Dietary Antioxidants in Coffee Leaves: Impact of Botanical Origin and Maturity on Chlorogenic Acids and Xanthones

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Taxon Sampling and Plant Material
2.3. Preparation of Plant Material
2.4. Coffee Leaf Extraction
2.5. Validation of the Ultra-High Performance Liquid Chromatography-DAD method
2.6. Description of Ultra-High Performance Liquid Chromatography-DAD
2.7. Description of Ultra-High Performance Liquid Chromatography Electrospray Ionization Mass Spectrometry
2.8. Statistical Analysis
3. Results and Discussion
3.1. Validation of the UHPLC-DAD Method
3.2. Quantification of Alkaloids, Chlorogenic acids, Mangiferin, and Isomangiferin in Coffee Leaves
3.2.1. Alkaloids
3.2.2. Chlorogenic Acids
3.2.3. Xanthones
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Han, X.Z.; Shen, T.; Lou, H.X. Dietary polyphenols and their biological significance. Int. J. Mol. Sci. 2007, 8, 950–988. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Perez-Jimenez, J.; Fezeu, L.; Touvier, M.; Arnault, N.; Manach, C.; Hercberg, S.; Galan, P.; Scalbert, A. Dietary intake of 337 polyphenols in French adults. Am. J. Clin. Nutr. 2011, 93, 1220–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, C.; Fukushima, Y.; Kishimoto, Y.; Suzuki-sugihara, N.; Saita, E. Estimated dietary polyphenol intake and major food and beverage sources among elderly Japanese. Nutrients 2015, 7, 10269–10281. [Google Scholar] [CrossRef] [Green Version]
- Ovaskainen, M.; Törrönen, R.; Koponen, J.M.; Sinkko, H.; Hellström, J.; Reinivuo, H.; Mattila, P. Dietary intake and major food sources of polyphenols in Finnish adults. J. Nutr. 2008, 138, 562–566. [Google Scholar] [CrossRef]
- International Coffee Organization. Total Production by All Exporting Countries; International Coffee Organization: London, UK, 2018. [Google Scholar]
- Ludwig, I.A.; Clifford, M.N.; Lean, M.E.J.; Ashihara, H.; Crozier, A. Coffee: Biochemistry and potential impact on health. Food Funtion 2014, 5, 1695–1717. [Google Scholar] [CrossRef]
- Campa, C.; Petitvallet, A. Beneficial compounds from coffee leaves. In Achieving Sustainable Cultivation of Coffee; Lashermes, P., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018. [Google Scholar]
- Ashihara, H.; Monteiro, A.; Gillies, F.M.; Crozier, A. Biosynthesis of caffeine in leaves of coffee. Plant Physiol. 1996, 111, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Campa, C.; Mondolot, L.; Rakotondravao, A.; Bidel, L.P.; Gargadennec, A.; Couturon, E.; La Fisca, P.; Rakotomalala, J.; Jay-allemand, C.; Davis, A.P. A survey of mangiferin and hydroxycinnamic acid ester accumulation in coffee (Coffea) leaves: Biological implications and uses. Ann. Bot. 2012, 110, 595–613. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, M.T.S.; De Almeida, R.F.; Soto, G.; De Melo, E.; Filho, V.; Ulrich, C.M.; Owen, R.W. Quantitation by HPLC-UV of mangiferin and isomangiferin in coffee (Coffea arabica) leaves from Brazil and Costa Rica after solvent extraction and infusion. Food Anal. Methods 2016, 9, 2649–2655. [Google Scholar] [CrossRef]
- de Almeida, R.F.; Trevisan, M.T.S.; Thomaziello, R.A.; Breuer, A.; Klika, K.D.; Ulrich, C.M.; Owen, R.W. Nutraceutical compounds: Echinoids, flavonoids, xanthones and caffeine identified and quantitated in the leaves of Coffea arabica trees from three regions of Brazil. Food Res. Int. 2019, 115, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Farah, A.; Donangelo, C.M. Phenolic compounds in coffee. Braz. J. Plant Physiol. 2006, 18, 23–36. [Google Scholar] [CrossRef]
- Stalmach, A.; Steiling, H.; Williamson, G.; Crozier, A. Bioavailability of chlorogenic acids following acute ingestion of coffee by humans with an ileostomy. Arch. Biochem. Biophys. 2010, 501, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Mondolot, L.; La Fisca, P.; Buatois, B.; Talansier, E.; De Kochko, A.; Campa, C. Evolution in caffeoylquinic acid content and histolocalization during Coffea canephora leaf development. Ann. Bot. 2006, 98, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Campa, C.; Urban, L.; Mondolot, L.; Fabre, D.; Roques, S.; Lizzi, Y.; Aarrouf, J.; Doulbeau, S.; Breitler, J.; Letrez, C.; et al. Juvenile coffee leaves acclimated to low light are unable to cope with a moderate light increase. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, M.; Sousa, M.; Nascimento, M. Xanthone Derivatives: New Insights in Biological Activities. Curr. Med. Chem. 2005, 12, 2517–2538. [Google Scholar] [CrossRef]
- Joubert, E.; Gelderblom, W.C.A.; Louw, A.; de Beer, D. South African herbal teas: Aspalathus linearis, Cyclopia spp. and Athrixia phylicoides—A review. J. Ethnopharmacol. 2008, 119, 376–412. [Google Scholar] [CrossRef]
- Barreto, J.C.; Trevisan, M.T.S.; Hull, W.E.; Erben, G.; De Brito, E.; Pfundstein, B.; Wurtele, G.; Spiegelhalder, B.; Owen, R.W. Characterization and quantitation of polyphenolic compounds in bark, kernel, leaves and peel of mango (Mangifera indica L.). J. Agric. Food Chem. 2008, 56, 5599–5610. [Google Scholar] [CrossRef]
- Nehlig, A. Interindividual Differences in Caffeine Metabolism and Factors Driving Caffeine Consumption. Pharmacol. Rev. 2018, 70, 384–411. [Google Scholar] [CrossRef] [Green Version]
- Allred, K.F.; Yackley, K.M.; Vanamala, J.; Allred, C.D. Trigonelline Is a Novel Phytoestrogen in Coffee Beans. J. Nutr. Biochem. Mol. Genet. Mech. 2009, 139, 1833–1838. [Google Scholar] [CrossRef] [Green Version]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Antioxidant and prooxidant properties of caffeine, theobromine and xanthine. Med. Sci. Monit. 2003, 9, BR325–BR330. [Google Scholar] [PubMed]
- Tellone, E.; Galtieri, A.; Giardina, B.; Russo, A.; Bellocco, E.; Barreca, D.; Ficarra, S. Antioxidant Activity of Caffeine: A Focus on Human Red Blood Cells and Correlations with Several Neurodegenerative Disorders. In Coffee in Health and Disease Prevention; Elsevier Inc.: London, UK, 2015. [Google Scholar]
- Chen, X.; Mu, K.; Kitts, D.D. Characterization of phytochemical mixtures with inflammatory modulation potential from coffee leaves processed by green and black tea processing methods. Food Chem. 2019, 271, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Ma, Z.; Kitts, D.D. Effects of processing method and age of leaves on phytochemical profiles and bioactivity of coffee leaves. Food Chem. 2018, 249, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Kirkpatrick, J.; Kuhnert, N.; Roozendaal, H.; Salgado, P.R.; Kiikpatrick, J.; Kuhnert, N.; Roozendaal, H.; Salgado, P.R. LC-MSn analysis of the cis isomers of chlorogenic acids. Food Chem. 2008, 106, 379–385. [Google Scholar] [CrossRef]
- Gumustas, M.; Kurbanoglu, S.; Uslu, B.; Ozkan, S.A. UPLC versus HPLC on Drug Analysis: Advantageous, Applications and Their Validation Parameters. Chromatographia 2013, 76, 1365–1427. [Google Scholar] [CrossRef]
- Perrone, D.; Donangelo, C.M.; Farah, A. Fast simultaneous analysis of caffeine, trigonelline, nicotinic acid and sucrose in coffee by liquid chromatography–mass spectrometry. Food Chem. 2008, 110, 1030–1035. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, H.; Jeong, I.; Hong, S.; Oh, M.; Park, K.; Shim, J.; Abd El-Aty, A.M. Determination of chlorogenic acids and caffeine in homemade brewed coffee prepared under various conditions. J. Chromatogr. B 2017, 1064, 115–123. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.; Knight, S.; Kuhnert, N. Hierarchical Scheme for LC-MS Identification of Chlorogenic Acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Zheng, X.; Ashihara, H. Distribution, biosynthesis and function of purine and pyridine alkaloids in Coffea arabica seedlings. Plant Sci. 2004, 166, 807–813. [Google Scholar] [CrossRef]
- Ky, C.; Louarn, J.; Dussert, S.; Guyot, B.; Hamon, S.; Noirot, M. Caffeine, trigonelline, chlorogenic acids and sucrose diversity in wild Coffea arabica L. and C. canephora P. accessions. Food Chem. 2001, 75, 223–230. [Google Scholar] [CrossRef]
- Farah, A.; Monteiro, M.C.C.; Calado, V.; Franca, A.S.S.; Trugo, L.C.C. Correlation between cup quality and chemical attributes of Brazilian coffee. Food Chem. 2006, 98, 373–380. [Google Scholar] [CrossRef]
- Nagai, C.; Rakotomalala, J.J.; Katahira, R.; Li, Y.; Yamagata, K.; Ashihara, H. Production of a new low-caffeine hybrid coffee and the biochemical mechanism of low caffeine accumulation. Euphytica 2008, 164, 133–142. [Google Scholar] [CrossRef]
- Priolli, R.H.G.; Mazzafera, P.; Siqueira, W.J.; Möller, M.; Zucchi, M.I.; Ramos, L.C.S.; Gallo, P.B.; Colombo, C.A. Caffeine inheritance in interspecific hybrids of Coffea arabica × Coffea canephora (Gentianales, Rubiaceae). Genet. Mol. Biol. 2008, 31, 498–504. [Google Scholar] [CrossRef]
- Cheng, B.; Furtado, A.; Smyth, H.E.; Henry, R.J. Influence of genotype and environment on coffee quality. Trends Food Sci. Technol. 2016, 57, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Ashihara, H.; Sano, H.; Crozier, A. Caffeine and related purine alkaloids: Biosynthesis, catabolism, function and genetic engineering. Phytochemistry 2008, 69, 841–856. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, S.T.V.; Guedes, R.N.C.; Demuner, A.J.; Lima, E.R. Effect of coffee alkaloids and phenolics on egg-laying by the coffee leaf miner Leucoptera coffeella. Bull. Entomol. Res. 2008, 98, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Oestreich-Janzen, S. Chemistry of Coffee. In Comprehensive Natural Products II Chemistry and Biology; Elsevier: Kidlington, UK, 2010. [Google Scholar]
- Koshiro, Y.; Jackson, M.C.; Katahira, R.; Wang, M.L.; Nagai, C.; Ashihara, H. Biosynthesis of chlorogenic acids in growing and ripening fruits of Coffea arabica and Coffea canephora plants. Zeitschrift für Naturforschung C 2007, 62, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Campa, C.; Doulbeau, S.; Dussert, S.; Hamon, S.; Noirot, M. Qualitative relationship between caffeine and chlorogenic acid contents among wild Coffea species. Food Chem. 2005, 93, 135–139. [Google Scholar] [CrossRef]
- Shang, X.; Pan, H.; Li, M.; Miao, X.; Ding, H. Lonicera japonica Thunb.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2011, 138, 1–21. [Google Scholar] [CrossRef]
- The Commission of the PPRC. Pharmacopoeia of People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2015. [Google Scholar]
- Imran, M.; Arshad, M.S.; Butt, M.S.; Kwon, J.H.; Arshad, M.U.; Sultan, M.T. Mangiferin: A natural miracle bioactive compound against lifestyle related disorders. Lipids Health Dis. 2017, 16, 1–17. [Google Scholar] [CrossRef]


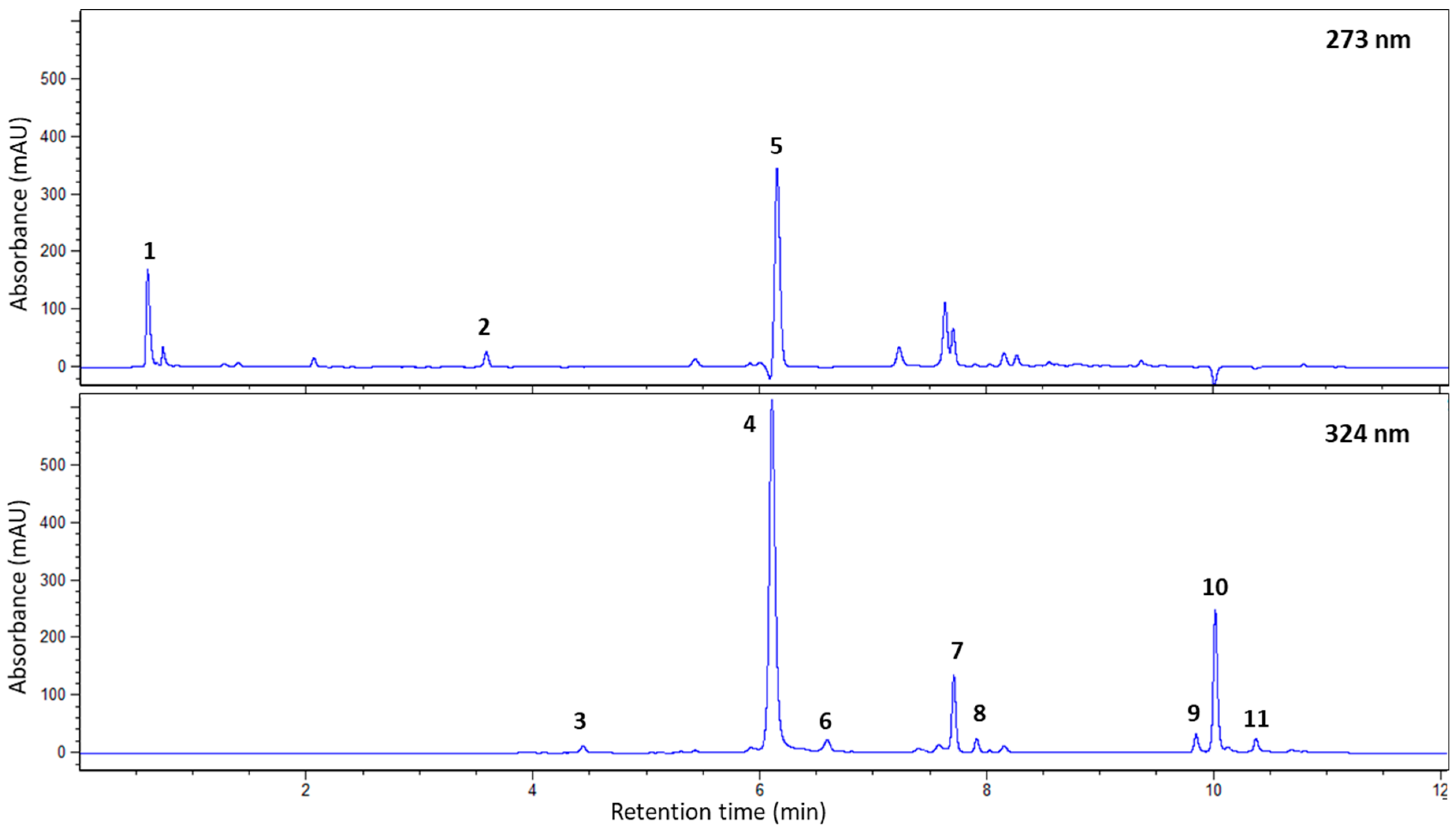{kind=link}
| Analyte | λmax (nm) | Retention Time (min) | Calibration Curve | LOD 1 (µg/mL) | LOQ 2 (µg/mL) | ||
|---|---|---|---|---|---|---|---|
| Intercept | Slope | r2 | |||||
| Trigonelline | 254 | 0.60 | 9.025 | 5.375 | 0.9996 | 0.110 | 0.365 |
| Mangiferin Isomangiferin | 258 | 7.71 7.85 | 4.986 | 16.409 | 0.9987 | 0.025 | 0.083 |
| Theobromine | 273 | 3.65 | 1.482 | 17.420 | 0.9999 | 0.023 | 0.075 |
| Caffeine | 6.24 | 32.541 | 14.371 | 0.9999 | 0.003 | 0.012 | |
| 3-CQA | 324 | 4.49 | 8.217 | 8.924 | 0.9999 | 0.066 | 0.220 |
| 5-CQA | 6.18 | 23.912 | 8.487 | 0.9998 | 0.089 | 0.297 | |
| 4-CQA | 6.66 | 0.710 | 9.330 | 0.9999 | 0.044 | 0.145 | |
| 3,4-DiCQA | 9.88 | 1.354 | 8.913 | 0.9995 | 0.016 | 0.054 | |
| 3,5-DiCQA | 10.05 | 0.771 | 11.102 | 0.9999 | 0.108 | 0.359 | |
| 4,5-DiCQA | 10.41 | −1.908 | 11.486 | 0.9993 | 0.057 | 0.190 | |
| Species | Coffee Genotype | Trigonelline | Theobromine | Caffeine * | Total Alkaloids | ||||
|---|---|---|---|---|---|---|---|---|---|
| Young | Mature | Young | Mature | Young | Mature | Young | Mature | ||
| C. arabica | var. Bourbon | 11.718 ± 0.544 | 2.955 ± 0.207 | 0.811 ± 0.029 | 0.045 ± 0.005 | 14.936 ± 0.019 a | 5.993 ± 0.508 e | 27.465 ± 0.592 | 8.993 ± 0.720 |
| C. canephora | CIFC 2975 | 7.879 ± 0.209 | 5.979 ± 0.017 | 0.169 ± 0.001 | 0.046 ± 0.000 | 13.186 ± 0.423 b | 7.527 ± 0.032 d | 21.234 ± 0.633 | 13.552 ± 0.049 |
| CIFC 829/1 | 4.817 ± 0.019 | 4.157 ± 0.300 | 4.142 ± 0.040 | 0.806 ± 0.076 | 10.043 ± 0.043 c | 2.805 ± 0.077 f | 19.002 ± 0.102 | 7.769 ± 0.454 | |
| C. eugenioides | CIFC 1634/11 | 14.596 ± 0.143 | 11.232 ± 0.001 | - | - | - | - | 14.596 ± 0.143 | 11.232 ± 0.001 |
| CIFC 241/43 | 9.932 ± 0.021 | 7.781 ± 0.002 | - | - | - | - | 9.932 ± 0.021 | 7.781 ± 0.002 | |
| C. racemosa | CIFC 1693/76 | 11.188 ± 0.222 | 9.836 ± 0.177 | 1.532 ± 0.026 | 0.549 ± 0.003 | - | - | 12.721 ± 0.245 | 10.385 ± 0.180 |
| CIFC 13969 | 11.090 ± 0.245 | 9.736 ± 0.092 | - | - | - | - | 11.090 ± 0.245 | 9.736 ± 0.092 | |
| Species | Coffee Genotypes | 3-CQA | 5-CQA | 4-CQA | 3,4-DiCQA | ||||
|---|---|---|---|---|---|---|---|---|---|
| Young | Mature | Young | Mature | Young | Mature | Young | Mature | ||
| C. arabica | var. Bourbon | 0.881 ± 0.070 | 1.429 ± 0.103 | 56.790 ± 3.640 | 14.678 ± 1.617 | 1.586 ± 0.110 | 1.853 ± 0.155 | 1.798 ± 0.133 | 0.431 ± 0.040 |
| C. canephora | CIFC 2975 | 0.906 ± 0.031 | 0.873 ± 0.002 | 22.880 ± 0.796 | 13.874 ± 0.033 | 0.906 ± 0.006 | 1.035 ± 0.008 | 1.003 ± 0.007 | 0.441 ± 0.000 |
| CIFC 829/1 | 1.395 ± 0.002 | 1.217 ± 0.031 | 29.458 ± 0.155 | 16.346 ± 0.798 | 3.850 ± 0.003 | 2.602 ± 0.175 | 1.001 ±0.007 | 0.498 ± 0.018 | |
| C. eugenioides | CIFC 1634/11 | 0.521 ± 0.003 | 0.402 ± 0.004 | 49.581 ± 0.299 | 31.451 ± 0.538 | 3.148 ± 0.000 | 2.872 ± 0.040 | 0.549 ± 0.001 | 0.257 ± 0.005 |
| CIFC 241/43 | 0.716 ± 0.006 | 0.450 ± 0.005 | 34.283 ± 0.657 | 17.667 ± 0.200 | 5.096 ± 0.039 | 3.388 ± 0.040 | 0.421 ± 0.012 | 0.044 ± 0.000 | |
| C. racemosa | CIFC 1693/76 | 3.007 ± 0.030 | 2.816 ± 0.018 | 21.123 ± 0.129 | 15.574 ± 0.002 | 8.359 ± 0.112 | 6.546 ± 0.099 | 0.797 ± 0.009 | 0.628 ± 0.032 |
| CIFC 13969 | 1.618 ± 0.016 | 2.234 ± 0.006 | 46.145 ± 0.752 | 24.781 ± 0.129 | 8.232 ± 0.050 | 6.282 ± 0.032 | 2.044 ± 0.101 | 1.768 ± 0.043 | |
| Species | Coffee Genotypes | 3,5-DiCQA | 4,5-DiCQA | Total CGAs * | |||||
| Young | Mature | Young | Mature | Young | Mature | ||||
| C. arabica | var. Bourbon | 11.266 ± 0.591 | 0.318 ± 0.030 | 1.217 ± 0.045 | 0.499 ± 0.067 | 73.539 ± 4.590 b | 19.208 ± 2.012 i | ||
| C. canephora | CIFC 1693/76 | 9.952 ± 0.184 | 2.994 ± 0.034 | 0.085 ± 0.001 | 0.095 ± 0.003 | 35.732 ± 1.027 g | 19.312 ± 0.080 i | ||
| CIFC 13969 | 11.035 ± 0.070 | 1.934 ± 0.019 | 0.185 ± 0.003 | 0.066 ± 0.013 | 46.924 ± 0.239 d | 22.663 ± 1.042 i | |||
| C. eugenioides | CIFC 2975 | 6.513 ± 0.136 | 3.495 ± 0.302 | 0.704 ± 0.118 | 0.344 ± 0.019 | 40.503 ± 0.533 ef | 29.402 ± 0.473 h | ||
| CIFC 829/1 | 20.309 ± 0.423 | 6.739 ± 0.001 | 2.488 ± 0.080 | 2.308 ± 0.105 | 80.836 ± 1.422 a | 44.112 ± 0.180 de | |||
| C. racemosa | CIFC 1634/11 | 6.837 ± 0.065 | 1.348 ± 0.011 | 1.150 ± 0.013 | 0.272 ± 0.003 | 61.782 ± 0.382 c | 36.603 ± 0.600 fg | ||
| CIFC 241/43 | 2.388 ± 0.068 | 0.084 ± 0.000 | 0.914 ± 0.033 | 0.091 ± 0.003 | 43.818 ± 0.814 de | 21.724 ± 0.248 i | |||
| Species | Coffee Genotypes | Mangiferin * | Isomangiferin | Total Xanthones | |||
|---|---|---|---|---|---|---|---|
| Young | Mature | Young | Mature | Young | Mature | ||
| C. arabica | var. Bourbon | 14.714 ± 0.838 e | 2.222 ± 0.326 f | 1.076 ± 0.038 | 0.111 ± 0.002 | 15.789 ± 0.876 | 2.333 ± 0.328 |
| C. canephora | CIFC 2975 | - | - | - | - | - | - |
| CIFC 829/1 | - | - | - | - | - | - | |
| C. eugenioides | CIFC 1634/11 | 53.875 ± 0.168 b | 30.195 ± 0.714 d | 9.644 ± 0.026 | 5.392 ± 0.128 | 63.519 ± 0.194 | 35.587 ± 0.842 |
| CIFC 241/43 | 76.686 ± 1.176 a | 32.605 ± 0.081 c | 12.959 ± 0.335 | 5.411 ± 0.147 | 89.645 ± 1.511 | 38.016 ± 0.228 | |
| C. racemosa | CIFC 1693/76 | - | - | - | - | - | - |
| CIFC 13969 | - | - | - | - | - | - | |
| Species | Coffee Genotypes | Decrease during Leaf Aging (%) | ||
|---|---|---|---|---|
| 5-CQA | 3,5-DiCQA | Xanthones | ||
| C. arabica | var. Bourbon | 74 | 97 | 85 |
| C. eugenioides | CIFC 241/43 | 37 | 46 | 44 |
| CIFC 1634/11 | 48 | 67 | 58 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, Â.; Colomban, S.; Azinheira, H.G.; Guerra-Guimarães, L.; Do Céu Silva, M.; Navarini, L.; Resmini, M. Dietary Antioxidants in Coffee Leaves: Impact of Botanical Origin and Maturity on Chlorogenic Acids and Xanthones. Antioxidants 2020, 9, 6. https://doi.org/10.3390/antiox9010006
Monteiro Â, Colomban S, Azinheira HG, Guerra-Guimarães L, Do Céu Silva M, Navarini L, Resmini M. Dietary Antioxidants in Coffee Leaves: Impact of Botanical Origin and Maturity on Chlorogenic Acids and Xanthones. Antioxidants. 2020; 9(1):6. https://doi.org/10.3390/antiox9010006
Chicago/Turabian StyleMonteiro, Ângelo, Silvia Colomban, Helena G. Azinheira, Leonor Guerra-Guimarães, Maria Do Céu Silva, Luciano Navarini, and Marina Resmini. 2020. "Dietary Antioxidants in Coffee Leaves: Impact of Botanical Origin and Maturity on Chlorogenic Acids and Xanthones" Antioxidants 9, no. 1: 6. https://doi.org/10.3390/antiox9010006