1. Introduction
Ultraviolet B (UVB)-irradiation (280–320 nm) primarily induces cell damage by augmenting intracellular reactive oxygen species (ROS) levels. The radiation easily penetrates the stratum corneum, where it increases the risk of photoaging, with symptoms characterized by thickening of the epidermis, discoloration, skin wrinkling, loss of elasticity and skin cell growth retardation [
1]. Recent developments in marine bioresource technology have substantiated the therapeutic potential of marine natural products to be used as cosmeceuticals, functional foods, new drugs and drug leads. Fucoidan is a fucose-rich sulfated polysaccharide found in brown algae that is renowned for its versatile biologic activities. High molecular weight fucoidans closely resemble the structure of heparin and are effective anticoagulants [
2]. Nevertheless, low molecular weight fucoidans are reported to possess antioxidant, anti-inflammatory, anticancer and anti-microbial activities [
3,
4]. Fucoidan mainly contains α-1,3 and α-1,4-linked fucose units with sulfate ester substituents at the 2, 3 and/or 4 positions. However, the chains could also contain monosaccharides such as glucose, mannose, xylose, arabinose and hexuronic acid. The monomer composition, connectivity, branching, sulfation pattern, and molecular weight are species-related [
2]. The conventional method of refining fucoidan involves hot water extraction of dry algal powder under mildly acidic pH, followed by ethanol precipitation. The precipitate undergoes additional fractionation via ultrafiltration, size exclusion or anion exchange chromatography. This method lacks efficiency based on the low extraction yield, convenience, time consumption and selectivity over molecular weights.
As fucoidan is mainly localized to the cell wall of brown algae, degradation of the cell wall using enzymes is a well-planned extraction strategy conserving polymer integrity, and thereby its biofunctional properties. The use of enzymes, including Celluclast, Termamyl, Ultraflo, Amyloglucosidase, and Viscozyme, is advisable to obtain fucoidan whose biofunctional properties are preserved [
5]. Gradient alcohol precipitation is a fractionation method used for fractionating polysaccharides with gradually narrowed down molecular weight distributions [
6]. It is a relatively inexpensive and simple method compared to ultrafiltration and chromatography. The precipitation is achieved by incorporating organic solvents such as methanol, ethanol, isopropanol, 1-butanol and acetone or concentrated inorganic salts such as ammonium sulfate ((NH
4)
2SO
4) and polymerizing agents such as polyethylene glycol (PEG). The use of this method has been previously reported for fractionating fucoidan [
7]. The present study was undertaken as a part of a project to investigate possible industrial uses of
S. horneri as a sustainable approach for managing its large biomass. The extraction of fucoidan enriched crude polysaccharides followed an optimized green approach using enzymes, while the fractionation followed a step gradient ethanol precipitation.
2. Materials and Methods
Fucoidan standard, KBr (FTIR grade), deuterium oxide, 2′,7′-dichlorodihydrofluorescein diacetate (DCFH2-DA) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), o-Toluidine blue, trifluoroacetic acid and 2-mercaptoethanolwere purchased from Sigma-Aldrich (St. Louis, MO, USA). Celluclast was obtained from Novozyme Co. (Bagsvaerd, Denmark). Chloroform, methanol and ethanol were of analytical grade. Dulbecco’s Modified Eagle Medium (DMEM), fetal bovine serum (FBS) and penicillin/streptomycin mixture were purchased from GIBCO INC., (Grand Island, NY, USA). Primary and ary antibodies were purchased from Cell Signalling Technology, Inc. (Beverly, MA, USA) and Santa Cruz Biotechnology (Santa Cruz, CA, USA).
2.1. Preparation of Fucoidan Fraction
2.1.1. Extraction of Fucoidan Enriched Polysaccharide
Washed and dried S. horneri samples collected off Jeju coast were provided to us by Seojin Biotech Company limited. The samples were pulverized using an MF 10 basic, IKA microfine grinder (Werke, Germany). Depigmentation was carried out using a solvent system of chloroform and methanol 1:1. Next, the dried powder was soaked in a solution of ethanol containing 10% formaldehyde for 3 h at 40 °C. The dried powder was washed twice with 80% ethanol. After evaporating off any remaining solvent, the sample powder was suspended in 5 L of deionized water at a 1:10 (kg/L) ratio. The pH was adjusted to 4.5 by adding diluted HCl while equilibrating at 50 °C in a shaking incubator for 1 h. Celluclast was added at a 0.5% sample ratio and kept for 8 h under continuous agitation at 50 °C. The mixture was filtered through a muslin cloth. Celluclast was heat-denatured at 100 °C for 10 min. The extract was neutralized at room temperature by adding diluted NaOH and centrifuged at 5000× g for 20 min to remove unfiltered particles. The supernatant (4.5 L) was frozen and lyophilized to reduce the volume to 1 L.
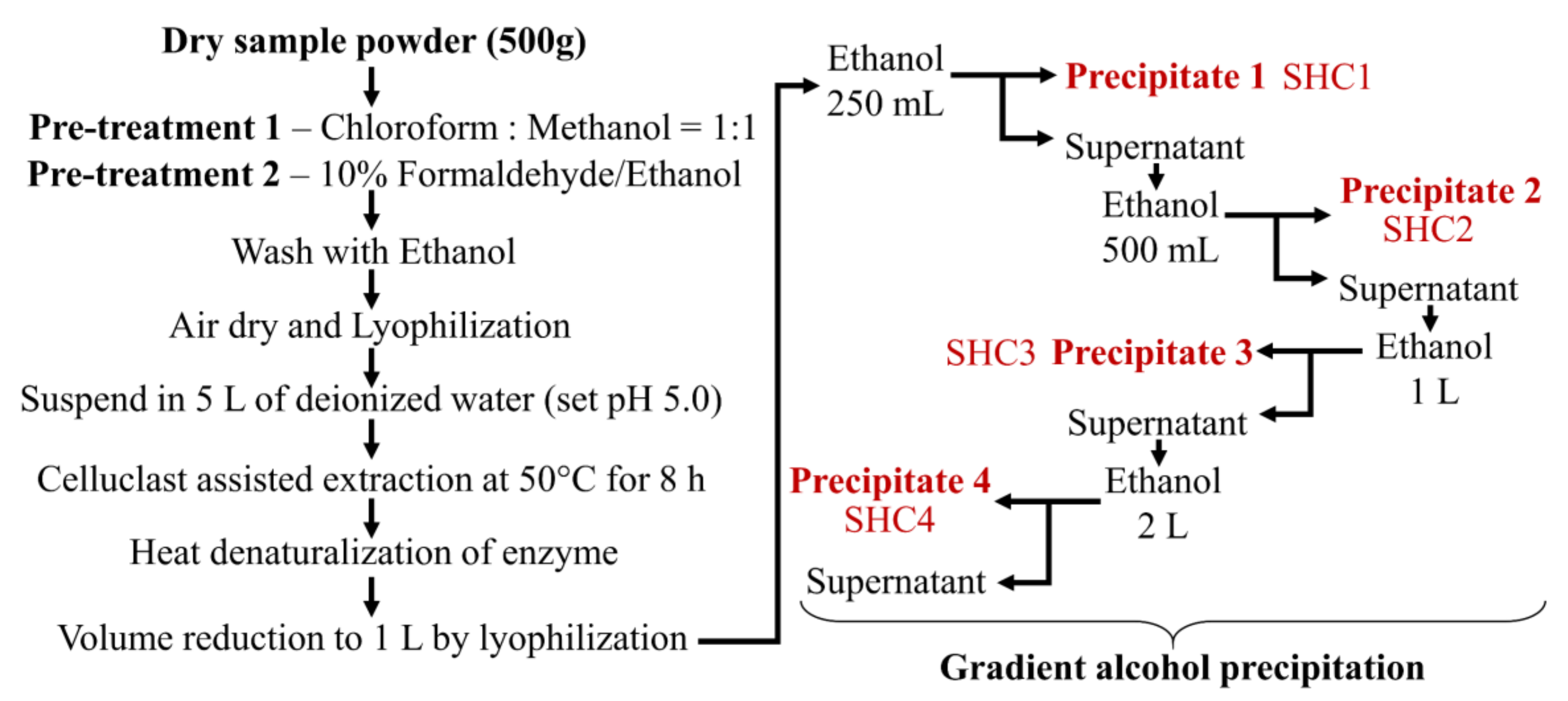
2.1.2. Step Gradient Ethanol Precipitation
The ratio of ethanol was determined following optimization studies. As the first step in gradient ethanol precipitation, 250 mL of ethanol was gently added to 1 L of the extract while stirring. The mixture was incubated at 4 °C for 12 h, allowing it to equilibrate while precipitating the polysaccharides (
Figure 1). After, the mixture was centrifuged at 5000×
g for 20 min at 4 °C to obtain the first precipitate designated as SHC1. Sequentially the second, third and fourth precipitates were collected by, respectively adding 500 mL, 1 L and 2 L of ethanol to the supernatant after each precipitation step. All precipitates were dually washed with 95% ethanol (homogenization) and centrifuged to recover the polymer. Finally, the precipitates were dissolved in deionized water and dialyzed using 3.5-kD molecular weight cutoff dialysis membranes (Spectra/Por, Los Angeles, CA, USA). Polysaccharide fractions were lyophilized and stored at −20 °C for proceeding experiments.
2.2. Analysis of Molecular Weight (MW) Distribution
Approximate molecular weight distribution, homogeneity, and separation efficiency of the polysaccharide fractions were analyzed by an agarose gel electrophoresis method [
3]. Briefly, markers and samples (1 mg mL
−1) were electrophoresed in 1% agarose gels in Tris-Borate-EDTA running buffer (pH 8.3) at 100 V for 20 min. The gel was stained with 0.02% toluidine blue and 0.5% Triton X-100 in 3% acetic acid and de-stained with 3% acetic acid.
2.3. Fourier-Transform Infrared Spectroscopy (FTIR) and Monosaccharide Composition Analysis
Polysaccharide powders were cast into KBr pellets and analyzed by a VERTEX 70v FTIR spectrometer (Bruker, Germany) [
3]. For the monosaccharide composition analysis, polysaccharides were hydrolyzed with 4 M of trifluoroacetic acid and separated on a CarboPac PA1 column integrated to a Dionex ED50 Detector (HPAEC-PAD) (Dionex, Sunnyvale, CA, USA). A standardized monosaccharide mixture was used as the reference standard [
3].
2.4. H1 Nuclear Magnetic Resonance (NMR) Analysis
The selected polysaccharide fraction, SHC4, was deuterium exchanged by co-lyophilizing with deuterium oxide, dissolved in deuterium oxide, and analyzed by a JNM-ECX400, 400 MHz spectrometer (JEOL, Tokyo, Japan).
2.5. In Vivo Cell Culture
2.5.1. Cell Maintenance
HaCaT keratinocytes (Korean Cell Line Bank, Seoul, Korea) were maintained in DMEM containing 10% FBS and 1% penicillin-streptomycin. Cells were sub-cultured once every two days. Exponentially growing cells were used for seeding (2 × 105 cells mL−1). After 24 h, wells were treated with different sample concentrations and further incubated for 24 h. Cytotoxicity was measured by MTT assay. Briefly, cells were incubated with MTT for 4 h, dissolved in dimethyl sulfoxide (DMSO), and the absorbance readings were taken at 540 nm using a SpectraMax® M2 system (Molecular Devices, Sunnyvale, CA, USA).
2.5.2. UVB Exposure and Oxidative Stress Analysis
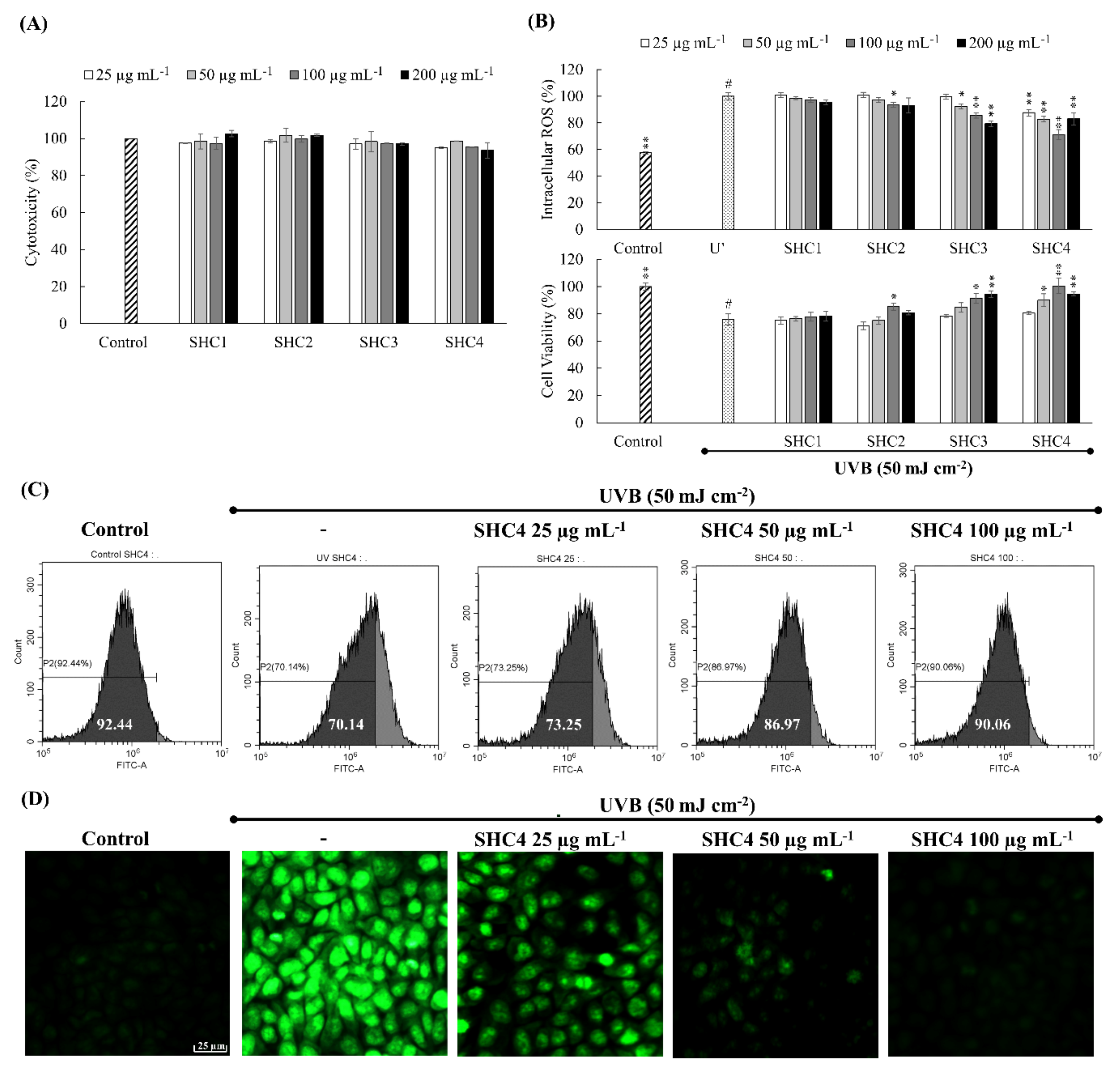
HaCaT keratinocytes were incubated for 2 h with different concentrations of samples. The cell culture media was withdrawn from the wells, once washed, and then filled with PBS. Wells except the control were exposed to UVB (50 mJ cm
−2) by a UVP CL-1000L ultraviolet cross-linker (Upland, CA, USA). Immediately after the irradiation, PBS was withdrawn, once washed, and replaced with serum-free culture media. Intracellular ROS levels were measured after incubating for an hour (DCFH2-DA assay) and cell viability was measured by MTT assay after incubating for 24 h [
1]. SHC4 that indicated superior antioxidant and cytoprotective effects were further verified by DCFH2-DA staining of cells and analysis by fluorescence microscopy (EVOS FL Auto 2 Imaging, ThermoFisher Scientific, CA, USA) and flow cytometry (CytoFLEX, Beckman Coulter, PA, USA) based on methods described in our previous publications [
8].
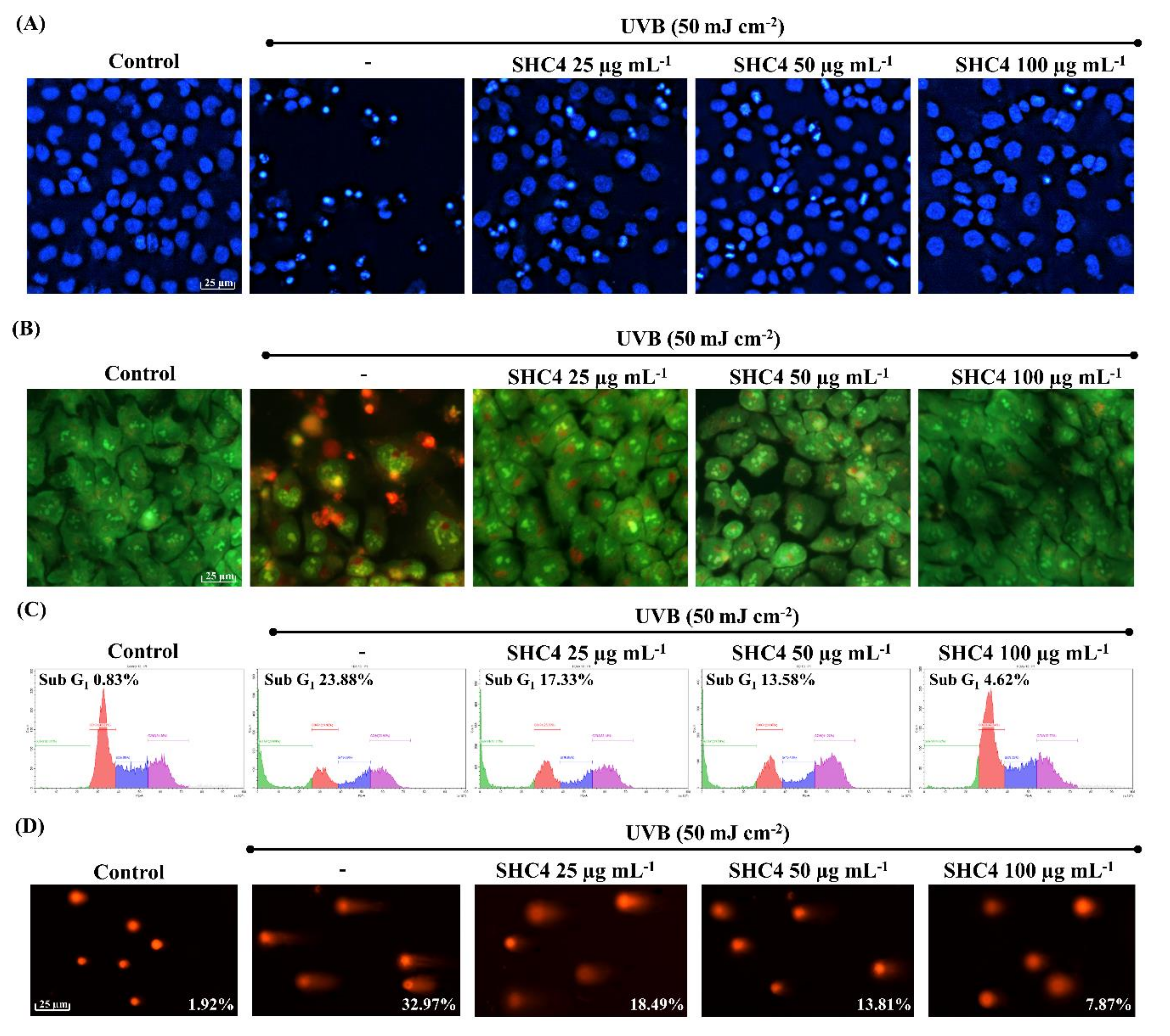
2.5.3. Analysis of Nuclear Morphology
Cells were stained with either Hoechst 33342 or mixture of acridine orange and ethidium bromide according to the method described in our previous publication [
9]. Nuclear morphologies were visualized on an EVOS FL Auto 2 Imaging microscope (Thermo-Fisher, Waltham, MA, USA).
2.5.4. Western Blot Analysis
HaCaT cells after initial sample treatment and UVB-exposure were used for the experiments following a 24 h incubation period. Western blot analysis was done following our previously reported method [
10]. A nuclear and cytoplasmic extraction kit, NE-PER
® (Thermo Scientific, Rockford, IL, USA), was used to isolate cytosolic and nucleic proteins. Proteins were standardized using a BCA
TM protein assay kit (Pierce, Rockford, IL, USA). Electrophoresis was carried out using 10% SDS-polyacrylamide gels. SuperSignal™ West Femto Maximum Sensitivity Substrate (Thermo, Burlington, ON, Canada) was used for the detection of protein bands on a Core Bio DavinchChemi imaging system (Seoul, Korea).
2.5.5. Cell Cycle Analysis
Harvested cells were permeabilized in 70% ethanol for 30 min, incubated in PBS containing EDTA, RNase and propidium iodide at 37 °C for 30 min, and analyzed by a Beckman Coulter CytoFLEX flow cytometer (Brea, CA, USA). A dual-region gating approach, initial FS vs. SS to remove derbies and FL3-area vs. FL3-hight to eliminate doublets, was applied. Sub-G1 apoptotic cell population percentages were used for data compression.
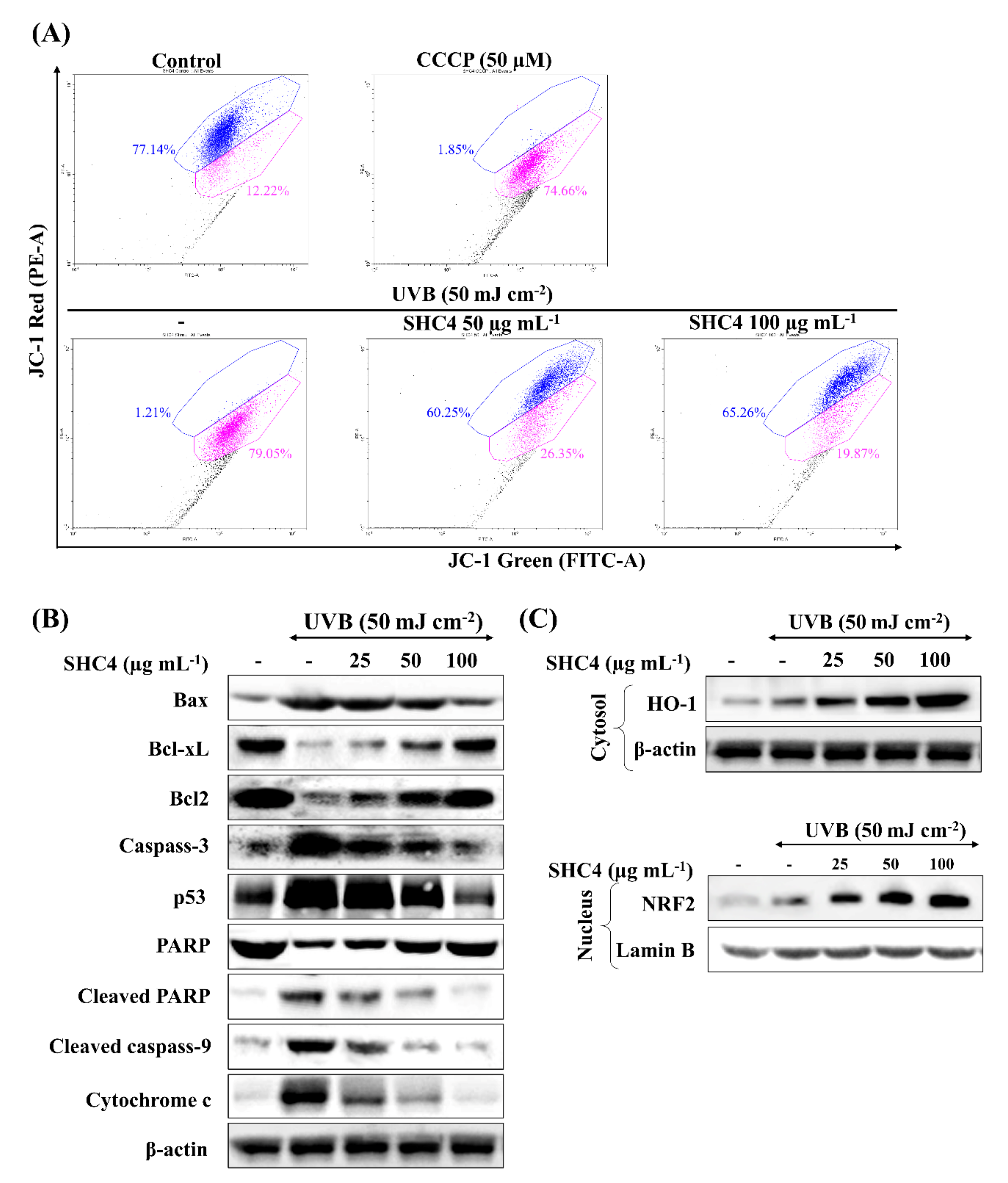
2.5.6. JC-1 Assay
Mitochondria membrane potential was ratiometrically measured by flow cytometry using MitoProbe JC-1 Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) following the manufacturer’s instructions. Cultured cells were initially subjected to sample treatment and UVB-exposure. After 4 h, cells were harvested for the assay.
2.5.7. Comet Assay
DNA damage in individual cells was evaluated by alkaline comet assay following the procedure outlined in our previous publication [
9]. Briefly, cells were harvested and suspended in low-melting agarose at 37 °C and gently applied on the surface of agarose coated slides. The slides were overnight immersed in alkaline lysis buffer at 4 °C. Electrophoresis was performed at 4 °C in an alkaline running buffer. Finally, the slides were submerged in a chilled neutralization buffer and stained with ethidium bromide. Comet tails were pictured on an EVOS FL Auto 2 Imaging microscope and tail DNA contents were quantified using OpenComet plugin in NIH Image J software (US National Institutes of Health, Bethesda, MD, USA).
2.6. Statistical Analysis
The data from experiments are presented as mean ± standard error (SE). Statistical comparisons were carried out by one-way analysis of variance by Duncan’s test using PASW Statistics 19.0 software (SPSS, Chicago, IL, USA). The level of significance was set at * p < 0.05 and ** p < 0.01.
4. Discussion
The edible brown seaweed
Sargassum horneri has long been acknowledged for its biofunctional effects. It is abundant along the Jeju island coasts in South Korea. Currently, the native
S. horneri population is threatened by the massive invasion of
S. horneri from the east coast of China [
22]. This not only threatens the coastal ecosystem but also reduces the attraction for tourists. Climate changes have contributed to warmer temperatures and increased rainfall, which increase fertilizer runoff from paddy fields and the aquaculture industry, adding nutrients to coastal areas. The drastic change observed in
S. horneri growth pattern is an example of how human activities alter the environmental balance. Other than clearing up the coasts, the utilization of
S. horneri in the food industry and for obtaining functional ingredients such as polyphenols, fucoidan and alginic acid could be an effective and beneficial strategy.
Celluclast is a food-grade cellulase that hydrolyzes cell wall cellulose, resulting in a higher extraction yield of plant materials. The water-based extract under pH 4.5 may contain ions and polar metabolites such as phenols, soluble polysaccharides and proteins. Alternatively, extraction at acidic pH greatly reduces alginate contamination. Based on preliminary investigations, the addition of CaCl
2 was not required as the formation of calcium alginate was negligible. Pretreatment with a mixture of chloroform and methanol (1:1) is reported to reduce lipophilic contaminants [
23]. The chloroform and methanol (1:1) extract is currently studied for its potential bioactive natural products. Pretreatment with 10% formaldehyde in ethanol is reported to reduce the polyphenolic contamination of polysaccharides [
23]. Otherwise, the contamination could lead to unreliable observations when assessing the bioactivities of polysaccharides. Further, washing with 80% ethanol not only removes any remaining formaldehyde but also removes polar compounds, such as phenolic acids. The reduced levels of polyphenol, protein and ash content (a measure of mineral content) in polysaccharide fractions obtained by step gradient ethanol precipitation suggested the efficacy of the reported method. The current pretreatment and extraction strategy was developed based on observations by several previous studies and preliminary analysis [
3,
11,
14,
24].
Crude fucoidan is generally obtained by adding a large volume of ethanol to the water-based extract. Subsequent fractionation is done by filtering precipitated polysaccharides through a series of molecular weight cutoff membranes (ultrafiltration) or by chromatography using size exclusion or anion exchange columns [
23]. Ultrafiltration and chromatography are not suitable for industrial-scale preparations considering the investment, cost of equipment and operation delays. The step gradient ethanol precipitation method offers convenience over conventional fucoidan purification techniques. However, the technique has not widely been used for fractionating fucoidans except for that in one reported study [
7]. The gradient alcohol precipitation method is commonly used in dextran fractionation and less frequently for purifying other polysaccharides such as starch, hemicellulose, glucan, fructan, pectin, arabinan and pullulan [
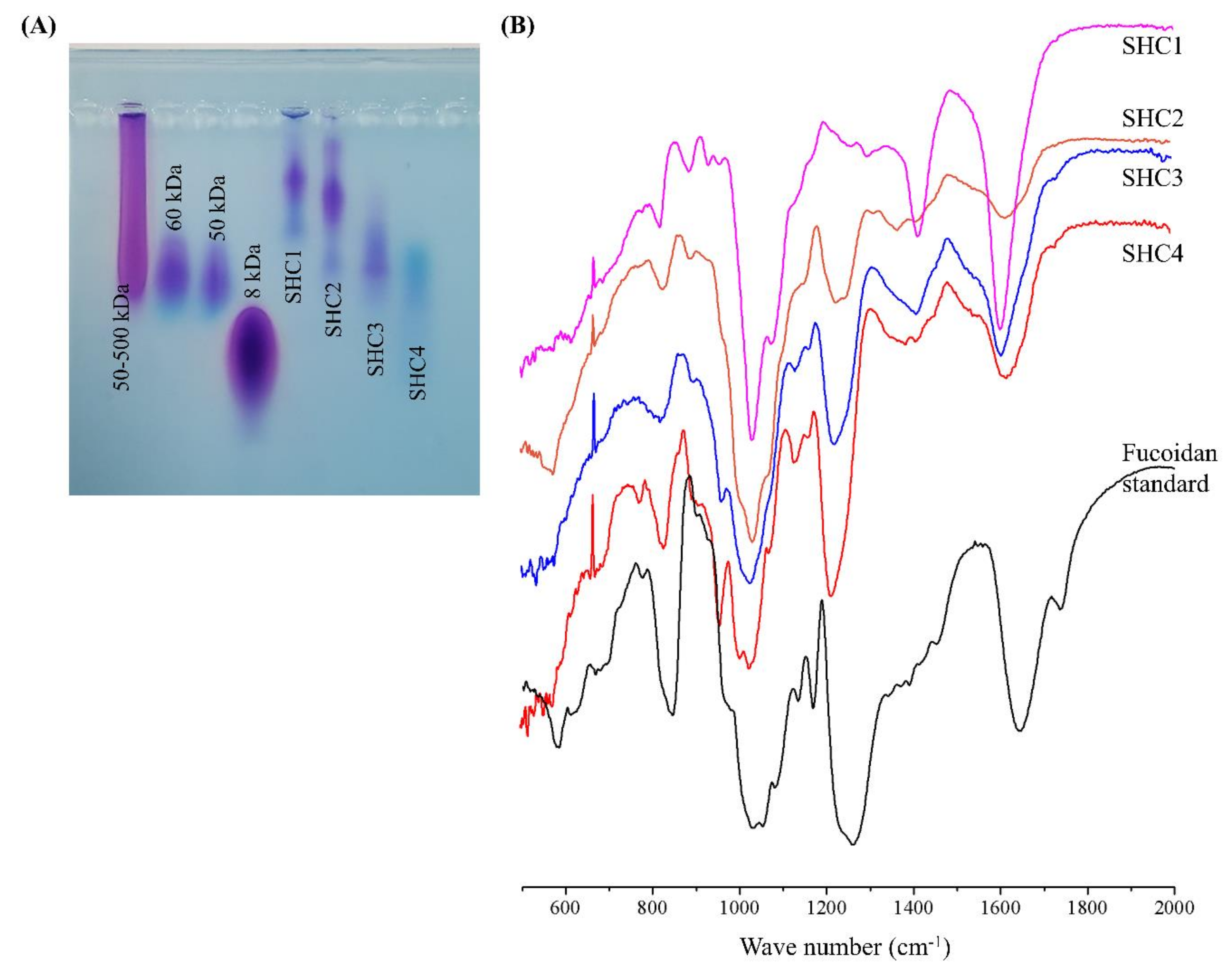
6]. Based on agarose gel electrophoresis, four different fractions with approximate molecular weight distributions of 230, 205, 90 and 60 kDa were successfully obtained using step gradient ethanol precipitation. The high yield of the SHC2 fraction suggested that most fucoidans in
S. horneri had a molecular weight range of approximately 205 kDa. Furthermore, low molecular weight polysaccharide fractions indicated a higher degree of sulfation. This further relates to the solubility of polysaccharides, where the ionic characteristics of polysaccharides increase with an increased degree of sulfation. Hence, the step gradient ethanol precipitation method could be said to separate fucoidans based on both the molecular weight and solubility.
The structural features of the fucoidan heteropolysaccharide vary considerably depending on seaweed species and environmental conditions. These features include monosaccharide composition, connectivity, chain length, degree of sulfation and connectivity of sulfate groups. Due to its heterogeneous nature, a full-scale analysis of connectivity remains challenging. Based on the present observations, the fractions were identified as fucoidans by FTIR and chemical composition analysis. Though it is not a generally accepted explanation, we propose that fucoidans could be ranked based on their quality, with the major parameters including the amount of fucose and degree of sulfation. According to many studies, the fucose amount is directly proportional to the degree of sulfation, which suggests that most sulfate groups remain attached to fucose units in fucoidan [
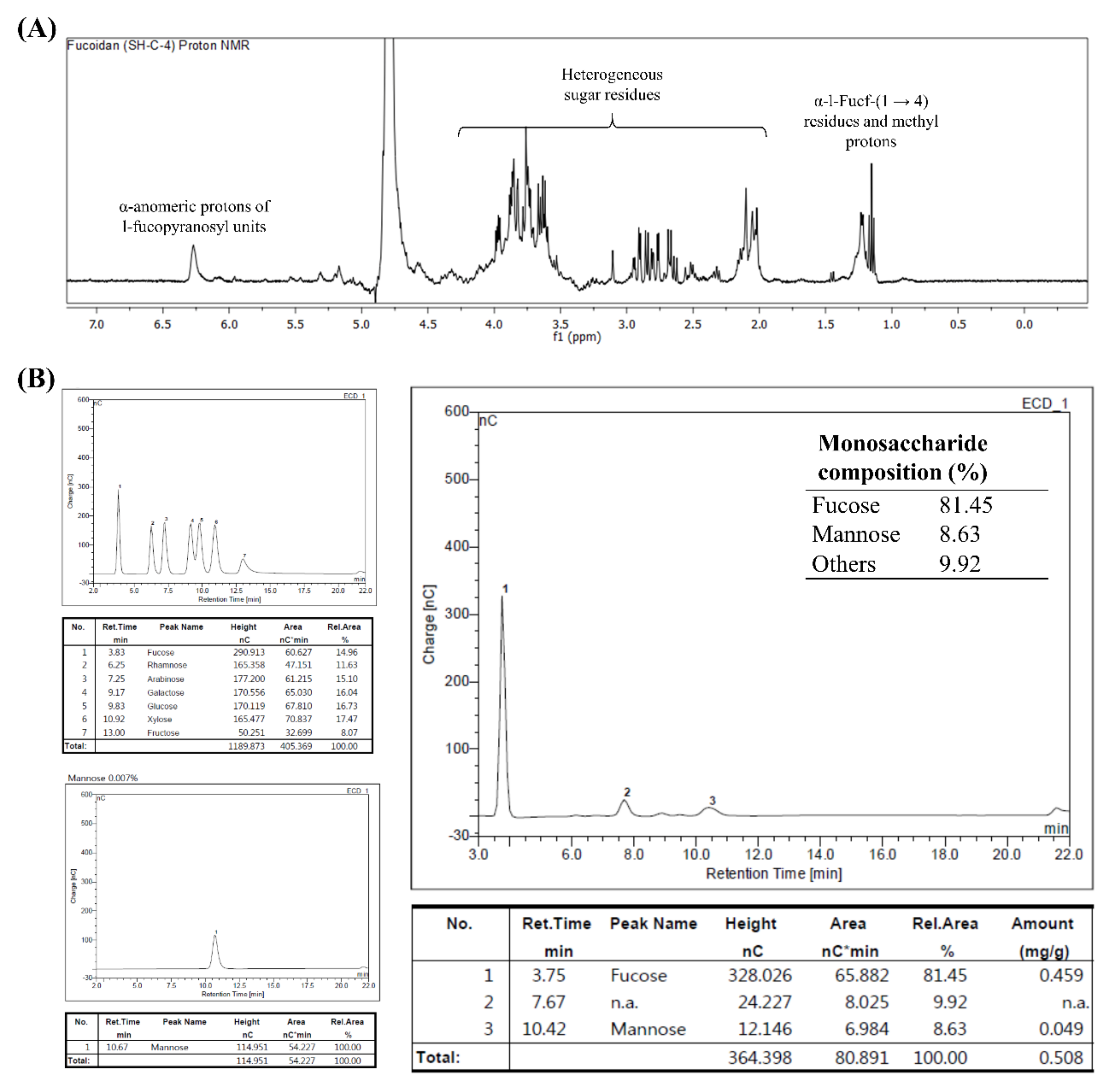
3]. The fucoidan fraction that had the most prominent antioxidant effects, SHC4, was found to contain 45.96 ± 0.36% polysaccharides, of which, 81.45% was fucose, which corresponded to 37.43% of the fraction weight with a sulfate content of 28.01 ± 0.50%.
1H NMR data revealed the presence of α-
l-Fucp-(1→4) residues, which are a major feature of fucoidans. Hence, we designated SHC4 as a high-quality fucoidan. The lack of data on monosaccharide and sulfate group connectivity patterns is a limitation of the present study. The connectivity “methylation analysis” requires the fucoidan to be further fractionated, obtaining a less heterogeneous polymer mix.
Fucoidan is renowned for its potential antioxidant properties among a wide range of bioactivities [
23]. The dose-dependent reduction of intracellular ROS levels in all fractions indicated the protective effect of fucoidans. Considering the slope observed for each fraction dose, the rate of the intracellular ROS reduction increased in succession, with the SHC4 fraction showing the most reduction. This suggested that low molecular weight and high-quality fucoidans had superior antioxidant properties. The increments observed for cell viability suggested that the protective effects of SHC4 were within the 25–100 µg mL
−1 concentration range. The minor reduction in cell viability at 200 µg mL
−1 concentration could be associated with toxic effects of sulfated polysaccharides under high concentrations. Further bioassays were conducted to assess the protective effects of SHC4 against UVB-induced oxidative stress and apoptosis. There was a reduction in UVB-induced apoptotic body formation, accumulation of Sub-G
1 apoptotic cells and DNA damage, clarifying the potential antioxidant activity of SHC4. Further studies were then performed to understand the effects of SHC4 on the regulation of apoptosis.
The mitochondria-mediated apoptosis pathway is considered a major signaling route mediating apoptosis; the underlying mechanism can vary based on the type of cell, stimulus and other factors. Oxidative stress resulting from multiple factors, including the induction of UVB-radiation is reported to activate the mitochondria-mediated apoptosis pathway [
18]. The pathway is initiated with the permeabilization of the mitochondrial outer membrane. The permeabilization is controlled by Bcl-2 family proteins, which includes anti-apoptotic proteins (Bcl-2 and Bcl-xL) and pro-apoptotic proteins (Bax, Bak, Bok, Bad, Bid, Bik, Bim, Bmf, Puma and Noxa). Their primary function is disrupting the functions of the Bcl-2 family proteins, thereby promoting the release of apoptogenic proteins present in the intermembrane space. Released apoptogenic proteins include cytochrome c, apoptosis-inducing factor (AIF) and endonuclease G. Cytochrome c with pro-caspase 9 and apoptosis protease activating factor (APAF-1) form an ‘apoptosome’. The apoptosome promotes caspase 9 activation, in turn activating effector caspases, which collectively execute apoptosis. Both AIF and endonuclease G contribute to DNA fragmentation and chromosomal condensation, features that are a hallmark of apoptosis [
25]. Apart from the aforementioned mediators, permeabilization of the mitochondrial outer membrane causes the release of numerous proteins that antagonize the activation of caspases. Pro-apoptotic stimuli inducing p53 further aggravate mitochondrial pathway-mediated apoptosis. p53 plays a crucial role in ultraviolet-induced apoptosis in HaCaT keratinocytes [
26]. Under physiological conditions, p53 is maintained at a low concentration, inhibiting its transcriptional activity. Activation and post-translational stabilization of p53 drives the expression of pro-apoptotic factors, provoking death pathways. Caspases, consisting of initiator and effector caspases, are proteases that mediate cell death. Initiator caspases, such as caspase-9, activate death receptors, whereas effector caspases directly mediate the cleavage of numerous cytoplasmic and nuclear substrates. Cytochrome c activates caspase-9 and subsequently activates downstream effector caspases such as caspase-3, −6 and −7 [
9]. The cleavage of PARP, a nuclear enzyme that maintains DNA stability, repair and transcription, is another crucial feature of apoptosis. Effector caspases cause the cleavage of PARP, thereby inhibiting its catalytic activity [
9]. The present observations clarified the effect of UVB on the initiation of mitochondria-mediated apoptosis proteins and the dose-dependent attenuation effects of SHC4. There could be numerous alternative pathways, other than the prominent mitochondria-mediated apoptosis that regulate oxidative stress-induced cell death. Hence, further studies could broaden the understanding of the effects of fucoidan on UVB-induced apoptosis.
Based on previous studies, fucoidans are reported to upregulate gene expression of HO-1 and SOD-1 in HaCaT keratinocytes via upregulating the transcription factor Nrf2 [
21]. Similar results were observed during the present analysis, where SHC4 treatment caused a dose-dependent increase of HO-1 production in the cytosol and Nrf2 levels in the nucleus. This suggested that the protective effects of fucoidan against UVB-induced oxidative stress were mediated by Nrf2/HO-1 signaling.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}