
Reviewing the Current Understanding of Replant Syndrome in Orchards from a Soil Microbiome Perspective

1
Department of Horticulture and Landscape Architecture, Colorado State University, Fort Collins, CO 80523, USA
2
Agricultural Research Service, United States Department of Agriculture, Fort Collins, CO 80526, USA
*
Author to whom correspondence should be addressed.
Appl. Microbiol. 2023, 3(3), 856-866; https://doi.org/10.3390/applmicrobiol3030059
Submission received: 11 July 2023
/
Revised: 31 July 2023
/
Accepted: 1 August 2023
/
Published: 4 August 2023
(This article belongs to the Special Issue Microbiome in Ecosystem 2.0)
{kind=link}
{kind=link}
Abstract
:Replant syndrome (RS) of fruit and nut trees causes reduced tree vigor and crop productivity in orchard systems due to repeated plantings of closely related tree species. Although RS etiology has not been clearly defined, the causal agents are thought to be a complex of soil microorganisms combined with abiotic factors and susceptible tree genetics. Different soil disinfection techniques alleviate RS symptoms by reducing the loads of the deleterious microbiome; however, the positive effect on crop growth is temporary. The goals of this paper are: (1) to conceptualize the establishment of the syndrome from a microbiome perspective and (2) to propose sustainable solutions to develop a beneficial microbiome to inhibit the onset of RS.
1. Introduction
The agronomic challenges that arise during the re-establishment of a closely related tree fruit/nut species are collectively known as replant syndrome (RS). RS symptoms can be observed in the first replant generation (second orchard generation) and persist for several years or even decades [1,2]. RS has been vaguely characterized by reduced tree growth, lifespan, fruit yield, and fruit quality attributes such as soluble sugar level and sugar–acid ratio [2,3,4,5]. More descriptive characterizations have included reduced branching, shortened internodes, deformed leaves, root necrosis/discoloration, and reduced root growth [4,5,6,7]. However, even these more descriptive symptoms are not diagnostic [8] and often trees grown in replant soils need to be compared to trees grown in previously non-orchard soils in order to fully grasp the detrimental consequences of replant syndrome. Furthermore, most reports of RS longevity in the soil are merely anecdotal, since controlled research paired with accurate and detailed, multi-decade cropping histories has been practically impossible to obtain [9,10].
In addition to ambiguous symptom descriptions, there is no consensus on terminology as synonyms for RS include soil sickness, soil fatigue, replant problem, replant disease [11], soil exhaustion, replant disorder [12], and specific replant disease [13]. Here, the term replant syndrome is used since the condition’s onset is driven by repeated monocropping [14,15]. Additionally, while a “disease” has distinguishing symptoms typically derived from a single known cause (i.e., a specific pathogen), a “syndrome” refers to a group of signs, phenomena, or symptoms that occur together [16] with an uncertain underlying primary cause.
There is no unanimous agreement on the etiology of RS, which has largely remained an enigma for over 300 years [7,17]. The mechanisms by which consecutive monocultures give rise to a decline in crop productivity are still subject to debate. While RS has been reported in many crops, its negative impacts have been more notable among fruit/nut trees in the Rosaceae family such as almond [18], apple [12], cherry [19], pear [19], and peach [6,11]. Citrus species (Rutaceae family) are greatly affected by RS [20,21]. Multiple factors such as autotoxin production leading to accumulation, an imbalance of soil nutrients, and an imbalance of the microbial community structure have been credited with exacerbating RS [20]. While abiotic factors such as autotoxins and nutrient imbalance decrease soil fertility [22], they are not necessarily the direct cause of RS, but rather, may increase the survival and competitiveness of phytopathogens [23]. A critique of the idea that autotoxicity relates to RS is that fallow periods of up to three years fail to suppress RS and improve tree growth [24]. While chemicals causing autotoxicity are unlikely to be stable enough to persist for years, they may result in longer-term shifts in the soil microbiome [23]. Thus, instead of one specific phytopathogen, the primary cause of RS is suspected to consist of a complex of soil phytopathogens which have been shown to be enriched by autotoxins [10,25]. For example, Panax notoginseng was found to produce autotoxic ginsenosides which enriched potential phytopathogens (Alternaria, Cylindrocarpon, Fusarium, Gibberella, and Phoma); meanwhile, relative abundances of beneficial taxa (Acremonium, Mucor, and Ochroconis) decreased [25]. As the microbiome shifts, plant-growth-promoting microbes could become outnumbered by phytopathogens.
Microbiome shifts as the possible underlying cause of RS is further supported by studies showing that plants grown in autoclaved RS soil experience a remarkable increase in growth relative to plants grown in untreated replant soils [11,26,27]. Similarly, fruit tree biomass has been shown to increase in RS soils treated with chemical fumigation, resulting in reduced microbial biomass carbon with no apparent effect on other soil properties (basal respiration, ergosterol content, pH, electrical conductivity, and most nutrient and metal contents) [22]. Finally, when apple trees exhibiting RS symptoms were transplanted from RS soils into healthy soils the RS symptoms reversed [23].
This reversibility is particularly interesting given the identification/involvement of several potential soilborne plant pathogens in RS. For example, phytopathogens frequently associated with RS are oomycetes Pythium and Phytophthora, bacterial taxa from actinomycetes and genera of Bacillus and Pseudomonas, and the root lesion nematode [5,22]. Fungal suspects are Cylindrocarpon, Rhizoctonia, Fusarium sp., Alternaria sp., Myrothecium verrucaria, and Mycelia sterilia with many of these taxa being frequently isolated from the rhizosphere (soil surrounding plant roots) [6,23]. However, microbe–microbe and microbe–plant interactions are complex, and site-to-site variation has yielded contradicting results. For instance, Phytopythium vexans was found to be a pathogen in one site but acted as a biological control at a different location [6]. Virulence differences of P. vexans strains compounded with different abiotic or biotic soil factors could explain these discrepancies [6].
In summary, the diversity and abundance of phytopathogens cause RS, with abiotic factors and autotoxicity instigated by the previous monocrop acting as positive feedback mechanisms for phytopathogen recruitment and development. RS etiology appears to depend not only on the presence of phytopathogens but also on the overall balance of the soil microbial community.
2. Developing a Soil Microbiome Model to Understand Replant Syndrome
Tree growth in an orchard’s second generation (replanting) is not as vigorous as its first generation. Often, young, transplanted trees die in sites exhibiting severe RS [7]. Additionally, peach seedlings display reduced height and trunk width compared to a control group grown in fumigated soil in as little as 10 weeks [6]. Some studies have found that shoot growth was reduced by 66.9–71% with shoot masses staying consistently low after multiple replanting generations [2]. Less severe cases have noted that trees can overcome an initial delay in growth, eventually reaching the size and annual yields of those grown in healthy soils [28,29]. Nonetheless, recovery is time-consuming, taking valuable years and resources, which ultimately reduces the profitability of the orchard [4]. In these less-severe sites, the fruiting of trees can be delayed 2 to 3 years and still never attain comparable yields to those of the first cycle of planting [7]. Even in instances where RS causes a reduction in fruit yield or a shortened production life without ending in plant death, the resulting reduction in profits has been estimated at 10–20% [6].
RS can persist in fallowed soil for several years or even decades following the removal of the first established orchard [12,24]. It is believed that RS symptoms can be observed even if the roots of previous plants were in an area for only a few months. When young saplings are transplanted, the young root systems interact with populations of phytopathogens from the plant matter residue of the previous trees. There are examples of literature indicating younger plants are more susceptible to diseases compared to their adult counterparts [27]. As such, peach saplings are known to struggle in RS soil.
Traditionally, it is believed that RS primarily affects the next cropping cycle if the consecutive plant species are closely related (i.e., peaches following peaches). Specific RS, like specific apple replant disease, is a buildup of non-generalist pathogens tailored for the genotype of the host tree with host plant residues playing a key role [8]. This would support the possibility that with a decrease in the number of tree hosts, there would be a decrease in the specific replant microbes. Nonetheless, even with the removal of tree hosts, specific phytopathogens can be sequestered in plant residues until complete decomposition. A non-competing concept is that the pathogen build-ups are often composed of ubiquitous generalists [30]. For example, a build-up of phytopathogenic nematodes has been found to be partially responsible for the nonspecific replant symptoms [8]. Once the orchard is newly planted, the RS microbiome will exponentially colonize these recently introduced tree hosts.
Orchard management practices use natural tree physiology to dictate the processes to which the peach tree should direct its energy [31]. Traditional horticultural practices in orchards do not focus on encouraging the tree host to expend its energy in recruiting beneficial microbes for the sake of immune defense. However, recent studies have investigated sustainable techniques like intercropping and how different cover crops influence soil microbial communities in apple orchards [32]. Plants have been found to use between 5–25% of all photosynthetic net fixation of CO2 for root exudation of carboxylates [33], which are critical for attracting plant-growth-promoting rhizobacteria to the plant [34]. Consequently, there is a possibility that the RS microbiome develops instead of a beneficial microbiome tailored to the peach plant, negatively impacting the peach orchard even within the first generation. The disease may not be readily apparent, and the damage observed may be misidentified as part of the aging process. It is generally agreed that the process of RS is initiated by repeated monoculture, and here it is highlighted that RS begins to establish, even in the first generation, if the fruit trees are relatively asymptomatic [35]. Incidentally, significant shifts in the soil microbial community have been detected between non-cultivated, first-year, and second-year apple trees of the first planting [35]. In support of our hypothesis, when second-generation apple trees were planted in steamed disinfected soils where first-generation apple trees had been grown for only three years, the increase in growth was equal to that achieved in non-cultivated soils [35]. In short, the precursor phytopathogenic replant microbes existed in the soil before the orchard was established and, with time, the environment began to evolve virulent traits that were increasingly effective, building an inhospitable environment for the next planting of fruit trees.
Agricultural practices such as pruning initiate a stress response, which stimulates growth to replace the lost biomass [36]. As a result, exposed tissue can become infected [37]. It is known that common pathogens, like the Cytospora leucostoma, have great difficulty colonizing trees except through open wounds induced by injuries such as drought injury, winter injury, or pruning [38]. These wounds allow repeated recolonization/co-colonization of multiple strains of phytopathogens which should increase their virulence as observed in other pathogens [39]. Here, it is posited that the replant microbiome virulence levels build up gradually over time, and microbial populations approach higher levels as the first-generation plants are maturing for the first cycle of growth. The chronological age of a plant has been correlated with increased pathogen resistance [40]. Although immune signaling can increase from early developmental stages to reproductive stages, the fitness of a plant’s immune system decreases during the reproductive stage as a possible function of host senescence [40]. These findings need to be correlated for fruit trees with a longer lifespan.
Here, the proposed model (Figure 1) is based on broad patterns in an attempt to link tree development through time and the RS microbiome build-up to monitor the development of RS. The purpose of this model is to represent a hypothetical replant situation. Peach, Prunus persica, was selected as the example. The first generation of an orchard is defined as an area where peaches have not been grown previously. The timeline starts with the trees planted from seedlings or transplanted saplings (Figure 1a). First-generation orchards do not exhibit replant symptoms [41], since neither allelochemicals nor the replant microbiome are present in the soil in detrimental concentrations [6,42]. Typically, peach trees take 1–3 years to be established in the soil and have the potential to provide a commercial crop during the second year [32]. As the tree roots are established, the tree canopy is trimmed and trained to bear larger branches that can hold a heavy load of fruit [43]. This is a large energy expenditure since the more trimming, the more vigorous epicormic growth occurs [44]. Peak fruit set starts at 4 years of age for what is considered a mature tree [45,46], and peaks at eight years with yields being around 50–150 pounds of fruit per year [47]. After year 8, the fruit set decreases, with year 12 possibly having minimal fruit sets. In orchards, dwarfing rootstocks are used to reduce vegetative vigor by controlling root growth, which in turn can divert sugars to fruit production, especially in young trees [32]. Peach dwarfing rootstocks typically live about 10–15 years in an intensive orchard setting [47].
Microbial communities in the bulk soil are extremely diverse, with estimates of 10 billion bacteria classified under thousands of different species in just 1 g of soil [48]. Plants secrete root exudates to culture beneficial microbes in the rhizosphere that are tailored to the plant’s needs [49]. However, in addition to symbionts, phytopathogens can also be attracted to this chemical communication (Figure 1b) [50]. Thus, the precursor microbes that make up the RS microbiome are most likely already present in the bulk soil and can proliferate as the composition of the bulk soil shifts [51].
3. Breaking the Cycle of Replanting Syndrome
Several solutions have been proposed to combat the problem of RS, each with varying degrees of success. These solutions may be viewed in two ways: (1) a single application of a pre-plant soil disinfection strategy and (2) a continuously implemented biological strategy that increases either plant or microbial diversity. Soil disinfection methods include chemical fumigation, solarization, anaerobic soil disinfestation, autoclaving, soil amendments (Brassica napus seed meal, biochar), or even soil replacement in severe cases. Biological strategies are polyculture (cover crops or intercropping), rootstocks, or plant-growth-promoting inoculations and use concepts drawn from the intermediate disturbance hypothesis (IDH). These strategies are designed to avoid the shift towards an RS microbiome.
Pre-plant soil disinfection strategies typically yield more consistent successes, even if temporary, while a biological strategy that increases diversity often varies in success. Sterilization is defined as a process that effectively eradicates all viable microorganisms (including bacterial spores) from a surface or product [52]. Since sterilization of bulk soil is incredibly challenging, the term “soil disinfection” is used here in place of “soil sterilization” to convey a process that reduces the microbial load of a surface [53]. Although other soil microbe eradication techniques such as microwaving and gamma radiation exist, methods such as soil replacement, chemical fumigation, and solarization are the most common practices implemented in orchards for soil disinfection. Pre-plant fumigation has the remarkable ability to reduce RS; however, its benefits are temporary, and it is primarily a pre-plant method. Although chloropicrin has shown effectiveness in reducing RS that was not nematode related, the fumigant was deemed “unpleasant to handle” [54]. Preliminary field and greenhouse trials testing Vorlex have shown promising results and could be an alternative fumigant to chloropicrin for ameliorating RS soils [54]. Currently, chloropicrin is heavily restricted, and Vorlex’s registration has been cancelled since 1992. Methyl bromide is a chemical fumigant that was used for RS until 2005 but has since been phased out by U.S. and European governments, since it was found to deplete the ozone layer [55]. Other chemical fumigants, such as Methyl iodide, have been shown to be as effective against RS as methyl bromide [55]. Although methyl iodide does not deplete the ozone layer and was approved by the Environmental Protection Agency in 2008, by 2011 the Pesticide Action Network of North America characterized the fumigant as a neurotoxin and carcinogen [56]. This led Arysta LifeScience to withdraw methyl iodide from the United States and other markets [56]. Chemical fumigants are becoming more restricted since they are considered non-sustainable methods for soil remediation [57]. Solarization, the technique of trapping the sun’s radiation in the soil using tarps, has reduced soil fungal phytopathogens such as Fusaruim spp., Verticillium spp., and Ilyonectria mors-panacis (responsible for RS in ginseng) [58]. Anaerobic soil disinfestation builds upon solarization through the addition of carbon substrates and water to the soil, which increases soil temperature and slows down gas exchange [59]. Anaerobic soil disinfestation has demonstrated potential for reducing soil microbial loads (fungi, oomycetes, bacteria, and nematodes) in different soil types and is comparable to soil fumigation [59]. Anaerobic soil disinfestation has been shown to increase trunk cross-sectional area in almond trees by 148–214% compared to controls [59]. Autoclaving the soil as a pre-planting method has increased peach tree biomass [25]. Gamma radiation appears to be the most effective method for soil sterilization [60], but this method is impractical at an orchard scale. In an attempt to reduce RS-related microbes, the complete removal of the RS soils and replacement with healthy/non-pathogenic soil has been in practice [61]. Nonetheless, an inoculation of merely 1% of RS soil is sufficient for the associated microbes to re-establish and reduce tree growth [62]. Soil amendments with Brassica napus seed meal were effective starting in the third year of application [24]. Additionally, soil amendments of pinewood biochar (10–20% (v/v)) led to an increase in total peach biomass compared to the untreated control [41]. Although effective in reducing RS, these pre-plant soil disinfection strategies are a temporary solution, which provide some relief from RS symptoms.
In terms of the microbiome, how RS develops could follow the intermediate disturbance hypothesis (IDH), which posits that local species diversity is optimized when environmental disturbances are not drastic in terms of magnitude and occur at a regular interval [63]. Although both “magnitude” and “regular interval” are ambiguous [64], the management practices of an orchard—such as irrigation, fertilizer, and pesticides—might provide an ideal environment for pathogens and microbial competitors to enhance their virulence and colonization of the rhizosphere. Since microbes can quickly undergo multiple generations, they can evolve in a relatively short time span. If placed in an ideal setting, bacteria can, therefore, evolve resistance to antibacterial within 10 days [65]. The bulk orchard soil of an orchard experiences much less disturbance than annual crops, so 12 years should be sufficient time for the convergent evolution of several microbes to develop virulent functionalities towards their host.
Biological strategies (cover crops and rootstocks) used to remedy RS are continuously implemented, with success being site-dependent unlike pre-plant soil disinfection strategies [66,67,68]. These strategies aim to increase diversity in the field by using genetically distinct rootstocks and cover crops, which in turn can increase microbial diversity [40]. Sustainable practices such as increasing plant diversity through polyculture, crop rotation, intercropping, and cover crops have been shown to improve soil health unlike monoculture. Cover crops can improve soil health by increasing nitrogen levels (legumes) or increasing antimicrobial glucosinolates (Brassica). Furthermore, the planting of multiple genetically distinct species from the previous crop in polyculture can dilute the build-up of autotoxic compounds by contributing a mix of different plant residues [69]. Although one year of using wheat as a cover crop gave rise to enhanced vegetative growth and apple tree yield, it was not as effective as methyl bromide [24]. A cover crop of wheat showed promising results, but to further reduce RS there needs to be an antimicrobial aspect as well. Incorporation of cover crops which are resistant to generalists phytopathogens, such as nematode-resistant cowpea Vigna unguiculata (L.), have been shown to increase tomato yields more than the growth and incorporation of susceptible cowpea or non-incorporation of cowpea [70]. Other promising cover crops that may be used to manage generalist phytopathogens such as plant-parasitic nematodes are Crotalaria spp. and Tagetes spp. [70,71,72].
The development of genetic tools such as rootstocks have shown potential. Peach rootstocks with resistance to root-knot nematodes have been developed [73,74]. Furthermore, peach rootstocks—such as Evrica, PAC 9801-02, ROOTPAC® 40, and Tetra—appear to be tolerant to replant soils [75]. Additionally, the drawbacks of monoculture can be mitigated by using rootstocks that are genetically different from their scions which could be used to promote plant diversity while maintaining the same fruit crop type in the orchard [76]. However, RS-resistant rootstocks need to be able to tolerate regional abiotic conditions such as climate, soil type, pH, salinity, etc. [77].
Beneficial microbe inoculums with antimicrobial properties have been developed to enhance crop productivity [78], but these are still in development for RS. Generalized conclusions have surmised that more than 60% of the strains isolated from healthy soils corresponded to Pseudomonas sp. [79]. More specifically, Pseudomonas putida has been found to isolate suppressed replant-contributing phytopathogens—such as the growth of Rhizoctonia and Pythium spp. in vitro—and could control Rhizoctonia root rot for apple trees [12]. Arbuscular mycorrhiza fungi (AMF) form symbiosis with the roots of approximately 80% of studied land plants [80]. Arbuscular mycorrhiza has been tested by using inoculations of Acauloapora scrobiculata in replant soils, resulting in significantly increased shoot biomass and root phosphorus, potassium, calcium, copper, zinc, iron, and boron concentrations [81].
The effect of soil disinfection is effective but temporary and requires a complimentary technique. Figure 2 conceptualizes how the phytopathogen load of peach orchard (orange) soils gradually increase once a tree of the same genotype is re-planted. However, it is possible that, even in untreated soils, the phytopathogen concentrations could plateau. The microbial composition of the rhizosphere, in terms of both bacterial and fungal communities, has been found to be highly variable and to change over seasons and years [4], which could indicate that the players that cause RS shift even within the same site.
The rhizosphere may not be space-limited, but rather nutrient-limited, since direct observations of roots have shown the majority of the root surface is open space and remains uncolonized [82]. This would mean that the rhizosphere has a carrying capacity, and the total abundance of rhizosphere microorganisms may be consistent with changes occurring in the composition of the rhizosphere [82]. As soil disinfection lowers the phytopathogen population in replant soils of a peach orchard (blue), there is temporary relief from RS. This population could eventually recover, and disinfected replant soils may require continuous measures to increase microbial diversity, such as those highlighted previously.
RS is a multifaceted issue, thus requiring a multifaceted solution. For example, combinations of cover cropping and Brassica napus seed meal soil amendment improved the initial peach growth equivalent to a fumigation treatment using 1,3-dichloropropene-chloropicrin [24]. However, using an autoclave as the pre-plant soil disinfection method prior to having the cover crops established, the soil was not amended in a way that was conducive to inducing a biomass increase in the following peach tree planting as compared to the non-autoclaved with no cover crop controls. Additionally, in the same study, not all cover crops induced peach growth equally [26]. Soil disinfection can be challenging to incorporate in multifaceted approaches, since this strategy can decrease not only phytopathogens but beneficial bacteria like nitrogen-fixing Rhizobium [83]. A common goal of soil disinfection is to reduce all microbial life, which can be accomplished by heating moist soil to 63 °C for 30 min as it is known to eliminate most pathogenic fungi, bacteria, and viruses [84]. However, solarization practices which induced soil temperatures that did not exceed 41 °C at depths of 30–46 cm still greatly reduced soil population densities of fungal phytopathogens such as Verticillium dahlia Kleb., Pythium ultimum Trow., Rhizcotonia solani Kuehn, and Thielaviopsis basicola [85]. Similar solarization studies also found that lethal temperatures for thermal sensitive phytopathogens have been reported to be less than 41 °C (ED90 of Verticillium dahlia after 14 h at 37 °C, 50–100% mortality of Rosellinia necatrix Berl. ex Prill after 4 h at 38 °C, mycelium mortality of Phytophthora cinnamomi after 1–2 h at 38–40 °C, Macrophomina phaseolina and Pythium aphanidermatum (strongly declined after 24 h at 40 °C) [83]. Rhizobium spp. have an upper-temperature limit range of 37–47 °C with some strains still capable of nodulation at 45 °C [86]. Although solarization heat treatment has been shown to decrease soil abundances of Rhizobium spp., these bacteria quickly recovered after the establishment of a legume crop [83]. Therefore, a multifaceted solution including soil disinfection and retaining beneficial microbes may benefit if soil temperatures do not go above 41 °C. Regardless, strategies which mitigate RS are not always as effective when they are combined unless all the factors are considered.
4. Conclusions
Arriving at a solution to RS will require experts to reach a consensus on RS-related terminology, develop explicit descriptions for its symptoms, define its etiology, and identify its primary phytopathogens. Consistent terminology would facilitate compiling the literature. Explicit symptom descriptions may aid in detangling compound issues like depleted soil nutrients and autotoxicity, each of which can lead to reduced overall plant biomass in monocultures. The buildup of RS-causing microbes needs to be reduced. The soil disinfection method outlined should also allow the survival of beneficial microbes in the soil instead of aiming for the total elimination of the soil’s microbial load. Once the microbial load of RS soils is reduced, then multiple continuous biological methods should be used to keep RS under control. Such methods include RS-resistant rootstocks, poly-cropping, and inoculations of beneficial microbes. Continuous efforts to use these biological methods to increase plant/microbe diversity is critical, since RS-causing microbes will continuously attempt to build up in the soil throughout this time as well.
Author Contributions
Conceptualization, D.R.N., D.K.M. and J.M.V., writing—original draft preparation, D.R.N. and J.M.V. writing—review and editing, D.R.N., D.K.M. and J.M.V.; visualization, D.R.N.; supervision, J.M.V.; and funding acquisition, J.M.V. All authors have read and agreed to the published version of the manuscript.
Funding
Time dedicated in preparing this review was funded by National Institute of Food and Agriculture/United States Department of Agriculture through a Western Sustainable Agriculture Research Education (SARE), project #SW20-910 with title ‘Developing sustainable peach orchard soil microbiome management practices to control replant disease syndrome’.
Data Availability Statement
No new data were created in the making of this manuscript.
Acknowledgments
The present articles contents are solely the responsibility of the authors and do not necessarily represent the official views of the USDA. Special thanks to Madelene Shehan for reviewing and editing the manuscript for grammar, sentence structure, and syntax.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Simon, M.; Lehndorff, E.; Wrede, A.; Amelung, W. In-field heterogeneity of apple replant disease: Relations to abiotic soil properties. Sci. Hortic. 2020, 259, 108809. [Google Scholar] [CrossRef]
- Mahnkopp, F.; Simon, M.; Lehndorff, E.; Pätzold, S.; Wrede, A.; Winkelmann, T. Induction and diagnosis of apple replant disease ARD): A matter of heterogeneous soil properties? Sci. Hortic. 2018, 241, 167–177. [Google Scholar] [CrossRef]
- He, J.-D.; Shu, B.; Wu, Q.-S. Mycorrhizosphere of fruit crops: Nature and properties. In Fruit Crops; Elsevier: Alpharetta, GA, USA, 2020; pp. 325–338. [Google Scholar]
- Rumberger, A.; Merwin, I.A.; Thies, J.E. Microbial community development in the rhizosphere of apple trees at a replant disease site. Soil Biol. Biochem. 2007, 39, 1645–1654. [Google Scholar] [CrossRef]
- Yang, J.-I.; Ruegger, P.M.; McKenry, M.V.; Becker, J.O.; Borneman, J. Correlations between root-associated microorganisms and peach replant disease symptoms in a California soil. PLoS ONE 2012, 7, e46420. [Google Scholar] [CrossRef] [Green Version]
- Liu, E.-T.; Wang, G.-S.; Li, Y.-Y.; Shen, X.; Chen, X.-S.; Song, F.-H.; Wu, S.-J.; Chen, Q.; Mao, Z.-Q. Replanting affects the tree growth and fruit quality of Gala apple. J. Integr. Agric. 2014, 13, 1699–1706. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M. Elucidation of the microbial complex having a causal role in the development of apple replant disease in Washington. Phytopathology 1998, 88, 930–938. [Google Scholar] [CrossRef] [Green Version]
- Jaffee, B.; Abawi, G.; Mai, W. Role of soil microflora and Pratylenchus penetrans in an apple replant disease. Phytopathology 1982, 72, 247–251. [Google Scholar] [CrossRef]
- Westerveld, S.M.; Shi, F. The history, etiology, and management of ginseng replant disease: A Canadian perspective in review. Can. J. Plant Sci. 2021, 101, 886–901. [Google Scholar] [CrossRef]
- Zhao, Y.-P.; Lin, S.; Chu, L.; Gao, J.; Azeem, S.; Lin, W. Insight into structure dynamics of soil microbiota mediated by the richness of replanted Pseudostellaria heterophylla. Sci. Rep. 2016, 6, 26175. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Winkelmann, T. Biofumigation for Fighting Replant Disease—A Review. Agronomy 2020, 10, 425. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M.; Manici, L.M. Apple replant disease: Role of microbial ecology in cause and control. Annu. Rev. Phytopathol. 2012, 50, 45–65. [Google Scholar] [CrossRef]
- Caruso, F.L.; Neubauer, B.F.; Begin, M.D. A histological study of apple roots affected by replant disease. Can. J. Bot. 1989, 67, 742–749. [Google Scholar] [CrossRef]
- Ma, W.; Liao, X.; Wang, C.; Zhang, Y. Effects of four cropping patterns of lilium brownii on rhizosphere microbiome structure and replant disease. Plants 2022, 11, 824. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Chen, J.; Khan, M.U.; Wang, J.; Wu, H.; Xiao, Z.; Zhang, Z.; Lin, W. Rhizosphere fungal community dynamics associated with Rehmannia glutinosa replant disease in a consecutive monoculture regime. Phytopathology 2018, 108, 1493–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agmon-Levin, N.; Mosca, M.; Petri, M.; Shoenfeld, Y. Systemic lupus erythematosus one disease or many? Autoimmun. Rev. 2012, 11, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Sewell, G. Effects of Pythium species on the growth of apple and their possible causal role in apple replant disease. Ann. Appl. Biol. 1981, 97, 31–42. [Google Scholar] [CrossRef]
- Browne, G.; Lampinen, B.; Holtz, B.; Doll, D.; Upadhyaya, S.; Schmidt, L.; Bhat, R.; Udompetaikul, V.; Coates, R.; Hanson, B. Managing the almond and stone fruit replant disease complex with less soil fumigant. Calif. Agric. 2013, 67, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Mai, W.; Abawi, G. Determining the cause and extent of apple, cherry, and pear replant diseases under controlled conditions. Phytopathology 1978, 68, 1540–1544. [Google Scholar] [CrossRef]
- Chen, T.; Lin, S.; Wu, L.; Lin, W.; Sampietro, D.A. Soil sickness: Current status and future perspectives. Allelopath. J. 2015, 36, 167–196. [Google Scholar]
- Thakur, K.K.; Sharma, D.P. Pot-culture studies on soil and leaf nutrients status of peach in response to different soil management techniques for preventing replant disease. J. Pharm. Innov. 2018, 7, 1046–1053. [Google Scholar]
- Spath, M.; Insam, H.; Peintner, U.; Kelderer, M.; Kuhnert, R.; Franke-Whittle, I.H. Linking soil biotic and abiotic factors to apple replant disease: A greenhouse approach. J. Phytopathol. 2015, 163, 287–299. [Google Scholar] [CrossRef]
- Winkelmann, T.; Smalla, K.; Amelung, W.; Baab, G.; Grunewaldt-Stöcker, G.; Kanfra, X.; Meyhöfer, R.; Reim, S.; Schmitz, M.; Vetterlein, D. Apple replant disease: Causes and mitigation strategies. Curr. Issues Mol. Biol. 2019, 30, 89–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Dai, S.; Wang, B.; Jiang, Y.; Ma, Y.; Pan, L.; Wu, K.; Huang, X.; Zhang, J.; Cai, Z. Autotoxic ginsenoside disrupts soil fungal microbiomes by stimulating potentially pathogenic microbes. Appl. Environ. Microbiol. 2020, 86, e00130-20. [Google Scholar] [CrossRef]
- Mazzola, M.; Mullinix, K. Comparative field efficacy of management strategies containing Brassica napus seed meal or green manure for the control of apple replant disease. Plant Dis. 2005, 89, 1207–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newberger, D.R.; Minas, I.S.; Manter, D.K.; Vivanco, J.M. A Microbiological Approach to Alleviate Soil Replant Syndrome in Peaches. Microorganisms 2023, 11, 1448. [Google Scholar] [CrossRef]
- Li, K.; DiLegge, M.J.; Minas, I.S.; Hamm, A.; Manter, D.; Vivanco, J.M. Soil sterilization leads to re-colonization of a healthier rhizosphere microbiome. Rhizosphere 2019, 12, 100176. [Google Scholar] [CrossRef]
- Arneson, P.; Mai, W. Root diseases of fruit trees in New York State. VII. Costs and returns of preplant soil fumigation in a replanted apple orchard. Plant Dis. Rep. 1976, 60, 1054–1057. [Google Scholar]
- Foy, C.L.; Drake, C.R.; Pirkey, C.L. Impact of herbicides applied annually for twenty-three years in a deciduous orchard. Weed Technol. 1996, 10, 587–591. [Google Scholar] [CrossRef]
- Forge, T.; Neilsen, G.; Neilsen, D. Organically acceptable practices to improve replant success of temperate tree-fruit crops. Sci. Hortic. 2016, 200, 205–214. [Google Scholar] [CrossRef]
- Chalmers, D.; Mitchell, P.; Van Heek, L. Control of Peach Tree Growth and Productivity by Regulated Water Supply, Tree Density, and Summer Pruning1. J. Am. Soc. Hortic. Sci. 1981, 106, 307–312. [Google Scholar] [CrossRef]
- Li, X.; Chu, Y.; Jia, Y.; Yue, H.; Han, Z.; Wang, Y. Changes to bacterial communities and soil metabolites in an apple orchard as a legacy effect of different intercropping plants and soil management practices. Front. Microbiol. 2022, 13, 956840. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Römheld, V. The release of root exudates as affected by the plant’s physiological status. In The Rhizosphere: Biochemistry and Organic Substances at The Soil-Plant Interface; Marcel Dekker Inc.: New York, NY, USA, 2000; pp. 41–93. [Google Scholar]
- Hassan, M.K.; McInroy, J.A.; Kloepper, J.W. The interactions of rhizodeposits with plant growth-promoting rhizobacteria in the rhizosphere: A review. Agriculture 2019, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M. Transformation of soil microbial community structure and Rhizoctonia-suppressive potential in response to apple roots. Phytopathology 1999, 89, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Suchocka, M.; Swoczyna, T.; Kosno-Jończy, J.; Kalaji, H.M. Impact of heavy pruning on development and photosynthesis of Tilia cordata Mill. trees. PLoS ONE 2021, 16, e0256465. [Google Scholar] [CrossRef]
- Zhang, L.; Koc, A.; Wang, X.; Jiang, Y. A review of pruning fruit trees. IOP Conf. Ser. Earth Environ. Sci. 2018, 153, 062029. [Google Scholar] [CrossRef]
- Alfieri, S., Jr.; Seymour, C.; French, W. Cytospora Canker of Peach in Florida. In Proceedings of the Florida State Horticultural Society, Miami, FL, USA, 6–8 November 1973. [Google Scholar]
- Chao, Y.; Marks, L.R.; Pettigrew, M.M.; Hakansson, A.P. Streptococcus pneumoniae biofilm formation and dispersion during colonization and disease. Front. Cell. Infect. Microbiol. 2015, 4, 194. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Lu, Y.-J.; Chen, H.; Day, B. The lifecycle of the plant immune system. Crit. Rev. Plant Sci. 2020, 39, 72–100. [Google Scholar] [CrossRef] [PubMed]
- Atucha, A.; Litus, G. Effect of biochar amendments on peach replant disease. HortScience 2015, 50, 863–868. [Google Scholar] [CrossRef]
- Lü, L.-H.; Wu, Q.-S. Mitigation of replant disease by mycorrhization in horticultural plants: A review. Folia Hortic. 2018, 30, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Teskey, B.J. Tree Fruit Production; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Dejong, T.M.; Negron, C.M.; Favreau, R.; Day, K.; Costes, E.; Lopez, G. Using concepts of shoot growth and architecture to understand and predict responses of peach trees to pruning. In Proceedings of the 7th International Peach Symposium. Acta Hortic. 2012, 962, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Paço, T.A.; Ferreira, M.I.; Conceição, N. Peach orchard evapotranspiration in a sandy soil: Comparison between eddy covariance measurements and estimates by the FAO 56 approach. Agric. Water Manag. 2006, 85, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Nuzzo, V.; Palese, A.M.; Xiloyannis, C.; Celano, G.; Zukowskyj, P.; Dichio, B. Net CO2 storage in Mediterranean olive and peach orchards. Sci. Hortic. 2005, 107, 17–24. [Google Scholar] [CrossRef]
- Warmund, M. Fruit Production; University of Missouri Extension: Columbia, MO, USA, 2009; pp. 1–20. [Google Scholar]
- Badri, D.; Vivanco, J. Regulation and function of root exudates Plant. Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Steinkellner, S.; Lendzemo, V.; Langer, I.; Schweiger, P.; Khaosaad, T.; Toussaint, J.-P.; Vierheilig, H. Flavonoids and strigolactones in root exudates as signals in symbiotic and pathogenic plant-fungus interactions. Molecules 2007, 12, 1290–1306. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Chen, Q.; Zhang, Q.; Long, C.; Jia, W.; Cheng, X. Keystone species affect the relationship between soil microbial diversity and ecosystem function under land use change in subtropical China. Funct. Ecol. 2021, 35, 1159–1170. [Google Scholar] [CrossRef]
- Silindir, M.; Özer, A.Y. Sterilization methods and the comparison of E-beam sterilization with gamma radiation sterilization. Fabad J. Pharm. Sci. 2009, 34, 43–53. [Google Scholar]
- McDonnell, G.; Burke, P. Disinfection: Is it time to reconsider Spaulding? J. Hosp. Infect. 2011, 78, 163–170. [Google Scholar] [CrossRef]
- Ross, R.G.; Delbridge, R.; Kimpinski, J.; McRae, K. Control of apple replant disease in Nova Scotia by soil fumigation with Vorlex and chloropicrin. Can. J. Plant Pathol. 1983, 5, 177–180. [Google Scholar] [CrossRef]
- Eayre, C.; Sims, J.; Ohr, H.; Mackey, B. Evaluation of methyl iodide for control of peach replant disorder. Plant Dis. 2000, 84, 1177–1179. [Google Scholar] [CrossRef] [Green Version]
- Guthman, J.; Brown, S. How Midas Lost Its Golden Touch: Neoliberalism and Activist Strategy in the Demise of Methyl Iodide in California. In The New Food Activism: Opposition, Cooperation, and Collective Action; Alkon, A.H., Guthman, J., Eds.; University of California Press: Oakland, CA, USA, 2017; pp. 80–106. [Google Scholar]
- Hestmark, K.; Fernández-Bayo, J.; Harrold, D.; Randall, T.; Achmon, Y.; Stapleton, J.; Simmons, C.; VanderGheynst, J. Compost induces the accumulation of biopesticidal organic acids during soil biosolarization. Resour. Conserv. Recycl. 2019, 143, 27–35. [Google Scholar] [CrossRef]
- Westerveld, S.M.; Riddle, R.N.; Shi, F. Efficacy of fumigants and biofumigants for the control of replant disease of American ginseng (Panax quinquefolius). Can. J. Plant Pathol. 2023, 45, 405–419. [Google Scholar] [CrossRef]
- Browne, G.; Ott, N.; Poret-Peterson, A.; Gouran, H.; Lampinen, B. Efficacy of anaerobic soil disinfestation for control of Prunus replant disease. Plant Dis. 2018, 102, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroetmann, I.; Kämpfer, P.; Dott, W. The efficiency of sterilization methods for different soils. Zentralbl. Hyg. Umweltmed. 1994, 195, 111–120. [Google Scholar] [PubMed]
- Kelderer, M.; Manici, L.M.; Caputo, F.; Thalheimer, M. Planting in the ‘inter-row’to overcome replant disease in apple orchards: A study on the effectiveness of the practice based on microbial indicators. Plant Soil 2012, 357, 381–393. [Google Scholar] [CrossRef]
- Bent, E.; Loffredo, A.; Yang, J.-I.; McKenry, M.V.; Becker, J.O.; Borneman, J. Investigations into peach seedling stunting caused by a replant soil. FEMS Microbiol. Ecol. 2009, 68, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Santillan, E.; Wuertz, S. Microbiome assembly predictably shapes diversity across a range of disturbance frequencies in experimental microcosms. NPJ Biofilms Microbiomes 2022, 8, 41. [Google Scholar] [CrossRef]
- Sheil, D.; Burslem, D.F. Disturbing hypotheses in tropical forests. Trends Ecol. Evol. 2003, 18, 18–26. [Google Scholar] [CrossRef]
- Dößelmann, B.; Willmann, M.; Steglich, M.; Bunk, B.; Nübel, U.; Peter, S.; Neher, R.A. Rapid and consistent evolution of colistin resistance in extensively drug-resistant Pseudomonas aeruginosa during morbidostat culture. Antimicrob. Agents Chemother. 2017, 61, e00043-17. [Google Scholar] [CrossRef] [Green Version]
- Yim, B.; Nitt, H.; Wrede, A.; Jacquiod, S.; Sørensen, S.J.; Winkelmann, T.; Smalla, K. Effects of soil pre-treatment with Basamid® granules, Brassica juncea, Raphanus sativus, and Tagetes patula on bacterial and fungal communities at two apple replant disease sites. Front. Microbiol. 2017, 8, 1604. [Google Scholar] [CrossRef]
- Gu, Y.-H.; Mazzola, M. Modification of fluorescent pseudomonad community and control of apple replant disease induced in a wheat cultivar-specific manner. Appl. Soil Ecol. 2003, 24, 57–72. [Google Scholar] [CrossRef]
- Roberts, P.; Matthews, W., Jr.; Ehlers, J. Root-knot nematode resistant cowpea cover crops in tomato production systems. Agron. J. 2005, 97, 1626–1635. [Google Scholar] [CrossRef]
- Hooks, C.R.; Wang, K.-H.; Ploeg, A.; McSorley, R. Using marigold (Tagetes spp.) as a cover crop to protect crops from plant-parasitic nematodes. Appl. Soil Ecol. 2010, 46, 307–320. [Google Scholar] [CrossRef]
- Wang, K.-H.; Sipes, B.; Schmitt, D. Crotalaria as a cover crop for nematode management: A review. Nematropica 2002, 32, 35–58. [Google Scholar]
- Abd Alhady, M.R.A. In vitro propagation for peach rootstock (Nemaguard). Egypt. J. Desert Res. 2018, 68, 1–13. [Google Scholar] [CrossRef]
- Wani, I.A.; Ahanger, R.A.; Bhat, H.A.; Lone, A.A.; Bhat, T.A.; Malik, I.A.; Hassan, G. Rootstocks of almond. J. Plant Dev. Sci. Vol. 2012, 4, 137–150. [Google Scholar]
- Schneider, S.M.; Rosskopf, E.N.; Leesch, J.G.; Chellemi, D.O.; Bull, C.T.; Mazzola, M. United States Department of Agriculture—Agricultural Research Service research on alternatives to methyl bromide: Pre-plant and post-harvest. Pest Manag. Sci. Former. Pestic. Sci. 2003, 59, 814–826. [Google Scholar] [CrossRef]
- Cesarano, G.; Zotti, M.; Antignani, V.; Marra, R.; Scala, F.; Bonanomi, G. Soil sickness and negative plant-soil feedback: A reappraisal of hypotheses. J. Plant Pathol. 2017, 99, 545–570. [Google Scholar]
- Jiménez, S.; Pinochet, J.; Romero, J.; Gogorcena, Y.; Moreno, M.Á.; Espada, J.L. Performance of peach and plum based rootstocks of different vigour on a late peach cultivar in replant and calcareous conditions. Sci. Hortic. 2011, 129, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; von Wettberg, E.J.; Miller, A.J. Rootstocks: Diversity, domestication, and impacts on shoot phenotypes. Trends Plant Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Wang, Y.; Li, W.; Xu, X.; Qiu, C.; Wu, T.; Wei, Q.; Ma, F.; Han, Z. Progress of apple rootstock breeding and its use. Hortic. Plant J. 2019, 5, 183–191. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Lifshitz, R.; Zablotowicz, R.M. Free-living bacterial inocula for enhancing crop productivity. Trends Biotechnol. 1989, 7, 39–44. [Google Scholar] [CrossRef]
- Benizri, E.; Piutti, S.; Verger, S.; Pages, L.; Vercambre, G.; Poessel, J.; Michelot, P. Replant diseases: Bacterial community structure and diversity in peach rhizosphere as determined by metabolic and genetic fingerprinting. Soil Biol. Biochem. 2005, 37, 1738–1746. [Google Scholar] [CrossRef]
- Gao, W.-Q.; Lü, L.-H.; Srivastava, A.; Wu, Q.-S.; Kuča, K. Effects of mycorrhizae on physiological responses and relevant gene expression of peach affected by replant disease. Agronomy 2020, 10, 186. [Google Scholar] [CrossRef] [Green Version]
- Lǚ, L.-H.; Zou, Y.-N.; Wu, Q.-S. Mycorrhizas mitigate soil replant disease of peach through regulating root exudates, soil microbial population, and soil aggregate stability. Commun. Soil Sci. Plant Anal. 2019, 50, 909–921. [Google Scholar] [CrossRef]
- Weller, D.M. Biological control of soilborne plant pathogens in the rhizosphere with bacteria. Ann. Rev. Phytopathol. 1988, 26, 379–407. [Google Scholar] [CrossRef]
- D’Addabbo, T.; Miccolis, V.; Basile, M.; Candido, V. Soil solarization and sustainable agriculture. In Sociology, Organic Farming, Climate Change and Soil Science; Springer Dordrecht: Dordrecht, The Netherlands, 2010; pp. 217–274. [Google Scholar]
- Baker, K.F.; Chandler, P.A. UC system for producing healthy container-grown plants. AIBS Bull. 1957, 8, 46. [Google Scholar]
- Pullman, G.; DeVay, J.; Garber, R. Soil solarization and thermal death: A logarithmic relationship between time and temperature for four soilborne plant pathogens. Phytopathology 1981, 71, 959–964. [Google Scholar] [CrossRef]
- Patel, A.K.; Banjare, U.; Kumari, A.; Singh, R.K.; Pandey, K.D. Temperature tolerant Rhizobium leguminosorum bv. viciae strains with plant growth promotion traits. J. Pure Appl. Microbiol. 2020, 14, 2603–2609. [Google Scholar] [CrossRef]
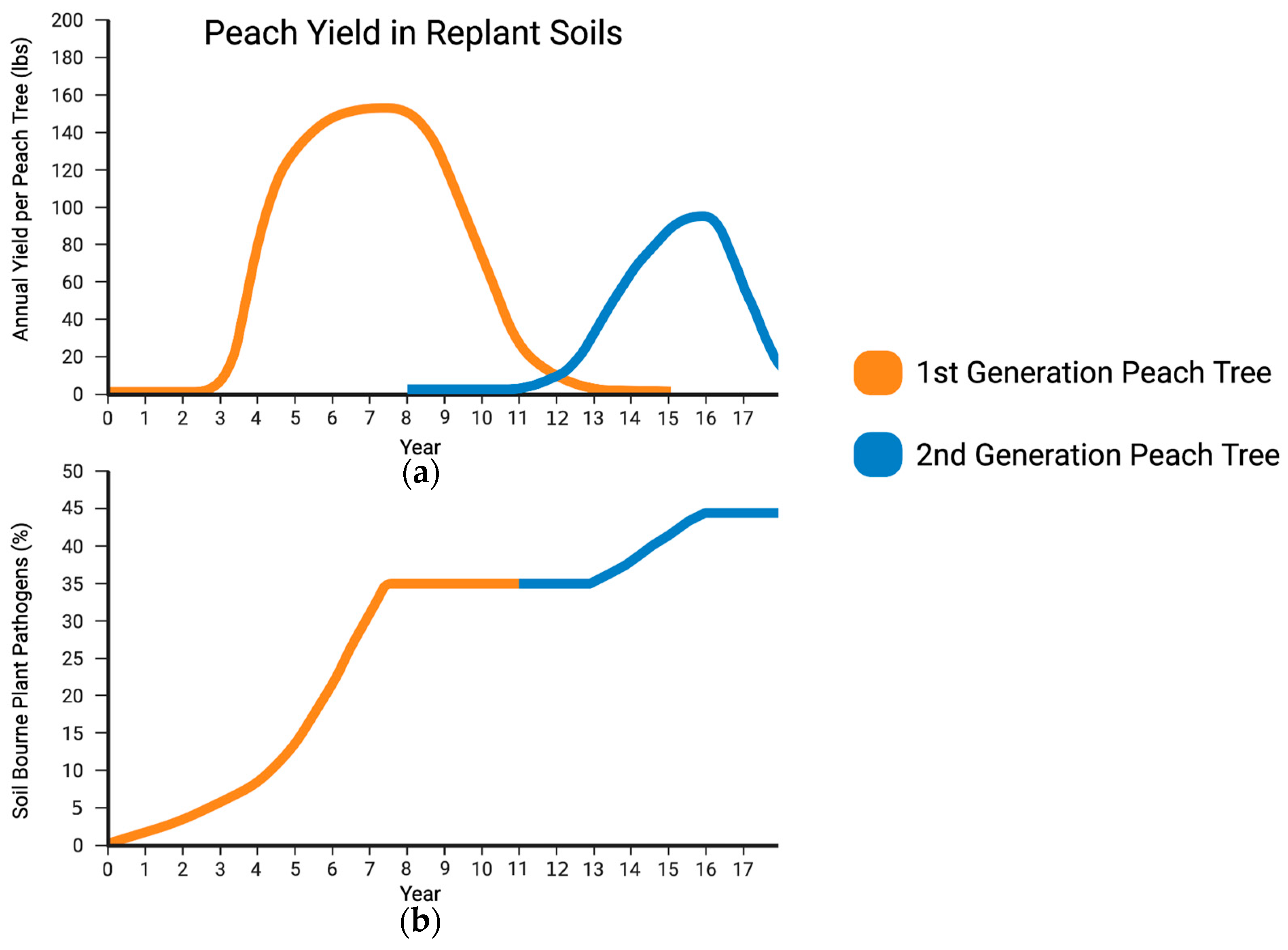
Figure 1.
Proposed concept of the development of a replant syndrome microbiome. (a) First-generation peach trees show expected and typical growth. In the first replanting or second generation, the symptoms of the replant microbiome are observed by the impact it has on the developing fruit tree’s crop yield. (b) A replant microbiome is established within the first generation of a monocropping orchard. Replant symptoms are immediately evident on newly planted saplings since the pathogens in the soil microbiome have been established previously. Phytopathogens that make up the replant microbiome can potentially specialize to be specific to the orchard genotype, and the pathogenic microbial load is at its peak biomass when crop production is at its highest.
Figure 1.
Proposed concept of the development of a replant syndrome microbiome. (a) First-generation peach trees show expected and typical growth. In the first replanting or second generation, the symptoms of the replant microbiome are observed by the impact it has on the developing fruit tree’s crop yield. (b) A replant microbiome is established within the first generation of a monocropping orchard. Replant symptoms are immediately evident on newly planted saplings since the pathogens in the soil microbiome have been established previously. Phytopathogens that make up the replant microbiome can potentially specialize to be specific to the orchard genotype, and the pathogenic microbial load is at its peak biomass when crop production is at its highest.

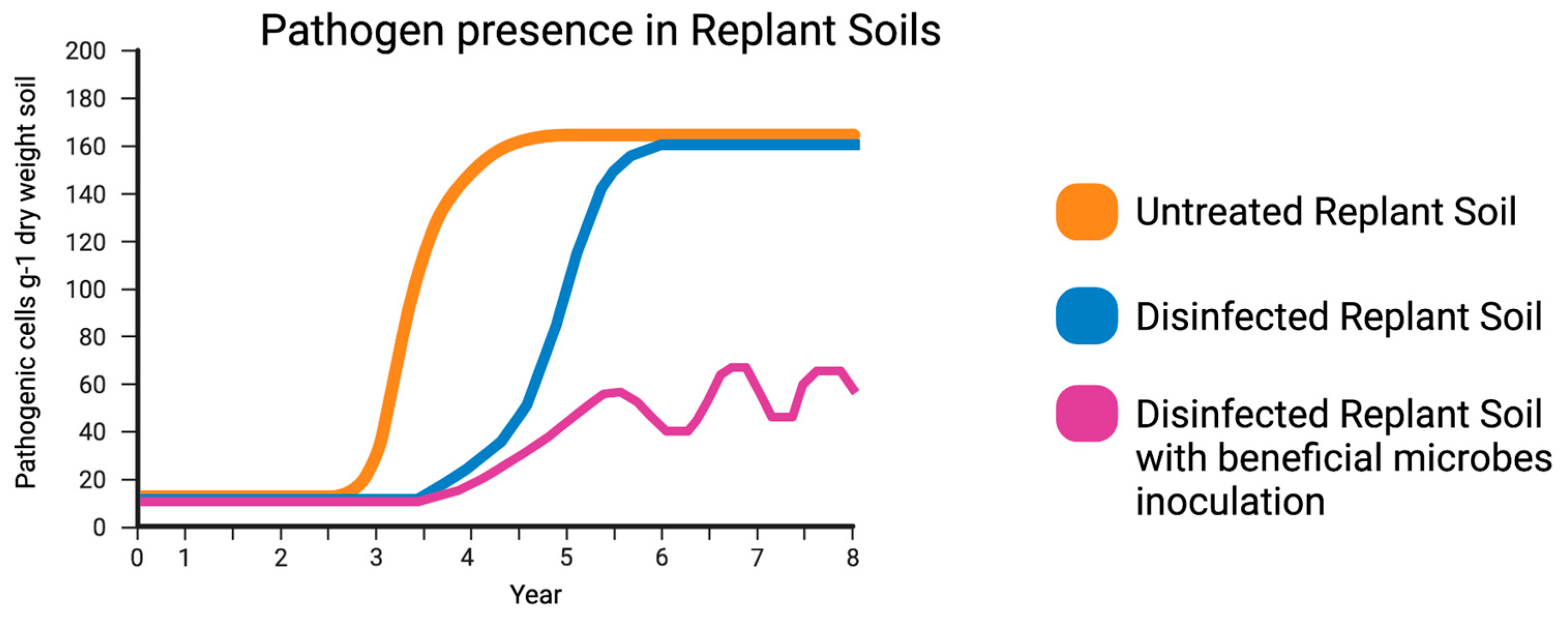
Figure 2.
Proposed concept of the relative pathogenic gene expression or pathogenic cells g-1 dry weight soil. Phytopathogen load in untreated replant soils of a peach orchard (orange) gradually increases once the host peach tree is planted with possibly plateauing. Phytopathogen load of disinfected replant soils of a peach orchard (blue) show temporary RS relief. Disinfected replant soil with repeated inoculations of beneficial microbes (violet) would fail to remove all phytopathogens but could control RS microbiome populations.
Figure 2.
Proposed concept of the relative pathogenic gene expression or pathogenic cells g-1 dry weight soil. Phytopathogen load in untreated replant soils of a peach orchard (orange) gradually increases once the host peach tree is planted with possibly plateauing. Phytopathogen load of disinfected replant soils of a peach orchard (blue) show temporary RS relief. Disinfected replant soil with repeated inoculations of beneficial microbes (violet) would fail to remove all phytopathogens but could control RS microbiome populations.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Newberger, D.R.; Manter, D.K.; Vivanco, J.M. Reviewing the Current Understanding of Replant Syndrome in Orchards from a Soil Microbiome Perspective. Appl. Microbiol. 2023, 3, 856-866. https://doi.org/10.3390/applmicrobiol3030059
AMA Style
Newberger DR, Manter DK, Vivanco JM. Reviewing the Current Understanding of Replant Syndrome in Orchards from a Soil Microbiome Perspective. Applied Microbiology. 2023; 3(3):856-866. https://doi.org/10.3390/applmicrobiol3030059
Chicago/Turabian StyleNewberger, Derek R., Daniel K. Manter, and Jorge M. Vivanco. 2023. "Reviewing the Current Understanding of Replant Syndrome in Orchards from a Soil Microbiome Perspective" Applied Microbiology 3, no. 3: 856-866. https://doi.org/10.3390/applmicrobiol3030059