression in rainbow trout (Oncorhynchus mykiss) by dietary L-tryptophan. J. Exp. Biol. 2001, 204, 3867–3876. [Google Scholar] [CrossRef]
- Mojjada, S.K.; Dash, B.; Pattnaik, P.; Anbarasu, M.; Joseph, I. Effect of stocking density on growth and survival of hatchery reared fry of Asian sea bass, Lates calcarifer (Bloch) under captive conditions. Ind. J. Fish. Sci. 2013, 60, 71–75. [Google Scholar]
- Manley, C.B.; Rakocinski, C.F.; Lee, P.G.; Blaylock, R.B. Stocking density effects on aggressive and cannibalistic behaviors in larval hatchery-reared spotted seatrout, Cynoscion nebulosus. Aquaculture 2014, 420–421, 89–94. [Google Scholar] [CrossRef]
- Rowland, S.J.; Mifsud, C.; Nixon, M.; Boyd, P. Effects of stocking density on the performance of the Australian freshwater silver perch (Bidyanus bidyanus) in cages. Aquaculture 2006, 253, 301–308. [Google Scholar] [CrossRef]
- Lefrançois, C.; Claireaux, G.; Mercier, C.; Aubin, J. Effect of density on the routine metabolic expenditure of farmed rainbow trout (Oncorhynchus mykiss). Aquaculture 2001, 195, 269–277. [Google Scholar] [CrossRef]
- Król, J.; Zakęś, Z. Effect of dietary L-tryptophan on cannibalism, survival and growth in pikeperch Sander lucioperca (L.) post-larvae. Aquac. Int. 2016, 24, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kailasam, M.; Sethi, S.N.; Sukumaran, K.; Biswas, G.; Subburaj, R.; Thiagarajan, G.; Ghoshal, T.K.; Vijayan, K.K. Effect of dietary L-tryptophan on cannibalism, growth and survival of Asian seabass, Lates calcarifer (Bloch, 1790) fry. Ind. J. Fish. Sci. 2017, 64, 28–32. [Google Scholar] [CrossRef]
- Höglund, E.; Øverli, Ø.; Winberg, S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef] [Green Version]
- Hseu, J.R.; Lu, F.I.; Su, H.M.; Wang, L.S.; Tsai, C.L.; Hwang, P.P. Effects of exogenous tryptophan on cannibalism, survival and growth in juvenile grouper, Epinephelus coioides. Aquaculture 2003, 218, 251–263. [Google Scholar] [CrossRef]
- Höglund, E.; Bakke, M.J.; Øverli, Ø.; Winberg, S.; Nilsson, G.E. Suppression of aggressive behaviour in juvenile Atlantic cod (Gadus morhua) by L-tryptophan supplementation. Aquaculture 2005, 249, 525–531. [Google Scholar] [CrossRef]
- Laranja, J.L.Q., Jr.; Quinitio, E.T.; Catacutan, M.R.; Coloso, R.M. Effects of dietary L-ryptophan on the agonistic behavior, growth and survival of juvenile mud crab Scylla serrata. Aquaculture 2010, 310, 84–90. [Google Scholar] [CrossRef]
- Harlioğlu, M.M.; Harlioglu, A.G.; Yonar, S.M.; Duran, T.C. Effects of dietary L-tryptophan on the agonistic behavior, growth, and survival of freshwater crayfish Astacus leptodactylus Eschscholtz. Aquac. Int. 2014, 22, 733–748. [Google Scholar] [CrossRef]
- Coloso, R.M.; Murillo-Gurrea, D.P.; Borlongan, I.G.; Catacutan, M.R. Tryptophan requirement of juvenile Asian sea bass Lates calcarifer. J. Appl. Ichthyol. 2004, 20, 43–47. [Google Scholar] [CrossRef]
- Johnston, W.L.; Atkinson, J.L.; Hilton, J.W.; Were, K.E. Effect of dietary tryptophan on plasma and brain tryptophan, brain serotonin, and brain 5-hydroxyindoleacetic acid in rainbow trout. J. Nutr. Biochem. 1990, 1, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Lepage, O.; Tottmar, O.; Winberg, S. Elevated dietary intake of L-tryptophan counteracts the stress induced elevation of plasma cortisol in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 2002, 205, 3679–3687. [Google Scholar] [CrossRef]
- de Pedro, N.; Pinillos, M.L.; Valenciano, A.I.; Alonso-Bedate, M.; Delgardo, M.J. Inhibitory effects of serotonin on feeding behavior in goldfish: Involvement of CRF. Peptides 1998, 19, 505–511. [Google Scholar] [CrossRef]
- Papoutsoglou, S.E.; Karakatsouli, N.; Chiras, G. Dietary L-tryptophan and tank colour effects on growth performance of rainbow trout (Oncorhynchus mykiss) juveniles reared in a recirculating water system. Aquac. Eng. 2005, 32, 277–284. [Google Scholar] [CrossRef]
- Lovell, T. Nutrition and Feeding of Fish, 2nd ed.; Springer Science+Business Media: New York, NY, USA; Kluwer Academic Publishers: Boston, MA, USA, 1998. [Google Scholar]
- Khan, M.S.K.; Salin, K.R.; Yakupitiyage, A.; Tsusaka, T.W.; Shrestha, S.; Siddique, M.A.M. Effect of stocking density and tank colour on nursery growth performance, cannibalism and survival of the Asian seabass Lates calcarifer (Bloch, 1790) in a recirculating aquaculture system. Aquac. Res. 2022, 53, 2472–2483. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association/American Water Works Association/Water Environment Federation: Denver, CO, USA, 2005; Volume 21, pp. 258–259. [Google Scholar]
- Khan, M.S.K.; Siddique, M.A.M.; Zamal, H. Replacement of fish meal by plant protein sources in Nile tilapia (Oreochromis niloticus) diet: Growth performance and utilization. Iran. J. Fish. Sci. 2013, 12, 864–872. [Google Scholar]
- Baras, E.; Kestemont, P.; Mélard, C. Effect of stocking density on the dynamics of cannibalism in sibling larvae of Perca fluviatilis under controlled conditions. Aquaculture 2003, 219, 241–255. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Cardinaletti, G.; Sigelaki, I.; Polzonetti-Magni, A. Comparative efficacy of clove oil and 2-phenoxyethanol as anesthetics in the aquaculture of European sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus aurata) at different temperatures. Aquaculture 2005, 246, 467–481. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2000; 238p. [Google Scholar]
- Siddique, M.A.M.; Aktar, M. Changes of Nutritional Value of Three Marine Dry Fishes (Johnius dussumieri, Harpodon nehereus and Lepturacanthus savala) during storage. Food Nutr. Sci. 2011, 2, 1082–1087. [Google Scholar] [CrossRef] [Green Version]
- Siddique, M.A.M.; Mojumder, P.; Zamal, H. Proximate composition of three commercially available marine dry fishes (Harpodon nehereus, Johnius dussumieri and Lepturacanthus savala). Am. J. Food Technol. 2012, 7, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Papoutsoglou, S.E.; Karakatsouli, N.; Koustas, P. Effects of dietary L-tryptophan and lighting conditions on growth performance of European seabass (Dicentrarchus labrax) juveniles reared in a recirculating water system. J. Appl. Ichthyol. 2005, 21, 520–524. [Google Scholar] [CrossRef]
- Pinchasov, Y.; Fancher, B.I.; Burke, W.H.; Jensen, L.S. Glycolic acid and tryptophan effects on feed intake and hypothalamic indolamines in chicks. Physiol. Behav. 1989, 45, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Young, S.N. Behavioral effects of dietary neurotransmitter precursor: Basic and clinical aspects. Neurosci. Biobehav. Rev. 1996, 20, 313–323. [Google Scholar] [CrossRef]
- Mélard, C.; Baras, E.; Mary, L.; Kestemont, P. Relationship between stocking density, growth, cannibalism and survival rate in intensively cultured larvae and juveniles of perch Perca fluvialitilis. Ann. Zool. Fennici 1996, 33, 643–651. [Google Scholar]
- Khan, M.S.K.; Salin, K.R.; Yakupitiyage, A.; Siddique, M.A.M. Effect of stocking densities on the growth performance, cannibalism and survival of Asian seabass Lates calcarifer (Bloch, 1790) fry in different nursery rearing system. Aquac. Res. 2021, 52, 5332–5339. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Mamcarz, A.; Kujawa, R.; Skrzypczak, A. Development of cannibalism in larval northern pike, Esox lucius (Esocidae). Ital. J. Zool. 1998, 65, 261–263. [Google Scholar] [CrossRef]
- Szkudlarek, M.; Zakęś, Z. Effect of stocking density on survival and growth performance of pikeperch, Sander lucioperca (L.), larvae under controlled conditions. Aquac. Int. 2007, 15, 67–81. [Google Scholar] [CrossRef]
- Solomon, R.J.; Udoji, F.C. Canibalism among cultured African catfishes (Heterobranchus longifillis and Clarias gariepinus). Nat. Sci. 2011, 9, 1–13. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Stocking Density | Doses of Dietary TRP (%) | ||
|---|---|---|---|
| Control (Diet) | Diet-1 (1.00%) | Diet-2 (1.50%) | |
| 0.5 ind./L | 0.41 * + 0 | 0.41 + 0.59 | 0.41 + 1.09 |
| 1.5 ind./L | 0.41 * + 0 | 0.41 + 0.59 | 0.41 + 1.09 |
| Parameter | Mean ± SE | Interaction Effects (Two-Way Factorial p-Value) | ||||
|---|---|---|---|---|---|---|
| Experimental Diet | Stocking Density | |||||
| Control | Diet-1 | Diet-2 | 0.5 ind./L | 1.5 ind./L | ||
| Temperature (°C) | 29.10 ± 0.00 | 29.10 ± 0.00 | 29.10 ± 0.00 | 29.10 ± 0.00 | 29.10 ± 0.00 | 0.300 |
| DO (mg/L) | 6.78 ± 0.06 b | 6.69 ± 0.06 a | 6.73 ± 0.05 ab | 6.86 ± 0.02 B | 6.61 ± 0.02 A | 0.120 |
| pH | 7.55 ± 0.00 | 7.55 ± 0.00 | 7.55 ± 0.00 | 7.55 ± 0.00 B | 7.54 ± 0.00 A | 0.397 |
| NH3 (mg/L) | 0.06 ± 0.00 | 0.06 ± 0.00 | 0.07 ± 0.00 | 0.06 ± 0.00 A | 0.07 ± 0.00 B | 0.178 |
| NO2 (mg/L) | 0.55 ± 0.03 | 0.56 ± 0.03 | 0.53 ± 0.03 | 0.49 ± 0.01 A | 0.60 ± 0.01 B | 0.451 |
| Dependent Variable | Experimental Diet | Stocking Density | Interaction Effects (Two-Way Factorial p-Value) | |||
|---|---|---|---|---|---|---|
| Control | Diet-1 | Diet-2 | 0.5 ind./L | 1.5 ind./L | ||
| Final body length (TLf, cm) | 10.43 ± 0.16 a | 10.68 ± 0.26 a | 11.64 ± 0.35 b | 10.48 ± 0.25 A | 11.35 ± 0.22 B | 0.708 |
| Final body weight (BWf, g) | 16.61 ± 1.96 a | 18.35 ± 2.67 a | 22.93 ± 2.67 b | 14.21 ± 1.10 A | 24.38 ± 1.28 B | 0.432 |
| Weight gain (WG, g) | 16.32 ± 1.96 a | 18.06 ± 2.67 a | 22.64 ± 2.67 b | 13.92 ± 1.10 A | 24.09 ± 1.28 B | 0.432 |
| Specific growth rate (SGR, % day−1) | 8.91 ± 0.27 a | 9.09 ± 0.34 a | 9.63 ± 0.27 b | 8.60 ± 0.26 A | 9.82 ± 0.11 B | 0.371 |
| Feed conversion ratio (FCR) | 0.94 ± 0.04 b | 0.81 ± 0.04 a | 0.87 ± 0.07 ab | 0.96 ± 0.03 B | 0.79 ± 0.03 A | 0.163 |
| Protein efficiency ratio (PER) | 2.55 ± 0.10 a | 2.98 ± 0.16 b | 2.82 ± 0.20 ab | 2.50 ± 0.07 A | 3.07 ± 0.12 B | 0.086 |
| Yield (g) | 4245 ± 1069 a | 5862 ± 1715 b | 5481 ± 1462 b | 2069 ± 99 A | 8322 ± 508 B | 0.003 |
| Cannibalism (%) | 26.36 ± 5.18 b | 17.03 ± 3.35 a | 35.00 ± 2.34 c | 18.28 ± 3.11 A | 33.98 ± 2.56 B | 0.001 |
| Mortality (%) | 7.17 ± 0.53 | 6.86 ± 0.76 | 5.83 ± 0.78 | 5.44 ± 0.50 A | 7.80 ± 0.32 B | 0.363 |
| Survival rate (SR, %) | 66.47 ± 5.52 b | 76.11 ± 3.90 c | 59.17 ± 3.01 a | 76.28 ± 2.88 B | 58.22 ± 2.49 A | 0.006 |
| CV for length (TLf, %) | 16.14 ± 3.84 | 21.51 ± 3.51 | 21.61 ± 1.93 | 14.14 ± 2.06 A | 25.37 ± 1.53 B | 0.397 |
| CV for weight (BWf, %) | 46.88 ± 13.23 a | 74.88 ± 7.44 b | 67.04 ± 7.67 b | 42.78 ± 6.74 A | 82.69 ± 3.02 B | 0.050 |
| Component (%) | Mean ± SE | Interaction Effects (Two-Way Factorial p-Value) | ||||
|---|---|---|---|---|---|---|
| Experimental Diet | Stocking Density | |||||
| Control | Diet-1 | Diet-2 | 0.5 ind./L | 1.5 ind./L | ||
| Moisture | 77.88 ± 0.25 | 77.82 ± 0.39 | 77.02 ± 0.26 | 77.36 ± 0.30 | 77.79 ± 0.23 | 0.164 |
| Ash | 1.28 ± 0.01 | 1.30 ± 0.03 | 1.33 ± 0.01 | 1.32 ± 0.02 | 1.29 ± 0.01 | 0.050 |
| Lipid | 0.44 ± 0.06 a | 0.41 ± 0.05 a | 0.57 ± 0.09 b | 0.40 ± 0.04 A | 0.54 ± 0.07 B | <0.001 |
| Protein | 18.13 ± 0.26 | 18.33 ± 0.29 | 18.59 ± 0.23 | 18.53 ± 0.26 | 18.17 ± 0.15 | 0.510 |
| Fiber | 0.05 ± 0.00 | 0.05 ± 0.00 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.00 | 0.294 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.S.K.; Salin, K.R.; Yakupitiyage, A.; Tsusaka, T.W.; Nguyen, L.T.; Siddique, M.A.M. L-Tryptophan Mitigates Cannibalism and Improves Growth of Asian seabass, Lates calcarifer Reared in a RAS System. Aquac. J. 2023, 3, 168-180. https://doi.org/10.3390/aquacj3030014
Khan MSK, Salin KR, Yakupitiyage A, Tsusaka TW, Nguyen LT, Siddique MAM. L-Tryptophan Mitigates Cannibalism and Improves Growth of Asian seabass, Lates calcarifer Reared in a RAS System. Aquaculture Journal. 2023; 3(3):168-180. https://doi.org/10.3390/aquacj3030014
Chicago/Turabian StyleKhan, Md. Shahzad Kuli, Krishna R. Salin, Amararatne Yakupitiyage, Takuji W. Tsusaka, Loc Thai Nguyen, and Mohammad Abdul Momin Siddique. 2023. "L-Tryptophan Mitigates Cannibalism and Improves Growth of Asian seabass, Lates calcarifer Reared in a RAS System" Aquaculture Journal 3, no. 3: 168-180. https://doi.org/10.3390/aquacj3030014