
Evaluating the Efficacy of Common Treatments Used for Vairimorpha (Nosema) spp. Control

1
Entomology and Nematology Department, University of Florida, Gainesville, FL 32611, USA
2
Department of Horticulture, Oregon State University, Corvallis, OR 97331, USA
*
Author to whom correspondence should be addressed.
Appl. Sci. 2023, 13(3), 1303; https://doi.org/10.3390/app13031303
Submission received: 30 November 2022
/
Revised: 9 January 2023
/
Accepted: 10 January 2023
/
Published: 18 January 2023
(This article belongs to the Special Issue Frontier Research in Apiculture (Diagnosis and Control of Bee Diseases, Bee Products, Environmental Monitoring))
Abstract
:Vairimorpha (formerly Nosema) apis and V. ceranae are microsporidian pathogens that are of concern for managed honey bee colonies. Multiple treatments have been proposed to be effective in reducing the prevalence and intensity of Vairimorpha spp. infections. Here, we test the efficacy of these products in one lab-based experiment and three field experiments. In the lab experiment, we found no reductions in Vairimorpha spp. prevalence (proportion of individuals infected with Vairimorpha spp.) or intensity (number of Vairimorpha spp. spores per individual), but we did find a decrease in honey bee survival after treatment with Fumagilin-B, Honey-B-Healthy®, and Nozevit Plus. The first field experiment showed increased Vairimorpha spp. intensity in colonies treated with Fumagilin-B and HiveAlive® compared to a negative control (sucrose syrup alone). The second field experiment showed a weak reduction in Vairimorpha spp. intensity after 3 weeks post treatment with Fumagilin-B compared to Nozevit. However, Vairimorpha spp. intensity returned to levels comparable to those of other treatment groups after 5 weeks post treatment and remained similar to those of other groups for the duration of the experiment. The final field trial showed no positive or negative effects of treatment with Fumagilin-B or Nosevit on Vairimorpha spp. prevalence or intensity. These findings raise questions regarding the efficacy of the products currently being used by beekeepers to control Vairimorpha spp. We argue that the observed reduction of Vairimorpha spp. is more likely relevant to the phenology of spore prevalence and intensity in honey bee colonies than to chemical treatment.
Keywords:
honey bee; Vairimorpha spp.; Fumagilin-B; HiveAlive®; Nozevit; treatment; prevalence; intensity1. Introduction
The western honey bee, Apis mellifera, is an economically important pollinator in agricultural systems and faces stressors such as pathogens, parasites, and pests [1]. Vairimorpha (formerly Nosema, [2]) spp. are microsporidian parasites that reproduce in the midgut of honey bees and have been shown to be detrimental to honey bee health [3,4]. Infections with Vairimorpha spp. alter behavior, reduce immunity to other pathogens, increase energetic demand, and lead to increased mortality of individuals bees [5,6]. To date, three species of Vairimorpha have been found to infect honey bees; V. apis, V. ceranae, and V. neumanni [7,8, 9]. These species differ in virulence, geographic range, and clinical signs associated with infection in honey bees [10].
In North America, fumagillin is the only approved antibiotic treatment for Vairimorpha spp. infections [11]. Fumagillin is a non-specific antibiotic derived from Aspergillus fumigatus [12]. Historically, a product called Fumagilin-B (Medivet Pharmaceuticals Ltd., High River, AD, Canada) was the only fumagillin product used in North America. In 2018, Medivet ceased production of this treatment, and Fumadil-B (KBNP, Inc., Anyang, Republic of Korea) is now the commonly used fumagillin-based treatment. While fumagillin products have been shown by some to be effective at reducing Vairimorpha spp. prevalence and intensity [13,14], others have shown that treatment with fumagillin does not always control Vairimorpha spp. infection [15,16,17,18].
Some researchers have proposed that proper honey bee nutrition can prevent negative impacts of Vairimorpha spp. in honey bee colonies. Colonies with a high pollen diet have been shown to cope with Vairimorpha ceranae infection better than colonies that consume less pollen [19,20]. Furthermore, nutritional stress can change honey bee gut microbiota and suppress immune function, thus favoring V. ceranae infection [21]. Manufacturers of certain natural plant products, such as Nozevit (Apivita, Varaždin, Croatia) and Honey-B-Healthy® (Honey-B-Healthy, Inc, Cumberland, MD, USA), claim that treatments improve honey bee nutrition and ultimately colony survival following Vairimorpha spp. infection [17,22]. Similarly, another feeding supplement called HiveAlive® (Advance Science Ltd., Galway, Ireland) is purported to support good nutrition and intestinal well-being of honey bees, thereby reducing V. ceranae infection over time [23]. Many beekeepers throughout the world feed Nozevit, Honey-B-Healthy®, and HiveAlive® to their honey bee colonies several times each year with the intention of fortifying their immune response and protecting them from mortality caused by Vairimorpha spp.
In the present study, we tested Vairimorpha spp. infection control in honey bees by treatments (both registered and unregistered for Vairimorpha spp. control) commonly used by beekeepers. We conducted four separate experiments to test the efficacy of these treatments. In the first experiment, we compared the prevalence and intensity of V. ceranae and the subsequent survival of caged honey bees exposed to V. ceranae and treated with Fumagilin-B, Nozevit Plus, and Honey-B-Healthy®. In the second experiment, honey bee colonies were treated during the winter with Fumagilin-B and HiveAlive® and sampled monthly until spring. In the third and fourth experiments, colonies were treated during the winter with Nozevit Plus and Fumagilin-B following label recommendations for either “fall” or “spring” treatments. The objective of these experiments was to evaluate the efficacy of different treatments to reduce the prevalence (proportion of individuals infected with Vairimorpha spp.) and intensity (number of Vairimorpha spp. spores per individual) of Vairimorpha spp. infection, and to improve the health of honey bee colonies.
2. Materials and Methods
2.1. Experiment 1: Laboratory Cage Study for Evalting Efficacy of Fumagilin-B, Nozevit Plus, and Honey-B-Healthy® against Vairimorpha ceranae
In July 2014, capped brood combs were obtained from honey bee colonies at Oregon State University apiaries (Corvallis, OR, USA). We placed the combs in an incubator under simulated hive conditions (33 °C, 55% RH) to facilitate adult worker bee emergence. Twenty-four hours later, we gently brushed newly emerged bees into a large container and mixed them gently by hand. After the bees were mixed, we placed 250 individual bees inside cylindrical wire cages (63.06 cm3) and returned them to the incubator, as per [19]. The caged bees were immediately provided with ad libitum access to a glass vial containing 25 mL of a V. ceranae spore/50% sucrose solution dosed at 40,000 spores/bee. The vials were covered with two layers of cheesecloth and then secured, inverted, and placed upon the top of the cage. Each cage also contained 25 g of finely ground wildflower pollen mixed with a 33% sucrose solution in a 2:1 (weight/volume) ratio. Prior to the experiment, the wildflower pollen was sent to the USDA National Science Laboratory (Gastonia, NC, USA) for pesticide analysis to assess the possible presence of pesticide residues. We found 34 ppb of fluvalinate, 3.7 ppb of chlorfenopyr, 2.6 ppb of trifluralin, and trace concentrations of all other pesticides that were tested for. The results of this pesticide residue analysis are presented as Supplementary Table S1 in the Supplemental Materials.
The spore concentration of the V. ceranae solution was formulated and purified by centrifugation following the methods of [24]. Briefly, the contents of infected gut samples were collected and centrifuged at 5000 rpm for five minutes at room temperature to produce a pellet of spores. After discarding the supernatant, the pellet was resuspended in distilled water by vortexing. This process was repeated 2–3 times. DNA analysis was performed using the methods of [25] to confirm that only V. ceranae spores were present in the inoculum. Once the 25 mL solution containing V. ceranae was completely consumed, the caged bees were provided ad libitum access to water and 50% sucrose syrup (weight/volume). Three days after inoculation of bees with V. ceranae spores, we provided 25 mL of sucrose syrup containing one of four treatments to the cages. There were five cage replicates per treatment for the following four treatments: Fumagilin-B, Nozevit Plus, Honey-B-Healthy®, and a negative control (provided only 50% sucrose syrup). We prepared all treatments in 50% sucrose syrup according to the product label. Once the bees completely consumed the 25 mL of treated sucrose syrup, the caged bees were again given ad libitum access to water and sucrose syrup. In all, there were four treatments and five replicates (cages) of each treatment, for a total of 20 cages.
Once a week, we measured consumption of the pollen by bees and replaced unconsumed pollen with fresh pollen mixed with sucrose solution as described previously. Bee mortality was recorded every other day and dead bees were removed at the time of diet replacement for convenience. We measured consumption of both water and sucrose solution and replaced them on alternate days. At 16 and 23 days after spore inoculation, we removed 25 bees at random from each experimental cage for infection analysis. The abdomens of the bees were used to estimate V. ceranae prevalence (presence or absence of spores) and intensity (# of spores/bee) by light microscopy techniques as described by [26]. Each bee abdomen was checked individually for V. ceranae infection.
2.2. Experiment 2: Field Treatment Using HiveAlive® and Fumagilin-B
In December 2017, 90 colonies located in Waldo, Florida were identified and grouped into three apiaries of 30 colonies per apiary. As honey bee colony dynamics can influence Vairimorpha spp. prevalence and intensity [27], we ensured that all 90 colonies were of similar size and strength through equalization of the number of brood and adult bees prior to study initiation. These colonies were maintained by a local commercial beekeeper following best management practices common in the region (swarms controlled, pests managed, fed when necessary, etc.). Thirty colonies received HiveAlive® according to the product label. Another thirty colonies were treated with Fumagilin-B according to the product label. The remaining 30 colonies belonged to the negative control group and received untreated sugar syrup. All colonies were fed at the same time and the same amount of syrup, with at least 4 L of syrup per seasonal feeding.
On the day of first treatment, baseline data from all colonies were collected by sampling for Vairimorpha spp. and Varroa destructor (a parasitic mite of honey bees). Samples of adult bees were collected by shaking brood frames with nurse-aged bees from the combs onto a pan. About 300 bees were poured from the pan into sampling jars containing 70% ethanol. Later, Vairimorpha spp. intensity was estimated using the methods described by [27] from 100 bee pooled sub-samples. The infestation of V. destructor was determined as described by [28]. Vairimorpha spp. and V. destructor levels were monitored for every colony every 4–6 weeks from December 2017–May 2018.
2.3. Experiments 3 and 4: Winter Field Treatment Using Fumagilin-B and Nozevit at Fall and Spring Recommended Treatments
In January 2009 and December 2009 for Experiment 3 and 4 respectively, honey bee colonies in Umatilla, Florida and Windsor, Florida were assessed for the presence of Vairimorpha spp. infection.
2.4. Colony Selection
In Experiment 3, colonies were first randomly assigned to treatment groups, and then sampled for Vairimorpha spp. infection. The intensity of Vairimorpha spp. was not different among colonies or between treatments at the beginning of the experiment. In Experiment 4, we chose colonies that had a mild to moderate Vairimorpha spp. infection (~50,000 to 500,000 spores per bee) as candidates for the study. Of these positively infected candidates, 50 queen right colonies of similar colony strength were selected and divided into the five treatment groups. All hives were equalized for honey stores within the supers. Each plot of ten hives was located in an open sunny field and placed on wooden pallets, approximately 15 m apart from other plots of colonies.
2.5. Vairimorpha spp. Treatments and Experimental Design
Negative control treatment groups received only sucrose syrup at each feeding application. We created two treatments for Nozevit and Fumagilin-B (Medivet Pharmaceuticals Ltd., Alberta Canada). We mixed dosages according to product labels from both products and made applications two (equivalent to a labeled “spring” treatment) and four (equivalent to a labeled “fall” treatment) times, seven days apart from one another. The two applications (“spring” treatment) were followed by two feedings of only sugar syrup when the “fall” treatment group received its third and fourth treatment. In total, we used five treatments in both experiments: a negative control, “spring” treatments (two applications) of Nozevit and Fumagilin-B, and “fall” treatments (four applications) of Nozevit and Fumagilin-B.
2.6. Sampling for Vairimorpha spp. Infection
For each sample of bees for Vairimorpha spp. assessment, we collected 100 bees per hive and placed them in 100 mL containers with 70% ethanol and returned them to the laboratory. There, we removed the abdomens of 100 bees per sample and combined them with 100 mL distilled water. We placed the solution into a sterilized Cooks Power Blender (JC Penny, Manchester, CT, USA) and blended for 30 s until an even suspension was formed. Vairimorpha spp. intensity was estimated using the methods described [26] from 100 bee pooled sub-samples. Using the 5-square method reported by [26], we calculated the number of spores per bee for each sample. In Experiment 3, we sampled baseline Vairimorpha spp. levels, again one week after the second treatment of all colonies, then one week after the fourth treatment of applicable colonies, and finally, three weeks after the fourth treatment. In Experiment 4, we again sampled Vairimorpha spp. at a baseline, followed by one week post treatment, two weeks post treatment, and a final sampling four weeks after the initial treatment.
2.7. Colony Assessment
The colony assessors were blind to the treatment group assignments and had not visited the site during treatment application. In Experiment 3, two observers estimated the area covered by bees and brood of all combs in the hives at the end of the experiment, and the average estimation of the observers was calculated [29]. In Experiment 4, a baseline and final assessment were made by one assessor.
2.8. Statistical Analyses
Statistical analyses for all experiments were performed using R version 4.1.1. The effect of time was analyzed as a factor in all analyses where applicable. In Experiment 1, we used linear models (normal error structures) to test for relationships between the Vairimorpha spp. treatment and average bee consumption of pollen, water, and sucrose using the following model structure in the lme4 package [30]: [response variable = treatment * time]. To compare bee survival across treatments, we used a Cox proportional hazards model in the survival package [31]. Bees that survived until the end of the experiment (Day 28) and those that were removed for V. ceranae intensity and prevalence were treated as censored cases.
To test the effects of Fumagilin-B and HiveAlive® on Vairimorpha spp. intensity, we used a linear mixed model in the lme4 package [30] using the log spore load + 1 as the response variable. Prevalence of Vairimorpha spp. and V. destructor in Experiment 2 were analyzed using general linear mixed models with binomial error distributions. For all response variables in Experiment 2, the error structures were: [response variable = treatment * time + colony (random effect)]. In Experiments 3 and 4, we analyzed the effect of the treatments (Fumagilin-B at fall and spring treatments, Nozevit at fall and spring treatments, and the negative control group) on estimations of bees and brood using linear mixed models (normal error structures). The prevalence of Vairimorpha spp. was analyzed using general linear mixed models with the error structures: [response variable = treatment * time + colony (random effect)]. Vairimorpha spp. intensity (log + 1 transformed) in Experiments 3 and 4 was analyzed using linear mixed models with the same error structures as prevalence. Tukey’s honest significant difference (HSD) was used on the models where multiple comparisons were made.
3. Results
3.1. Experiment 1: Laboratory Cage Study Evaluating Available Treatments against Vairimorpha ceranae
3.1.1. Consumption of Pollen, Sucrose Solution, and Water
On average, bees consumed 18.4 mL of sucrose syrup, 43.2 mL of water, and 3.9 g of pollen over the course of 28 days. There was no evidence that the consumption of pollen (F3,233 = 0.09; p = 0.967), sucrose syrup (F3,233 = 0.35; p = 0.791) or water (F3,233 = 0.48, p = 0.699) was significantly different between bees in the various treatment groups. Trace amounts of several pesticides were found in the wildflower pollen that was provided to bees in experimental cages (Supplementary Table S1).
3.1.2. V. ceranae Prevalence and Intensity
There was no evidence for an effect of an interaction between treatment and time on V. ceranae prevalence (χ2 = 3.33; df = 3; p = 0.344; Table 1) or for differences between treatments (χ2 = 5.2; df = 3; p = 0.158) or time (χ2 = 0.51; df = 1; p = 0.475). There was no evidence of significant differences in intensity of V. ceranae infection between the treatments (F3,34 = 0.66; p =0.586). The median V. ceranae infection intensity for Honey-B-Healthy® treatment was 2.8 × 106 spores/bee, while Nozevit Plus, Fumagilin-B, and negative control treatments had a median infection intensity of 2.22 × 106, 1.13 ×106, and 1.83 × 106 spores/bee, respectively. There was evidence that V. ceranae intensity increased over time (F3,36 = 4.21, p = 0.049), but there was no evidence of an interaction between treatment and time of sampling (F3,36 = 1.06, p = 0.38).
3.1.3. Survival Analysis
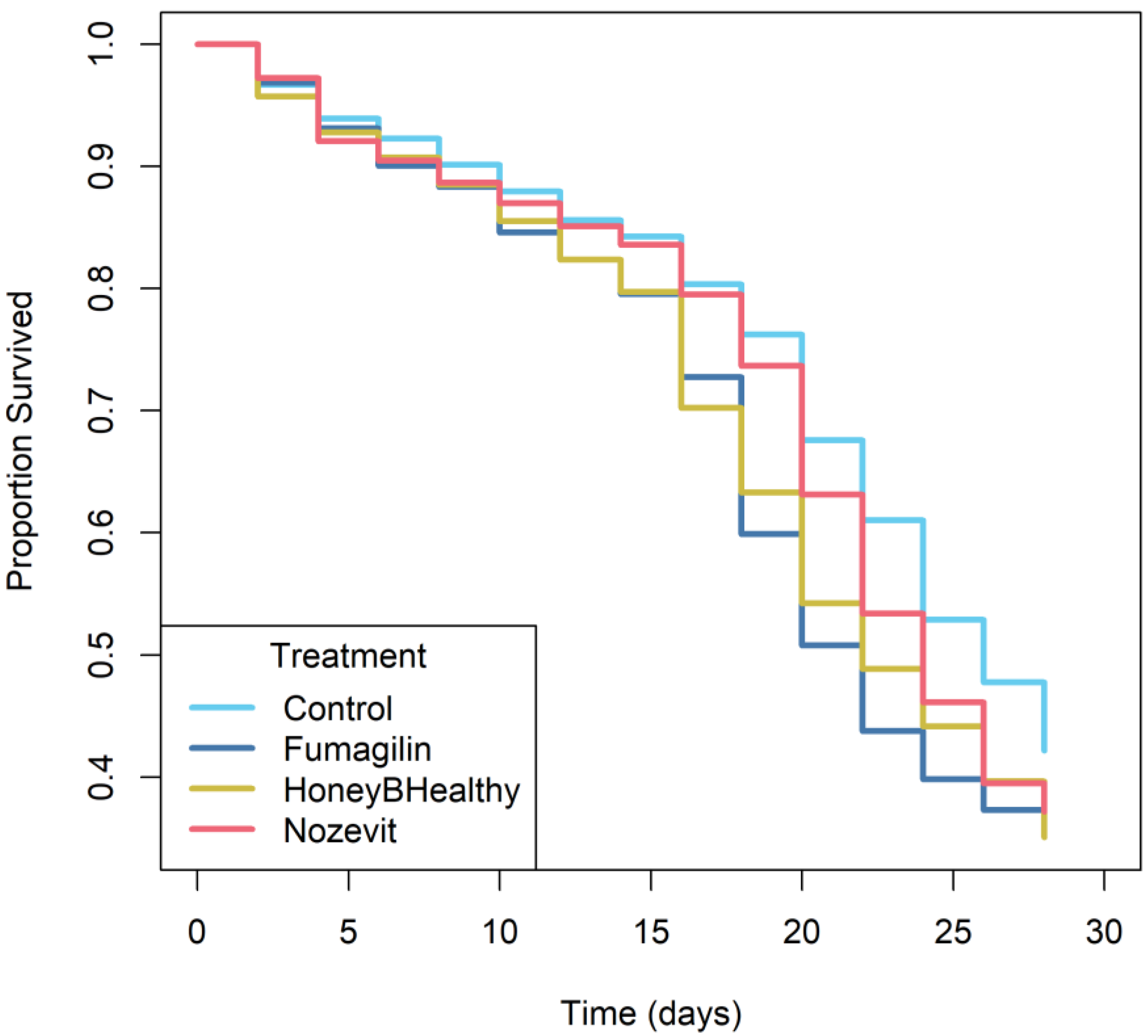
Kaplan–Meier survival curves were used to plot the survival data (Figure 1) and a Cox proportional hazards model was used to compare the survival curves of the various treatments. The Cox proportional hazards model indicated that there was evidence for reduction in survival among bees that were fed with all treatments compared to the negative control group (χ2 = 10.8; df = 3; p = 0.013). Kaplan–Meier curves showed that bees in the negative control group had the greatest survival, followed by that of bees in the Nozevit Plus and Honey-B-Healthy® group, and the Fumagilin-B treatment group (Figure 1).
3.2. Experiment 2: Field Experiment—HiveAlive® and Fumagilin-B
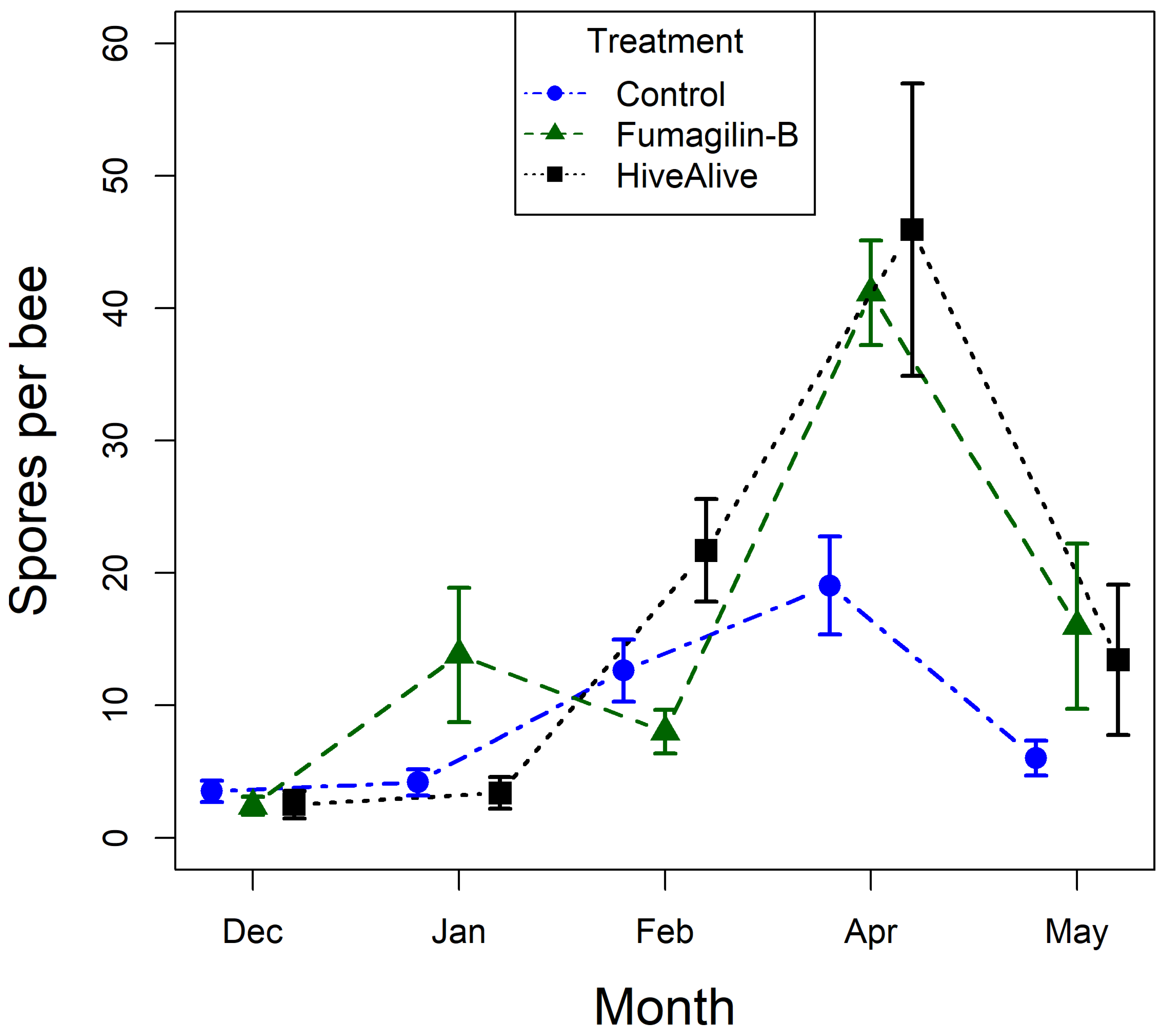
Overall, there was strong evidence that the number of V. destructor per 100 bees across all treatments decreased over the course of the experiment (χ24,439 = 42.19, p < 0.001), and there was an effect of treatment (χ22,439 = 7.87, p = 0.02), but there was no evidence of an interaction between treatment and time (χ28,439 = 5.49, p = 0.705) (Figure 2). Multiple comparisons showed that HiveAlive had a lower prevalence of V. destructor than controls, and Fumagilin-B was not different from either one.
Generally, Vairimorpha spp. intensity in bees increased over the course of the experiment, regardless of treatment, then decreased in the final month of the experiment. There was strong evidence of an interaction between treatment and time on Vairimorpha spp. intensity (F8,438 = 2.88, p = 0.004). In the multiple comparison breakdown, there was strong evidence of an increase in Vairimorpha spp. intensity in April for the bees fed the Fumagilin-B treatment (z = 2.72, p = 0.006) and the Hive Alive® treatment (z = 3.27, p = 0.001). In contrast, there was no evidence of a corresponding increase in intensity in bees in the negative control group (Figure 2).
3.3. Experiment 3: Winter Field Treatment using Fumagilin-B and Nosevit at Fall and Spring Recommended Treatments
At the end of the experiment, there was no evidence that colonies from the treatment groups differed in the observed number of bees (F4,39 = 0.32, p = 0.866) or brood (F4,39 = 0.73, p = 0.58) compared to those of the negative control.
At the end of the experiment, there was an average Vairimorpha spp. spore load of 516,666.67 spores per bee, with a prevalence of 0.84 across all treatments. There was no evidence that Vairimorpha spp. intensity was affected by treatment (F4,45 = 0.69, p = 0.604), but there was very strong evidence that Vairimorpha spp. intensity was affected by time (F3,131 = 24.39, p < 0.001). There was no evidence of an interaction between treatment and time on Vairimorpha spp. intensity (F12,127 = 1.22, p = 0.279). Similarly, there was very strong evidence that Vairimorpha spp. prevalence was only affected by the time variable (χ2 = 24.27, p < 0.001), with no differences between treatments (χ2 = 4.17, p = 0.384); Figure S1.
Reducing treatments to just the ingredient (negative control, Fumagilin-B, and Nozevit), regardless of solution preparation, showed weak evidence of an interaction between the treatment and time (F6,136 = 2.07, p = 0.056) on Vairimorpha spp. intensity. Again, there was strong evidence of an effect of time, where Vairimorpha spp. intensity dropped after the first week in all treatments (F3,136 = 10.79, p < 0.001). There was no effect of treatment (F2,46 = 1.39, p = 0.259).
3.4. Experiment 4: Repeat of Winter Field Treatment using Fumagilin-B and Nozevit at Fall and Spring Recommended Treatments
Overall, the number of observed bees and brood increased over time (bees: F1,93 = 125.05, p < 0.001; brood: F1,93 = 181.67, p < 0.001), but there was no evidence of an effect from treatments on these variables (bees: p = 0.279; brood: p = 0.564). For both bees and brood, there was no evidence of an interaction between treatment and time (bees: p = 0.257; brood: p = 0.584).
Across all treatments and observations, there was an average Vairimorpha spp. intensity of 673,500 spores per bee, with a prevalence of 0.8. There was no evidence of an effect of treatment (F2,195 = 5.8, p = 0.214) on Vairimorpha spp. intensity or the interaction between treatment and time (F12,195 = 14.32, p = 0.281), but very strong evidence that Vairimorpha spp. intensity was affected by time (F3,195 = 40.39, p < 0.001). Multiple comparisons showed an increase in Varirmorpha spp. intensity from week 1 to 2, then intensity decreased again after week 2. For Vairimorpha spp. prevalence, there was again no evidence for an effect of treatment χ2 = 7.33, p = 0.119) or the interaction between treatment and time (χ2 = 10.43, p = 0.578), but there was very strong evidence of an effect of time (χ2 = 26.17, p < 0.001).
4. Discussion
Many consider the control of pathogens such as Vairimorpha spp. essential for honey bee health [11] and important for reducing yearly honey bee colony losses. Our results provide weak or no evidence of any available registered or unregistered treatments to reduce Vairimorpha spp. prevalence and intensity. Laboratory exposure of bees to Vairimorpha spp. and subsequent treatment with various therapeutics showed no evidence of treatment effects on Vairimorpha spp. spore prevalence or intensity (Experiment 1). Subsequent field experiments showed increased Vairimorpha spp. intensity following exposure to HiveAlive® and Fumagilin-B (Experiment 2), weak short-term benefits of Fumagilin-B (Experiment 3), or no effects of treatments at all (Experiment 4). In all experiments, time was the principal factor that predicted Vairimorpha spp. infection.
Fumagilin-B, Honey-B-Healthy®, and Nozevit treatments reduced the survival of bees when compared to that of bees fed only sugar water (the negative control group). We speculate that, in a colony setting, bees would likely interact with treatments mixed into supplemental sugar syrup differently than they would in the laboratory cage setting. In cages, for instance, honey bees are forced to feed on the provided treatments (no choice scenario), thus maximizing the treatment received per bee. In the field, however, honey bees would have access to ample pollen, honey, and nectar stores, in addition to supplemental sucrose. Therefore, caged bees (in a no choice/forced feeding scenario) might be exposed to higher amounts of treatment than bees in the field colonies, which could lead to reductions in honey bee survival. As for Fumagilin-B specifically, [12] reviewed negative effects of the compound on honey bee health and found fumagillin, along with its often overlooked counterion (dicyclohexylamine) in salt form, to be toxic to honey bees. More recently, researchers have found significant reductions in mortality following treatment with dicyclohexylamine in isolation; this counterion is present in fumagillin-DCH salt and in Fumagilin-B [32].
In the first field experiment (Experiment 2), bees treated with Fumagilin-B and HiveAlive® showed increased Vairimorpha spp. intensity in April compared to that of bees in the negative control group. Previous studies have found that the death of beneficial gut bacteria resulting from exposure to antibiotics leads to increased vulnerability to Vairimorpha spp. infection and antibiotic resistant pathogens in honey bees [33,34,35]. Given that the gut microbial communities of honey bees have high specificity and stability [34], it could take time for populations of beneficial microbes to rebuild post treatment, allowing opportunistic parasites to become established in the meantime. Thus, if antibiotic treatments become necessary, subsequent probiotic treatments and protein nutrition may also be essential to prevent establishment of parasites e.g., [36,37,38]. We hypothesize that this disruption of gut microbial communities led to higher Vairimorpha spp. intensity in the treatment groups seen in this study.
The Vairimorpha species dominant in each experiment could influence the efficacy of the treatments used in these experiments. Vairimorpha ceranae and V. apis have been shown to differ in their respective biology and their impacts on honey bee colonies [10,39,40]. In Experiment 1, we confirmed V. ceranae as the dominant species of Vairimorpha via molecular detection. However, the species of Vairimorpha was not confirmed in Experiment 2. Nevertheless, this experiment was performed between late 2017 and early 2018. At that time, V. ceranae was the dominant species in honey bees throughout most of the world [40]. Experiments 3 and 4 were performed in early 2009 and from late 2009–early 2010, respectively. During these years, V. ceranae was present but likely not the dominant species. There may have been a mix of V. apis and V. ceranae infection in bees in these studies [10].
Vairimorpha apis infections in honey bees have a predictable phenology [40,41,42,43], where infections generally decrease in the summer and increase throughout fall and winter, with a peak in early spring [44,45]. This seasonality was particularly evident in Experiment 2, where a large peak was seen in April, particularly in colonies treated with Fumagilin-B and HiveAlive® (Figure 2). In contrast, V. ceranae has been shown to be present year-round [46,47] and have higher thermal tolerance than V. apis [48,49,50]. While V. ceranae appears not to have an easily predictable phenology, it can have periods of high infection followed by periods of low infection prevalence and intensity [51]. Thus, optimal management of honey bee colonies with the intention to outlast periods of high V. ceranae infection may be more effective than relying on treatments to reduce infection.
Proper nutrition appears to be effective for mitigating the effects of Vairimorpha spp. infection. In general, increased pollen consumption leads to improved outcomes following Vairimorpha spp. infection [19,20,52,53,54]. Despite this, it is unclear if pollen substitutes are as effective as natural pollen consumption by honey bees for the reduction of spore loads [reviewed in 54]. In periods of pollen dearth, certain pollen substitutes would likely improve outcomes for bees with Vairimorpha spp. infection [55,56], but might also lead to higher Vairimorpha spp. intensities. It is difficult to recommend specific pollen substitutes since the ingredients used and the formulation processes in the different products are highly variable.
Although fumagillin and other treatments designed to reduce Vairimorpha spp. infections have been shown to be effective in certain circumstances, there are many factors that can affect colonies’ responsiveness to treatments. Based on the findings described here, it is not feasible to recommend one specific treatment over another for Vairimorpha spp. control; rather, we recommend improving colony health and reducing pest pressure to ensure colonies are not afflicted with multiple stressors. Future research should focus on examining timing of treatments with predicted Vairimorpha spp. peaks. For this research, sampling for Vairimorpha spp. could be conducted throughout the year to predict when peaks would occur, as geography and climate play a significant role. Then, treatments could be applied the following year after understanding the phenology of Vairimorpha spp. Additionally, future research might further explore how certain products work on a preventative basis rather than a treatment basis, such as the use of propolis as a preventative treatment for Vairimorpha spp. infections [57]. Other preventative treatments that take Vairimorpha spp. seasonality into account could also prove to be effective. Such information would be valuable for both researchers and beekeepers.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app13031303/s1, Figure S1: Number of spores per bee in Experiment 3; Table S1: Pesticide results.
Author Contributions
Conceptualization, C.P., C.J., R.S. and J.D.E.; Methodology, C.J., R.S. and J.D.E.; Formal analysis, C.P.; Investigation, C.J.; Resources R.S. and J.D.E.; Data curation, C.P., C.J. and J.D.E.; Writing—original draft, C.P.; Writing—review & editing, C.P., C.J., R.S. and J.D.E.; Visualization, C.P.; Supervision, C.J., R.S. and J.D.E.; Project administration, J.D.E.; Funding acquisition, C.J., R.S. and J.D.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the USDA National Institute of Food and Agriculture Multistate Project (1005822), the Florida State Beekeepers Association and the Florida Department of Agriculture and Consumer Services. The funding sources had no involvement in any aspects of the study.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data in this manuscript are available in a public repository at: https://doi.org/10.5281/zenodo.7534789.
Acknowledgments
We gratefully thank Straughn Farms and Billy Rhodes for providing many experimental honey bee colonies during our field experiments. Additionally, we thank Walter Taylor, Jon Elmquist, Branden Stanford, and Michelle Weschler for their help sampling experimental honey bee colonies and counting Vairimorpha spp. spores.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boncristiani, H.; Ellis, J.D.; Bustamante, T.; Graham, J.; Jack, C.; Kimmel, C.B.; Mortensen, A.; Schmehl, D.R. World honey bee health: The global distribution of western honey bee (Apis mellifera L.) pests and pathogens. Bee World 2020, 98, 2–6. [Google Scholar] [CrossRef]
- Tokarev, Y.S.; Huang, W.-F.; Solter, L.F.; Malysh, J.M.; Becnel, J.J.; Vossbrinck, C.R. A formal redefinition of the genera Nosema and Vairimorpha (Microsporidia: Nosematidae) and reassignment of species based on molecular phylogenetics. J. Invertebr. Pathol. 2020, 169, 107279. [Google Scholar] [CrossRef]
- Genersch, E. Honey bee pathology: Current threats to honey bees and beekeeping. Appl. Microbiol. Biotechnol. 2010, 87, 87–97. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Sinpoo, C.; Paxton, R.J.; Disayathanoowat, T.; Krongdang, S.; Chantawannakul, P. Impact of Nosema ceranae and Nosema apis on individual worker bees of the two host species (Apis cerana and Apis mellifera) and regulation of host immune response. J. Insect Physiol. 2018, 105, 1–8. [Google Scholar] [CrossRef]
- Paris, L.; El Alaoui, H.; Delbac, F.; Diogon, M. Effects of the gut parasite Nosema ceranae on honey bee physiology and behavior. Curr. Opin. Insect Sci. 2018, 26, 149–154. [Google Scholar] [CrossRef]
- Higes, M.; Martín, R.; Meana, A. Nosema ceranae, a new microsporidian parasite in honeybees in Europe. J. Invertebr. Pathol. 2006, 92, 93–95. [Google Scholar] [CrossRef]
- Huang, W.-F.; Jiang, J.-H.; Chen, Y.-W.; Wang, C.-H. A Nosema ceranae isolate from the honeybee Apis mellifera. Apidologie 2007, 38, 30–37. [Google Scholar] [CrossRef]
- Chemurot, M.; De Smet, L.; Brunain, M.; De Rycke, R.; de Graaf, D.C. Nosema neumanni n. Sp. (Microsporidia, Nosematidae), a new microsporidian parasite of honeybees, Apis mellifera in Uganda. Eur. J. Protistol. 2017, 61, 13–19. [Google Scholar] [CrossRef]
- Paxton, R.J.; Klee, J.; Korpela, S.; Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 2007, 38, 558–565. [Google Scholar] [CrossRef]
- Burnham, A.J. Scientific advances in controlling Nosema ceranae (Microsporidia) infections in honey bees (Apis mellifera). Front. Vet. Sci. 2019, 6, 79. [Google Scholar] [CrossRef] [Green Version]
- van den Heever, J.P.; Thompson, T.S.; Curtis, J.M.; Ibrahim, A.; Pernal, S.F. Fumagillin: An overview of recent scientific advances and their significance for apiculture. J. Agric. Food Chem. 2014, 62, 2728–2737. [Google Scholar] [CrossRef] [PubMed]
- Webster, T.C. Fumagillin affects Nosema apis and honey bees (Hymonopterai Apidae). J. Econ. Entomol. 1994, 87, 601–604. [Google Scholar] [CrossRef]
- Goodman, M.; Houten, A.T.; Perry, J.; Blackmann, R. Cost benefit analysis of using fumagillin to treat nosema. N. Z. Beekeep. 1990, 208, 11–12. [Google Scholar]
- Williams, G.R.; Sampson, M.A.; Shutler, D.; Rogers, R.E. Does fumagillin control the recently detected invasive parasite Nosema ceranae in western honey bees (Apis mellifera)? J. Invertebr. Pathol. 2008, 99, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-F.; Solter, L.F.; Yau, P.M.; Imai, B.S. Nosema ceranae escapes fumagillin control in honey bees. PLoS Pathog. 2013, 9, e1003185. [Google Scholar] [CrossRef] [Green Version]
- Holt, H.L.; Grozinger, C.M. Approaches and challenges to managing Nosema (Microspora: Nosematidae) parasites in honey bee (Hymenoptera: Apidae) colonies. J. Econ. Entomol. 2016, 109, 1487–1503. [Google Scholar] [CrossRef]
- Bartlett, L.J. Frontiers in effective control of problem parasites in beekeeping. Int. J. Parasitol. Parasites Wildl. 2022, 17, 263–272. [Google Scholar] [CrossRef]
- Jack, C.J.; Uppala, S.S.; Lucas, H.M.; Sagili, R.R. Effects of pollen dilution on infection of Nosema ceranae in honey bees. J. Insect Physiol. 2016, 87, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, A.G.; Toth, A.L. Feedbacks between nutrition and disease in honey bee health. Curr. Opin. Insect Sci. 2018, 26, 114–119. [Google Scholar] [CrossRef]
- Castelli, L.; Branchiccela, B.; Garrido, M.; Invernizzi, C.; Porrini, M.; Romero, H.; Santos, E.; Zunino, P.; Antúnez, K. Impact of nutritional stress on honeybee gut microbiota, immunity, and Nosema ceranae infection. Microb. Ecol. 2020, 80, 908–919. [Google Scholar] [CrossRef]
- Botías, C.; Martín-Hernández, R.; Meana, A.; Higes, M. Screening alternative therapies to control Nosemosis type C in honey bee (Apis mellifera iberiensis) colonies. Res. Vet. Sci. 2013, 95, 1041–1045. [Google Scholar] [CrossRef]
- Charistos, L.; Parashos, N.; Hatjina, F. Long term effects of a food supplement HiveAliveTM on honey bee colony strength and Nosema ceranae spore counts. J. Apic. Res. 2015, 54, 420–426. [Google Scholar] [CrossRef]
- Fries, I.; Chauzat, M.P.; Chen, Y.P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; Mcmahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standard methods for Nosema research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Hamiduzzaman, M.M.; Guzman-Novoa, E.; Goodwin, P.H. A multiplex PCR assay to diagnose and quantify Nosema infections in honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 105, 151–155. [Google Scholar] [CrossRef]
- Cantwell, G.E. Standard methods for counting Nosema spores. Amer. Bee J. 1970, 110, 222–223. [Google Scholar]
- Jack, C.J.; Lucas, H.M.; Webster, T.C.; Sagili, R.R. Colony level prevalence and intensity of Nosema ceranae in honey bees (Apis mellifera L.). PLoS ONE 2016, 11, e0163522. [Google Scholar] [CrossRef] [Green Version]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B. Standard methods for varroa research. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Delaplane, K.S.; Van Der Steen, J.; Guzman-Novoa, E. Standard methods for estimating strength parameters of APIS mellifera colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Bates, D.; Kliegl, R.; Vasishth, S.; Baayen, H. Parsimonious mixed models. arXiv 2015, arXiv:1506.04967. [Google Scholar]
- Therneau, T.M.; Lumley, T. Package ‘survival’. R Top Doc 2015, 128, 28–33. [Google Scholar]
- van den Heever, J.P.; Thompson, T.S.; Otto, S.J.; Curtis, J.M.; Ibrahim, A.; Pernal, S.F. The effect of dicyclohexylamine and fumagillin on Nosema ceranae-infected honey bee (Apis mellifera) mortality in cage trial assays. Apidologie 2016, 47, 663–670. [Google Scholar] [CrossRef]
- Li, J.H.; Evans, J.D.; Li, W.F.; Zhao, Y.Z.; DeGrandi-Hoffman, G.; Huang, S.K.; Li, Z.G.; Hamilton, M.; Chen, Y.P. New evidence showing that the destruction of gut bacteria by antibiotic treatment could increase the honey bee’s vulnerability to Nosema infection. PLoS ONE 2017, 12, e0187505. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Fadhil, N.H.; Powell, J.E.; Kwong, W.K.; Moran, N.A. Long-term exposure to antibiotics has caused accumulation of resistance determinants in the gut microbiota of honeybees. MBio 2012, 3, e00377-12. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D.; Lopez, D.L. Bacterial probiotics induce an immune response in the honey bee (Hymenoptera: Apidae). J. Econ. Entomol. 2004, 97, 752–756. [Google Scholar] [CrossRef]
- Audisio, M.C. Gram-positive bacteria with probiotic potential for the Apis mellifera L. honey bee: The experience in the northwest of Argentina. Probiotics Antimicrob. Proteins 2017, 9, 22–31. [Google Scholar] [CrossRef]
- Daisley, B.A.; Pitek, A.P.; Chmiel, J.A.; Al, K.F.; Chernyshova, A.M.; Faragalla, K.M.; Burton, J.P.; Thompson, G.J.; Reid, G. Novel probiotic approach to counter Paenibacillus larvae infection in honey bees. ISME J. 2020, 14, 476–491. [Google Scholar] [CrossRef] [Green Version]
- Milbrath, M.O.; van Tran, T.; Huang, W.-F.; Solter, L.F.; Tarpy, D.R.; Lawrence, F.; Huang, Z.Y. Comparative virulence and competition between Nosema apis and Nosema ceranae in honey bees (Apis mellifera). J. Invertebr. Pathol. 2015, 125, 9–15. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Bartolomé, C.; Chejanovsky, N.; Le Conte, Y.; Dalmon, A.; Dussaubat, C.; García-Palencia, P.; Meana, A.; Pinto, M.A.; Soroker, V.; et al. Nosema ceranae in Apis mellifera: A 12 years postdetection perspective. Environ. Microbiol. 2018, 20, 1302–1329. [Google Scholar] [CrossRef] [Green Version]
- Gisder, S.; Hedtke, K.; Möckel, N.; Frielitz, M.-C.; Linde, A.; Genersch, E. Five-year cohort study of Nosema spp. in Germany: Does climate shape virulence and assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 2010, 76, 3032–3038. [Google Scholar] [CrossRef] [Green Version]
- Traver, B.E.; Fell, R.D. Prevalence and infection intensity of Nosema in honey bee (Apis mellifera L.) colonies in Virginia. J. Invertebr. Pathol. 2011, 107, 43–49. [Google Scholar] [CrossRef]
- Traver, B.E.; Williams, M.R.; Fell, R.D. Comparison of within hive sampling and seasonal activity of Nosema ceranae in honey bee colonies. J. Invertebr. Pathol. 2012, 109, 187–193. [Google Scholar] [CrossRef]
- Bailey, L. The epidemiology and control of Nosema disease of the honey-bee. Ann. Appl. Biol. 1955, 43, 379–389. [Google Scholar] [CrossRef]
- Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, S73–S79. [Google Scholar] [CrossRef]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; Del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Botías, C.; Bailón, E.G.; Martínez-Salvador, A.; Prieto, L.; Meana, A.; Higes, M. Microsporidia infecting Apis mellifera: Coexistence or competition. Is Nosema ceranae replacing Nosema apis? Environ. Microbiol. 2012, 14, 2127–2138. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Meana, A.; García-Palencia, P.; Marín, P.; Botías, C.; Garrido-Bailón, E.; Barrios, L.; Higes, M. Effect of temperature on the biotic potential of honeybee microsporidia. Appl. Environ. Microbiol. 2009, 75, 2554–2557. [Google Scholar] [CrossRef] [Green Version]
- Higes, M.; García-Palencia, P.; Botías, C.; Meana, A.; Martín-Hernández, R. The differential development of microsporidia infecting worker honey bee (Apis mellifera) at increasing incubation temperature. Environ. Microbiol. Rep. 2010, 2, 745–748. [Google Scholar] [CrossRef]
- Gisder, S.; Schueler, V.; Horchler, L.L.; Groth, D.; Genersch, E. Long-term temporal trends of Nosema spp. infection prevalence in Northeast Germany: Continuous spread of Nosema ceranae, an emerging pathogen of honey bees (Apis mellifera), but no general replacement of Nosema apis. Front. Cell. Infect. Microbiol. 2017, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- Copley, T.R.; Chen, H.; Giovenazzo, P.; Houle, E.; Jabaji, S.H. Prevalence and seasonality of Nosema species in Québec honey bees. Can. Entomol. 2012, 144, 577–588. [Google Scholar] [CrossRef]
- Azzouz-Olden, F.; Hunt, A.; DeGrandi-Hoffman, G. Transcriptional response of honey bee (Apis mellifera) to differential nutritional status and Nosema infection. BMC Genom. 2018, 19, 1–20. [Google Scholar] [CrossRef] [PubMed]
- de Jong, E.W.; DeGrandi-Hoffman, G.; Chen, Y.; Graham, H.; Ziolkowski, N. Effects of diets containing different concentrations of pollen and pollen substitutes on physiology, Nosema burden, and virus titers in the honey bee (Apis mellifera L.). Apidologie 2019, 50, 845–858. [Google Scholar] [CrossRef]
- Noordyke, E.R.; Ellis, J.D. Reviewing the efficacy of pollen substitutes as a management tool for improving the health and productivity of western honey bee (Apis mellifera) colonies. Front. Sustain. Food Syst. 2021, 437, 772897. [Google Scholar] [CrossRef]
- Fleming, J.C.; Schmehl, D.R.; Ellis, J.D. Characterizing the impact of commercial pollen substitute diets on the level of Nosema spp. In honey bees (Apis mellifera L.). PLoS ONE 2015, 10, e0132014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, A.N.; Jack, C.J.; Bustamante, T.A.; Schmehl, D.R.; Ellis, J.D. Effects of supplemental pollen feeding on honey bee (Hymenoptera: Apidae) colony strength and Nosema spp. Infection. J. Econ. Entomol. 2019, 112, 60–66. [Google Scholar] [CrossRef]
- Naree, S.; Ellis, J.D.; Benbow, M.E.; Suwannapong, G. The use of propolis for preventing and treating Nosema ceranae infection in western honey bee (Apis mellifera Linnaeus, 1787) workers. J. Apic. Res. 2021, 60, 686–696. [Google Scholar] [CrossRef]
Figure 1.
Proportion of honey bees that survived Vairimorpha ceranae infection and subsequent treatment in Experiment 1. Treatments are negative control (light blue), Fumagilin (dark blue), Honey-B-Healthy (yellow), and Nozevit Plus (red).
Figure 1.
Proportion of honey bees that survived Vairimorpha ceranae infection and subsequent treatment in Experiment 1. Treatments are negative control (light blue), Fumagilin (dark blue), Honey-B-Healthy (yellow), and Nozevit Plus (red).

Figure 2.
Vairimorpha spp. spores per bee (×50,000) in Experiment 2. Shapes and error bars represent mean and standard error. Treatments are: control (blue circles), Fumagilin-B® (green triangles), and Hive Alive (black squares).
Figure 2.
Vairimorpha spp. spores per bee (×50,000) in Experiment 2. Shapes and error bars represent mean and standard error. Treatments are: control (blue circles), Fumagilin-B® (green triangles), and Hive Alive (black squares).


{kind=link}
{kind=link}
Table 1.
Experiment 1: Prevalence (percent of infected individuals) of Vairimorpha spp. in honey bees provided various treatments. The sampling occurred on two days (16 and 23) post treatment. Prevalence means with different letters are significantly different at α ≤ 0.05.
Table 1.
Experiment 1: Prevalence (percent of infected individuals) of Vairimorpha spp. in honey bees provided various treatments. The sampling occurred on two days (16 and 23) post treatment. Prevalence means with different letters are significantly different at α ≤ 0.05.
| Treatment | Sampling Day | Prevalence (%) |
|---|---|---|
| Control | 16 | 2.4 a |
| Fumagilin-B | 16 | 2.4 a |
| Honey-B-Healthy® | 16 | 7.2 a |
| Nozevit Plus | 16 | 6.4 a |
| Control | 23 | 4.0 a |
| Fumagilin-B | 23 | 2.4 a |
| Honey-B-Healthy® | 23 | 2.4 a |
| Nozevit Plus | 23 | 3.2 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Prouty, C.; Jack, C.; Sagili, R.; Ellis, J.D. Evaluating the Efficacy of Common Treatments Used for Vairimorpha (Nosema) spp. Control. Appl. Sci. 2023, 13, 1303. https://doi.org/10.3390/app13031303
AMA Style
Prouty C, Jack C, Sagili R, Ellis JD. Evaluating the Efficacy of Common Treatments Used for Vairimorpha (Nosema) spp. Control. Applied Sciences. 2023; 13(3):1303. https://doi.org/10.3390/app13031303
Chicago/Turabian StyleProuty, Cody, Cameron Jack, Ramesh Sagili, and James D. Ellis. 2023. "Evaluating the Efficacy of Common Treatments Used for Vairimorpha (Nosema) spp. Control" Applied Sciences 13, no. 3: 1303. https://doi.org/10.3390/app13031303
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.