
Ensiling Typha (Typha latifolia) Forage with Different Additives for Ruminant Feeding: In Vitro Studies

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Typha Samples Collection, Silage Preparation, and Silage Processing
2.2. In Vitro Fermentations: Donor Animals and Experimental Procedures
2.3. Chemical Analyses
2.4. Calculations and Statistical Analyses
3. Results and Discussion
3.1. Chemical Composition and Characteristics of Silages
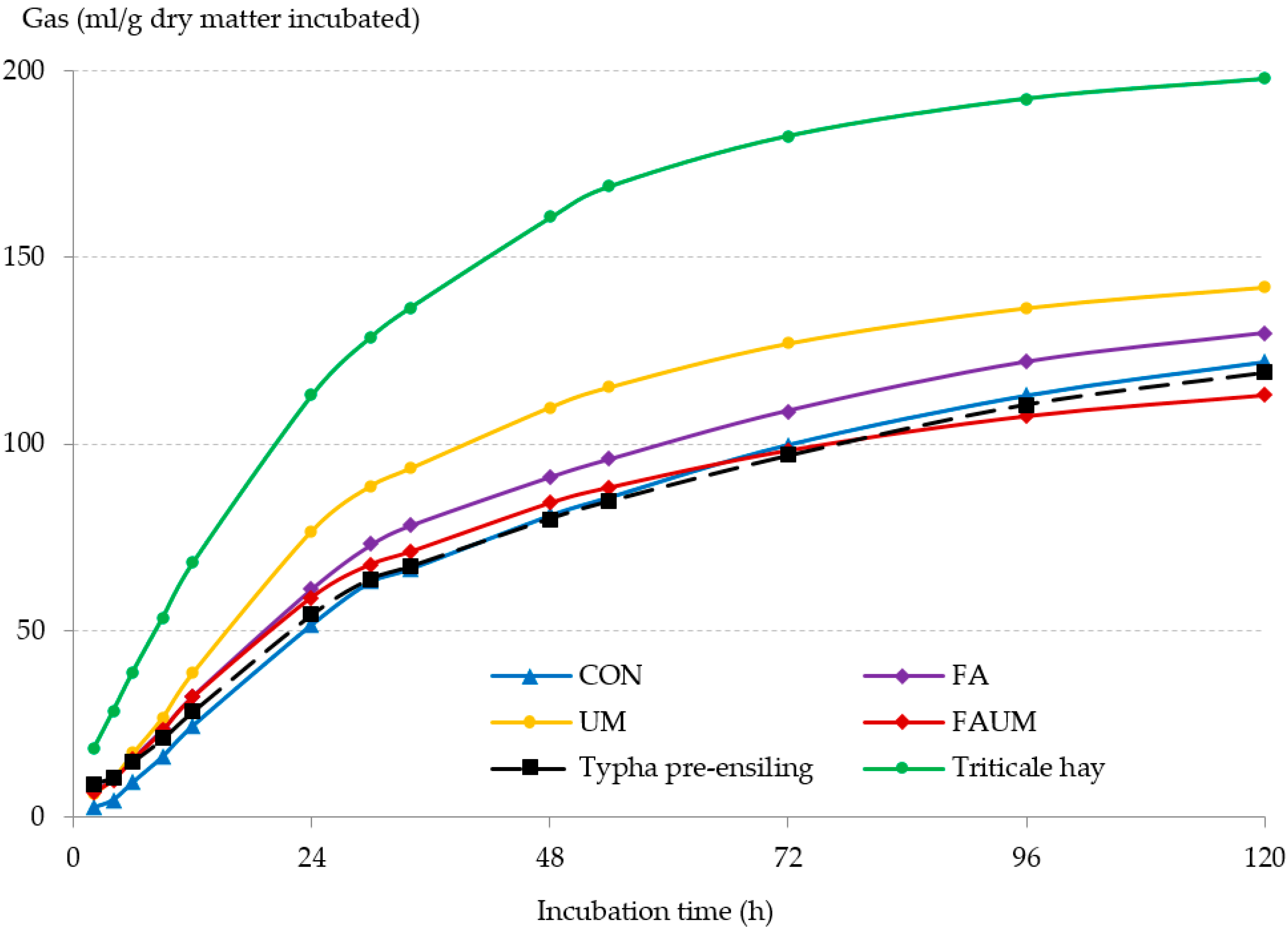
3.2. In Vitro Ruminal Fermentation of Silages: Ruminal Gas Production Kinetics
3.3. In Vitro Ruminal Fermentation of Silages: Fermentative Parameters
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dilshad, R.; Khan, K.U.R.; Saeed, L.; Sherif, A.E.; Ahmad, S.; Ovatlarnporn, C.; Jawad, N.; Hussain, M.; Ghalloo, B.A.; Basit, A.; et al. Chemical composition and biological evaluation of Typha domingensis pers. to ameliorate health pathologies: In vitro and in silico approaches. BioMed Res. Int. 2022, 2022, 8010395. [Google Scholar] [CrossRef]
- He, D.; Simoneit, B.R.; Jara, B.; Jaffé, R. Gas chromatography mass spectrometry based profiling of alkyl coumarates and ferulates in two species of cattail (Typha domingensis P., and Typha latifolia L.). Phytochem. Lett. 2015, 13, 91–98. [Google Scholar] [CrossRef]
- Iglesias, E.; Loureiro, M.L.; Escribano, F. Conference Workshop AERNA 2018 Household Perceptions on Biogas as a Sustainable Energy Source. A focus group study in Hadejia Valley, Nigeria. Available online: http://typhaproject.com/wp-content/uploads/2019/04/Iglesias-Loureiro-Escribano-AERNA-2018-V2.pdf (accessed on 18 April 2023).
- de Evan, T.; Marcos, C.N.; Carro, M.D.; Alao, J.S.; Makinde, J.O.; Rufai, M.A.; Iglesias, E.; Escribano, F. Nutritive value of typha for ruminants. In Proceedings of the 44th Annual Conference of the Nigerian Society for Animal Production, Abuja, Nigeria, 17–21 March 2019; pp. 48–50. Available online: www.typhaproject.com (accessed on 12 April 2023).
- WingChing-Jones, R.; Leal, J. Conservación del forraje de la Typha domingensis Typhaceae. Ensilaje y henificación. Cuadernos Investig. UNED 2018, 10, 107–114. (In Spanish) [Google Scholar] [CrossRef]
- Musa, A.R.; Garba, Y. Nutritive value of untreated and molasses-urea treated Typha (Typha domingensis) silage. FUDMA J. Agric. Agric. Technol. 2022, 8, 70–76. [Google Scholar] [CrossRef]
- Makinde, O.J.; Musa, A.R.; Sunday, A.J.; Escribano, F.; Kohn, R.; Iglesias, E.; Maidala, A.; Amos, M.; Chana, M.; Hannatu, C.; et al. Cattail (Typha domingensis) silage improves feed intake, blood profile, economics of production, and growth performance of beef cattle. Trop. Anim. Health Prod. 2022, 54, 48. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Musa, A.R.; de Evan, T.; Alao, J.S.; Iglesias, E.; Escribano, F.; Carro, M.D. Effects of different additives on the quality of typha grass (Typha latifolia) silages. In Proceedings of the 25th Annual Conference of ASAN 2020, Abuja, Nigeria, 1–4 November 2020; pp. 589–592. [Google Scholar]
- Samarasinghe, M.B.; Larsen, M.; Johansen, M.; Waldemar, P.; Weisbjerg, M.R. Effects of shredding on silage density and fermentation quality. Grass For. Sci. 2019, 74, 244–2453. [Google Scholar] [CrossRef]
- Goering, M.K.; Van Soest, P.J. Forage Fiber Analysis (Apparatus, Reagents, Procedures and Some Applications). In Agricultural Handbook; Agriculture Handbook Nº 379; Agricultural Research Services: Washington, DC, USA, 1970. [Google Scholar]
- AOAC. Association of Official Analytical Chemists. Official Methods of Analysis, 18th ed.; 9th rev.; AOAC: Gaithersburg, MD, USA, 2015. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.B.; Van Soest, P.J. The detergent system of analysis and its application to human foods. In The Analysis of Dietary Fiber in Food; James, W.P.T., Theander, O., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1981; pp. 123–142. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- García-Martínez, R.; Ranilla, M.J.; Tejido, M.L.; Carro, M.D. Effects of disodium fumarate on in vitro rumen microbial growth, methane production and fermentation of diets differing in their forage concentrate ratio. Br. J. Nutr. 2005, 94, 71–77. [Google Scholar] [CrossRef]
- Taylor, K.A. A simple colorimetric assay for muramic acid and lactic acid. Appl. Biochem. Biotechnol. 1996, 56, 49–58. [Google Scholar] [CrossRef]
- Krishnamoorthy, U.; Soller, H.; Steingass, H.; Menke, K.H. A comparative study on rumen fermentation of energy supplements in vitro. J. Anim. Physiol. Anim. Nutr. 1991, 65, 28–35. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT® Users Guide, version 9.3; SAS Inst. Inc.: Cary, NC, USA, 2017. [Google Scholar]
- France, J.; Dijkstra, J.; Dhanoa, M.S.; Lopez, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: Derivation of models and other mathematical considerations. Br. J. Nutr. 2000, 83, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kung, L., Jr.; Robinson, J.R.; Ranjit, N.K.; Chen, J.H.; Golt, C.M.; Pesek, J.D. Microbial populations, fermentation endproducts, and aerobic stability of corn silage treated with ammonia or a propionic acid-based preservative. J. Dairy Sci. 2000, 83, 1479–1486. [Google Scholar] [CrossRef]
- Yunus, M.; Ohba, N.; Shimojo, M.; Furuse, M.; Masuda, Y. Effects of Adding Urea and Molasses on Napiergrass Silage Quality. Anim. Biosci. 2000, 13, 1542–1547. [Google Scholar] [CrossRef]
- Dos Santos, A.P.M.; Santos, E.M.; Oliveira, J.S.D.; Ribeiro, O.L.; Perazzo, A.F.; Pinho, R.M.A.; Macêdo, A.J.D.S.; Pereira, G.A. Effects of urea addition on the fermentation of sorghum (Sorghum bicolor) silage. Afr. J. Range Forage Sci. 2018, 35, 55–62. [Google Scholar] [CrossRef]
- Stephanie, D.C.; Simon, R.L. Kinetic properties of Helicobacter pylori urease compared with Jack. bean urease. FEMS Microbiol. Lett. 1992, 99, 5–21. [Google Scholar] [CrossRef]
- Apajalahti, J.; Vienola, K.; Raatikainen, K.; Holder, V.; Moran, C.A. Conversion of Branched-Chain Amino Acids to Corresponding Isoacids—An in vitro Tool for Estimating Ruminal Protein Degradability. Front. Vet. Sci. 2019, 6, 311. [Google Scholar] [CrossRef]
- Hinds, M.A.; Bolsen, K.K.; Brethour, J.; Milliken, G.; Hoover, J. Effects of molasses/urea and bacterial inoculant additives on silage quality, dry matter recovery, and feeding value for cattle. Anim. Feed Sci. Technol. 1985, 12, 205–214. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe: Marlow, UK, 1991. [Google Scholar]
- Jaakkola, S.; Huhtanen, P.; Hissa, K. The effect of cell wall degrading enzymes or formic acid on fermentation quality and on digestion of grass silage by cattle. Grass Forage Sci. 1991, 46, 75–87. [Google Scholar] [CrossRef]
- Novoa-Garrido, M.; Marcos, C.N.; Carro, M.D.; Molina-Alcaide, E.; Larsen, M.; Weisbjerg, M.R. Preserving Porphyra umbilicalis and Saccharina latissima as Silages for Ruminant Feeding. Animals 2020, 10, 1957. [Google Scholar] [CrossRef] [PubMed]
- WingChing-Jones, R.; Leal-Rivera, J.C. Valoración agronómica y nutricional de la Typha domingensis, como alternativa de alimentación en animales rumiantes. Nutr. Anim. Trop. 2014, 8, 24–35. [Google Scholar]
- Han, L.; Zhou, H. Effects of ensiling processes and antioxidants on fatty acid concentrations and compositions in corn silages. J. Anim. Sci. Biotechnol. 2013, 4, 48. [Google Scholar] [CrossRef] [PubMed]
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstu_s from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef]
- Lunsin, R.; Duanyai, S.; Pilajun, R.; Duanyai, S.; Sombatsri, P. Effect of urea-and molasses-treated sugarcane bagasse on nutrient composition and in vitro rumen fermentation in dairy cows. Agric. Nat. Resour. 2018, 52, 622–627. [Google Scholar] [CrossRef]
- Phesatcha, K.; Wanapat, M. Improvement of nutritive value and in vitro ruminal fermentation of Leucaena silage by molasses and urea supplementation. Asian-Australas. J. Anim. Sci. 2016, 29, 1136. [Google Scholar] [CrossRef]
- Van Dung, D.; Shang, W.; Yao, W. Effect of crude protein levels in concentrate and concentrate levels in diet on in vitro fermentation. Asian-Australas. J. Anim. Sci. 2014, 27, 797. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, H.; Yu, Z. Effects of sucrose, formic acid and lactic acid bacteria inoculant on quality, in vitro rumen digestibility and fermentability of drooping wild ryegrass (Elymus nutans Griseb.) silage. J. Anim. Feed Sci. Technol. 2017, 26, 26–32. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, S.; Dong, Z.; Li, J.; Jia, Y.; Shao, T. Effect of storage time and the level of formic acid on fermentation characteristics, epiphytic microflora, carbohydrate components and in vitro digestibility of rice straw silage. Anim. Biosci. 2021, 34, 1038–1048. [Google Scholar] [CrossRef]
- Aksu, T.; Baytok, E.; Karslı, M.A.; Muruz, H. Effects of formic acid, molasses and inoculant additives on corn silage composition, organic matter digestibility and microbial protein synthesis in sheep. Small Rum. Res. 2006, 61, 29–33. [Google Scholar] [CrossRef]
- Chamberlain, D.G.; Thomas, P.C.; Wait, M.K. The rate of addition of formic acid to grass at ensilage and the subsequent digestion of the silage in the rumen and intestines of sheep. Grass Forage Sci. 1982, 37, 159–164. [Google Scholar] [CrossRef]


{kind=link}
| Silage Treatment | ||||||
|---|---|---|---|---|---|---|
| Item | CON | FA | UM | FAUM | SEM 2 | p = |
| Fresh matter (g/bag) | 410 a | 404 a | 440 b | 442 b | 3.4 | <0.001 |
| Recovery of dry matter (DM, %) | 87.7 | 96.8 | 88.9 | 94.1 | 2.43 | 0.089 |
| DM (%) | 30.4 a | 34.6 b | 32.6 ab | 34.6 b | 0.98 | 0.046 |
| pH | 5.39 b | 4.75 a | 8.50 d | 7.70 c | 0.114 | <0.001 |
| NH3-N (g/kg DM) 3 | 0.221 a | 0.00 a | 9.06 c | 7.22 b | 0.156 | <0.001 |
| VFA 4 (g/kg DM) | ||||||
| Acetic | 16.6 b | 8.86 a | 46.1 c | 13.7 ab | 1.15 | <0.001 |
| Propionic | 3.84 b | 0.22 a | 0.99 a | 0.88 a | 0.603 | 0.012 |
| Butyric | 17.2 c | 0.76 a | 11.8 bc | 4.84 ab | 2.34 | 0.005 |
| Isobutyric | 1.42 b | 0.19 a | 0.62 ab | 0.00 a | 0.262 | 0.0214 |
| Isovaleric | 0.32 b | 0.00 a | 0.69 c | 0.00 a | 0.0207 | <0.001 |
| Valeric | 2.73 b | 0.00 a | 0.00 a | 0.00 a | 0.144 | <0.001 |
| Total VFA | 42.0 b | 10.0 a | 60.2 c | 19.4 a | 3.78 | <0.001 |
| Lactic acid (g/kg DM) | 4.89 a | 14.5 a | 19.1 b | 17.0 ab | 2.15 | 0.028 |
| Silage Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|
| Item | CON | FA | UM | FAUM | SEM 2 | p = | Thypha Pre-Ensiling | Triticale Hay |
| Crude ash | 123 b | 117 a | 135 c | 123 b | 1.2 | <0.001 | 120 | 86.0 |
| Crude protein | 114 a | 115 a | 152 b | 222 c | 2.9 | <0.001 | 119 | 109 |
| Ether extract | 39.7 | 36.9 | 46.4 | 48.9 | 4.52 | 0.081 | 48.4 | 23.8 |
| Neutral detergent fibre | 694 bc | 702 c | 666 ab | 643 a | 7.6 | 0.002 | 691 | 587 |
| Acid detergent fibre | 411 | 404 | 415 | 395 | 10.8 | 0.581 | 401 | 315 |
| Acid detergent lignin | 69.2 ab | 63.1 a | 73.3 b | 65.8 a | 1.64 | 0.012 | 66.9 | 37.4 |
| Silage | ||||||||
|---|---|---|---|---|---|---|---|---|
| Item 2 | CON | FA | UM | FAUM | SEM 3 | p = | Typha Pre-Ensiling | Triticale Hay |
| PGP (mL/g DM) | 147 b | 152 b | 149 b | 126 a | 4.9 | 0.002 | 164 | 206 |
| c (%/h) | 4.09 a | 3.72 a | 4.21 ab | 4.94 b | 0.210 | 0.002 | 4.13 | 3.94 |
| Lag (h) | 3.77 c | 1.81 a | 2.78 b | 2.37 ab | 0.326 | 0.001 | 1.33 | 0.51 |
| AGPR (mL/h) | 2.25 a | 2.60 b | 3.29 c | 2.62 b | 0.066 | <0.001 | 2.35 | 5.16 |
| PDMD (%) | 61.7 a | 58.2 a | 77.1 c | 68.5 b | 0.90 | <0.001 | 65.2 | 68.0 |
| Silage Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|
| Item 2 | CON | FA | UM | FAUM | SEM 2 | p = | Typha Pre-Ensiling | Triticale Hay |
| Total VFA (mmol/g dry matter) | 4.33 a | 4.28 a | 5.31 b | 4.65 a | 0.105 | <0.001 | 4.00 | 5.74 |
| Molar proportions (mol/100 mol) | ||||||||
| Acetic (Ac) | 62.1 a | 63.9 b | 65.1 c | 63.3 b | 0.28 | <0.001 | 65.9 | 63.0 |
| Propionic (Pr) | 20.0 a | 22.1 c | 21.2 b | 22.5 c | 0.15 | <0.001 | 18.9 | 23.2 |
| Butyric | 11.7 c | 8.5 a | 9.3 ab | 9.6 b | 0.23 | <0.001 | 9.1 | 8.8 |
| Isobutyric | 1.84 b | 1.55 a | 1.27 a | 1.40 a | 0.070 | <0.001 | 1.21 | 1.25 |
| Isovaleric | 2.87 c | 2.58 b | 1.87 a | 2.05 a | 0.050 | <0.001 | 2.52 | 1.74 |
| Valeric | 1.55 d | 1.36 c | 1.09 a | 1.21 b | 0.027 | <0.001 | 1.76 | 2.06 |
| Ac/Pr (mol/mol) | 3.12 a | 2.91 b | 3.10 a | 2.83 b | 0.028 | <0.001 | 3.57 | 2.74 |
| NH3-N (mg/L) 3 | 257 a | 251 a | 314 b | 359 c | 2.0 | <0.001 | 246 | 250 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Evan, T.; Musa, A.R.; Marcos, C.N.; Alao, J.S.; Iglesias, E.; Escribano, F.; Carro, M.D. Ensiling Typha (Typha latifolia) Forage with Different Additives for Ruminant Feeding: In Vitro Studies. Appl. Sci. 2023, 13, 6546. https://doi.org/10.3390/app13116546
de Evan T, Musa AR, Marcos CN, Alao JS, Iglesias E, Escribano F, Carro MD. Ensiling Typha (Typha latifolia) Forage with Different Additives for Ruminant Feeding: In Vitro Studies. Applied Sciences. 2023; 13(11):6546. https://doi.org/10.3390/app13116546
Chicago/Turabian Stylede Evan, Trinidad, Ahmad Rufai Musa, Carlos N. Marcos, Johnson Sunday Alao, Eva Iglesias, Fernando Escribano, and Maria Dolores Carro. 2023. "Ensiling Typha (Typha latifolia) Forage with Different Additives for Ruminant Feeding: In Vitro Studies" Applied Sciences 13, no. 11: 6546. https://doi.org/10.3390/app13116546