
Beekeeping in the Desert: Foraging Activities of Honey Bee during Major Honeyflow in a Hot-Arid Ecosystem

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Area and Plant Species
2.2. Flowering Density
2.3. Honey Bee Subspecies
2.4. Foraging (Outgoing and Incoming Pollen-Gathering) Activities of Honey Bees
2.5. Collection of Meteorological Data
2.6. Thermal Window for Flight Activity
2.7. Statistical Analysis
3. Results
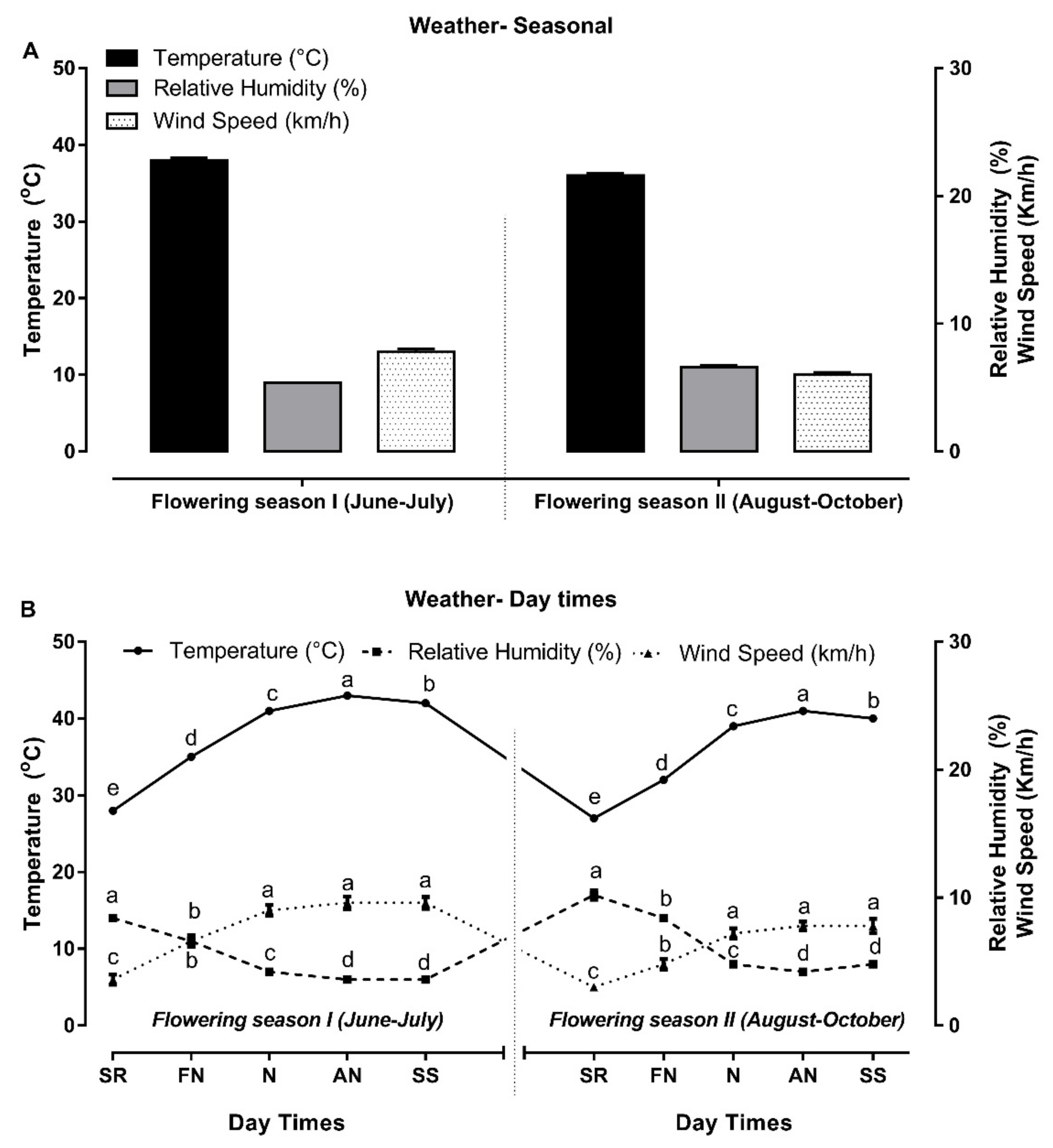
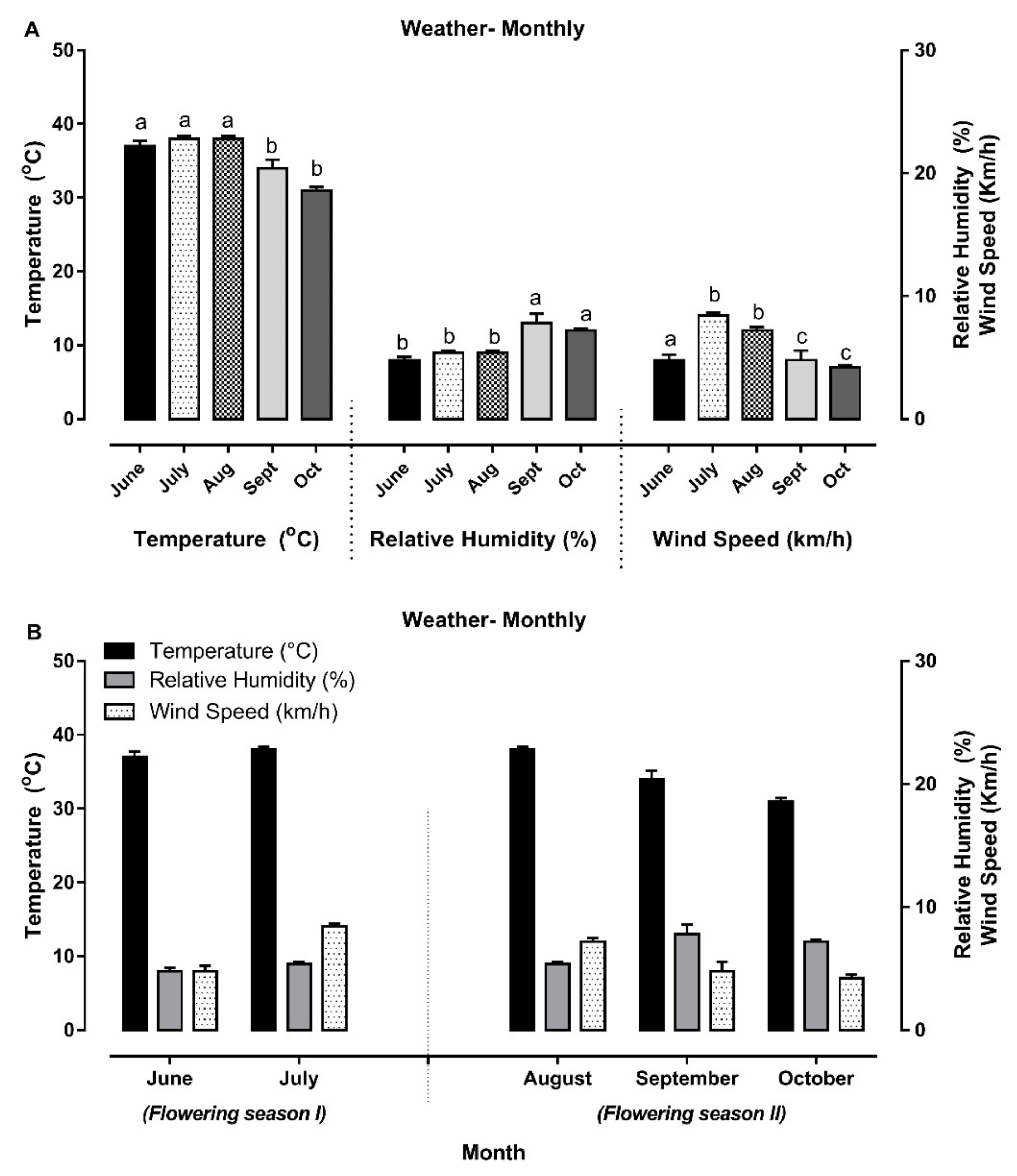
3.1. Meteorological Data
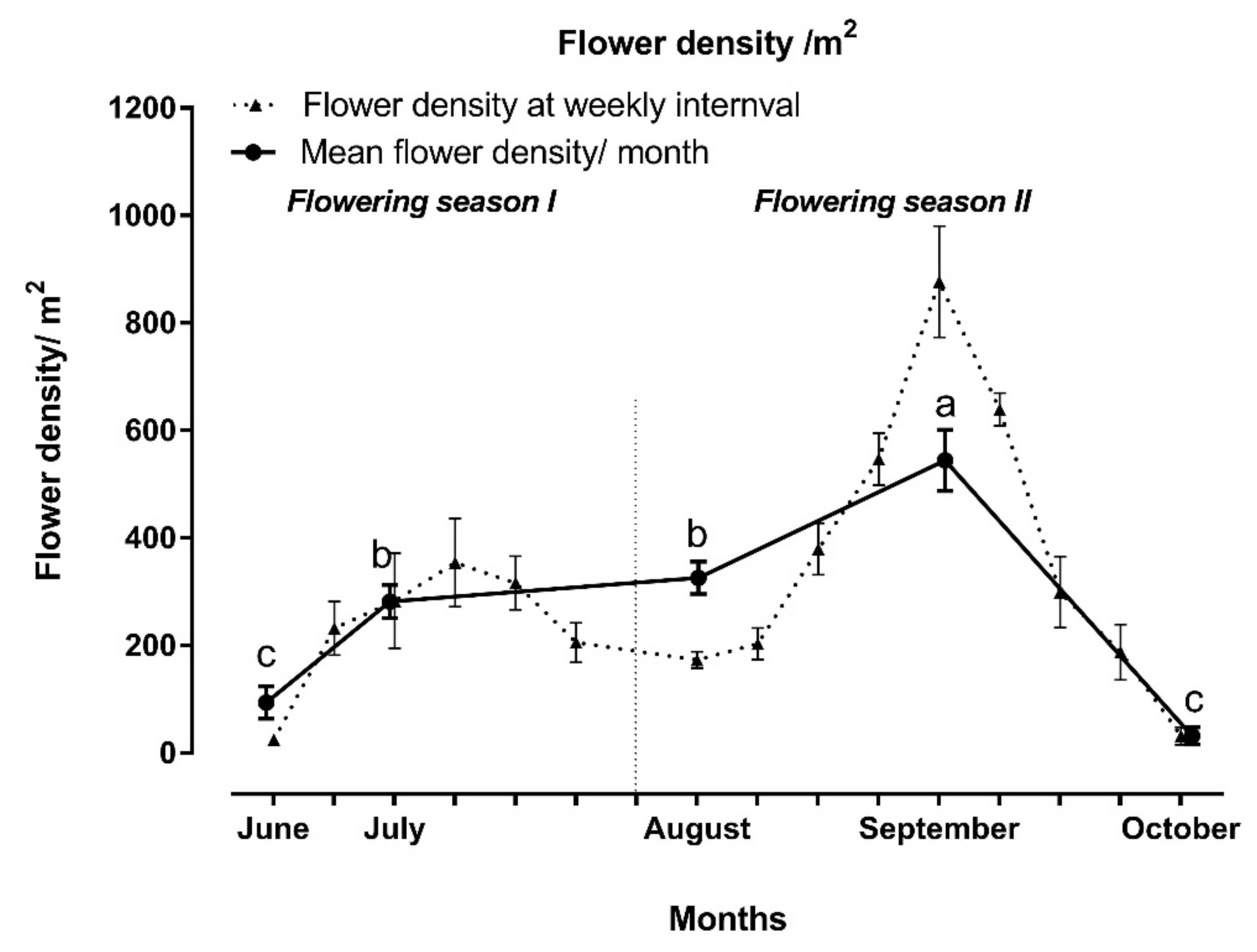
3.2. Flowering Density of Z. nummularia
3.3. Foraging Activities
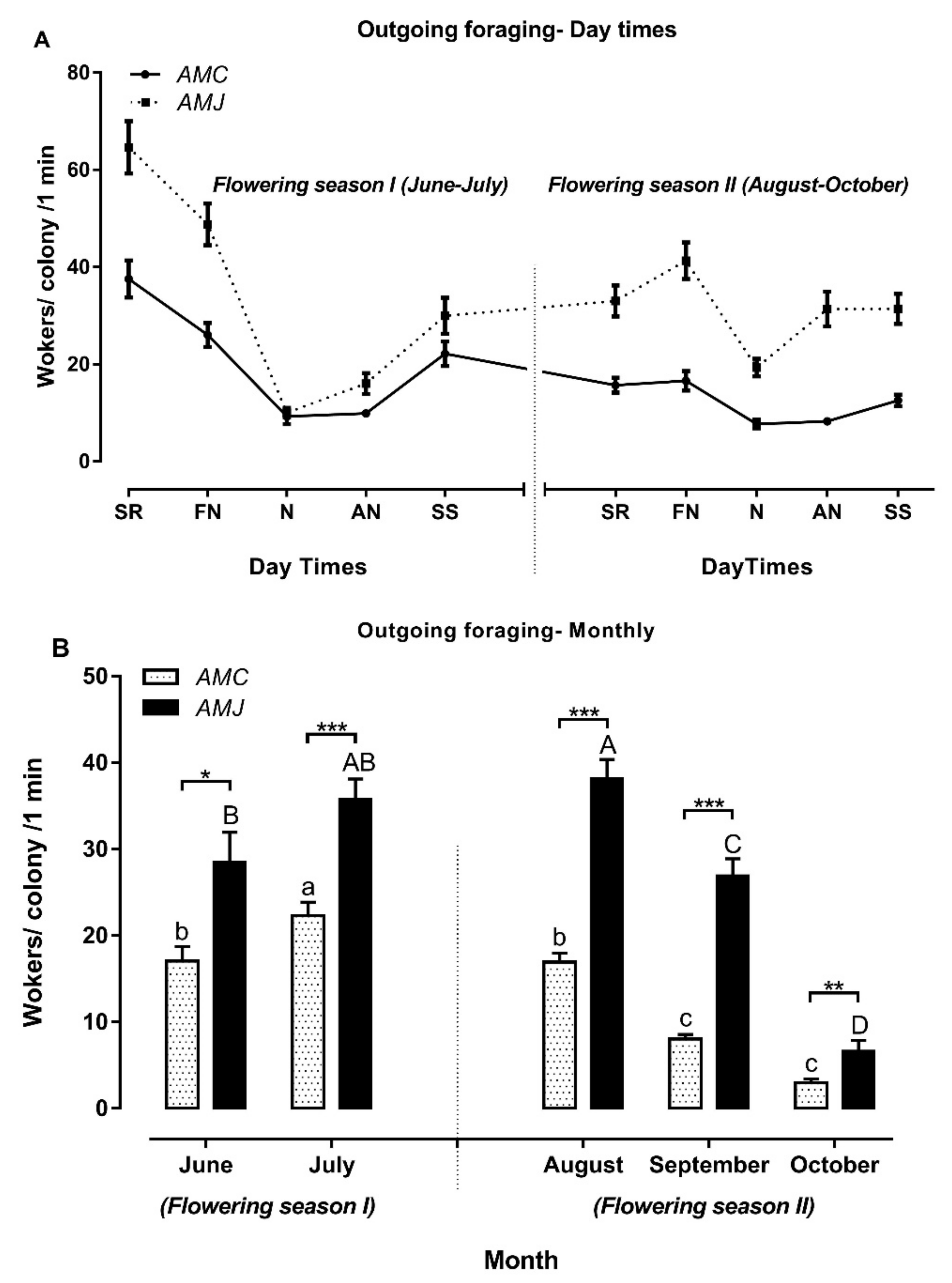
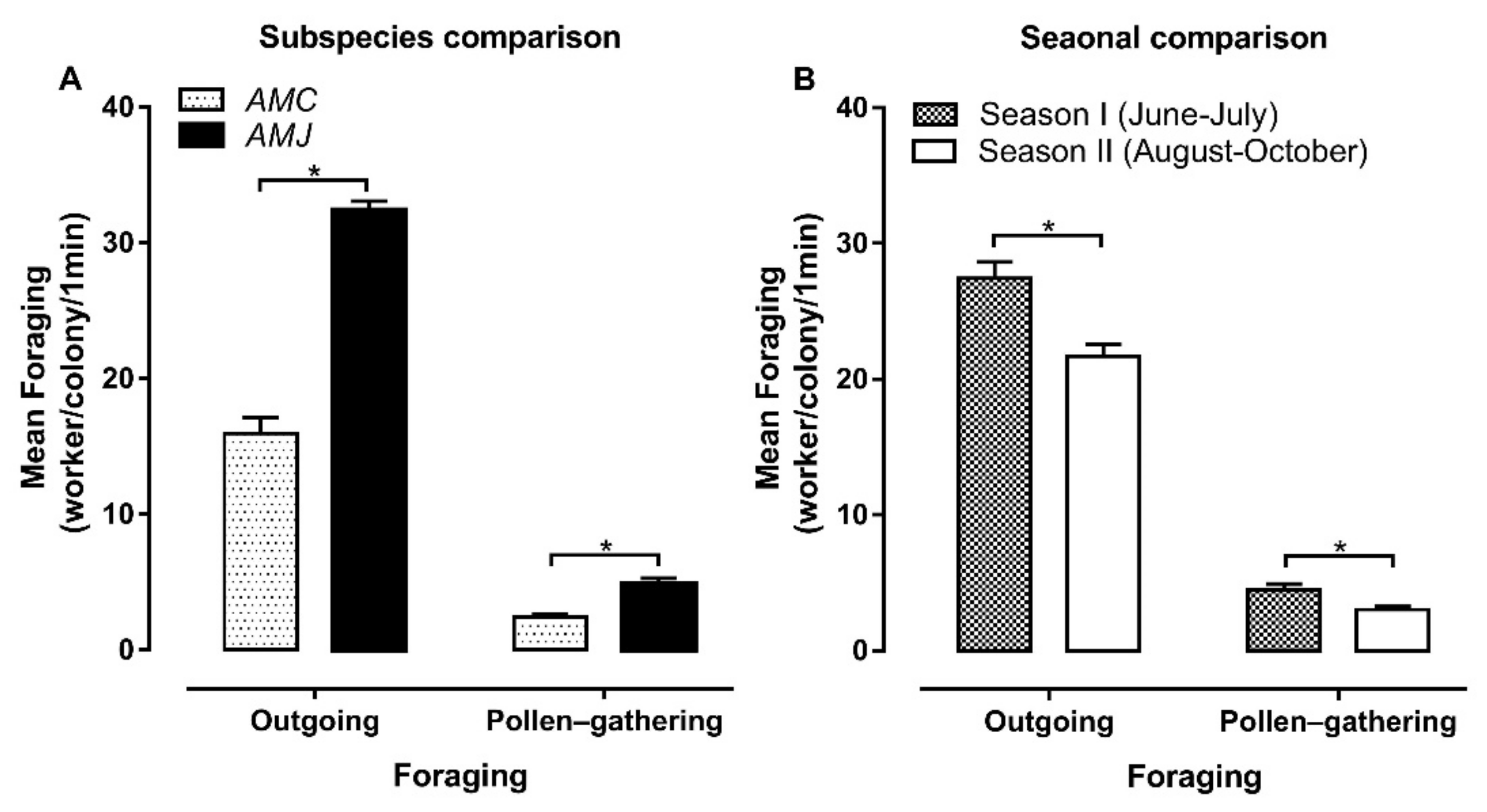
3.3.1. Outgoing Foraging
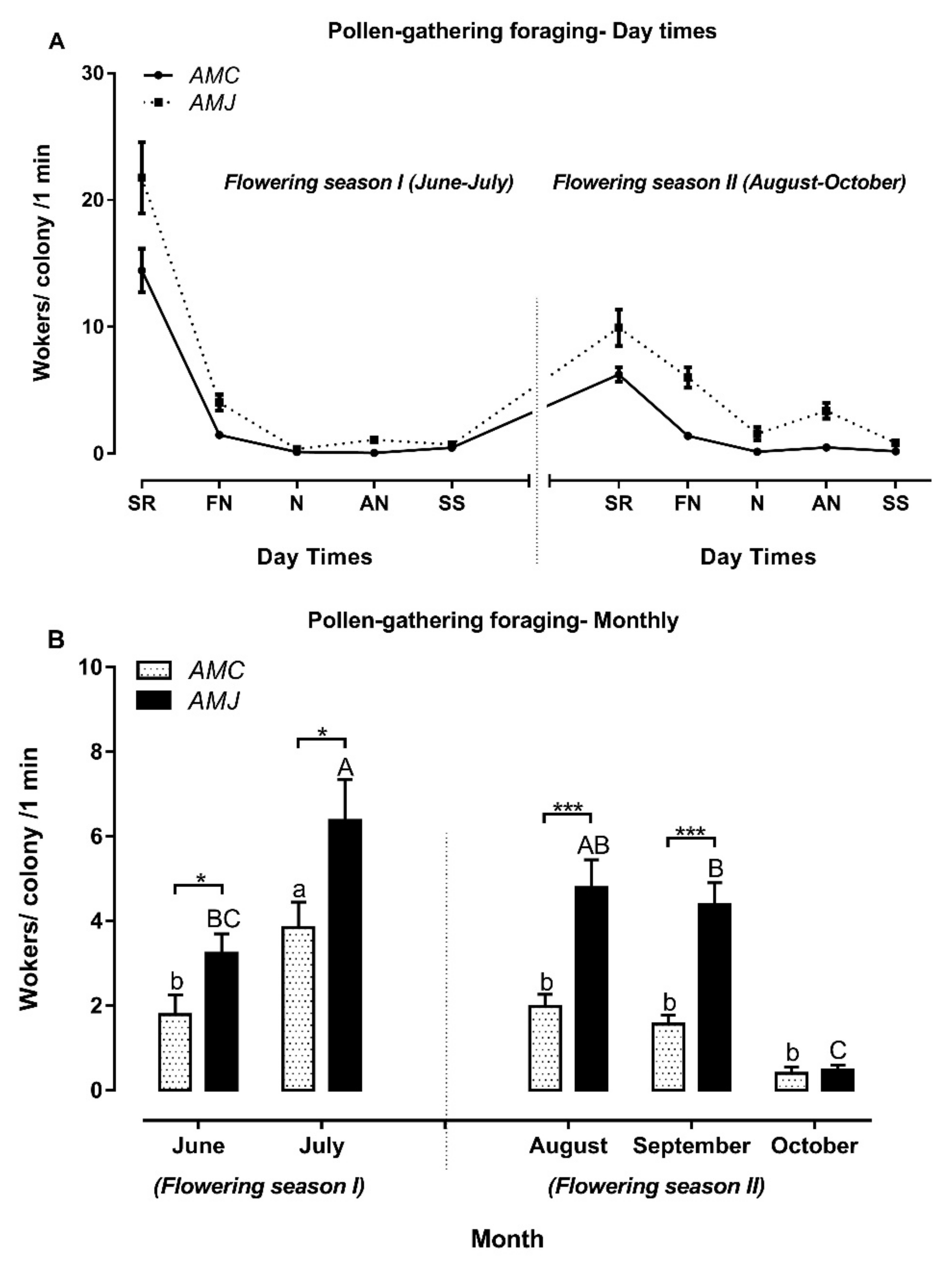
3.3.2. Pollen-Gathering Foraging
3.4. Correlation Analysis
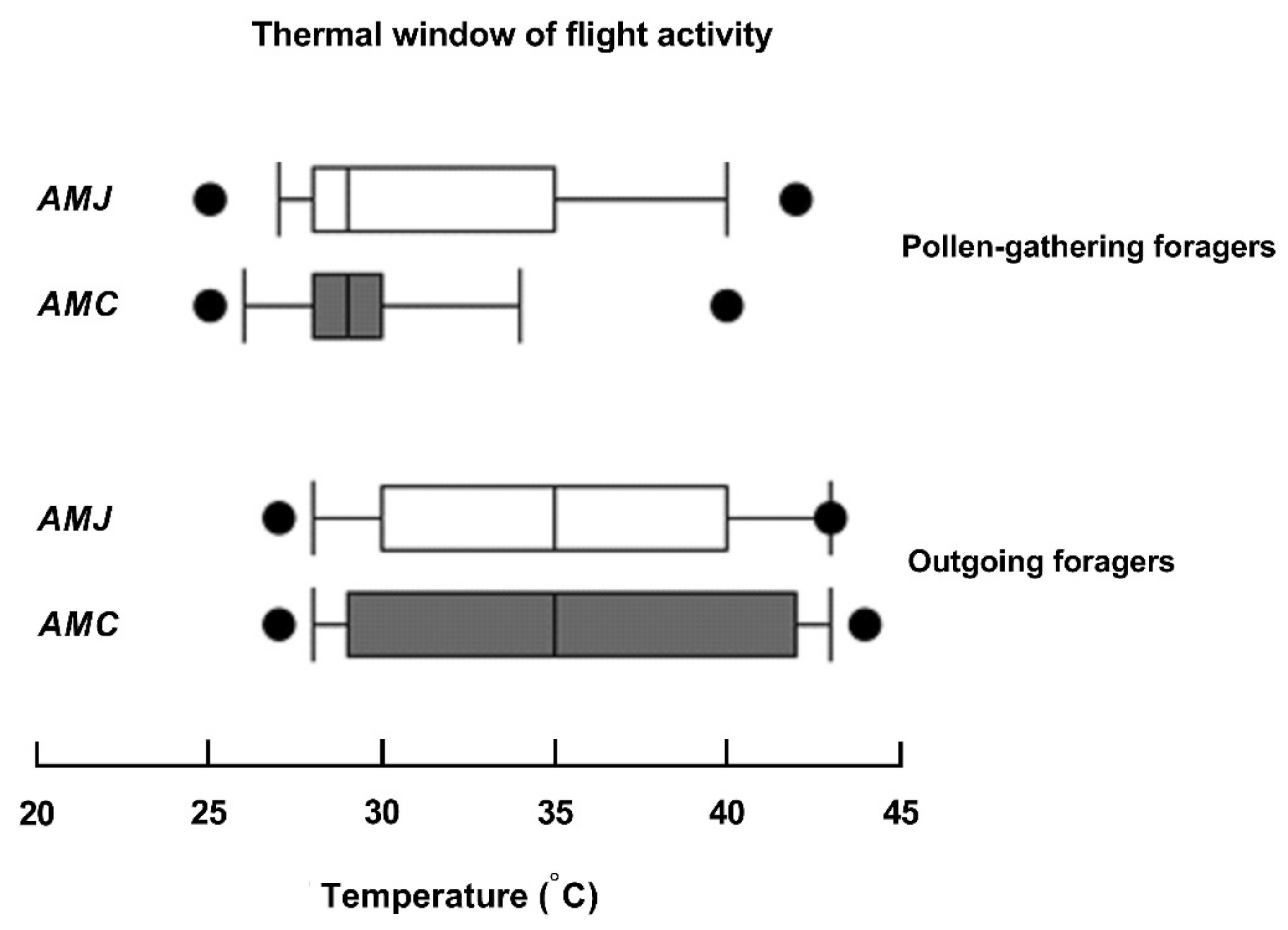
3.5. Thermal Windows of Flight Activity
4. Discussion
4.1. Flowering Density
4.2. Foraging Activities
4.3. Effect of Weather on Foraging
4.3.1. Foraging and Time of Day
4.3.2. Foraging and Flowering Season
4.4. Correlation and Thermal Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Attfield, H.H.D. A Beekeeping Guide for the Tropics and Subtropics; Vita Publication: Washington, DC, USA, 2001. [Google Scholar]
- Alqarni, A.S. Honeybee foraging, nectar secretion, and honey potential of wild jujube trees, Ziziphus nummularia. Neotrop. Entomol. 2015, 44, 232–241. [Google Scholar] [CrossRef]
- Hassan, A.M.A.; Giovanetti, M.; Raweh, H.S.A.; Owayss, A.A.; Ansari, M.J.; Nuru, A.; Radloff, S.E.; Alqarni, A.S. Nectar secretion dynamics of Ziziphus nummularia: A melliferous species of dry land ecosystems. Saudi J. Biol. Sci. 2017, 24, 1470–1474. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K. Rhamnaceae. In Mansfeld’s Encyclopedia of Agricultural and Horticultural Crops; Mansfeld, R., Hanelt, P., Institute of Plant Genetics and Crop Plant Research, Eds.; Springer: Berlin, Germany, 2001; pp. 1141–1150. [Google Scholar]
- Pandey, A.; Singh, R.; Radhamaniand, J.; Bhandari, D.C. Exploring the potential of Ziziphus nummularia (Burm. f.) Wight et Arn. from drier regions of India. Genet. Resour. Crop Evol. 2010, 57, 929–936. [Google Scholar] [CrossRef]
- Owayss, A.A. Diversity of insect floral visitors and their role in pollination of wild jujube, Ziziphus nummularia (Burm. F. Wight et Arn.) in hot-dry environment. Bull. Entomol. Soc. Egypt 2015, 92, 23–35. [Google Scholar]
- Adgaba, N.; Awad, A.M.; Al-Ghamdi, A.A.; Alqarni, A.S.; Radloff, S.E. Nectar of Ziziphus spina-christi (L.) willd (Rhamnaceae): Dynamics of secretion and potential for honey production. J. Apic. Sci. 2012, 56, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Tel-Zur, N.; Schenider, B. Floral biology of Ziziphus mauritiana (Rhamnaceae). Sex. Plant Reprod. 2009, 22, 73–85. [Google Scholar] [CrossRef]
- Singh, M.; Dwivedi, R.N. Farmers preferences on tree/crop species and livestock feeding—A study through pra approach. Progress. Agric. 2002, 2, 135–137. [Google Scholar]
- Shokrollahi, A.; Alizadeh, A.; Malekhosseini, Z.; Ranjbar, M. Removal of Bromocresol Green from aqueous solution via adsorption on Ziziphus nummularia as a new, natural, and low-cost adsorbent: Kinetic and thermodynamic study of removal process. J. Chem. Eng. Data 2011, 56, 3738–3746. [Google Scholar] [CrossRef]
- Chanda, S.; Kaneria, M.; Vaghasiya, Y.K. Evaluation of antimicrobial potential of some Indian medicinal plants against some pathogenic microbes. Indian J. Nat. Prod. Resour. 2011, 2, 225–228. [Google Scholar]
- Arndt, S.K.; Clifford, S.C.; Popp, M. Ziziphus—A Multipurpose Fruit Tree for Arid Regions. In Sustainable Land Use in Deserts; Springer: Berlin/Heidelberg, Germany, 2001; pp. 388–399. [Google Scholar]
- Saied, A.S.; Gebauer, J.; Hammer, K.; Buerkert, A. Ziziphus spina-christi (L.) Willd.: A multipurpose fruit tree. Genet. Resour. Crop Evol. 2008, 55, 929–937. [Google Scholar] [CrossRef]
- Bachaya, H.A.; Iqbal, Z.; Khan, M.N.; Sindhu, Z.U.; Jabbar, A. Anthelmintic activity of Ziziphus nummularia (bark) and Acacia nilotica (fruit) against Trichostrongylid nematodes of sheep. J. Ethnopharmacol. 2009, 123, 325–329. [Google Scholar] [CrossRef]
- Shah, A.H.; Tariq, M.; Al-Yahya, M.A. Studies on the alkaloidal fraction from the stem bark of Zizyphus nummularia. Fitoterapia 1990, 61, 452–469. [Google Scholar]
- Alfarhan, A.H. A floristic account on Raudhat Khuraim central province Saudi Arabia. Saudi J. Biol. Sci. 2001, 8, 80–103. [Google Scholar]
- Al-Farraj, M.M.; Al-Farhan, A.; Al-Yemeni, M. Ecological studies on Rawdhat system in Saudi Arabia I-Rawdhat Khorim. Pak. J. Bot. 1997, 29, 75–88. [Google Scholar]
- Shaltout, K.H.; Mady, M.A. Current situation of the raudha’s woody plant populations in the Central Saudi Arabia. Feddes Repert. 1993, 104, 503–509. [Google Scholar] [CrossRef]
- Brunet, J.; Sweet, H.R. Impact of insect pollinator group and floral display size on outcrossing rate. Evolution 2006, 60, 234–246. [Google Scholar] [CrossRef]
- Gómez, J.M.; Bosch, J.; Perfectti, F.; Fernández, J.; Abdelaziz, M. Pollinator diversity affects plant reproduction and recruitment: The tradeoffs of generalization. Oecologia 2007, 153, 597–605. [Google Scholar] [CrossRef]
- Awad, A.M.; Owayss, A.A.; Alqarni, A.S. Performance of two honey bee subspecies during harsh weather and Acacia gerrardii nectar-rich flow. Sci. Agric. 2017, 74, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Alqarni, A.S. Differential foraging of indigenous and exotic honeybee (Apis mellifera L.) races on nectar-rich flow in a subtropical ecosystem. Insects 2020, 11, 254. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shaara, H.F.; Owayss, A.A.; Ibrahim, Y.Y.; Basuny, N.K. A review of impacts of temperature and relative humidity on various activities of honey bees. Insectes Soc. 2017, 64, 455–463. [Google Scholar] [CrossRef]
- Nicolson, S.W.; Nepi, M.; Pacini, E. Nectaries and Nectar; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Langernberger, M.W.; Davis, A.R. Temporal changes in floral nectar production, reabsorption, and compostion associated with dichogamy in annual caraway (Carum carvi: Apiaceae). Am. J. Bot. 2002, 89, 1588–1598. [Google Scholar] [CrossRef]
- Rathcke, B.J. Nectar distributions, pollinator behavior, and plant reproductive success. In Effects of Resource Distribution on Animal–Plant Interactions; Hunter, M.D., Ohgushi, T., Price, P.W., Eds.; Academic Press: Boston, MA, USA, 1992; pp. 113–138. [Google Scholar]
- Boisvert, M.J.; Veal, A.J.; Sherry, D.F. Floral reward production is timed by an insect pollinator. Proc. R. Soc. B 2007, 274, 1831–1837. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.S.; McNally, L.C. Seasonal patterns of foraging activity in colonies of the African honey bee, Apis mellifera scutellata, in Africa. Insectes Soc. 1992, 39, 181–193. [Google Scholar] [CrossRef]
- Alqarni, A.S. Tolerance of summer temperature in imported and indigenous honeybee Apis mellifera L. races in central Saudi Arabia. Saudi J. Biol. Sci. 2006, 13, 123–127. [Google Scholar]
- Corbet, S.A.; Fussell, M.; Ake, R.; Fraser, A.; Gunsen, C.; Salvage, A.; Smithy, K. Temperature and pollination activity of social bees. Ecol. Entomol. 1993, 18, 17–30. [Google Scholar] [CrossRef]
- Owayss, A.A.; Elbanna, K.; Iqbal, J.; Abulreesh, H.H.; Organji, S.R.; Raweh, H.S.; Alqarni, A.S. In vitro antimicrobial activities of Saudi honeys originating from Ziziphus spina-christi L. and Acacia gerrardii Benth. trees. Food Sci. Nutr. 2020, 8, 390–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alqarni, A.S.; Balhareth, H.M.; Owayss, A.A. Performance evaluation of indigenous and exotic honey bee (Apis mellifera L.) races in Assir region, southwestern Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Beekman, M.; Sumpter, D.J.T.; Seraphides, N.; Ratnieks, F.L.W. Comparing foraging behaviour of small and large honey-bee colonies by decoding waggle dances made by foragers. Funct. Ecol. 2004, 18, 829–835. [Google Scholar] [CrossRef]
- Ali, M.A.M. Comparative study for evaluating two honey bee races, Apis mellifera jementica (indigenous race) and Apis mellifera carnica (carniolan race) in brood production, population development and foraging activity under the environmental conditions of the central region of the Kingdom of Saudi Arabia. Ann. Agric. Sci. 2011, 56, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Malerbo-Souza, D.T. The corn pollen as a food source for honeybees. Acta Sci. Agron. 2011, 33, 701–704. [Google Scholar] [CrossRef]
- Pearce, F.C.R.; Couvillon, M.J.; Ratnieks, F.L.W. Hive relocation does not adversely affect honey bee (Hymenoptera: Apidae) foraging. Psyche 2013, 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Woyke, J. Diurnal flight activity of African bees Apis mellifera adansonii in different seasons and zones of Ghana. Apidologie 1992, 23, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Dukku, U.; Russom, Z.; Domo, A.G. Diurnal and seasonal flight activity of the honeybee, Apis Mellifera L, and its relationship with temperature, light intensity and relative Humidity in the Savanna of Northern Nigeria. Glob. J. Sci. Front. Res. 2013, 13, 1–7. [Google Scholar]
- Alatar, A.A.; El-Sheikh, M.A.R.; Thomas, J.; Hegazy, A.K.; El Adawy, H.A. Vegetation, floristic diversity, and size-classes of Acacia gerrardii in an arid wadi ecosystem. Arid Land Res. Manag. 2015, 29, 335–359. [Google Scholar] [CrossRef]
- Hassan, A.M.A. Ecological Studies on Honey Bee Apis Mellifera L. Activities on Talh Trees Acacia Gerrardii (Benth.) and Use of Geographic Information Systems and Remote Sensing Technology to Assess Foraging Capacity for Honey Production. PhD Thesis, College of Food and Agriculture Sciences, King Saud University, Riyadh, Saudi Arabia, 2016. [Google Scholar]
- Silva, D.P.; Moisan-De Serres, J.; Souza, D.R.; Hilgert-Moreira, S.B.; Fernandes, M.Z.; Kevan, P.G.; Freitas, B.M. Efficiency in pollen foraging by honey bees: Time, motion and pollen depletion on flowers of Sisyrinchium palmifolium Linnaeus (Asparagales: Iridaceae). J. Pollinat. Ecol. 2013, 11, 27–32. [Google Scholar] [CrossRef]
- Adgaba, N.; Al-Ghamdi, A.; Tadesse, Y.; Getachew, A.; Awad, A.M.; Ansari, M.J.; Owayss, A.A.; Mohammed, S.E.A.; Alqarni, A.S. Nectar secretion dynamics and honey production potentials of some major honey plants in Saudi Arabia. Saudi J. Biol. Sci. 2017, 24, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Alqarni, A.S.; Hannan, M.A.; Owayss, A.A.; Engel, M.S. The indigenous honey bees of Saudi Arabia (Hymenoptera, Apidae, Apis mellifera jemenitica Ruttner): Their natural history and role in beekeeping. Zookeys 2011, 134, 83–98. [Google Scholar] [CrossRef] [Green Version]
- Abou-Shaara, H.F.; Al-Ghamdi, A.A.; Mohamed, A.A. Tolerance of two honey bee races to various temperature and relative humidity gradients. Environ. Exp. Bio. 2012, 10, 133–138. [Google Scholar]
- Pacini, E.; Nepi, M.; Vesprini, J.L. Nectar biodiversity: A short review. Plant Syst. Evol. 2003, 238, 7–21. [Google Scholar] [CrossRef]
- Alqarni, A.S.; Hassan, A.; Owayss, A. Evaluation of Acacia gerrardii benth. (Fabaceae: Mimosoideae) as a honey plant under extremely hot-dry conditions: Flowering phenology, nectar yield and honey potentiality. J. Anim. Plant Sci. 2015, 25, 1667–1674. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subspecies | Foraging Activity | Correlation Coefficient (r) and Significance | |||
|---|---|---|---|---|---|
| Temp. a | RH b | WS c | Outgoing | ||
| AMC | Outgoing | −0.0793 ** | 0.1105 ** | 0.1032 ** | - |
| Pollen-gathering | −0.1665 *** | 0.1661 ** | −0.0442 | 0.5120 ** | |
| AMJ | Outgoing | 0.0760 ** | −0.0662 | 0.0748 * | - |
| Pollen-gathering | −0.1307 ** | 0.1303 ** | −0.0629 | 0.5039 ** | |
| Weather factors | Temp. a | −7.993 ** | 0.5412 ** | ||
| RH b | −7.993 ** | −0.3786 ** | |||
| WS c | 0.5412 ** | −0.3786 ** | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqarni, A.S.; Iqbal, J.; Raweh, H.S.; Hassan, A.M.A.; Owayss, A.A. Beekeeping in the Desert: Foraging Activities of Honey Bee during Major Honeyflow in a Hot-Arid Ecosystem. Appl. Sci. 2021, 11, 9756. https://doi.org/10.3390/app11209756
Alqarni AS, Iqbal J, Raweh HS, Hassan AMA, Owayss AA. Beekeeping in the Desert: Foraging Activities of Honey Bee during Major Honeyflow in a Hot-Arid Ecosystem. Applied Sciences. 2021; 11(20):9756. https://doi.org/10.3390/app11209756
Chicago/Turabian StyleAlqarni, Abdulaziz S., Javaid Iqbal, Hael S. Raweh, Awad M. A. Hassan, and Ayman A. Owayss. 2021. "Beekeeping in the Desert: Foraging Activities of Honey Bee during Major Honeyflow in a Hot-Arid Ecosystem" Applied Sciences 11, no. 20: 9756. https://doi.org/10.3390/app11209756