
Augmented Retting Effect on Kenaf Fibers Using Alkalophilic Pectinase-Producing Bacteria in Combination with Water Solvents



1
Biotechnology Research Institute, Universiti Malaysia Sabah, Jalan UMS, Kota Kinabalu 88400, Malaysia
2
Farm Management Unit (FMU), Bangladesh Jute Research Institute, Manik Mian Avenue, Dhaka 1207, Bangladesh
*
Author to whom correspondence should be addressed.
Appl. Sci. 2022, 12(14), 7136; https://doi.org/10.3390/app12147136
Submission received: 1 June 2022
/
Revised: 4 July 2022
/
Accepted: 12 July 2022
/
Published: 15 July 2022
(This article belongs to the Section Chemical and Molecular Sciences)
Abstract
:A degumming approach is used in this paper with alkalophilic pectinase-producing bacteria (APPB) and two sources of water solvents to address the existing conventional water retting complexities of kenaf. The incorporation of APPB was confirmed based on their retting feasibilities and multiple cell-wall-degrading enzymatic delicacy. The combinations of APPB with seawater offered retting achievements within six-day retting in non-sterile conditions. These retting niches showed maximum (14.67 U/mL) pectinase activity with fiber separation feasibilities of 4.75 Fried test score. The yielded fiber composition analysis showed a higher cellulose composition (84.65%) and the least amount of hemicellulose, pectin, and ligneous gummy substances. The transmission electron microscopy scan of the yielded fibers showed smooth fiber surfaces, 84.20 µm fiber diameter, and 7.65 g/tex fine fiber compared with uninoculated and combinations of freshwater treatments. The FTIR spectra revealed the cellulosic discrepancies of the retting treatments by monitoring O-H and C=O stretching at ~3300 cm−1 and ~1730 cm−1 wavenumbers. These findings are compelling to yield kenaf fibers of quality considering the existing retting difficulties.
1. Introduction
Natural cellulosic bast fibers (NCBFs) are essential raw materials for the manufacturing of numerous commodities. The utilization of NCBFs has increased because of their biodegradability, affordability, and replenishing features, which are not offered by petroleum-based synthetic materials. Kenaf (Hibiscus cannabinus) is one of many commercial NCBF-producing plants and has great potential for industrial cellulosic textile fiber production. Due to its numerous positive aspects, it is considered the most promising field crop adapted to many ago-climatic conditions [1,2]. Similar to other dicotyledonous specimens, its cellulosic bast fibers inherently bond with hemicellulose, pectin, and ligneous non-cellulosic gums (NCGs) onto its cell walls’ matrix. The plants are required to undergo the obligatory degumming process, which facilitates the fiber separation and ascertains its qualities. The process confers benefits for several downstream biomaterials’ preparations [3,4].
Kenaf could be degummed through several processes, and conventional water retting (CWR) is widely used. It is achieved by the combined functions of retting solvents and microbial enzymatic activities. The mature harvested plants are steeped in natural freshwater bodies to depolymerize pectins and other non-cellulosic gummy substances as they could separate the fibers. The role of water as a retting solvent is specific to maceration, providing appropriate nutrients and media for the microbes’ sustenance and creating pathways to invade the cell walls. The process is favored over other degumming processes for managing a vast volume of plant materials at a low cost and without chemical treatments. It does not alter the basic fiber structures and can yield fibers with less or minimum processing during the product manufacturing steps [3,5]. Despite these feasibilities, the process is challenged by an inability to yield consistent quality fibers, freshwater scarcity, and eutrophication-related complexities. The process requires 20 to 25 volumes of water to biomass; typically, 10,000 kg of plant substrate requires approximately 432 m3 of water. Currently, this water is obtained from freshwater sources and after retting, it is discharged into aquatic ecosystems [6,7,8]. The scarcity of natural freshwater bodies for retting solvents severely affects the sustainability of its degumming process. Due to socio-economic and agro-climatic changes, the consequence of this scarcity tends to worsen. In addition, eutrophication and pungent odors are associated with this process as more than thirty types of plant organic compounds are directly dispersed to aquatic ecosystems [9,10,11]. The scarcity of freshwater retting niches leads to placing huge bast material in the same niche or compensating by reducing the cultivation portfolio. The used effluents accelerate oxygen depletion and cause threats to aquatic flora and fauna. The scope of other sources of water, such as seawater, as retting solvents have been demonstrated to be as viable as freshwater [12,13]. However, the inoculations of specific microbes into seawater to enhance the degumming process have not been reported.
Kenaf fibers are incompatible in several value-added commodity preparations since it has been challenging to obtain consistent quality fibers through the CWR process. This inconsistence in fiber quality is associated with the low metabolic activity of the microbial mixture. Such co-existence of microbial compositions releases intermediate-level enzymes, which linger in the process and yield inconsistent quality fibers [14,15]. In contrast, inadequate degumming affects the overall chemical compositions, and yields low-quality fibers that do not possess adequate economic value [16,17].
Water retting is primarily a macero-microbial process, predominantly accomplished with specific bacterial enzymes and solvent-derived maceration [18,19,20,21]. Naturally occurring microbiota produce the desired enzymes, and the rest act as non-producers [22]. In the last few decades, several studies have tried to incorporate bacterial enzymes or direct inoculum applications, including Bacillus, Enterobacteriaceae, and Pectobacterium, or consortia to leverage the CWR [18,23,24,25,26]. However, non-sterile niches confer a poorer degumming efficiency. Moreover, the sky-high enzyme costs hinder their application at the growers’ scale. Alkalophilic bacteria have been used in many biotechnological applications for their unique adaptability and metabolites. The utilizations of alkalophilic bacteria in kenaf degumming could serve as potent biological agents for the biodegradation of acidic pectin polysaccharides. They have been previously used for wastewater treatments, dye decolorization, and the degumming of ramie [27,28,29].
The scope of using APPB in combination with seawater as a kenaf retting agent remains unverified. Thus, this study aims to explore a bacterial class that can enhance the kenaf bio-degumming process in combination with retting solvents. Hence, the augmented bio-retting of kenaf was adopted in combination with isolated alkaliphilic pectinolytic bacteria and two sources water solvents. The feasibility would be comprehended by the fibers’ separation efficacy and fiber quality parameters. It was hypothesized that APPB with water solvents combinedly act as drivers of the kenaf retting process to obtain quality fibers. The findings are pertinent for selecting the specific class of bacteria with compatible retting solvents to face the existing kenaf degumming difficulties.
2. Materials and Methods
2.1. Obtaining Alkalophilic Pectinase-Producing Bacteria
The bacterial isolates APPB1 and APPB2 used in the augmented kenaf retting were isolated from wet paddy land sediments and kenaf retting niches. The strains were optimally grown in alkaline (~pH 9.0) conditions on Horikoshi (H1) media with 1% Na2CO3 solutions. The pectinolytic potency was confirmed by observing the hydrolysis zone on the yeast extract pectin (YEP) medium. In addition, polysaccharide-degrading enzymatic potency, i.e., pectinase, xylanase, and cellulase, were measured by the dinitro salicylic acid reagent (DNS) enzyme assay. The enzyme unit (U) was defined, which catalyzes the formation of one μmol of reducing sugar per minute under the specified assay conditions. The morpho-phylogy of APPB was confirmed using Berge’s Manual of Systematic Bacteriology as a reference [30]. Cell morphology was observed under a light stereomicroscope and scanning electron microscopic (Hitachi S3400-N, Tokyo, Japan) technology.
2.2. Experimental Procedure
Based on enzymatic and in situ degumming efficacies (data were not shown), the two superlatives’ alkalophilic pectinase-producing bacterial isolates (APPB1 and APPB2) were selected to test with six degumming treatments for the on-sterile retting (Table 1). The plant materials of kenaf (KE-3) and retting solvents used in this study were collected from local sources. The augmented system was integrated with a polyvinyl tank using freshly grown materials desized to ~150 cm long, with the central parts maintaining a 1:15 ratio of substrates to solvents. The processing parameters, including bath ratio, degumming period, and bacterial CFU/mL, were maintained based on in situ determinations, following the isolates’ maximum concentrations at the log phase for the selected APPB before application. The bacterial inoculum of both isolates grew for 16 h in the yeast extract pectin liquid media and was eventually used by maintaining 3.5 × 108 CFU/mL densities. In each tank, 1.5 L of inoculum was added to 15 L of water solvents for steeping one kg of plant materials of the respective isolates. During the retting treatments, the average daytime temperature prevailed at 29 °C with 5°C fluctuations at night, and the average humidity was 84%. The pH of the freshwater and seawater retting solvents were 7.2 and 8.1, respectively.
2.3. Determination of the Enzyme Activity
The quantitative pectinase, xylanase, and cellulase enzymatic activities of the isolates were determined by the dinitro salicylic acid reagent (DNS) enzyme assay. The method was adopted based on the reducing sugars produced for the enzymatic hydrolysis of the respective substrates, such as apple pectin, birchwood xylan, and carboxy methyl cellulose. The reducing sugar concentrations were estimated as D-galacturonic acid, D-xylose, and D-glucose equivalent standard curves by measuring the absorbable concentrations as followed by [31,32]. The pectinase enzyme activity of the retting effluents was estimated by using the method described by [33]. About 10 mL of retting effluent was treated with an equal volume of NaOH (100 °C for 5 min) to remove ammonia, followed by centrifugation (10,000× g for 15 min at 4 °C) and filtration with a 0.22-micron filter. A 2 mL of filtrate was added to an equal volume of 0.5% apple pectin solution maintained with a glycine–NaOH- buffer of pH 10. Then, the mixture was incubated at 45 °C for 30 min. About 1.5 mL of dinitro salicylic (DNS) reagents was added and heated at 100 °C for 15 min. Then, the mixture was diluted into a 25 mL ddH2O. The mixture’s absorbance at 530 nm was measured using a spectrophotometer to determine the amount of reducing sugars; D-galacturonic acid as standard and ddH2O were used as the reagent blank. Each enzyme activity unit was defined as the amount of enzyme that liberates one micromole of galacturonic acid per minute under assay conditions.
2.4. Fried Score Test
The degree of retting was determined by the modified Fried test, as described by [34]. The empirical test score level 0 to 6 was used to designate the bast fiber retting efficacy. After the retting treatment, the individual kenaf stems were cut into approximately 10 cm sections. Then, the portions were placed in a ten mL boiling water to a test tube with four stems pieces. The retted stems bearing tubes were wrapped and promptly agitated for exactly 10 s on a table-top vortex mixer and, thereafter, hand shaken forcefully four times in a vertical direction. Samples in the vials were numerically scored based on fiber release.
2.5. Physical Fibers’ Quality
The longitudinal features and surface morphology of the fibers were investigated using Hitachi S3400-N scanning electronic microscopic technology. Kenaf fiber physical quality, including the presence and absence of gums on the fiber surfaces, fiber bundle diameter, and fineness, was determined. Fiber Fineness tex (equal to g/1000 m of fiber length) was determined using Equation (1) as described by [35], following the substance density of natural fibers that amount to 1.45 g/cm3 [36,37].
where f = fineness (tex); d = diameter (mm); and ρ = substance density (1450 kg/m3).
2.6. Fiber Composition Analysis
The α-cellulose and hemicellulose were measured according to [38], with slight modifications, and acid soluble lignin was determined by the method described by [39]. The pectin content was determined by taking 50 g of constant dried grounded fibers; after that, it was sequentially washed and filtrated three times with 0.01 N, 0.05 N, and 0.3 N HCl. Then, the pooled samples made 500 mL; from it, 100–200 mL of aliquots were taken and 250 mL deionized water was added, neutralized with 1 N NaOH, and an additional 10 mL of 1 N NaOH, which stood overnight, was added. After that, 50 mL of 1 N acetic acid was added and 5 min later 25 mL of 1 N CaCl2 solution was added with constant stirring and allowed to stand for one hour. After boiling, it was filtrated through free weighed Whatman No. 1 filter paper with G3 glass suction. The precipitate was subsequently washed several times with near-boiling water to free it from chloride. The filter paper with Ca–pectate was then dried at 80 °C for 5 h and weighed for the estimation of pectin as Ca-pectate, following Equation (2).
2.7. FTIR Spectra Analysis
The FTIR technique was employed to monitor the spectral structure (peak shape, area, and positions) of the individual chemical groups of the yielded fibers compared with the unretted raw kenaf fiber [40]. The crushed samples were mixed well with potassium bromide and made in the form of thin pellets. The scanning was performed in the range from 4000 to 400 cm−1 at a resolution of 4 cm−1. The spectrum was captured at a rate of 32 scans per min. The FTIR spectra of the treated raw kenaf fibers were investigated on a Cary 63076 Spectrophotometer (Agilent Technologies, Santa Clara, CA, USA) and subsequently compared with unretted raw kenaf bast fibers.
3. Results and Discussion
3.1. Alakalophilic Pectinase-Producing Bacteria
The pure cultured alkali-tolerant bacterial colonies APPB1 and APPB2 were isolated from wet paddy land soil and kenaf retting niche. Both isolates were able to grow on an adapted alkaline (pH 9.0) nutrient agar medium. Screening by the plate assay method revealed that the cultured bacteria were able to grow in alkaline in pH media with 1% Na2CO3 solution. According to [41], bacteria and other microbes that are optimally grown at pH 9.0 or above are considered alkalophilic bacteria. These isolates did not necessarily require obligate alkaliphile; however, elucidation at multiple pH conditions showed optimal growth at an elevated pH (~9). Once they grew on H1 media, a lot of colonies were visible on the plates and countable numbers of colonies (30–300 cfu/mL) were observed in the diluted samples.
Alkalophilic bacteria can survive in harsh environmental conditions and maintain cellular homeostasis due to their proton-motive force for ATP synthesis [42]. Along with extreme alkaline niches, alkaliphilic bacteria are abundant in elevated pH niches, such as serpentinizing sites and wet paddy soil, where continuous bio-recycling occurred. However, the core task in advancing the retting process is to obtain strains capable of producing pectinase enzymes. The pectinolytic potency of the isolates was confirmed by their pectin hydrolysis potency in a solid pectin agar medium (Figure 1). The screening results show that both isolates were able to hydrolyze pectin; therefore, these isolates were designated as alkalophilic pectinase-producing bacteria (APPB). Even though multiple enzymes are required to degrade heterogeneous cell wall components, pectinase is the main retting enzyme determining the retting magnitude, and applications of high efficiency pectinolytic bacterial strains are advantageous in decreasing the retting period, thus enhancing fiber quality [25,43].
The quantitative multiple enzymatic activities of these isolates are presented in Figure 2. The majority of naturally occurring microorganisms do not produce enzymes in adequate levels for industrial use, and many of them lack the desired characteristics for applications. Hence, both the stains showed their enzymatic potency to be used as retting agents and are comparable with other reported results [25,44]. In this paper, the xylano-pectinolytic activity of both isolates was associated with cellulase activity (Figure 2).
The xylano-pectinolytic bacterial strains deprived of cellulase activities were considered critical for the degumming process. On the contrary, Zhang et al. [13] stated that, in addition to pectinases, hemicellulases and cellulases are also required to obtain good retting results. In general, cellulase are not preferred for degumming as those preferentially attack the fiber nodes or kink bands and weaken the fibers. Even so, microbial pectinases are well-mixed with cellulases and hemicellulases activities [15,45].
The bacterial extracellular enzymatic activity was considered the critical biocatalyst in the degradation and utilization of organic polymers. In this paper, the enzymatic activities of the isolates were estimated using the extracellular crude enzymes of all three enzymatic activities, which is consistent with the findings of other studies, when compared with the crude supernatant enzyme assay. The findings of the crude enzymatic activities of both isolates are consistent with the results of previous studies, such as those reported by [25,44,46] for Bacilillus spp.
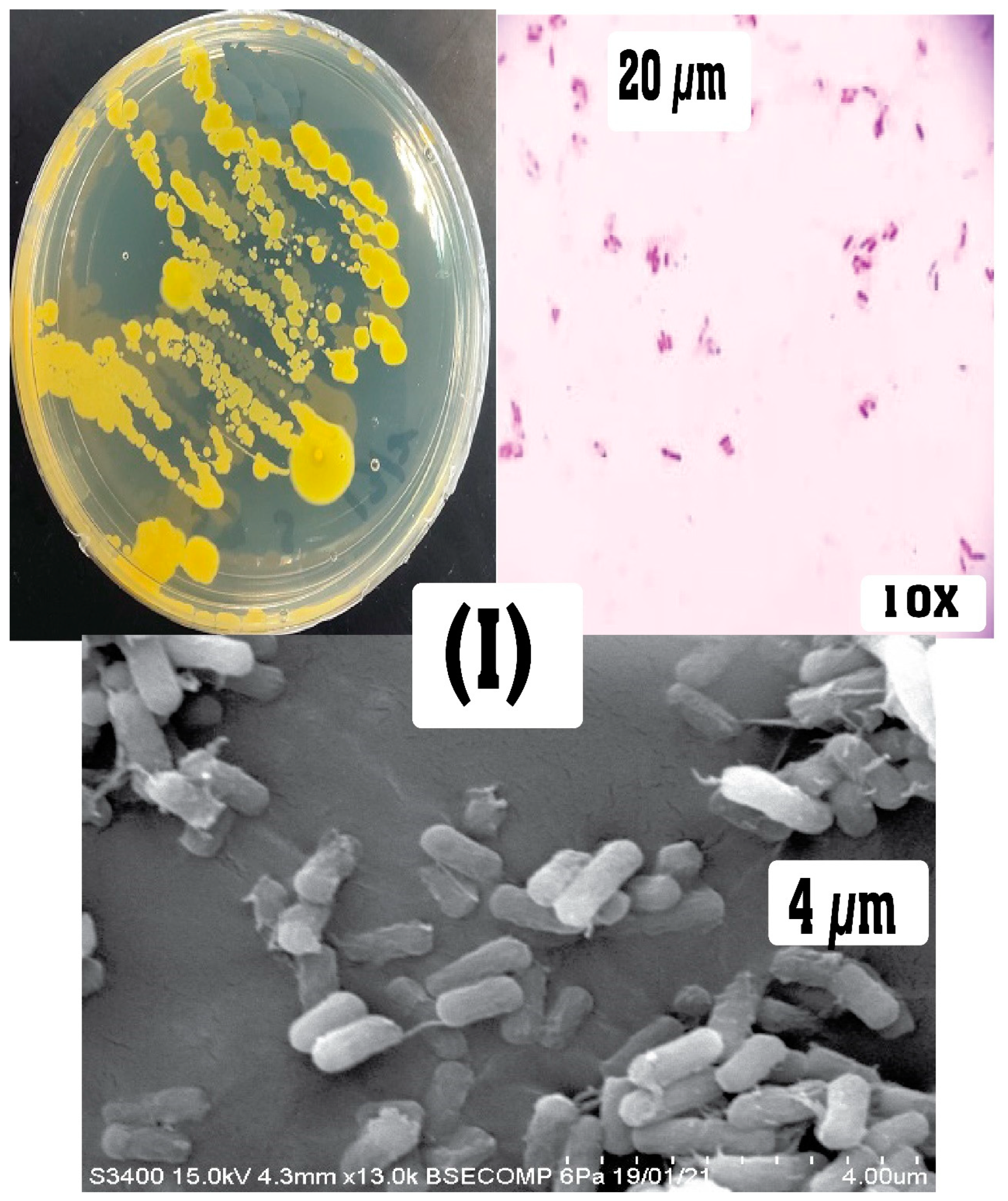
Both isolates were Gram-positive, rod-shaped bacilli, spore-forming, and able to grow in facultative anaerobic conditions, with differences in morphology and biochemical features. The colony of APPB1 had a marked yellowish color with slender cells, while APPB2 had a marked pale yellowish color with a robust cell structure (Figure 3). These cell and morphological features categorized the isolates in Firmicutes, a bacterial group that can survive in harsh environmental conditions and is widely used in many bioremediation applications [47,48]. The yellowish pigmentation is a good indicator of alkalophilic bacterial strains whose walls contain teichuronic acid. The teichuronopeptide consisted of polyglucuronic acid and other acidic amino acids, which assist them to withstand elevated pH levels. Additionally, for optimal bacterial growth at high pH, the acidic compounds and Na+ tend to be essential [49]. Bacterial strains from the bacilli are favored in retting due to their extracellular enzymes and spore-forming ability; among them, the alkali tolerant species are considered more feasible to be used as non-sterile retting as they prevent the admixture of unwanted endogenous colonies [13]. Moreover, bacterial pectinases withstand high pH levels and are viable in retting compared with other fungal sources [50,51].
3.2. Augmented Kenaf Retting Using APPB Combined with Water Solvents
The kenaf degumming efficacy of APPB with macerating solvents was verified in non-sterile retting conditions. The visual observations of the whole kenaf plants after 6 days of retting treatments are shown in Figure 4. The first steps of retting were initiated by the maceration of solvents, where the plants’ cell walls turn soft and soluble proteins make an appropriate environment for the microbes. The four significant retting steps involved in bio-degumming are: (i) absorption of solvents and forming of calcium oxalate crystals; (ii) removing the cortex; (iii) eliminating gums from the middle lamella; and, finally, (iv) eliminating gums from the fiber surface. The first two steps have solvent maceration effects, which also make favorable sites for microbial enzymatic activities [18,52]. The retting effect by visual observation produced a distinguished state in kenaf plant parts as uninoculated niches (UFW and USW) remained hard and intact with woody core portions compared with what was obtained with the inoculated treatments.
The incorporation of APPB1 with SW showed great value in the fiber bundle separation feasibility (Figure 4c), which is comparable with APPB1 with FW (Figure 4d). The trend of the fibers’ separation offered a clear indication of retting achievement. This might positively affect the inoculation of alkalophilic bacterial strains, as they prevent the unwanted endogenous microbial mixture [13]. Similar results were observed for the APPB2 with the same macerating solvents, while with FW, it starts to ret the fibers and additional treatment is required (Figure 4f). However, predicting the actual retting efficacy requires intensive investigation of the retting and fiber properties.
3.3. Pectinase Enzyme Activity
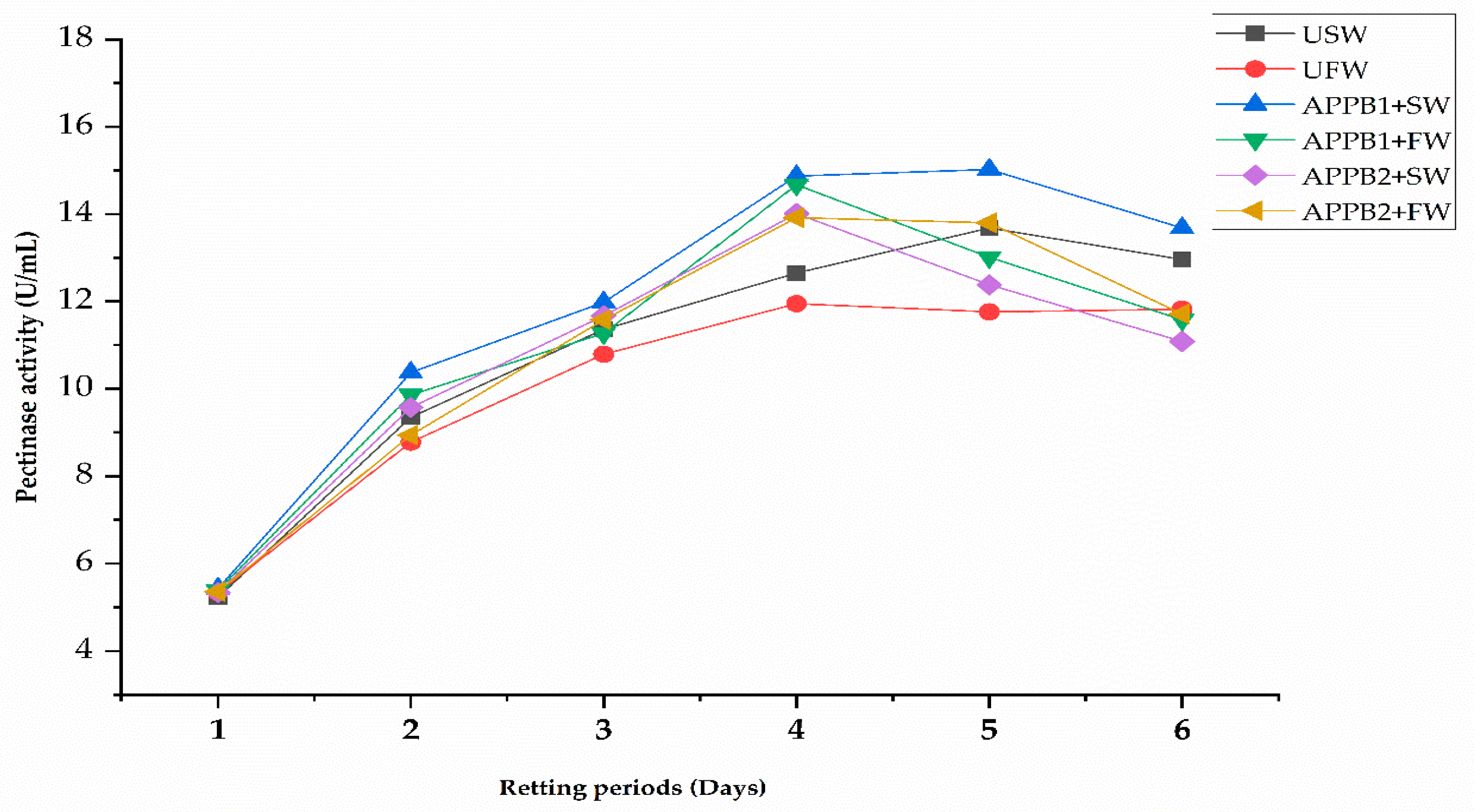
The APPB and solvent-treated kenaf retting niches were subjected to tab on their pectinase activity after being steeped for 24 h up to 6 days. The activity was counted based on the galacturonic acid contents of the pectic substances, using the retting effluents of each retting treatment in 24 h intervals. The result of each treatment is presented in Figure 5. A gradual increase in the pectinase activity was observed from days one to four. It plateaued on day five, and a slightly declining trend was observed irrespective of all treatments. Among the treatments, the degree of the activity varied, and APPB1 with seawater showed the highest (14.67 U/mL) pectinase activity. The determination of the pectinase enzymatic activity in the degumming cycle indicated the state of pectineus substance degradation, which was positively correlated with retting achievement [53,54]. However, this pectinase activity varied on several factors, including the occurrence of specific bacteria, the maceration effects of the solvents, and their biotic and abiotic factors.
The pectinase activity of the APPB inoculated retting niches was recorded to be comparatively higher than those of the uninoculated retting niches. This phenomenon might be due to the inoculation and proliferation of pectinolytic bacteria. Even though multiple enzymes are required to degrade kenaf heterogeneous NCGs, pectinase is the main retting enzyme determining the retting dynamics and applications of high-pectinase-potential bacterial strains were advantageous to yielding quality fiber. Previous studies revealed that the inoculation of pectinolytic bacteria significantly reduced the retting duration and achieved an optimal degree of retting in half of the time when compared with traditional retting without inoculum [23,55]. This pectinase activity was consistent with the visual degumming of kenaf plant materials. In addition, xylan is the major hemicellulose component of kenaf bast fibers, having 1,4 linked xylose units. Pectinase activities are also associated with hemicellulase and cellulase activities [45,56,57].
APPB with SW treatments showed the highest pectinase activity at the 4th day of retting. The first steps of retting were initiated by the maceration of solvents, where plants’ cell walls turn soft and soluble proteins make appropriate environments for the microbes. Previous studies have suggested that alkaliphiles favor Na+ for their optimum growth [58]. Other retting influencing factors that might have affected the degumming process include microbial growth characteristics and the maceration effects of the retting solvents. In the bio-retting process, endospore-forming species are highly efficient inoculants because of their higher survival rate in harsh retting conditions and, additionally, facultatively anaerobic strains benefit both aerobic and anaerobic phases [59,60].
3.4. Degree of Retting
The Fried test score of the six retting treatments is presented in Table 2. The results reveal that the uninoculated SW and FW treatments had a score of 1.5 and 1.0, indicating inferior separation feasibility and retting achievements. APPB with solvents scored from 3.5 to 4.75, clearly indicating the superior degree of kenaf fiber separation feasibility from its intact bindings. The highest score of APPB1 with SW might attributed to the compatibility of bacteria and solvents. However, the visual observations of kenaf-reverted stems (Figure 4) offered comparable findings to the results of the Fried test score and to the previous work reported by Zhang et al. [34]. However, the high test score supports that the inoculants successfully invaded the plants’ cuticle, thereby being admitted into the internal polysaccharide matrix [61]. The advancement of retting score in the APPB with SW treatment might have the combined positive effect of maceration and bacterial enzymes. The progression of retting was achieved by inundating the harvested plants with solvents where soluble cell wall compounds, such as sugars, nitrogenous substances, and proteins, are dissolved, allowing the microbial community to develop [52].
In this study, eye-estimated numerical values coupled with centrifugation forces were used to find the Fried test score. Although the chance of person-biased scores was higher to differentiate among the treatments effectively, it offered a clear insight into the retting achievement. In contrast, the expert growers confirmed the CWR achievement by touching and feeling the retted plants’ surfaces, rather than using any precise testing method.
3.5. Fiber Composition Analysis
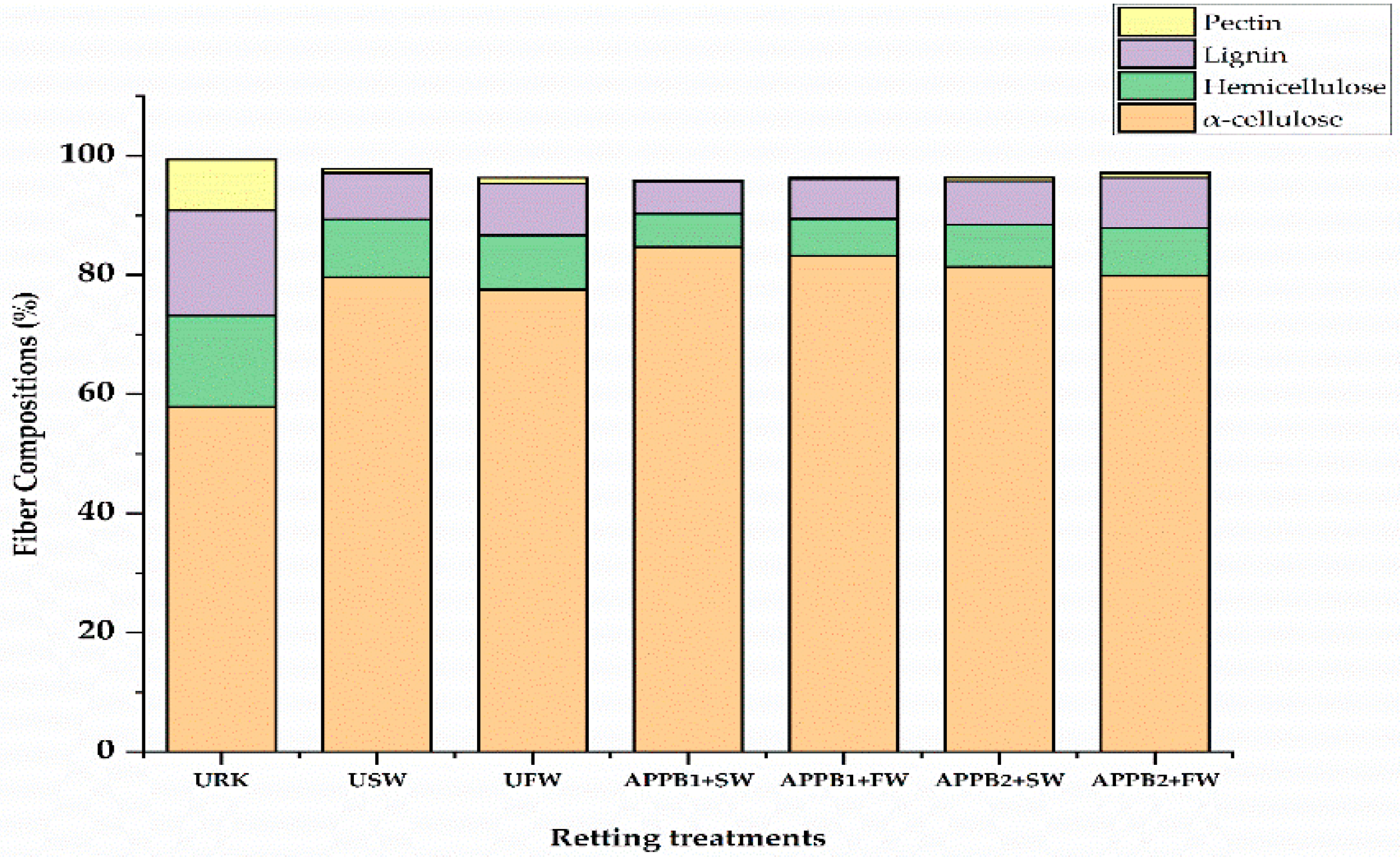
The fiber composition of kenaf-retted fibers from the non-sterilized retting treatments is shown in Figure 6. It was revealed that there was a significant increase in the cellulosic fiber compositions after retting compared with non-retted raw kenaf fiber (57.84%). The NCGs were eliminated at a differential rate, which affects the overall fiber compositions after treatment. The highest gum removal and elevated cellulose compositions (84.65%) were observed in the APPB-treated fibers combined with seawater treatments. However, the common phenomena of the retted fiber composition analysis showed that these retting treatments did not fully remove any of the major cell wall constituents.
The major chemical component of kenaf fibers that was significantly reduced due to retting was pectin, which plays a major role in achieving optimum degumming. Among the inoculated treatments, APPB1 with seawater yielded fibers showed maximum cellulose with the least gum’s compositions. This improvement in the cellulose composition of the retted kenaf fibers is indeed the result of the removal of non-cellulosic gums, including lignin, hemicellulose, and pectin. The variations in the balanced composition of NCGS and cellulose fibers were attributed to the lack of optimum degumming [62].
The inconsistency in kenaf fiber quality is the primary concern associated with the variations in its chemical compositions. This unsteadiness was primarily attributed to the lack of optimum degumming [62]. Moreover, under and over retting yielded inconsistent fiber compositions. Extended retting periods decompose the basic cellulosic fibers and under retting remained the gums unblemished. This is due to non-selective naturally occurring microflora, as some produce specific enzymes and the rest act as non-producers. This microbial mixture releases intermediate-level enzymes, which eventually delayed the process. The extended retting produced fiber rotting and degraded the basic cellulose components [14,15]. In contrast, inadequate degumming affects the overall chemical composition and affects the quality and end-uses [16,17].
3.6. Physical Fiber Quality
3.6.1. Surface Morphology
SEM images of retting-treated kenaf fiber surface analysis are shown in Figure 7. Figure 7a,b show uninoculated-treated fibers with FW and SW, respectively, indicating substantial gums are present on the fibers’ surfaces after six days of the retting treatments. Both treatments kept the fibers intact and required additional retting to remove the inherent gums. Figure 7c shows the APPB1-inoculated treatments combined with seawater solvents, revealing comparably smooth surfaces, which may be due to maximum NCG removal. In e, f, similar fibers on the surfaces of APPB2 combined with SW and FW indicated some degree of gums on their surfaces, while being cleaner than those of the uninoculated kenaf fibers.
The findings of the surface morphology revealed by the SEM technique after the retting treatments are consistent with the results obtained with the fiber composition analysis. The surfaces of the kenaf fibers retrieved from water retting did not show any mechanical damages, regardless of the presence and absence of adhered gums. These findings are consistent with the report by [63,64] of retted kenaf fibers.
3.6.2. Fiber Diameter and Fineness
The retted fibers’ diameter and their corresponding fineness are shown in Figure 8 and Table 3. The kenaf fibers’ bundle diameter retrieved from APPB and solvent-augmented retting, on the microscopic scale, ranged from 105.86 (0.10 mm) to 84.20 µm (0.08 mm). These findings are comparable to the degummed fibers (113 µm) reported by [65]. In textile technology, fiber fineness is described as weight per unit length, also known as linear density. It is the positive relationship between the fiber’s surface area and the substances present in its composition. The observations of the fibers’ diameters revealed that the uninoculated fibers possess higher fiber diameters, which, in turn, yielded coarser kenaf fibers (Table 3).
Among the retting treatments, the differentiations of non-cellulosic gums and the fiber dimensions were also observed. The surface and diameter of the natural fibers were aligned with the adjacent gums, and their elimination through the retting process reduced its dimensions and improved fineness [66]. The finer fibers showed the fibers possess a lower mass per unit of length. Natural cellulosic fibers with excellent fineness are suitable for fabrics and textiles. The fineness scores determined the end uses of the fiber as coarse fiber could use as rope, mat, or geotextile and fine fiber for textile fabric and clothing [67,68].
3.7. FTIR Spectra
The FTIR spectra of kenaf fiber obtained from six retting treatments at non-sterile retting are shown in Figure 9. FTIR is widely used to characterize natural fibers and obtain more in-depth information about their composition after various treatments. In this paper, FTIR spectroscopy investigation aimed to determine the functional groups of the cellulose-rich kenaf bast fibers after the augmented retting process. The scanning of grounded fiber samples (<1 mm) continued at a resolution of 4 cm−1 to depict the fiber’s constituents’ variation after retting treatments. The fibers constituents’ functional groups stretching pattern was used to differentiate the fiber quality. This non-destructive technique provides a strong perception of fiber compositions with reference to natural cellulosic fibers. In this paper, the spectra showed the degree of cellulose and non-cellulosic gummy substances, including hemicellulose, pectin, and lignin. The spectra were measured in comparison with the unretted raw kenaf bast fibers to illustrate the compositional changes after retting treatments and the variations among the treatments, ranging from 4000 to 500 cm−1 wave numbers.
The common phenomena revealed that yielded fibers possess the spectral intensity that prevails in untreated raw fibers, while their spectral stretching and intensity differed. The FTIR spectra of the kenaf bast samples showed that before retting the curve decreases in a much sharper shape than that produced after retting. Similar FTIR spectra of cellulose rich fibers were observed by [69]. The degree of spatial stretching of the curve among treatments was observed in each prominent peak points (a–k) at different wave lengths positions. FTIR spectroscopy can monitor the structure of individual chemical groups and each chemical group may have one or more specific absorption bands in the infrared spectra. The two main chemical groups observed thought the FTIR spectra due to the cellulosic effect were determined by the stretching of the two functional groups O-H and C=O [70].
The descriptions of each peak of the functional groups with reference to natural cellulosic fibers revealed a broad and intense peak “a” at ~3300 cm−1, which can be attributed to the characteristics of the cellulosic -OH bonds [71]. In this spectrum, the APPB1 with SW treatments showed the least sharp curve compared with those of all retting treatments. The spectra band “b” at ~3000 cm−1, which indicates the C-H stretching, showed that the retting treatments carried in non-sterile retting conditions on the fibers did not change the chemical structure of the treated fiber [72]. Polysaccharides, waxes, and oils, cellulose, and hemicellulose C–H symmetrical stretching and CH2 symmetrical stretching at ~2900 cm−1 indicated polysaccharide, wax, and oil were present. The peak “c” located at the ~1730 wave number in the raw kenaf fiber was assigned to the C=O stretching of the acetyl groups of hemicelluloses and p-coumaric acids of lignin [73]. This peak was prominent in the non-retted raw kenaf fibers, while the retted fibers were less intrusive but did not entirely disappear, indicating that the hemicellulose was still present in the fiber’s portions upon retting treatments. The prominent peak ‘e’ for raw kenaf fibers in the 1638–1543 wave numbers disappeared for solvent-retted fiber, indicating that sufficient NCGs were lost from the raw kenaf fibers upon retting treatments.
The peak between the 1000 and 1800 wave numbers assigned to the carbonyl group C=O represented ketone and aldehyde of aromatic lignin components. The absorption bands “g, h and i” resemble the scissoring, stretching, and bending vibrations of -CH2, -CH, and C-O bonds in cellulose. According to [74], the natural cellulose fibers’ crystalline and amorphous states were observed at peaks ‘f’ (~1400 cm−1) and ‘k’ (~850 cm−1). Natural cellulose fibers’ crystalline and amorphous states were observed at peaks ‘f’ (~1400 cm−1) and ‘k’ (~850 cm−1). The crystalline arrangements of natural fibers result from the intra and intermolecular arrangements through the hydroxyl (-OH) bonding and van der walls interactions originating from the cellulose portions [75,76]. In this case, the fluctuations of peak stretching arose due to the least amorphous gummy contents and increased crystalline cellulosic compositions among the treatments.
4. Conclusions
The bast cellulose fiber obtained thorough conventional water retting cannot yet satisfy the increasing needs and is itself facing scarcity difficulties. In this study, kenaf plants were successfully degummed by employing alkalophilic pectinase-producing bacteria with seawater. The enzymatic delicacy and bacteriological features of APPB were intended to be compatible with seawater to achieve successful retting. On the other hand, uninoculated retting niches (USW and UFW) kept the kenaf stem intact, indicating the unblemished non-cellulosic gummy substances within this retting tenure. When APPB were accompanied by seawater, optimal retting was achieved within six-day treatment cycles under non-sterile conditions. The visual degumming observations and the Fried score test revealed that the fibers could detach conveniently, comparable with the pectinase enzyme activity. The fiber composition analysis revealed increased cellulose fractions due to adequate pectin, hemicellulose, and lignin elimination. The SEM scans confirmed that APPB-inoculated retting niches yielded smooth fiber surfaces with the least gums present on the surfaces. In addition, the thinner bundle diameter with fine fibers showed the efficacy of the APPB-inoculated treatments.
Author Contributions
All the authors worked together for the conceptualization of this research project. The investigation of the project was mainly conducted by M.M.H. along with draft preparation. S.S. and V.K.S. reviewed and edited the manuscript. All authors provided their consent to the current version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research project was financed as part of the doctoral fellowship study No. NATP-2/PIU-BARC-44/2017/662(35) of the first author by the IFAD and World Bank-funded ‘National Agricultural Technology project-2′ (Project ID: P149553) of the People’s Republic of Bangladesh.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank to the ‘National Agricultural Technology project-2’ (Project ID: P149553), Government of the People’s Republic of Bangladesh, for their financial support as a foreign doctoral fellowship. We also thank the Biotechnology Research Institute and University Malaysia Sabah for infrastructure support and assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Papadopoulou, E.; Bikiaris, D.; Chrysafis, K.; Wladyka-Przybylak, M.; Wesolek, D.; Mankowski, J.; Kolodziej, J.; Baraniecki, P.; Bujnowicz, K.; Gronberg, V. Value-added industrial products from bast fiber crops. Ind. Crops Prod. 2015, 68, 116–125. [Google Scholar] [CrossRef]
- Tezara, C.; Siregar, J.P.; Lim, H.; Fauzi, F.; Yazdi, M.H.; Moey, L.; Lim, J. Factors that affect the mechanical properties of kenaf fiber reinforced polymer: A review. J. Mech. Eng. Sci. 2016, 10, 2159–2175. [Google Scholar]
- Narkpiban, K.; Poonsawat, T. Optimizing Cellulose Extraction from Kenaf (Hibiscus Cannabinus L.) Fiber by Selective Retting and Hydrothermal Pretreatment. J. Nat. Fibers 2020, 19, 700–713. [Google Scholar] [CrossRef]
- Zimniewska, M.; Wladyka-Przybylak, M.; Mankowski, J. Cellulosic bast fibers, their structure and properties suitable for composite applications. In Cellulose Fibers: Bio-and Nano-Polymer Composites; Springer: Berlin/Heidelberg, Germany, 2011; pp. 97–119. [Google Scholar]
- Ali, M.E.; Yong, C.K.; Ching, Y.C.; Chuah, C.H.; Liou, N.-S. Effect of single and double stage chemically treated kenaf fibers on mechanical properties of polyvinyl alcohol film. BioResources 2015, 10, 822–838. [Google Scholar] [CrossRef] [Green Version]
- Mondal, D.K.; Kaviraj, A. Ecotoxicological effects of jute retting on the survival of two freshwater fish and two invertebrates. Ecotoxicology 2008, 17, 207–211. [Google Scholar] [CrossRef]
- Nabiha Huda, A.; Tajaddin, R.M.; Ahmad, Z.; Nazrin, O.M. Determination of optimum debark time in retting process of kenaf fiber production. Int. Sustain. Civ. Eng. J. 2012, 1, 14–18. [Google Scholar]
- Ray, D.P.; Saha, S.C.; Sarkar, A.; Ghosh, R.K. Production of quality jute fibre through accelerated retting. Int. J. Bioresour. Sci. 2016, 3, 57. [Google Scholar] [CrossRef]
- Majumdar, B.; Chattopadhyay, L.; Barai, S.; Saha, A.; Sarkar, S.; Sarkar, S.; Mazumdar, S.; Saha, R.; Jha, S. Impact of conventional retting of jute (Corchorus spp.) on the environmental quality of water: A case study. Environ. Monit. Assess. 2019, 191, 440. [Google Scholar] [CrossRef]
- Zawani, Z.; Abdullah, L.C.; Abdan, K. Characterization of kenaf fibre-retting wastewater. Int. J. Sci. Res. 2015, 4, 314–317. [Google Scholar]
- Van Dam, J.E.; Bos, H.L. The environmental impact of fibre crops in industrial applications. In Hintergrundpapier zu: Van Dam, JEG; FAO: Rome, Italy, 2004. [Google Scholar]
- Hossain, M.M.; Siddiquee, S.; Kumar, V. Water Sources Derived Bio Retting Effect on Kenaf Fiber Compositions. J. Nat. Fibers 2021, 18, 1–14. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, R.; Chen, J.; Chen, J.; Feng, X. Seawater-retting treatment of hemp and characterization of bacterial strains involved in the retting process. Process Biochem. 2008, 43, 1195–1201. [Google Scholar] [CrossRef]
- Fernando, D.; Thygesen, A.; Meyer, A.S.; Daniel, G. Elucidating field retting mechanisms of hemp fibres for biocomposites: Effects of microbial actions and interactions on the cellular micro-morphology and ultrastructure of hemp stems and bast fibres. BioResources 2019, 14, 4047–4084. [Google Scholar] [CrossRef]
- Foulk, J.; Akin, D.; Dodd, R. Influence of pectinolytic enzymes on retting effectiveness and resultant fiber properties. BioResources 2008, 3, 155–169. [Google Scholar]
- Komuraiah, A.; Kumar, N.S.; Prasad, B.D. Chemical composition of natural fibers and its influence on their mechanical properties. Mech. Compos. Mater. 2014, 50, 359–376. [Google Scholar] [CrossRef]
- Sisti, L.; Totaro, G.; Vannini, M.; Celli, A. Retting process as a pretreatment of natural fibers for the development of polymer composites. In Lignocellulosic Composite Materials; Springer: Berlin/Heidelberg, Germany, 2018; pp. 97–135. [Google Scholar]
- Mao, K.; Chen, H.; Qi, H.; Qiu, Z.; Zhang, L.; Zhou, J. Visual degumming process of ramie fiber using a microbial consortium RAMCD407. Cellulose 2019, 26, 3513–3528. [Google Scholar] [CrossRef]
- Duan, S.W.; Cheng, L.F.; Feng, X.Y.; Yang, Q.; Liu, Z.Y.; Zheng, K.; Peng, Y.D. Insights on bio-degumming of kenaf bast based on metagenomic and proteomics. BMC Genom. 2020, 21, 121. [Google Scholar] [CrossRef] [Green Version]
- Visi, D.K.; D’Souza, N.; Ayre, B.G.; Webber, C.L., III; Allen, M.S. Investigation of the bacterial retting community of kenaf (Hibiscus cannabinus) under different conditions using next-generation semiconductor sequencing. J. Ind. Microbiol. Biotechnol. 2013, 40, 465–475. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, P.; Pan, C.; Du, R.; Ping, W.; Ge, J. Bacterial succession and metabolite changes during flax (Linum usitatissimum L.) retting with Bacillus cereus HDYM-02. Sci. Rep. 2016, 6, 31812. [Google Scholar] [CrossRef]
- Allison, S.D. Cheaters, diffusion and nutrients constrain decomposition by microbial enzymes in spatially structured environments. Ecol. Lett. 2005, 8, 626–635. [Google Scholar] [CrossRef]
- Ge, J.; Yang, Z.; Du, R.; Zhang, L.; Ping, W.; Zhao, D. Production of Pectinolytic Enzymes by Two Bacillus spp. Strains and Their Application in Flax Degumming. Trans. Tianjin Univ. 2019, 25, 413–419. [Google Scholar] [CrossRef]
- Ghazala, I.; Sayari, N.; Romdhane, M.B.; Ellouz-Chaabouni, S.; Haddar, A. Assessment of pectinase production by Bacillus mojavensis I4 using an economical substrate and its potential application in oil sesame extraction. J. Food Sci. Technol. 2015, 52, 7710–7722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, R.; Aktar, N.; Kabir, S.M.T.; Honi, U.; Halim, A.; Islam, R.; Sarker, M.D.H.; Haque, M.S.; Alam, M.M.; Islam, M.S. Pectinolytic Bacterial Consortia Reduce Jute Retting Period and Improve Fibre Quality. Sci. Rep. 2020, 10, 5174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Majumdar, B.; Saha, A.R. Biodegradation of Plant Pectin and Hemicelluloses with Three Novel Bacillus pumilus Strains and Their Combined Application for Quality Jute Fibre Production. Agric. Res. 2015, 4, 354–364. [Google Scholar] [CrossRef]
- Kohli, P.; Gupta, R. Alkaline pectinases: A review. Biocatal. Agric. Biotechnol. 2015, 4, 279–285. [Google Scholar] [CrossRef]
- Guadie, A.; Tizazu, S.; Melese, M.; Guo, W.; Ngo, H.H.; Xia, S. Biodecolorization of textile azo dye using Bacillus sp. strain CH12 isolated from alkaline lake. Biotechnol. Rep. 2017, 15, 92–100. [Google Scholar] [CrossRef]
- Zheng, L.; Du, Y.; Zhang, J. Degumming of ramie fibers by alkalophilic bacteria and their polysaccharide-degrading enzymes. Bioresour. Technol. 2001, 78, 89–94. [Google Scholar] [CrossRef]
- Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.-H.; Whitman, W.B. Bergey’s Manual of Systematic Bacteriology: Volume 3: The Firmicutes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; Volume 3. [Google Scholar]
- Cao, J.; Zheng, L.; Chen, S. Screening of pectinase producer from alkalophilic bacteria and study on its potential application in degumming of ramie. Enzym. Microb. Technol. 1992, 14, 1013–1016. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Silva, D.; Tokuioshi, K.; da Silva Martins, E.; Da Silva, R.; Gomes, E. Production of pectinase by solid-state fermentation with Penicillium viridicatum RFC3. Process Biochem. 2005, 40, 2885–2889. [Google Scholar] [CrossRef]
- Zhang, J. Biochemical Study and Technical Applications of Fungal Pectinase; Acta Universitatis Upsaliensis: Stockholm, Sweden, 2006. [Google Scholar]
- Munder, F.; Furll, C. Effective processing of bast fiber plants and mechanical properties of the fibers. In Proceedings of the 2004 ASAE Annual Meeting, Ottawa, ON, Canada, 1–4 August 2004; p. 1. [Google Scholar]
- Chen, Y.; Belsham, T.; Laguë, C.; Landry, H.; Peng, Q.; Zhong, W. Fineness and tensile properties of hemp (Cannabis sativa L.) fibres. Biosyst. Eng. 2011, 108, 9–17. [Google Scholar] [CrossRef]
- Madhu, P.; Sanjay, M.; Senthamaraikannan, P.; Pradeep, S.; Saravanakumar, S.; Yogesha, B. A review on synthesis and characterization of commercially available natural fibers: Part-I. J. Nat. Fibers 2018, 16, 1132–1144. [Google Scholar] [CrossRef]
- Punyamurthy, R.; Sampathkumar, D.; Srinivasa, C.V.; Bennehalli, B. Effect of alkali treatment on water absorption of single cellulosic abaca fiber. BioResources 2012, 7, 3515–3524. [Google Scholar]
- George, M.; Mussone, P.G.; Bressler, D.C. Surface and thermal characterization of natural fibres treated with enzymes. Ind. Crops Prod. 2014, 53, 365–373. [Google Scholar] [CrossRef]
- Van Nimmen, E.; De Clerck, K.; Verschuren, J.; Gellynck, K.; Gheysens, T.; Mertens, J.; Van Langenhove, L. FT-IR spectroscopy of spider and silkworm silks: Part I. Different sampling techniques. Vib. Spectrosc. 2008, 46, 63–68. [Google Scholar] [CrossRef]
- Takami, H.; Horikoshi, K. Analysis of the genome of an alkaliphilic Bacillus strain from an industrial point of view. Extremophiles 2000, 4, 99–108. [Google Scholar] [CrossRef]
- Preiss, L.; Hicks, D.B.; Suzuki, S.; Meier, T.; Krulwich, T.A. Alkaliphilic bacteria with impact on industrial applications, concepts of early life forms, and bioenergetics of ATP synthesis. Front. Bioeng. Biotechnol. 2015, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Saha, D.; Chattopadhyay, L.; Majumdar, B. Genome comparison identifies different bacillus species in a bast fibre-retting bacterial consortium and provides insights into pectin degrading genes. Sci. Rep. 2020, 10, 8169. [Google Scholar] [CrossRef]
- Cheng, L.; Duan, S.; Feng, X.; Zheng, K.; Yang, Q.; Xu, H.; Luo, W.; Peng, Y. Screening and identification of pectinolytic bacteria for ramie degumming. Text. Res. J. 2021, 91, 1056–1064. [Google Scholar] [CrossRef]
- Brühlmann, F.; Kim, K.S.; Zimmerman, W.; Fiechter, A. Pectinolytic enzymes from actinomycetes for the degumming of ramie bast fibers. Appl. Environ. Microbiol. 1994, 60, 2107–2112. [Google Scholar] [CrossRef] [Green Version]
- Attri, S.; Garg, G. Isolation of microorganisms simultaneously producing xylanase, pectinase and cellulase enzymes using cost effective substrates. J. Innov. Biol. 2014, 1, 45–50. [Google Scholar]
- Mukhopadhya, I.; Moraïs, S.; Laverde-Gomez, J.; Sheridan, P.O.; Walker, A.W.; Kelly, W.; Klieve, A.V.; Ouwerkerk, D.; Duncan, S.H.; Louis, P. Sporulation capability and amylosome conservation among diverse human colonic and rumen isolates of the keystone starch-degrader Ruminococcus bromii. Environ. Microbiol. 2018, 20, 324–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, C.; Filippidou, S.; Jamil, I.; Kooli, W.; House, G.L.; Estoppey, A.; Hayoz, M.; Junier, T.; Palmieri, F.; Wunderlin, T. Bacterial spores, from ecology to biotechnology. Adv. Appl. Microbiol. 2019, 106, 79–111. [Google Scholar] [PubMed]
- Aono, R.; Horikoshi, K. Carotenes produced by alkaliphilic yellow-pigmented strains of Bacillus. Agric. Biol. Chem. 1991, 55, 2643–2645. [Google Scholar] [CrossRef]
- Sohail, M.; Latif, Z. Phylogenetic analysis of polygalacturonase-producing Bacillus and Pseudomonas isolated from plant waste material. Jundishapur J. Microbiol. 2016, 9, e28594. [Google Scholar] [CrossRef] [Green Version]
- Hoondal, G.S.; Tiwari, R.P.; Tewari, R.; Dahiya, N.; Beg, Q.K. Microbial alkaline pectinases and their industrial applications: A review. Appl. Microbiol. Biotechnol. 2002, 59, 409–418. [Google Scholar] [CrossRef]
- Di Candilo, M.; Bonatti, P.M.; Guidetti, C.; Focher, B.; Grippo, C.; Tamburini, E.; Mastromei, G. Effects of selected pectinolytic bacterial strains on water-retting of hemp and fibre properties. J. Appl. Microbiol. 2010, 108, 194–203. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, Q.; Feng, X.; Duan, S.; Yang, Q.; Zheng, K.; Liu, Z.; Liu, Z.; Peng, Y. Screening a bacterium and its effect on the biological degumming of ramie and kenaf. Sci. Agric. 2018, 75, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Nath, M.; Majumdar, B.; Das, S.; Mazumdar, S.P.; Saha, A.; Sarkar, S. Optimization of Fermentation Conditions for pectin degrading enzyme production by pectinolytic microbial consortia used for jute retting. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 925–931. [Google Scholar] [CrossRef]
- Das, S.; Majumdar, B.; Saha, A.; Sarkar, S.; Jha, S.; Sarkar, S.; Saha, R. Comparative study of conventional and improved retting of jute with microbial formulation. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1351–1357. [Google Scholar] [CrossRef]
- Bajpai, P. Bioconversion of hemicelluloses. In Biotechnology for Pulp and Paper Processing; Springer: Berlin/Heidelberg, Germany, 2018; pp. 545–560. [Google Scholar]
- Song, Y.; Jiang, W.; Zhang, Y.; Ben, H.; Han, G.; Ragauskas, A.J. Isolation and characterization of cellulosic fibers from kenaf bast using steam explosion and Fenton oxidation treatment. Cellulose 2018, 25, 4979–4992. [Google Scholar] [CrossRef]
- Krulwich, T.A.; Guffanti, A.A.; Ito, M. pH tolerance in Bacillus: Alkaliphiles versus non-alkaliphiles. Bact. Responses pH 1999, 221, 167. [Google Scholar]
- Hu, X.; Mahillon, J. Life Cycle and Gene Exchange. In Endospore-Forming Soil Bacteria; Springer: Berlin/Heidelberg, Germany, 2011; pp. 89–113. [Google Scholar]
- Radhakrishnan, R.; Hashem, A.; Abd-Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, G.; Akin, D.E.; Hanlin, R.T.; Rodriguez, C.; Archibald, D.D.; Rigsby, L.L.; Eriksson, K.L. Identification and retting efficiencies of fungi isolated from dew-retted flax in the United States and europe. Appl. Environ. Microbiol. 1997, 63, 3950–3956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Jiang, W.; Nie, K.; Zhang, Y.; Ben, H.; Han, G.; Ragauskas, A.J. An alkali-free method to manufacture ramie fiber. Text. Res. J. 2019, 89, 3653–3659. [Google Scholar] [CrossRef]
- Rozyanty, A.; Zhafer, S.; Shayfull, Z.; Nainggolan, I.; Musa, L.; Zheing, L. Effect of water and mechanical retting process on mechanical and physical properties of kenaf bast fiber reinforced unsaturated polyester composites. Compos. Struct. 2021, 257, 113384. [Google Scholar] [CrossRef]
- Nie, K.; Liu, B.; Zhao, T.; Wang, H.; Song, Y.; Ben, H.; Ragauskas, A.J.; Han, G.; Jiang, W. A facile degumming method of kenaf fibers using deep eutectic solution. J. Nat. Fibers 2020, 19, 1115–1125. [Google Scholar] [CrossRef]
- Jiang, W.; Han, G.; Zhang, Y.; Liu, S.; Zhou, C.; Song, Y.; Zhang, X.; Xia, Y. Monitoring chemical changes on the surface of kenaf fiber during degumming process using infrared microspectroscopy. Sci. Rep. 2017, 7, 1240. [Google Scholar] [CrossRef] [Green Version]
- Beltran, R.; Hurren, C.J.; Kaynak, A.; Wang, X. Correlating the fineness and residual gum content of degummed hemp fibres. Fibers Polym. 2002, 3, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Misnon, M.I.; Islam, M.M.; Epaarachchi, J.A.; Lau, K.-T. Potentiality of utilising natural textile materials for engineering composites applications. Mater. Des. 2014, 59, 359–368. [Google Scholar] [CrossRef]
- Meshram, J.H.; Palit, P. Biology of industrial bast fibers with reference to quality. J. Nat. Fibers 2013, 10, 176–196. [Google Scholar] [CrossRef]
- Galiwango, E.; Rahman, N.S.A.; Al-Marzouqi, A.H.; Abu-Omar, M.M.; Khaleel, A.A. Isolation and characterization of cellulose and α-cellulose from date palm biomass waste. Heliyon 2019, 5, e02937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Wu, P. Investigation of the hydrogen-bond structure of cellulose diacetate by two-dimensional infrared correlation spectroscopy. Carbohydr. Polym. 2008, 74, 509–513. [Google Scholar] [CrossRef]
- Le Troedec, M.; Sedan, D.; Peyratout, C.; Bonnet, J.P.; Smith, A.; Guinebretiere, R.; Gloaguen, V.; Krausz, P. Influence of various chemical treatments on the composition and structure of hemp fibres. Compos. Part A Appl. Sci. Manuf. 2008, 39, 514–522. [Google Scholar] [CrossRef]
- Saelee, K.; Yingkamhaeng, N.; Nimchua, T.; Sukyai, P. An environmentally friendly xylanase-assisted pretreatment for cellulose nanofibrils isolation from sugarcane bagasse by high-pressure homogenization. Ind. Crops Prod. 2016, 82, 149–160. [Google Scholar] [CrossRef]
- Biagiotti, J.; Puglia, D.; Torre, L.; Kenny, J.M.; Arbelaiz, A.; Cantero, G.; Marieta, C.; Llano-Ponte, R.; Mondragon, I. A systematic investigation on the influence of the chemical treatment of natural fibers on the properties of their polymer matrix composites. Polym. Compos. 2004, 25, 470–479. [Google Scholar] [CrossRef]
- Hospodarova, V.; Singovszka, E.; Stevulova, N. Characterization of cellulosic fibers by FTIR spectroscopy for their further implementation to building materials. Am. J. Anal. Chem. 2018, 9, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Sheltami, R.M.; Abdullah, I.; Ahmad, I.; Dufresne, A.; Kargarzadeh, H. Extraction of cellulose nanocrystals from mengkuang leaves (Pandanus tectorius). Carbohydr. Polym. 2012, 88, 772–779. [Google Scholar] [CrossRef]
- Zhang, X.; Han, G.; Jiang, W.; Zhang, Y.; Li, X.; Li, M. Effect of steam pressure on chemical and structural properties of kenaf fibers during steam explosion process. BioResources 2016, 11, 6590–6599. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Formation of a pectin hydrolysis zone by alkalophilic pectinase-producing bacteria, APPB1 (b) and APPB2 (c), compared with a no hydrolysis zone (a).
Figure 1.
Formation of a pectin hydrolysis zone by alkalophilic pectinase-producing bacteria, APPB1 (b) and APPB2 (c), compared with a no hydrolysis zone (a).

Figure 2.
Quantitative pectinase, xylanase, and cellulase enzymatic activities of the alkalophilic bacterial isolates APPB1 and APPB2.
Figure 2.
Quantitative pectinase, xylanase, and cellulase enzymatic activities of the alkalophilic bacterial isolates APPB1 and APPB2.

Figure 3.
Physical characteristics of colony morphology, Gram staining (under stereo and light microscope), and SEM view of the APPB isolates on nutrient agar: (I) APPB1 and (II) APPB2.
Figure 3.
Physical characteristics of colony morphology, Gram staining (under stereo and light microscope), and SEM view of the APPB isolates on nutrient agar: (I) APPB1 and (II) APPB2.


Figure 4.
Augmented retting effect on kenaf plants. (a) Uninoculated with FW; (b) Uninoculated with SW. (c) APPB1 with SW. (d) APPB1 with FW. (e) APPB2 with SW. (f) APPB2 with FW.
Figure 4.
Augmented retting effect on kenaf plants. (a) Uninoculated with FW; (b) Uninoculated with SW. (c) APPB1 with SW. (d) APPB1 with FW. (e) APPB2 with SW. (f) APPB2 with FW.

Figure 5.
Pectinase enzyme activity during non-sterile kenaf retting using alkalophilic pectinolytic bacteria combined with retting solvents.
Figure 5.
Pectinase enzyme activity during non-sterile kenaf retting using alkalophilic pectinolytic bacteria combined with retting solvents.

Figure 6.
Compositions of the kenaf bast fibers retrieved from augmented retting in comparison with unretted raw kenaf.
Figure 6.
Compositions of the kenaf bast fibers retrieved from augmented retting in comparison with unretted raw kenaf.

Figure 7.
SEM images of retted kenaf fibers at a 100 µm magnification, showing the presence and absence of amounts of gummy substances on the fiber’s surfaces. (a) Uninoculated kenaf retted fiber with FW. (b) Uninoculated with SW. (c) APPB1 with SW. (d) APPB1 with FW. (e) APPB2 with SW. (f) APPB2 with FW.
Figure 7.
SEM images of retted kenaf fibers at a 100 µm magnification, showing the presence and absence of amounts of gummy substances on the fiber’s surfaces. (a) Uninoculated kenaf retted fiber with FW. (b) Uninoculated with SW. (c) APPB1 with SW. (d) APPB1 with FW. (e) APPB2 with SW. (f) APPB2 with FW.

Figure 8.
SEM images of the retted kenaf fibers at a 1 mm magnification, showing the fiber diameters. (a). Uninoculated with SW. (b) Uninoculated with FW. (c) APPB1 with SW. (d) APPB1 with FW. (e) APPB2 with SW. (f) APPB2 with FW.
Figure 8.
SEM images of the retted kenaf fibers at a 1 mm magnification, showing the fiber diameters. (a). Uninoculated with SW. (b) Uninoculated with FW. (c) APPB1 with SW. (d) APPB1 with FW. (e) APPB2 with SW. (f) APPB2 with FW.

Figure 9.
FTIR spectra retted kenaf fibers compared with the unretted raw kenaf bast fibers. The retted fibers recaptured from the six-day retting treatments in combination with APPB and retting solvents.
Figure 9.
FTIR spectra retted kenaf fibers compared with the unretted raw kenaf bast fibers. The retted fibers recaptured from the six-day retting treatments in combination with APPB and retting solvents.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of retting treatments used in augmented kenaf retting.
| Treatments | Combinations | Designations |
|---|---|---|
| T1 | Uninoculated with seawater | USW |
| T2 | Uninoculated with freshwater | UFW |
| T3 | APPB1 with seawater | APPB1 + SW |
| T4 | APPB1 with freshwater | APPB1 + FW |
| T5 | APPB2 with seawater | APPB2 + SW |
| T6 | APPB2 with sresh water | APPB2 + FW |
Table 2.
Fried test scores of the kenaf retting treatments.
| Retting Treatments | Test Score |
|---|---|
| USW | 1.50 (2,2,1,1) |
| UFW | 1.0 (1,1,1,1) |
| APPB1 + SW | 4.75 (5,5,4,5) |
| APPB1 + FW | 4.25 (5,4,4,4) |
| APPB2 + SW | 3.75 (4,4,4,3) |
| APPB2 + FW | 3.5 (4,4,3,3) |
Table 3.
Fiber diameter and fineness of the retted kenaf fibers.
| Treatments | Fiber Bundles Mean Diameter (µm) | Fineness Gtex (Equal to g/103 m of Fiber Length) |
|---|---|---|
| UFW | 105.86 | 12.74 |
| USW | 102.13 | 11.87 |
| APPB1 + SW | 84.20 | 7.65 |
| APPB1 + FW | 89.5 | 9.11 |
| APPB2 + SW | 93.83 | 10.02 |
| APPB2 + FW | 97.43 | 10.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hossain, M.M.; Subbiah, V.K.; Siddiquee, S. Augmented Retting Effect on Kenaf Fibers Using Alkalophilic Pectinase-Producing Bacteria in Combination with Water Solvents. Appl. Sci. 2022, 12, 7136. https://doi.org/10.3390/app12147136
AMA Style
Hossain MM, Subbiah VK, Siddiquee S. Augmented Retting Effect on Kenaf Fibers Using Alkalophilic Pectinase-Producing Bacteria in Combination with Water Solvents. Applied Sciences. 2022; 12(14):7136. https://doi.org/10.3390/app12147136
Chicago/Turabian StyleHossain, Mohammad Munir, Vijay Kumar Subbiah, and Shafiquzzaman Siddiquee. 2022. "Augmented Retting Effect on Kenaf Fibers Using Alkalophilic Pectinase-Producing Bacteria in Combination with Water Solvents" Applied Sciences 12, no. 14: 7136. https://doi.org/10.3390/app12147136
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.