
Neuronutraceuticals Modulate Lipopolysaccharide- or Amyloid-β 1-42 Peptide-Induced Transglutaminase 2 Overexpression as a Marker of Neuroinflammation in Mouse Microglial Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Mouse Primary Microglia Isolation
2.4. Western Blot Analysis
2.5. RNA Extraction and Semi-Quantitative RT-qPCR Analysis
2.6. Statistical Analysis
3. Results
3.1. Effects of Curcumin and PEA on LPS-Induced TG2 Expression Levels in BV2 Cells
3.1.1. Effects of Curcumin and PEA on LPS-Induced TG2 RNA Expression Levels in BV2 Cells
3.1.2. TG2 Protein Levels in BV2 Cells After Treatment with Neuronutraceuticals and LPS
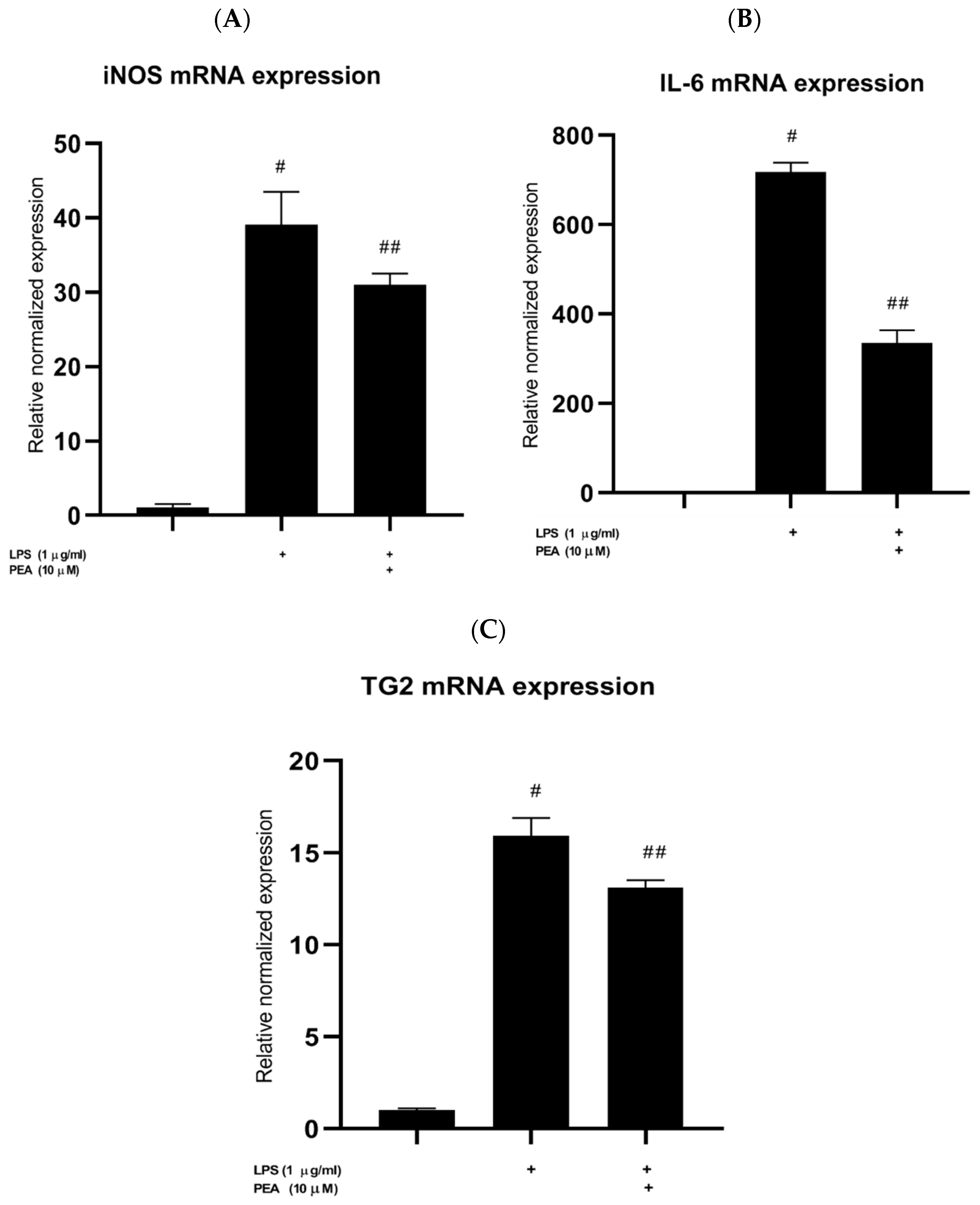
3.1.3. Effects of PEA on LPS-Induced TG2, Inflammatory Markers RNA Expression Levels in Murine Primary Microglial Cells
3.2. Effects of Curcumin and PEA on Aβ1-42-Induced TG2, Inflammatory or Neuroprotective Markers Expression Levels in BV2 Cells
3.2.1. Effects of Aβ1-42 on TG2 Protein Expression in BV2 Cells
3.2.2. Effects of Curcumin and PEA on Aβ1-42-Induced TG2, Inflammatory or Neuroprotective Markers RNA Expression Levels in BV2 Cells
3.2.3. TG2 Protein Levels in BV2 Cells after Treatment with Neuronutraceuticals and Aβ1-42
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Folk, J.E. Transglutaminases. Annu. Rev. Biochem. 1980, 49, 517–531. [Google Scholar] [CrossRef]
- Lorand, L.; Conrad, S.M. Transglutaminases. Mol. Cell Biol. 1984, 58, 9–35. [Google Scholar]
- Thomazy, V.; Fesus, L. Differential expression of tissue transglutaminase in human cells: An immunohistochemical study. Cell Tissue Res. 1989, 255, 215–224. [Google Scholar] [CrossRef]
- Fesus, L.; Thomazy, V.; Falus, A. Induction and activation of tissue transglutaminase during programmed cell death. FEBS (Fed. Eur. Biochem. Soc.) Lett. 1987, 224, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Piacentini, M.; Autuori, F.; Dini, L.; Farrace, M.G.; Ghibelli, L.; Piredda, L.; Fesus, L. “Tissue” transglutaminase is specifically expressed in neonatal rat liver cells undergoing apoptosis upon epidermal growth factor stimulation. Cell Tissue Res. 1991, 263, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Fesus, L.; Davies, P.J.A.; Piacentini, M. Apoptosis: Molecular mechanisms in programmed cell death. Eur. J. Cell Biol. 1992, 56, 170–177. [Google Scholar]
- Fesus, L.; Tarcsa, E. Formation of N-ε−(γ-glutamyl)-lysine isodipeptide in chinese hamster ovary cells. Biochem. J. 1989, 263, 843–848. [Google Scholar] [CrossRef]
- Melino, G.; Annichiarico-Petruzzelli, M.; Piredda, L.; Candi, E.; Gentile, V.; Davies, P.J.A.; Piacentini, M. Tissue transglutaminase and apoptosis: Sense and antisense transfection studies with human neuroblastoma cells. Mol. Cell. Biol. 1994, 14, 6584–6596. [Google Scholar] [CrossRef] [Green Version]
- Bowness, J.M.; Folk, J.E.; Timpl, R. Identification of a substrate site for liver transglutaminase on the aminopropeptide of type III collagen. J. Biol. Chem. 1987, 262, 1022–1024. [Google Scholar] [CrossRef]
- Slife, C.W.; Dorsett, M.D.; Tillotson, M.L. Subcellular localization and a large molecular weight substrate for the liver plasma membrane transglutaminase. J. Biol. Chem. 1986, 261, 3451–3456. [Google Scholar] [CrossRef]
- Tyrrel, D.J.; Sale, W.S.; Slife, C.W. Fibronectin is a component of the sodium dodecyl sulfate-insoluble transglutaminase substrate. J. Biol. Chem. 1988, 263, 8464–8469. [Google Scholar] [CrossRef]
- Kinsella, M.G.; Wight, T.N. Modulation of sulfated proteoglycan synthesis by bovine aortic endothelial cells during migration. J. Cell Biol. 1986, 162, 679–687. [Google Scholar] [CrossRef]
- Kinsella, M.G.; Wight, T.N. Formation of high weight dermatan sulfate proteoglycan in bovine aortic endothelial cell cultures. J. Biol. Chem. 1990, 265, 17891–17898. [Google Scholar] [CrossRef]
- Fesus, L.; Metsis, M.L.; Muszbek, L.; Koteliansky, V.E. Transglutaminase-sensitive glutamine residues of human plasma fibronectin revealed by studying its proteolytic fragments. Eur. J. Biochem. 1986, 154, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.; Rich, E.; Barsigian, C. Transglutaminase-mediated cross-linking of fibrinogen by human umbilical vein endothelial cells. J. Biol. Chem. 1989, 264, 20502–20508. [Google Scholar] [CrossRef]
- Sane, D.C.; Moser, T.L.; Greenberg, C.S. Vitronectin in the substratum of endothelial cells is cross-linked and phosphorylated. Biochem. Biophys. Res. Commun. 1991, 174, 465–469. [Google Scholar] [CrossRef]
- Gentile, V.; Thomazy, V.; Piacentini, M.; Fesus, L.; Davies, P.J.A. Expression of tissue transglutaminase in Balb-C 3T3 fibroblasts: Effects on cellular morphology and adhesion. J. Cell Biol. 1992, 119, 463–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, S.; Huang, W.; Bennett, D.A. Tissue transglutaminase and its product isopeptide are increased in Alzheimer’s disease and APPswe/PS1dE9 double transgenic mice brains. Mol. Neurobiol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelmus, M.M.; de Jager, M.; Smit, A.B.; van der Loo, R.J.; Drukarch, B. Catalytically active tissue transglutaminase colocalises with Aβ pathology in Alzheimer’s disease mouse models. Sci. Rep. 2016. [Google Scholar] [CrossRef] [Green Version]
- Ientile, R.; Currò, M.; Caccamo, D. Transglutaminase 2 and neuroinflammation. Amino Acids 2015, 47, 19–26. [Google Scholar] [CrossRef]
- Chih-Li, L.; Jen-Kun, L. Curcumin: A potential cancer chemopreventive agent through suppressing NF-kB signaling. J. Cancer Mol. 2008, 4, 11–16. [Google Scholar]
- Lee, J.; Kim, Y.S.; Choi, D.H.; Bang, M.S.; Han, T.R.; Joh, T.H.; Kim, S.Y. Transglutaminase 2 induces nuclear factor-kB activation via a novel pathway in BV-2 microglia. J. Biol. Chem. 2004, 279, 53725–53735. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Mehta, K. Tissue transglutaminase constitutively activates HIF-1a promoter and nuclear factor-kB via a non-canonical pathway. PLoS ONE 2012, 7, e49321. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Saydak, M.; Gentile, V.; Stein, J.P.; Davies, P.J.A. Isolation and characterization of the human tissue transglutaminase promoter. J. Biol. Chem. 1995, 270, 9748–9755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatta, N.G.; Cammarota, G.; Iannaccone, M.; Serretiello, E.; Gentile, V. Curcumin (Diferulolylmethane) reduces transglutaminase 2 overexpression induced by retinoic acid in human nervous cell lines. Neuroimmunomodulation 2016, 23, 188–193. [Google Scholar] [CrossRef]
- Alessio, N.; Belardo, C.; Trotta, M.C.; Paino, S.; Boccella, S.; Gargano, F.; Pieretti, G.; Ricciardi, F.; Marabese, I.; Luongo, L.; et al. Vitamin D Deficiency Induces Chronic Pain and Microglial Phenotypic Changes in Mice. Int. J. Mol. Sci. 2021, 22, 3604. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Luongo, L.; Boccella, S.; Giordano, M.E.; Romano, R.; Bellini, G.; Manzo, I.; Furiano, A.; Rizzo, A.; Imperatore, R.; et al. Palmitoylethanolamide induces microglia changes associated with increased migration and phagocytic activity: Involvement of the CB2 receptor. Sci. Rep. 2017, 7, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Boccella, S.; Iannotta, M.; De Gregorio, D.; Giordano, C.; Belardo, C.; Romano, R.; Palazzo, E.; Scafuro, M.A.; Serra, N.; et al. Palmitoylethanolamide Reduces Neuropsychiatric Behaviors by Restoring Cortical Electrophysiological Activity in a Mouse Model of Mild Traumatic Brain Injury. Front Pharmacol. 2017, 8, 95–119. [Google Scholar] [CrossRef] [Green Version]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The nuclear receptor peroxisome proliferator-activated receptor-alpha mediates the anti-inflammatory actions of palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes and Reference Sequence Data for Nucleotide Sequences | Forward | Reverse |
|---|---|---|
| TG2 NM_009373.3 | 5′-ACTTCGACGTGTTTGCCCACAT-3′ | 5′-TTGATGTCCTCAGTGCCACACT-3′ |
| IL6 NM_031168.2 | 5′-GATGGATGCTACCAAACTGGAT- 3′ | 5′-CCAGGTAGCTATGGTACTCCAGA-3′ |
| iNOS NM_010927.4 | 5′-TGAGCTCATCTTTGCCACCA-3′ | 5′-ACAGTTCCGAGCGTCAAAGA-3′ |
| IL-1β NM_008361.4 | 5′-TCGGACCCATATGAGCTGAAAG-3′ | 5′-CCACAGGTATTTTGTCGTTGCT-3′ |
| TREM2 NM_031254.3 | 5′-AGCCTGACTGGCTTGGTCAT-3′ | 5′-CCCAGTGCTTCAAGGCGTCA-3′ |
| ARG1 NM_007482.3 | 5′-CAGCCTCGAGGAGGGGTAGA-3′ | 5′-CCCGTGGTCTCTCACGTCAT-3′ |
| GAPDH NM_001289726.1 | 5′-GGGCATCTTGGGCTACACTGAGGACC-3′ | 5′-GGGGGCCGAGTTGGGATAGGG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatta, N.G.; Parente, A.; Guida, F.; Maione, S.; Gentile, V. Neuronutraceuticals Modulate Lipopolysaccharide- or Amyloid-β 1-42 Peptide-Induced Transglutaminase 2 Overexpression as a Marker of Neuroinflammation in Mouse Microglial Cells. Appl. Sci. 2021, 11, 5718. https://doi.org/10.3390/app11125718
Gatta NG, Parente A, Guida F, Maione S, Gentile V. Neuronutraceuticals Modulate Lipopolysaccharide- or Amyloid-β 1-42 Peptide-Induced Transglutaminase 2 Overexpression as a Marker of Neuroinflammation in Mouse Microglial Cells. Applied Sciences. 2021; 11(12):5718. https://doi.org/10.3390/app11125718
Chicago/Turabian StyleGatta, Nicola Gaetano, Andrea Parente, Francesca Guida, Sabatino Maione, and Vittorio Gentile. 2021. "Neuronutraceuticals Modulate Lipopolysaccharide- or Amyloid-β 1-42 Peptide-Induced Transglutaminase 2 Overexpression as a Marker of Neuroinflammation in Mouse Microglial Cells" Applied Sciences 11, no. 12: 5718. https://doi.org/10.3390/app11125718