

Redox Biomarkers and Matrix Remodeling Molecules in Ovarian Cancer
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Enrolled in the Study
2.2. Antioxidant Enzymes’ Activities
2.3. Reduced Glutathione (GSH) Content
2.4. Pro-Oxidant Enzymes
2.5. Nitrosative Stress Parameters
2.6. Protein Glycoxidation Products
2.7. Oxidative Damage of Lipids
2.8. Caspase Activity
2.9. Real-Time PCR Reaction
2.10. Metalloproteinases Activity
2.11. TCGA Data Analysis
2.12. Statistical Analysis
3. Results
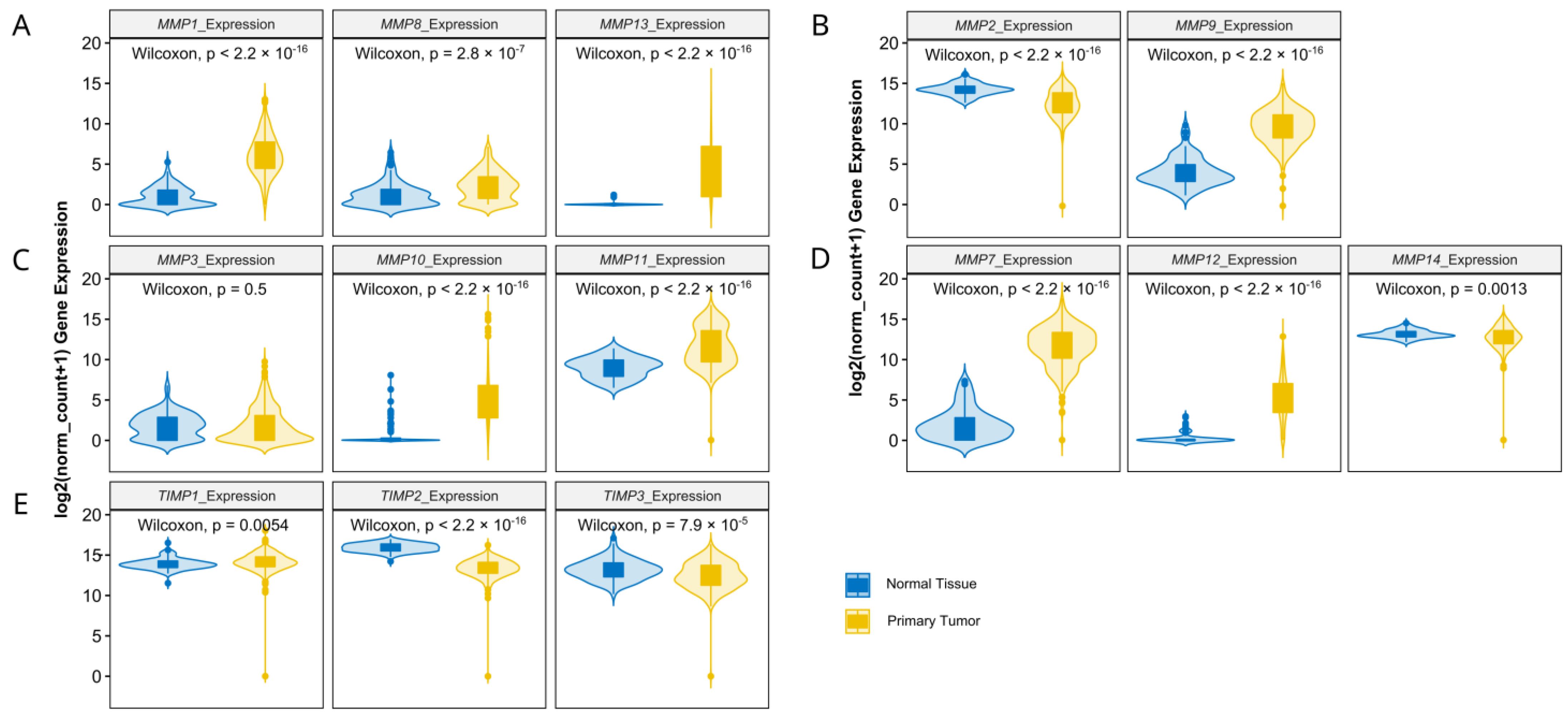
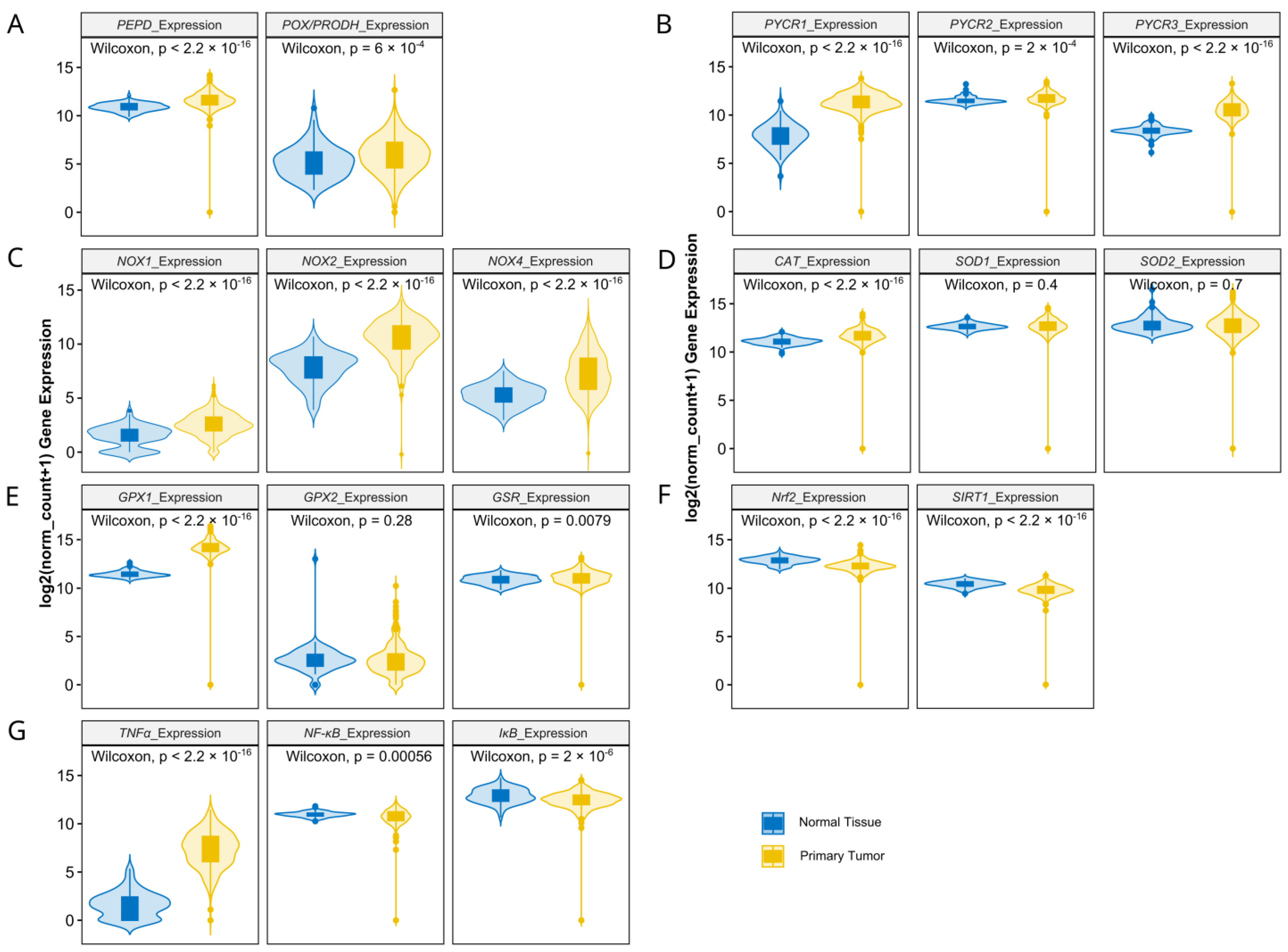
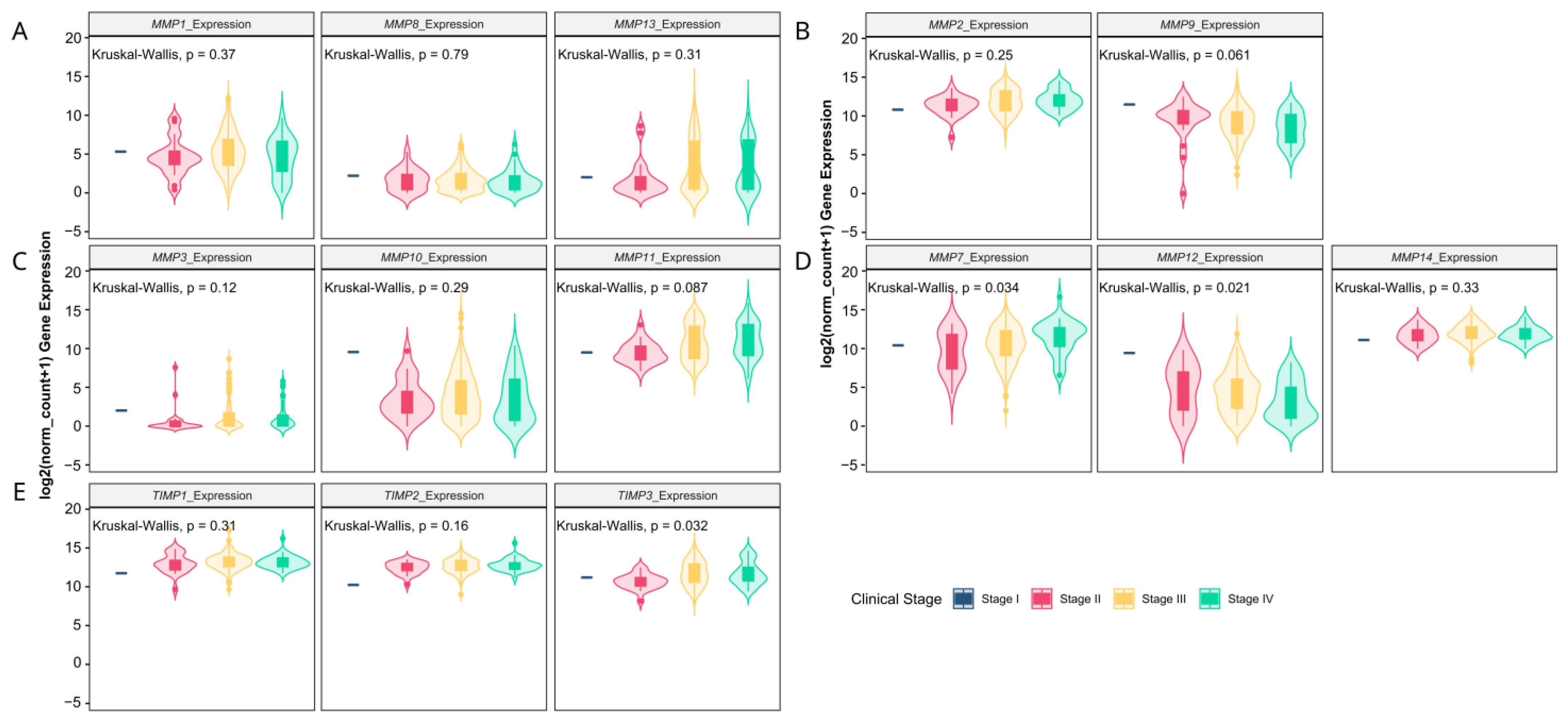
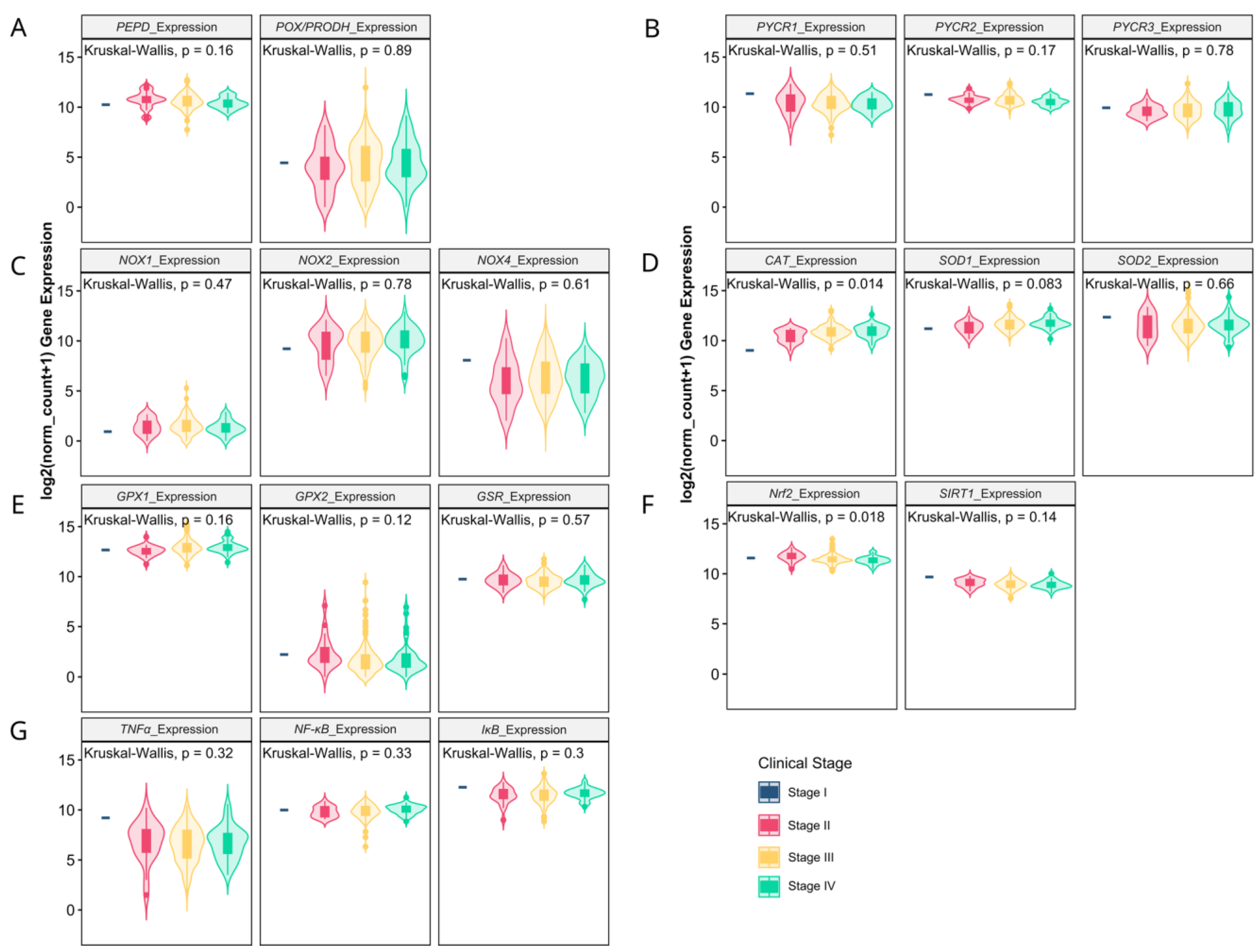
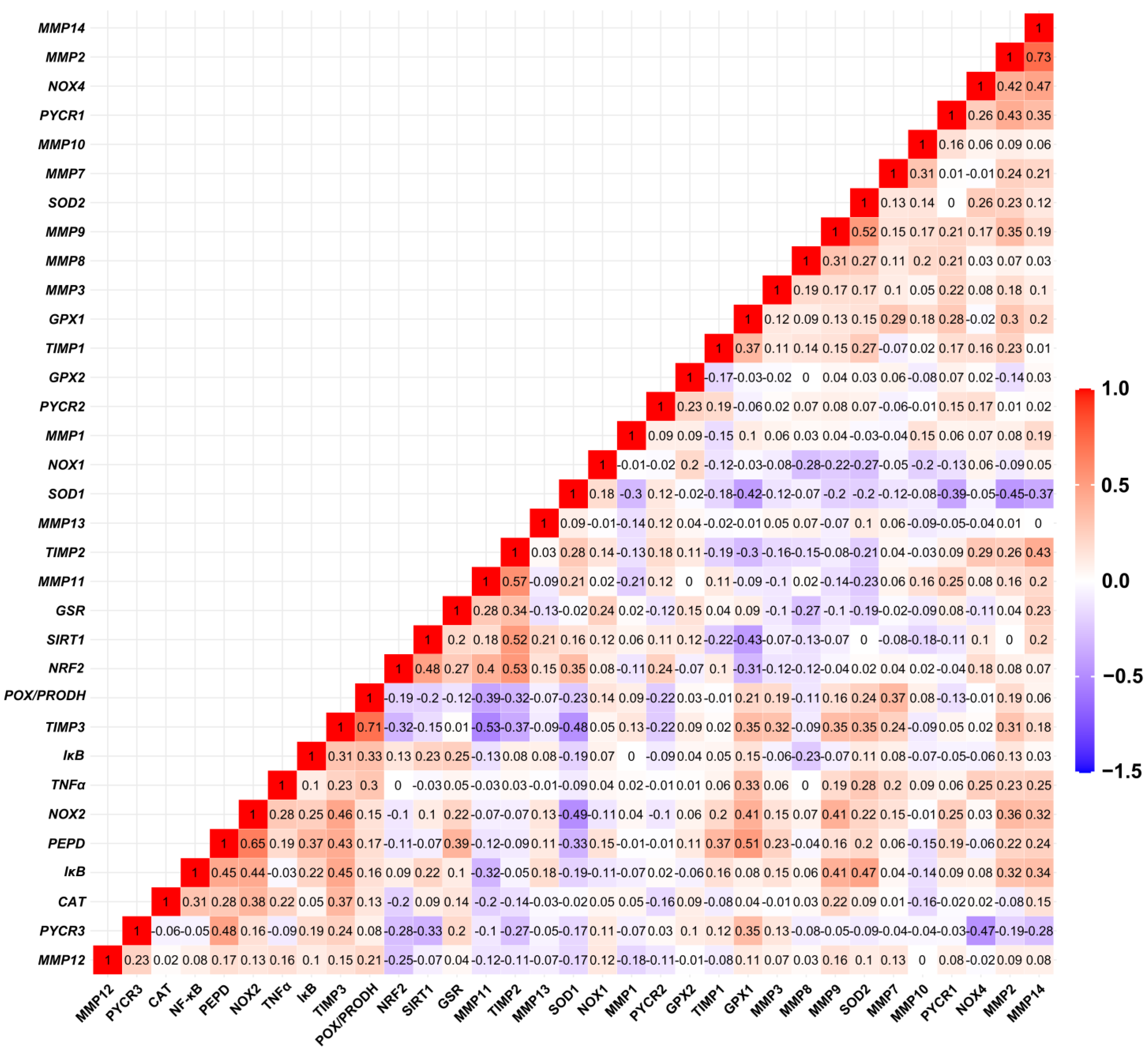
3.1. Matrix-Remodeling Associated and Redox-Related Gene Expression in TCGA Cohort
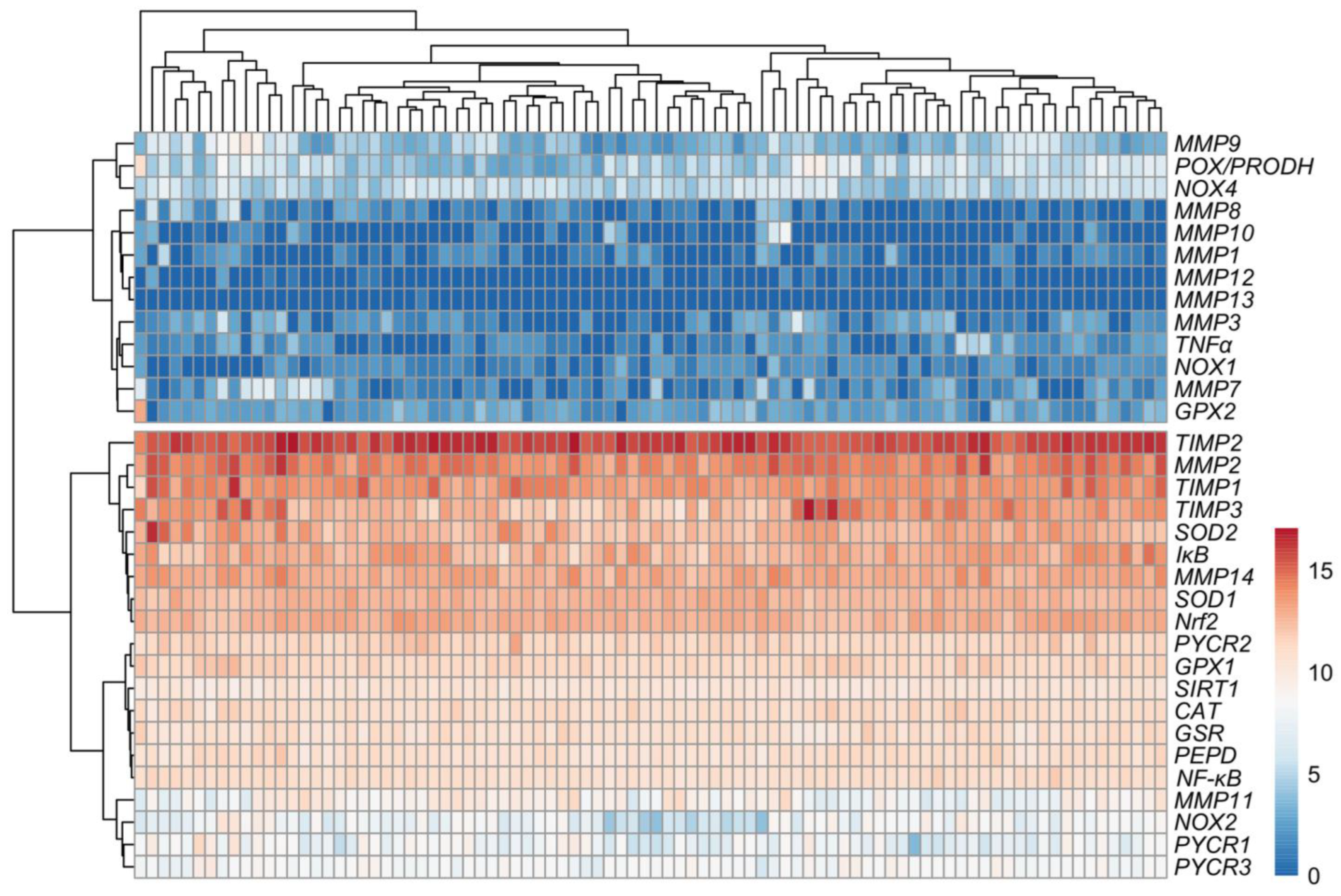
3.2. Relationship between Expression Levels of Matrix-Remodeling Associated and Redox-Related Gene Expression in TCGA Cohort
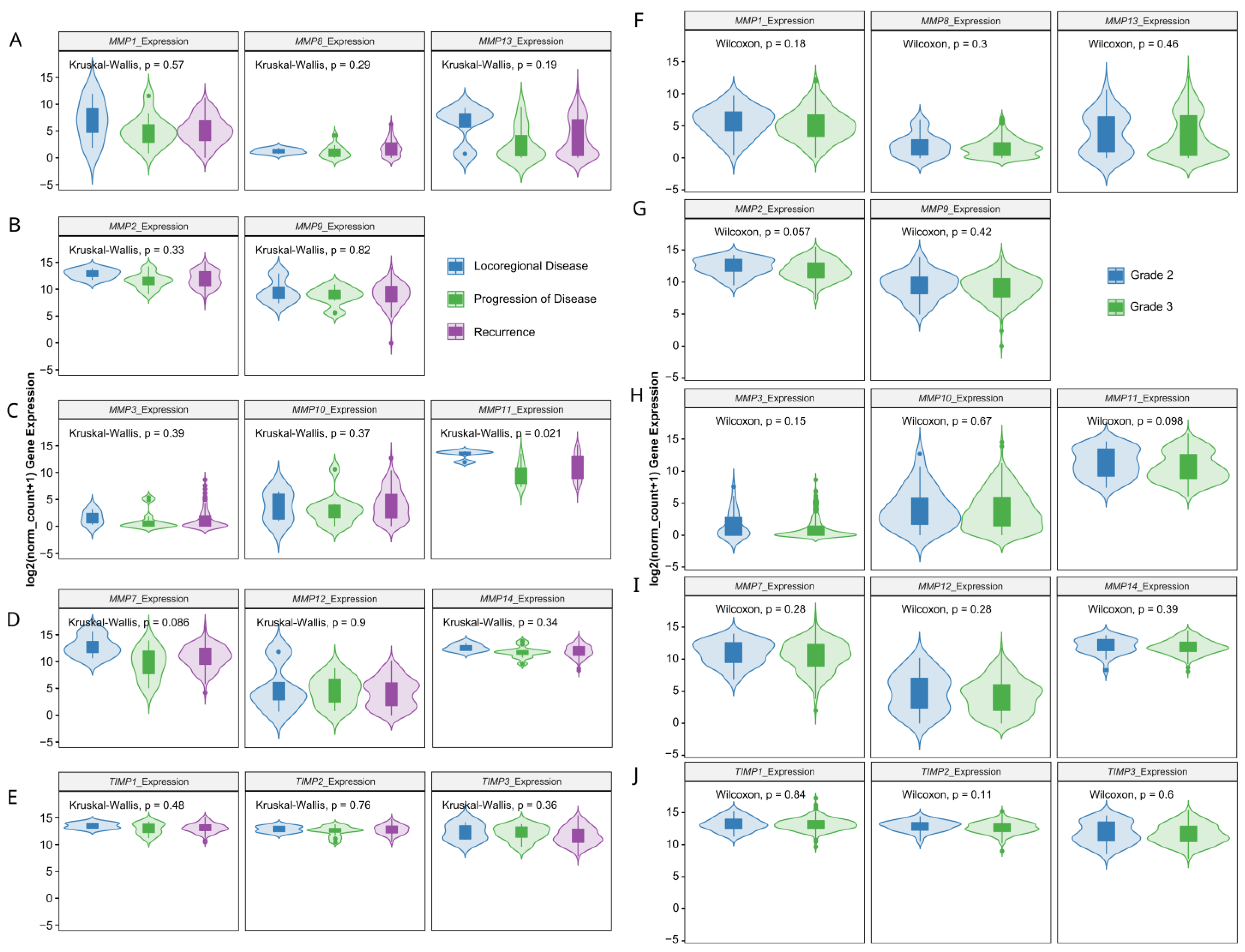
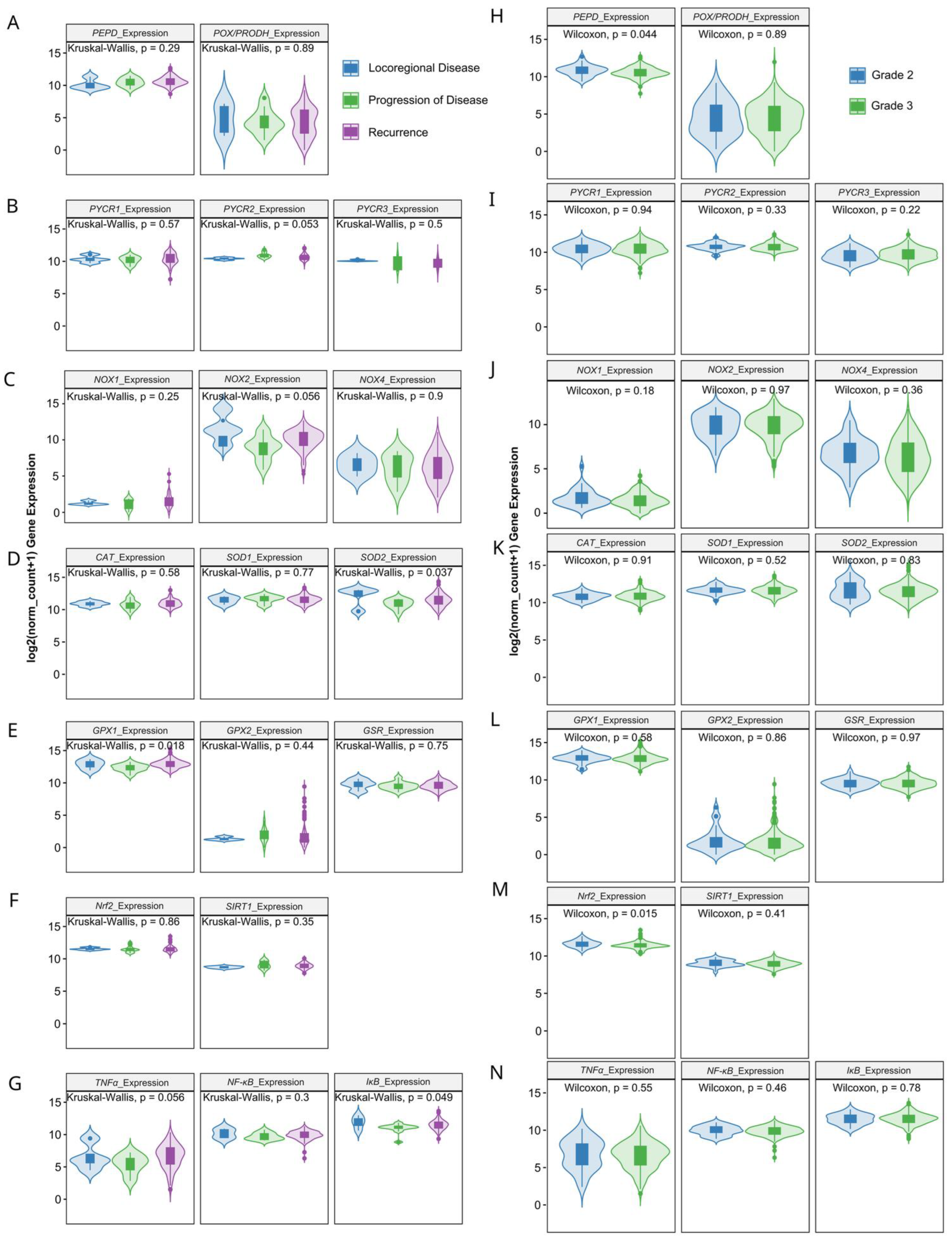
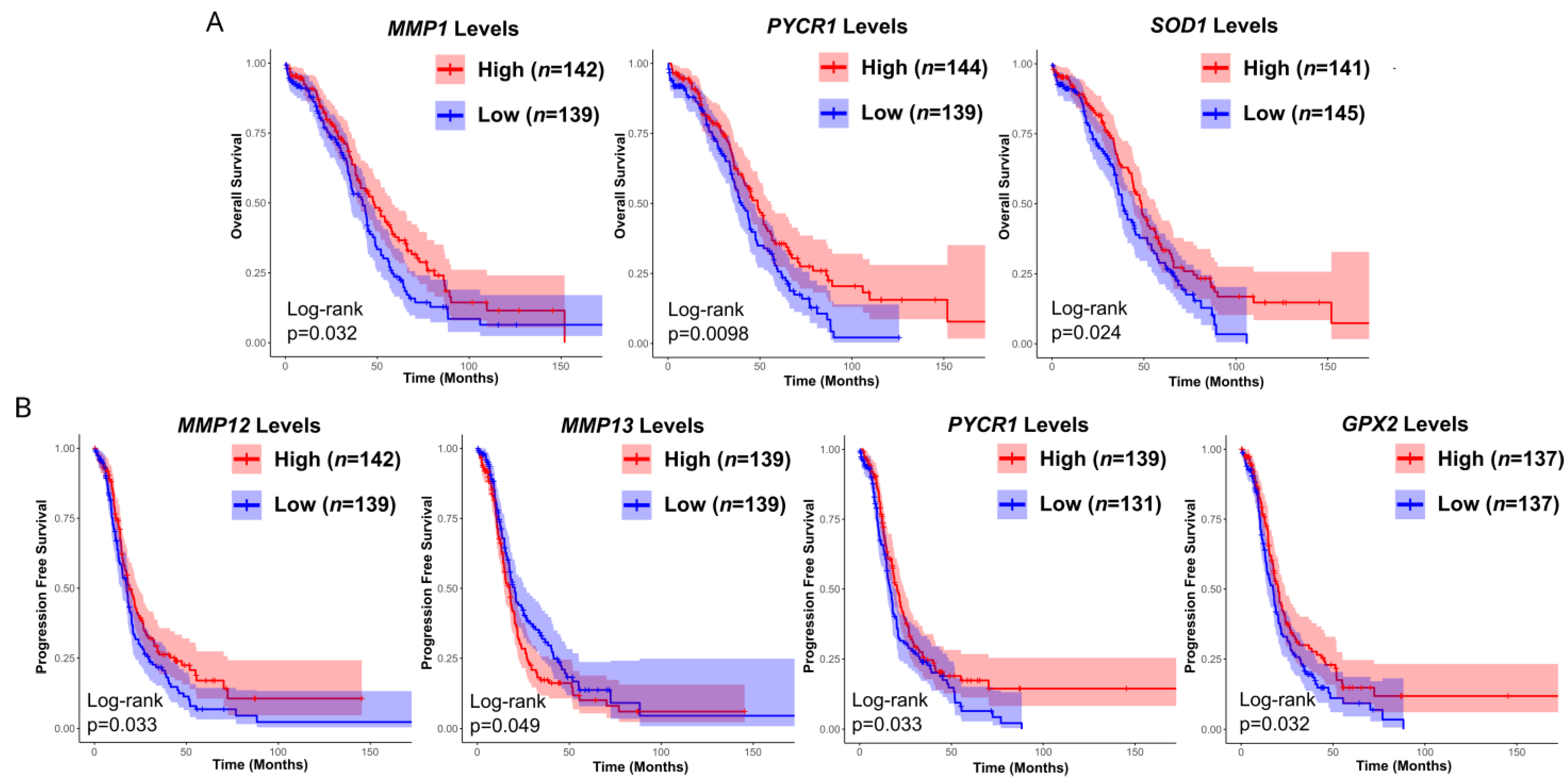
3.3. Prognostic Value of Matrix-Remodeling Associated and Redox-Related Gene Expression in Ovarian Cancer
3.4. Patient Characteristics
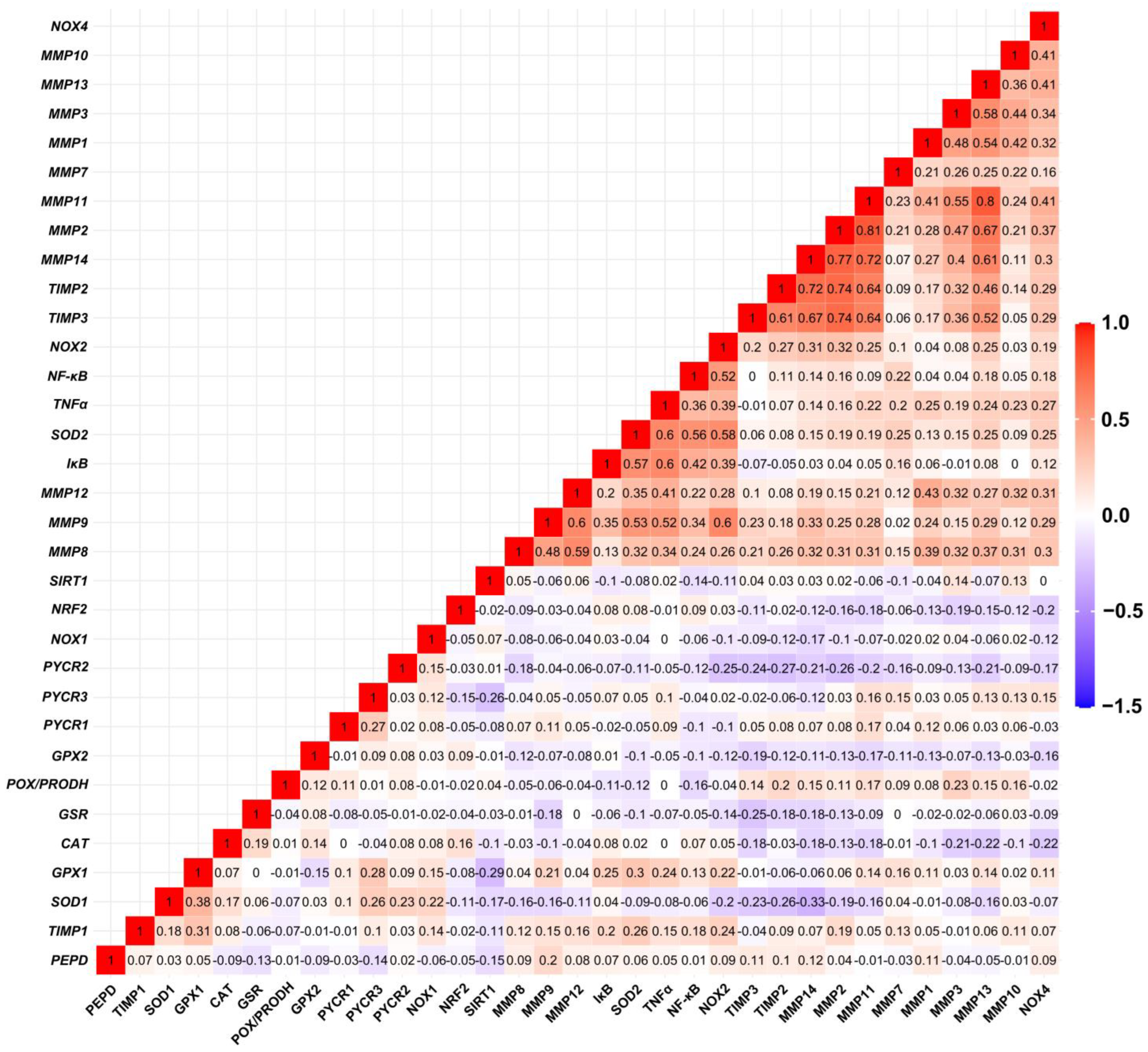
3.5. Matrix Remodeling-Associated and Redox-Related Gene Expression in HGSOC Patients
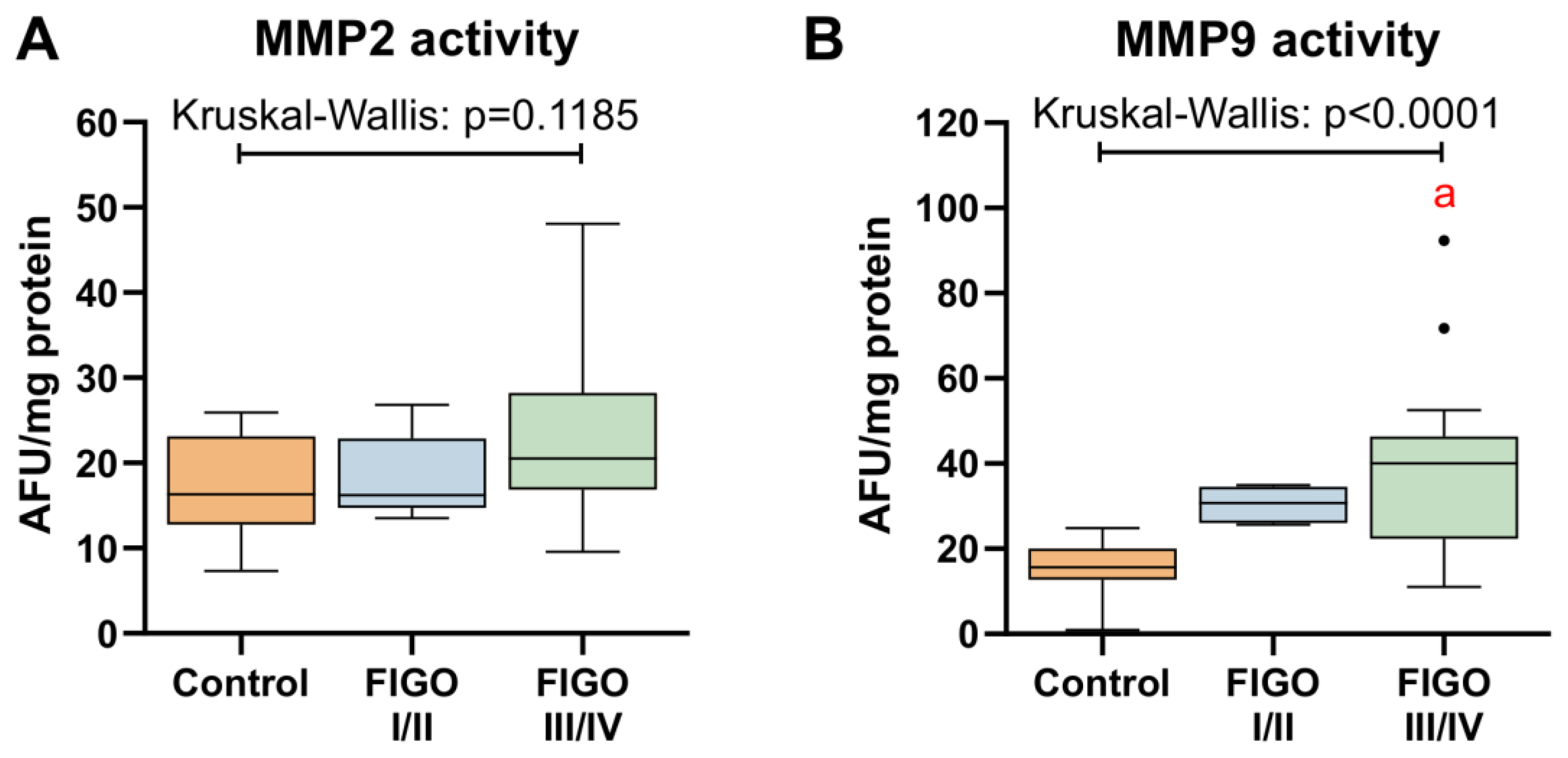
3.6. MMPs Activity in Ovarian Cancer
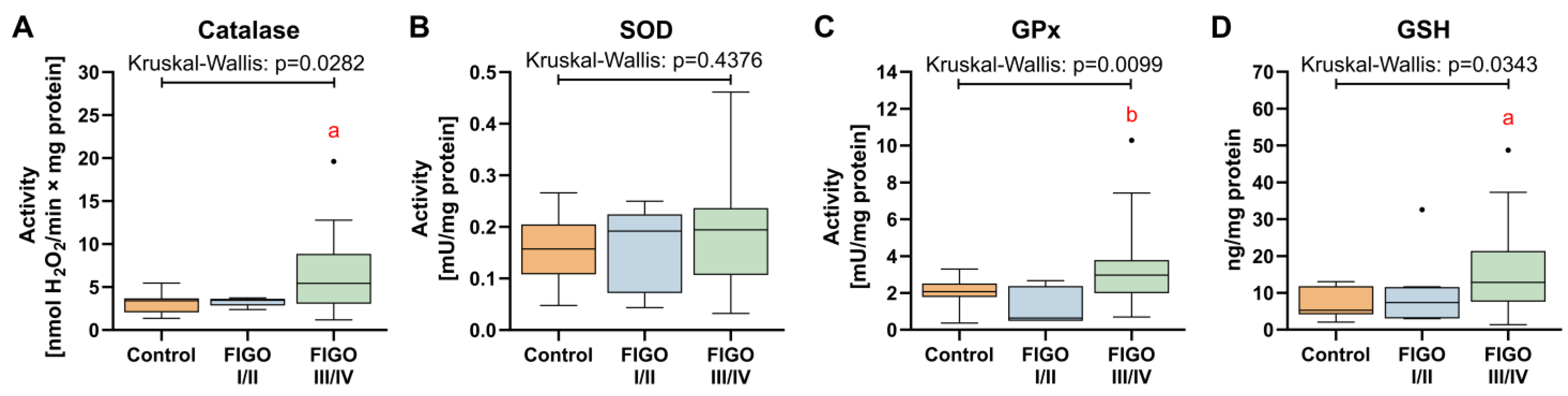
3.7. Antioxidant Defense System
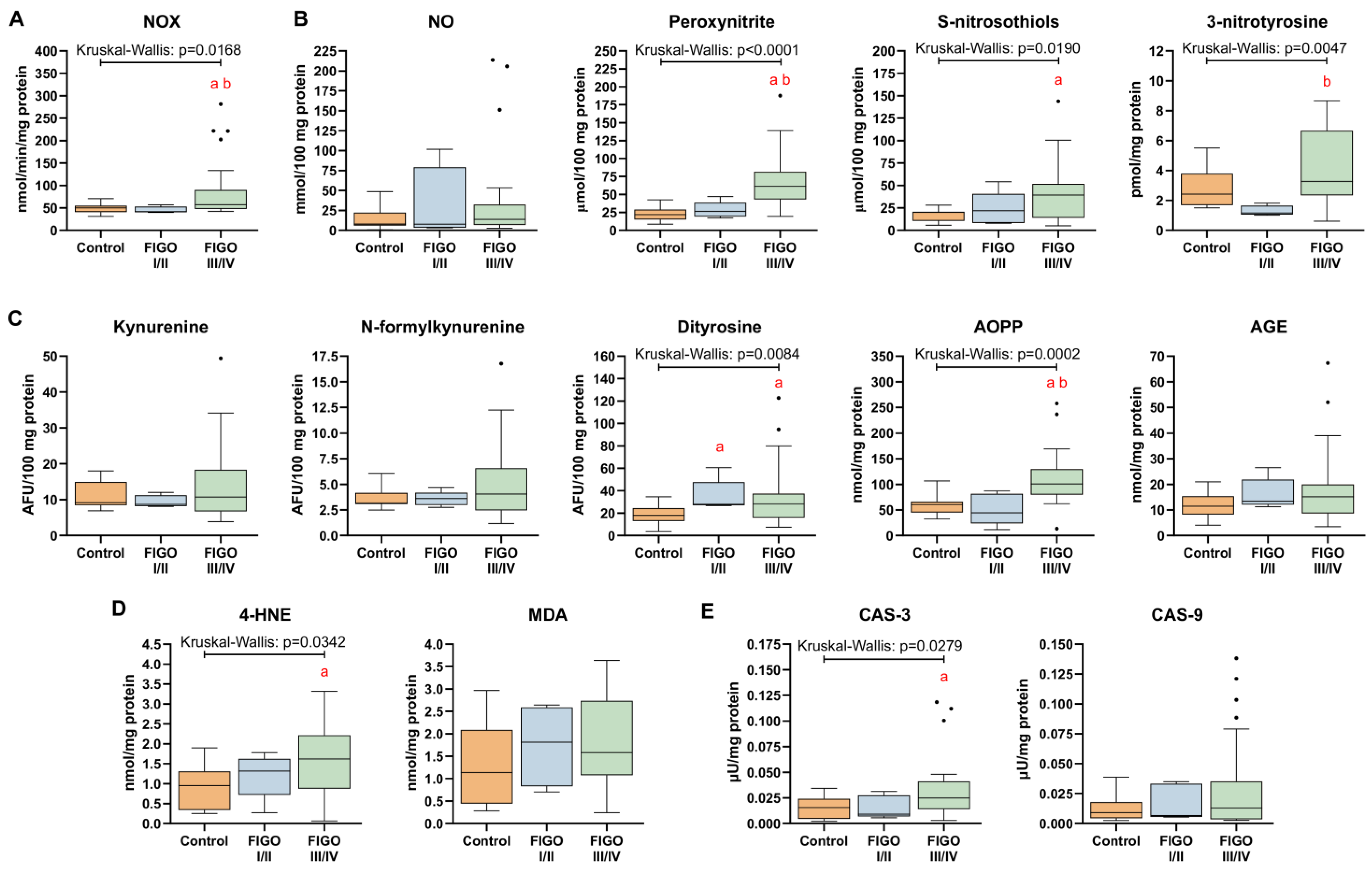
3.8. Markers of Nitrosative Stress and Oxidative Damage in HGSOC
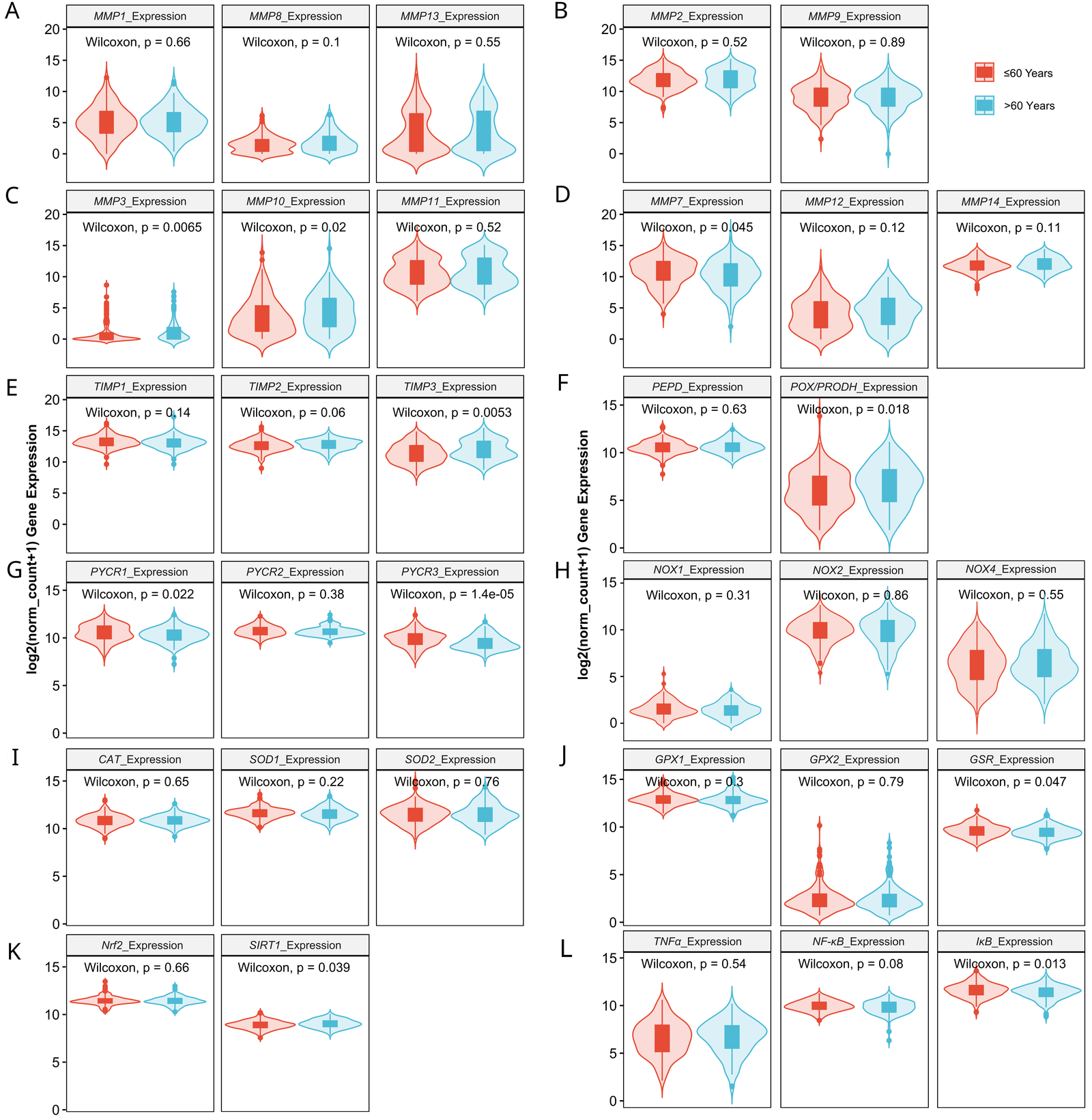
3.9. The Distribution of Redox Biomarkers in HGSOC by BMI, Patient’s Age and Presence of Metastases
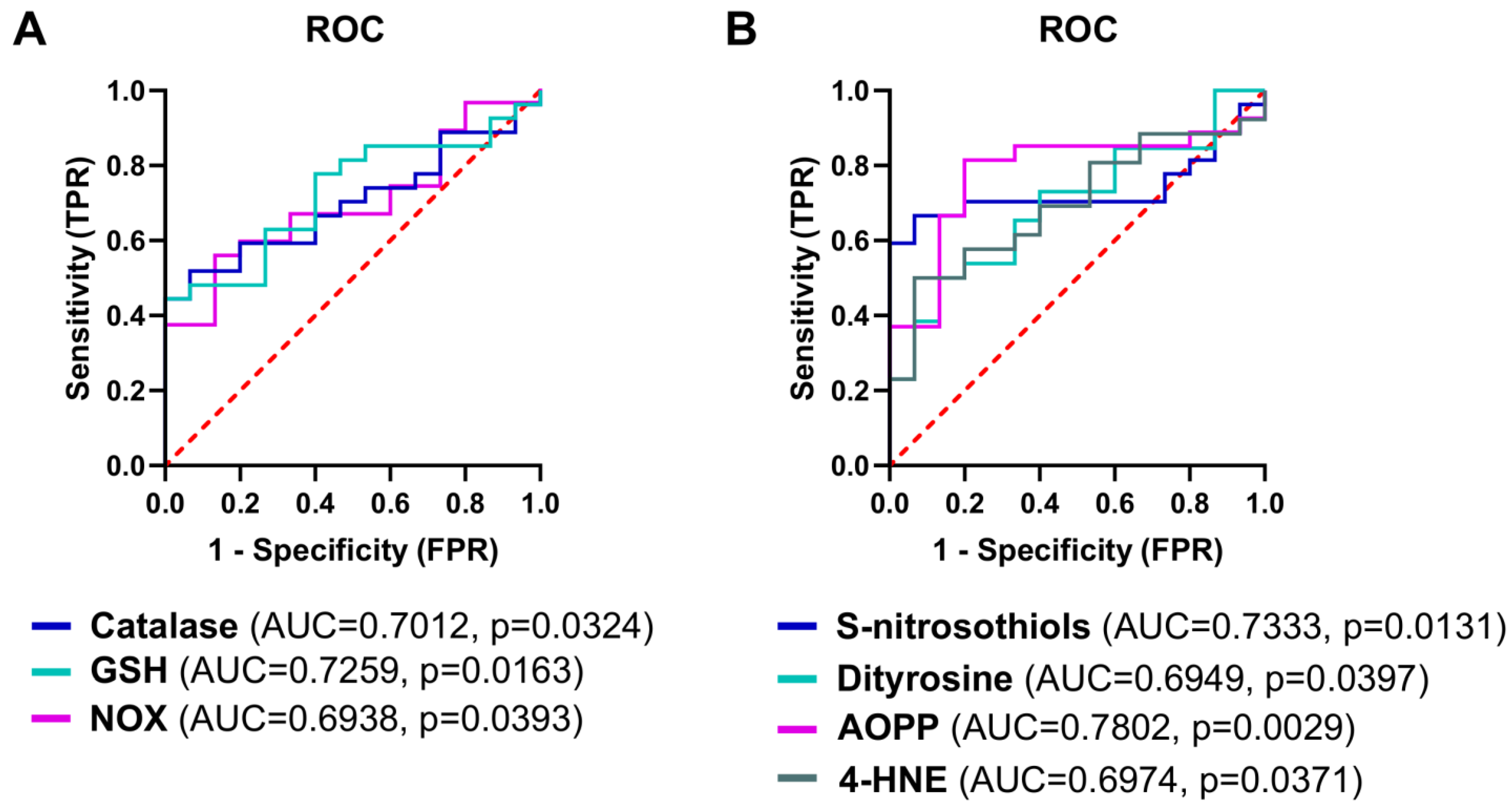
3.10. The Analysis of Redox Biomarkers for the Prediction of the HGSOC Occurrence
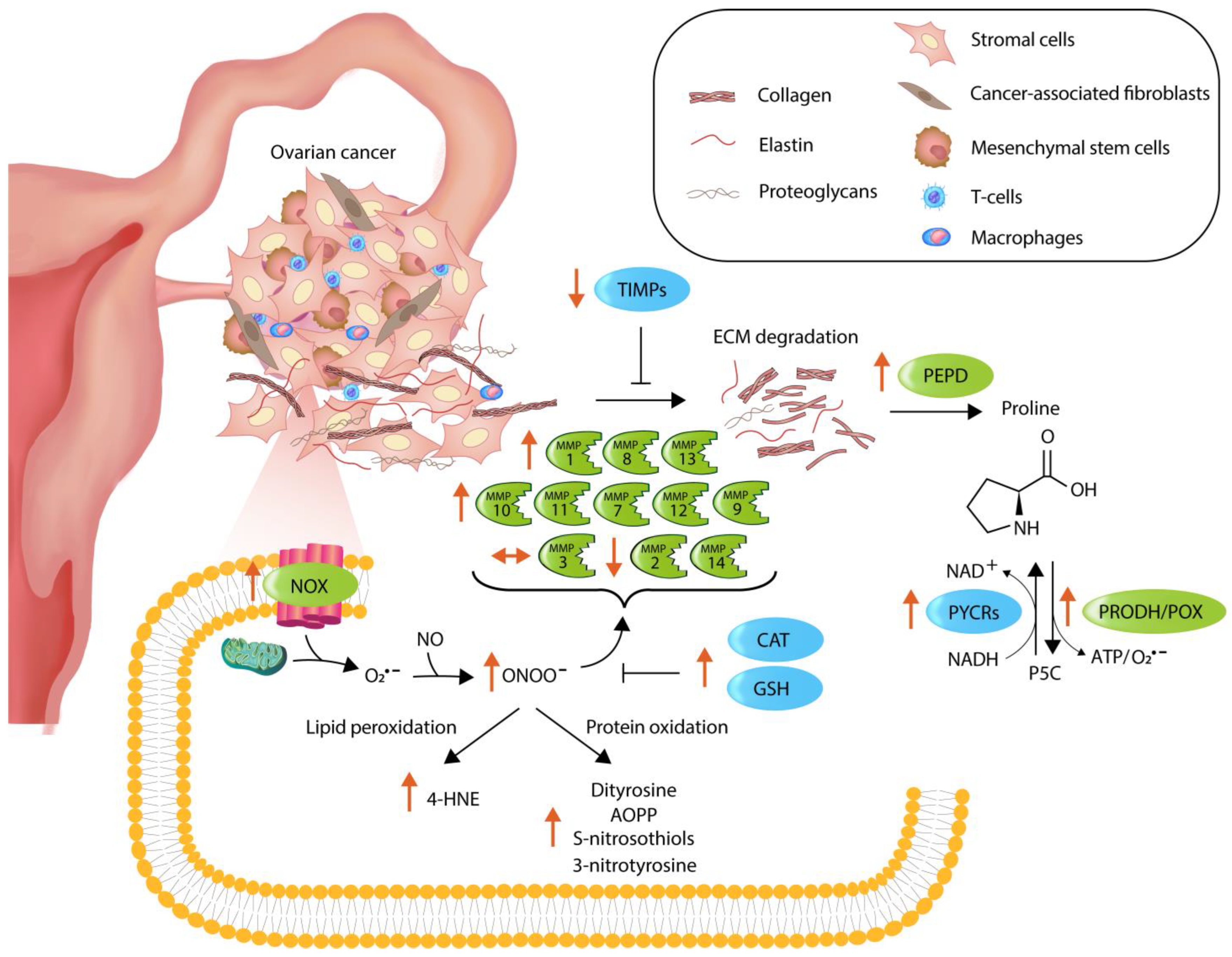
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, J.; Chan, W.C.; Ngai, C.H.; Lok, V.; Zhang, L.; Lucero-Prisno, D.E.; Xu, W.; Zheng, Z.J.; Elcarte, E.; Withers, M.; et al. Worldwide Burden, Risk Factors, and Temporal Trends of Ovarian Cancer: A Global Study. Cancers 2022, 14, 2230. [Google Scholar] [CrossRef] [PubMed]
- Kuroki, L.; Guntupalli, S.R. Treatment of epithelial ovarian cancer. BMJ 2020, 371, m3773. [Google Scholar] [CrossRef]
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 14, 9–32. [Google Scholar] [CrossRef]
- Desai, A.; Xu, J.; Aysola, K.; Qin, Y.; Okoli, C.; Hariprasad, R.; Chinemerem, U.; Gates, C.; Reddy, A.; Danner, O.; et al. Epithelial ovarian cancer: An overview. World J. Transl. Med. 2014, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, J.A.; Raja, F.A.; Fotopoulou, C.; Gonzalez-Martin, A.; Colombo, N.; Sessa, C. Newly diagnosed and relapsed epithelial ovarian carcinoma: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 24, vi24–vi32. [Google Scholar] [CrossRef]
- Harter, P.; Mouret-Reynier, M.A.; Pignata, S.; Cropet, C.; González-Martín, A.; Bogner, G.; Fujiwara, K.; Vergote, I.; Colombo, N.; Nøttrup, T.J.; et al. Efficacy of maintenance olaparib plus bevacizumab according to clinical risk in patients with newly diagnosed, advanced ovarian cancer in the phase III PAOLA-1/ENGOT-ov25 trial. Gynecol. Oncol. 2022, 164, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef]
- López-Otín, C.; Palavalli, L.H.; Samuels, Y. Protective roles of matrix metalloproteinases: From mouse models to human cancer. Cell Cycle 2009, 8, 3657–3662. [Google Scholar] [CrossRef]
- Eni-Aganga, I.; Lanaghan, Z.M.; Balasubramaniam, M.; Dash, C.; Pandhare, J. PROLIDASE: A Review from Discovery to its Role in Health and Disease. Front. Mol. Biosci. 2021, 8, 723003. [Google Scholar] [CrossRef]
- Behrman, H. Oxidative stress and the ovary. J. Soc. Gynecol. Investig. 2001, 8, S40–S42. [Google Scholar] [CrossRef]
- White, M.; Cohen, J.; Hummel, C.; Burky, R.; Cruz, A.; Farias-Eisner, R. The Role of Oxidative Stress in Ovarian Cancer. In Cancer; Elsevier: Amsterdam, The Netherlands, 2014; pp. 41–50. ISBN 9780124052055. [Google Scholar]
- Takiguchi, S.; Sugino, N.; Kashida, S.; Yamagata, Y.; Nakamura, Y.; Kato, H. Rescue of the corpus luteum and an increase in luteal superoxide dismutase expression induced by placental luteotropins in the rat: Action of testosterone without conversion to estrogen. Biol. Reprod. 2000, 62, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Badana, A.K.; Malla, R. Reactive Oxygen Species: A Key Constituent in Cancer Survival. Biomark. Insights 2018, 13, 1177271918755391. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.N.; Xie, L.Z.; Shen, Y.; Li, J.; Guo, Y.; Fu, Y.; Liu, F.Y.; Han, F.J. Insights into the Role of Oxidative Stress in Ovarian Cancer. Oxid. Med. Cell. Longev. 2021, 2021, 8388258. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shigdar, S.; Gantier, M.P.; Hou, Y.; Wang, L.; Li, Y.; Al Shamaileh, H.; Yin, W.; Zhou, S.F.; Zhao, X.; et al. Cancer stem cell targeted therapy: Progress amid controversies. Oncotarget 2015, 6, 44191–44206. [Google Scholar] [CrossRef]
- Veskoukis, A.S.; Nikolaidis, M.G.; Kyparos, A.; Kouretas, D. Blood reflects tissue oxidative stress depending on biomarker and tissue studied. Free Radic. Biol. Med. 2009, 47, 1371–1374. [Google Scholar] [CrossRef]
- Aebi, H. Catalase In Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. BBA Gen. Subj. 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef]
- Grisham, M.B.; Johnson, G.G.; Lancaster, J.R. Quantitation of nitrate and nitrite in extracellular fluids. Methods Enzymol. 1996, 268, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Kalousová, M.; Škrha, J.; Zima, T. Advanced glycation end-products and advanced oxidation protein products in patients with diabetes mellitus. Physiol. Res. 2002, 51, 597–604. [Google Scholar]
- Diplock, A.T.; Symons, M.C.R.; Rice-Evans, C.A. Laboratory Techniques in Biochemistry and Molecular Biology. Lab. Tech. Biochem. Mol. Biol. 1993, 24, 2. [Google Scholar]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Meki, A.R.M.A.; Esmail, E.E.D.F.; Hussein, A.A.; Hassanein, H.M. Caspase-3 and heat shock protein-70 in rat liver treated with aflatoxin B1: Effect of melatonin. Toxicon 2004, 43, 93–100. [Google Scholar] [CrossRef]
- Supruniuk, E.; Mikłosz, A.; Chabowski, A. Pyrroloquinoline quinone modifies lipid profile, but not insulin sensitivity, of palmitic acid-treated L6 myotubes. Int. J. Mol. Sci. 2020, 21, 8382. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, E45. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, M.; Żebrowska, E.; Nesterowicz, M.; Supruniuk, E.; Choromańska, B.; Chabowski, A.; Żendzian-Piotrowska, M.; Zalewska, A. α-Lipoic Acid Reduces Ceramide Synthesis and Neuroinflammation in the Hypothalamus of Insulin-Resistant Rats, While in the Cerebral Cortex Diminishes the β-Amyloid Accumulation. J. Inflamm. Res. 2022, 15, 2295–2312. [Google Scholar] [CrossRef]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef]
- Wang, S.; Liu, X. The UCSCXenaTools R package: A toolkit for accessing genomics data from UCSC Xena platform, from cancer multi-omics to single-cell RNA-seq. J. Open Source Softw. 2019, 4, 1627. [Google Scholar] [CrossRef]
- Stone, R.L.; Nick, A.M.; McNeish, I.A.; Balkwill, F.; Han, H.D.; Bottsford-Miller, J.; Rupaimoole, R.; Armaiz-Pena, G.N.; Pecot, C.V.; Coward, J.; et al. Paraneoplastic Thrombocytosis in Ovarian Cancer. N. Engl. J. Med. 2012, 366, 610–618. [Google Scholar] [CrossRef]
- Giannelli, G.; Bergamini, C.; Marinosci, F.; Fransvea, E.; Quaranta, M.; Lupo, L.; Schiraldi, O.; Antonaci, S. Clinical role of MMP-2/TIMP-2 imbalance in hepatocellular carcinoma. Int. J. Cancer 2002, 97, 425–431. [Google Scholar] [CrossRef]
- Zhang, S.; Li, L.; Lin, J.Y.; Lin, H. Imbalance between expression of matrix metalloproteinase-9 and tissue inhibitor of metalloproteinase-1 in invasiveness and metastasis of human gastric carcinoma. World J. Gastroenterol. 2003, 9, 899–904. [Google Scholar] [CrossRef]
- Bȩdkowska, G.E.; Gacuta, E.; Zajkowska, M.; Głazewska, E.K.; Osada, J.; Szmitkowski, M.; Chrostek, L.; Dabrowska, M.; Ławicki, S. Plasma levels of MMP-7 and TIMP-1 in laboratory diagnostics and differentiation of selected histological types of epithelial ovarian cancers. J. Ovarian Res. 2017, 10, 39. [Google Scholar] [CrossRef]
- Liu, W.; Phang, J.M. Proline dehydrogenase (oxidase) in cancer. BioFactors 2012, 38, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.S.; Agarwal, R.; Kaye, S.B. Mechanisms of transcoelomic metastasis in ovarian cancer. Lancet Oncol. 2006, 7, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef]
- Pietrzak, J.; Wosiak, A.; Szmajda-Krygier, D.; Świechowski, R.; Łochowski, M.; Pązik, M.; Balcerczak, E. Correlation of TIMP1-MMP2/MMP9 Gene Expression Axis Changes with Treatment Efficacy and Survival of NSCLC Patients. Biomedicines 2023, 11, 1777. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Zhang, Q.; Liu, F.; Zhou, D. Prognostic value of MMP-2 for patients with ovarian epithelial carcinoma: A systematic review and meta-analysis. Arch. Gynecol. Obstet. 2017, 295, 689–696. [Google Scholar] [CrossRef]
- Verma, S. Matrix metalloproteinases and gastrointestinal cancers: Impacts of dietary antioxidants. World J. Biol. Chem. 2014, 5, 355. [Google Scholar] [CrossRef]
- Lim, J.; Luderer, U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Shao, L.; Yuan, C.; Jiang, C.-Y.; Liu, J.-Y.; Gao, C.; Gao, L.; Cui, Y.-G.; Jiang, S.-W.; Liu, J.-Y.; et al. Implication of Differential Peroxiredoxin 4 Expression with Age in Ovaries of Mouse and Human for Ovarian Aging. Curr. Mol. Med. 2016, 16, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Senthil, K.; Aranganathan, S.; Nalini, N. Evidence of oxidative stress in the circulation of ovarian cancer patients. Clin. Chim. Acta 2004, 339, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Lewandowicz, G.M.; Britt, P.; Elgie, A.W.; Williamson, C.J.; Coley, H.M.; Hall, A.G.; Sargent, J.M. Cellular glutathione content, in vitro chemoresponse, and the effect of BSO modulation in samples derived from patients with advanced ovarian cancer. Gynecol. Oncol. 2002, 85, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Serpa, J. Glutathione in Ovarian Cancer: A Double-Edged Sword. Int. J. Mol. Sci. 2018, 19, 1882. [Google Scholar] [CrossRef]
- Takenaka, M.; Furui, T.; Suzuki, N.; Koike, T.; Aoki, H.; Morishige, K.-I. Glutathione as a Prognostic Biomarker and a Potential Therapeutic Target for Ovarian Cancer. Open J. Obstet. Gynecol. 2022, 12, 56–66. [Google Scholar] [CrossRef]
- Godwin, A.K.; Meister, A.; O’Dwyer, P.J.; Huang, C.S.; Hamilton, T.C.; Anderson, M.E. High resistance to cisplatin in human ovarian cancer cell lines is associated with marked increase of glutathione synthesis. Proc. Natl. Acad. Sci. USA 1992, 89, 3070–3074. [Google Scholar] [CrossRef]
- Surowiak, P.; Materna, V.; Kaplenko, I.; Spaczyński, M.; Dietel, M.; Lage, H.; Zabel, M. Augmented expression of metallothionein and glutathione S-transferase pi as unfavourable prognostic factors in cisplatin-treated ovarian cancer patients. Virchows Arch. 2005, 447, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhu, L.; Hao, B.; Gao, W.; Wang, Q.; Li, K.; Wang, M.; Huang, M.; Liu, Z.; Yang, Q.; et al. iNOS-derived nitric oxide promotes glycolysis by inducing pyruvate kinase M2 nuclear translocation in ovarian cancer. Oncotarget. 2017, 16, 33047–33063. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Li, X.; Sun, Y.; Li, L.; Zhang, Q.; Zhu, L.; Zhong, Z.; Wang, M.; Wang, Q.; Liu, Z.; et al. NOS1 expression promotes proliferation and invasion and enhances chemoresistance in ovarian cancer. Oncol. Lett. 2020, 19, 2989–2995. [Google Scholar] [CrossRef]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef] [PubMed]
- Tcyganov, E.N.; Sanseviero, E.; Marvel, D.; Beer, T.; Tang, H.Y.; Hembach, P.; Speicher, D.W.; Zhang, Q.; Donthireddy, L.R.; Mostafa, A.; et al. Peroxynitrite in the tumor microenvironment changes the profile of antigens allowing escape from cancer immunotherapy. Cancer Cell 2022, 40, 1173–1189.e6. [Google Scholar] [CrossRef]
- Mishra, D.; Patel, V.; Banerjee, D. Nitric Oxide and S-Nitrosylation in Cancers: Emphasis on Breast Cancer. Breast Cancer Basic Clin. Res. 2020, 14, 117822341988268. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yasuoka, H.; Tsujimoto, M.; Yoshidome, K.; Nakahara, M.; Nakao, K.; Nakamura, M.; Kakudo, K. Nitric oxide in breast cancer: Induction of vascular endothelial growth factor-C and correlation with metastasis and poor prognosis. Clin. Cancer Res. 2006, 12, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G. Oxidative Stress and Lipid Peroxidation Products in Cancer Progression and Therapy. ISRN Oncol. 2012, 2012, 137289. [Google Scholar] [CrossRef]
- Cristani, M.; Speciale, A.; Saija, A.; Gangemi, S.; Minciullo, P.; Cimino, F. Circulating Advanced Oxidation Protein Products as Oxidative Stress Biomarkers and Progression Mediators in Pathological Conditions Related to Inflammation and Immune Dysregulation. Curr. Med. Chem. 2016, 23, 3862–3882. [Google Scholar] [CrossRef]
- Zavadskiy, S.; Sologova, S.; Moldogazieva, N. Oxidative distress in aging and age-related diseases: Spatiotemporal dysregulation of protein oxidation and degradation. Biochimie 2022, 195, 114–134. [Google Scholar] [CrossRef]
- Harris, I.S.; Endress, J.E.; Coloff, J.L.; Selfors, L.M.; McBrayer, S.K.; Rosenbluth, J.M.; Takahashi, N.; Dhakal, S.; Koduri, V.; Oser, M.G.; et al. Deubiquitinases Maintain Protein Homeostasis and Survival of Cancer Cells upon Glutathione Depletion. Cell Metab. 2019, 29, 1166–1181.e6. [Google Scholar] [CrossRef]
- Pizzimenti, S.; Ciamporcero, E.; Daga, M.; Pettazzoni, P.; Arcaro, A.; Cetrangolo, G.; Minelli, R.; Dianzani, C.; Lepore, A.; Gentile, F.; et al. Interaction of aldehydes derived from lipid peroxidation and membrane proteins. Front. Physiol. 2013, 4, 62208. [Google Scholar] [CrossRef]
- Karihtala, P.; Kauppila, S.; Puistola, U.; Jukkola-Vuorinen, A. Divergent behaviour of oxidative stress markers 8-hydroxydeoxyguanosine (8-OHdG) and 4-hydroxy-2-nonenal (HNE) in breast carcinogenesis. Histopathology 2011, 58, 854–862. [Google Scholar] [CrossRef]
- Harrington, B.S.; Annunziata, C.M. NF-κB Signaling in Ovarian Cancer. Cancers 2019, 11, 1182. [Google Scholar] [CrossRef]
- Sun, N.K.; Huang, S.L.; Chang, T.C.; Chao, C.C.K. TLR4 and NFκB signaling is critical for taxol resistance in ovarian carcinoma cells. J. Cell. Physiol. 2018, 233, 2489–2501. [Google Scholar] [CrossRef]
- Gonzalez-Torres, C.; Gaytan-Cervantes, J.; Vazquez-Santillan, K.; Mandujano-Tinoco, E.A.; Ceballos-Cancino, G.; Garcia-Venzor, A.; Zampedri, C.; Sanchez-Maldonado, P.; Mojica-Espinosa, R.; Jimenez-Hernandez, L.E.; et al. NF-κB Participates in the Stem Cell Phenotype of Ovarian Cancer Cells. Arch. Med. Res. 2017, 48, 343–351. [Google Scholar] [CrossRef]
- Chei, S.; Oh, H.J.; Jang, H.; Lee, K.; Jin, H.; Choi, Y.; Lee, B.Y. Korean red ginseng suppresses the expression of oxidative stress response and NLRP3 inflammasome genes in aged C57BL/6 mouse ovaries. Foods 2020, 9, 526. [Google Scholar] [CrossRef]
- He, L.; Long, X.; Yu, N.; Li, Y.; Liu, X.; Cheng, X. Premature Ovarian Insufficiency (POI) Induced by Dynamic Intensity Modulated Radiation Therapy via P13K-AKT-FOXO3a in Rat Models. Biomed. Res. Int. 2021, 2021, 7273846. [Google Scholar] [CrossRef]
- Topcu, A.; Balik, G.; Atak, M.; Mercantepe, T.; Uydu, H.A.; Tumkaya, L. An investigation of the effects of metformin on ovarian ischemia-reperfusion injury in rats. Eur. J. Pharmacol. 2019, 865, 172790. [Google Scholar] [CrossRef]
- Mao, P.; Smith, L.; Xie, W.; Wang, M. Dying endothelial cells stimulate proliferation of malignant glioma cells via a caspase 3-mediated pathway. Oncol. Lett. 2013, 5, 1615–1620. [Google Scholar] [CrossRef]
- Donato, A.L.; Huang, Q.; Liu, X.; Li, F.; Zimmerman, M.A.; Li, C.Y. Caspase 3 promotes surviving melanoma tumor cell growth after cytotoxic therapy. J. Investig. Dermatol. 2014, 134, 1686–1692. [Google Scholar] [CrossRef]
- Hu, Q.; Peng, J.; Liu, W.; He, X.; Cui, L.; Chen, X.; Yang, M.; Liu, H.; Liu, S.; Wang, H. Elevated cleaved caspase-3 is associated with shortened overall survival in several cancer types. Int. J. Clin. Exp. Pathol. 2014, 7, 5057–5070. [Google Scholar]
- Huang, Q.; Li, F.; Liu, X.; Li, W.; Shi, W.; Liu, F.F.; O’Sullivan, B.; He, Z.; Peng, Y.; Tan, A.C.; et al. Caspase 3-mediated stimulation of tumor cell repopulation during cancer radiotherapy. Nat. Med. 2011, 17, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Luo, L.L.; Xu, J.J.; Xu, M.Y.; Zhang, X.M.; Zhou, X.L.; Liu, W.J.; Fu, Y.C. Obesity accelerates ovarian follicle development and follicle loss in rats. Metabolism 2014, 63, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Kompella, P.; Vasquez, K.M. Obesity and cancer: A mechanistic overview of metabolic changes in obesity that impact genetic instability. Mol. Carcinog. 2019, 58, 1531–1550. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.C.; Nagle, C.M.; Thrift, A.P.; Pharoah, P.D.P.; Pearce, C.L.; Zheng, W.; Painter, J.N.; Chenevix-Trench, G.; Fasching, P.A.; Beckmann, M.W.; et al. Adult body mass index and risk of ovarian cancer by subtype: A Mendelian randomization study. Int. J. Epidemiol. 2016, 45, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.L.Y.; Dunning, K.R.; Yang, X.; Russell, D.L.; Lane, M.; Norman, R.J.; Robker, R.L. High-fat diet causes lipotoxicity responses in cumulus—Oocyte complexes and decreased fertilization rates. Endocrinology 2010, 151, 5438–5445. [Google Scholar] [CrossRef] [PubMed]
- Chartoumpekis, D.V.; Ziros, P.G.; Psyrogiannis, A.I.; Papavassiliou, A.G.; Kyriazopoulou, V.E.; Sykiotis, G.P.; Habeos, I.G. Nrf2 represses FGF21 during long-term high-fat diet—Induced obesity in mice. Diabetes 2011, 60, 2465–2473. [Google Scholar] [CrossRef]
- More, V.R.; Xu, J.; Shimpi, P.C.; Belgrave, C.; Luyendyk, J.P.; Yamamoto, M.; Slitt, A.L. Keap1 knockdown increases markers of metabolic syndrome after long-term high fat diet feeding. Free Radic. Biol. Med. 2013, 61, 85–94. [Google Scholar] [CrossRef]
- Illesca, P.; Valenzuela, R.; Espinosa, A.; Echeverría, F.; Soto-Alarcon, S.; Ortiz, M.; Videla, L.A. Hydroxytyrosol supplementation ameliorates the metabolic disturbances in white adipose tissue from mice fed a high-fat diet through recovery of transcription factors Nrf2, SREBP-1c, PPAR-γ and NF-κB. Biomed. Pharmacother. 2019, 109, 2472–2481. [Google Scholar] [CrossRef]
- He, H.-J. Curcumin attenuates Nrf2 signaling defect, oxidative stress in muscle and glucose intolerance in high fat diet-fed mice. World J. Diabetes 2012, 3, 94. [Google Scholar] [CrossRef]
- Czogalla, B.; Kahaly, M.; Mayr, D.; Schmoeckel, E.; Niesler, B.; Kolben, T.; Burges, A.; Mahner, S.; Jeschke, U.; Trillsch, F. Interaction of ERα and NRF2 impacts survival in ovarian cancer patients. Int. J. Mol. Sci. 2019, 20, 112. [Google Scholar] [CrossRef]
- Sindan, N.; Bhandari, A.; Zhao, Y.; Lu, X.; Lv, J. Expression and localization of nuclear factor erythroid 2-related factor 2 in the ovarian tissues of mice at different ages. Exp. Ther. Med. 2018, 16, 3546–3552. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Zhou, Z.; Zhong, Y.; Sun, Y.; Wang, Y.; Zhu, Z.; Jiao, W.; Bai, M.; Sun, J.; Yin, H.; et al. Plasma activity of Thioredoxin Reductase as a Novel Biomarker in Gastric Cancer. Sci. Rep. 2019, 9, 19084. [Google Scholar] [CrossRef] [PubMed]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Ovarian Cancer | Control | ||
|---|---|---|---|---|
| FIGO I/II | FIGO III/IV | |||
| Total | n = 15 | n = 5 | n = 25 | - |
| Age | 54.2 ± 11.99 | 64.2 ± 8.927 | 61.75 ± 11.05 | 0.0730 |
| BMI (kg/m2) | 26.76 ± 3.155 | 27.96 ± 4.17 | 28.42 ± 5.49 | 0.7170 |
| Overweight/obese | n = 10 | n = 3 | n = 18 | - |
| PLT (×103 cells/mm3) | 231.1 ± 43.43 | 247.6 ± 65.21 | 366.13 ± 113.17 | < 0.0001 |
| Fibrinogen (mg/dL) | 330.7 ± 54.4 | 444 ± 51.69 | 442.1 ± 108.6 | 0.0019 |
| K+ (mEq/L) | 4.203 ± 0.3137 | 4.778 ± 0.3762 | 4.616 ± 0.547 | 0.0073 |
| TSH (µU/mL) | 1.572 ± 0.6521 | 1.731 ± 0.3752 | 2.024 ± 1.225 | 0.5790 |
| Glucose (mg/dL) | 91.33 ± 9.713 | 93.6 ± 9.397 | 94.28 ± 14.96 | 0.9637 |
| SBP (mmHg) | 131.1 ± 17.69 | 133.8 ± 14.13 | 141.72 ± 19.22 | 0.3282 |
| DBP (mmHg) | 86.31 ± 8.702 | 84.2 ± 9.859 | 82.78 ± 12.29 | 0.6014 |
| Ca125 (U/mL) | 19.06 ± 8.366 | 243.8 ± 133.5 | 1161.32 ± 1022.9 | 0.0083 |
| He4 (U/mL) | 35.90 ± 14.00 | 218.3 ± 172.9 | 794.4 ± 689.9 | 0.0409 |
| Primary tumor volume (cm3) 1 | - | 767.2 ± 959.6 | 279.2 ± 488.3 | 0.0572 |
| Nodal invasion | - | - | n = 16 | - |
| Cancer cells in peritoneal fluid | - | - | n = 13 | - |
| ‘Omental-cake’ 2 | - | - | n = 13 | - |
| Nodal invasion > omental invasion 3 | - | - | n = 3 | - |
| Nodal invasion < omental invasion 4 | - | - | n = 8 | - |
| Time of hospitalization (day) | 4.357 ± 1.865 | 6.8 ± 2.49 | 10.435 ± 4.99 | 0.0001 |
| Overweight (n = 9) vs. Lean (n = 9) | Obese (n = 10) vs. Lean (n = 9) | Age ≥ 60 (n = 19) vs. Age < 60 (n = 11) | ≥2 Pregnancies (n = 9) vs. ≤1 Pregnancy (n = 4) | N1 + N2 (Lymph Node Metastasis; n = 16) vs. N0 (n = 14) | Nodal Invasion < Omental Invasion (n = 8) vs. Nodal Invasion > Omental Invasion (n = 3) | |
|---|---|---|---|---|---|---|
| Fold Change (p Value) | ||||||
| MMP2 | 0.124 (p = 0.905) | −0.030 (p = 0.941) | −0.734 (p = 0.154) | −0.500 (p = 0.799) | 0.660 (p = 0.050) | 1.375 (p = 0.235) |
| MMP3 | 1.141 (p = 0.095) | −3.330 (p = 0.044) | −1.958 (p = 0.973) | −1.050 (p = 0.931) | 0.440 (p = 0.782) | −0.594 (p = 0.376) |
| MMP7 | 0.965 (p = 0.661) | −0.132 (p = 0.656) | −0.396 (p = 0.773) | −1.515 (p = 0.874) | −0.476 (p = 0.405) | −0.839 (p = 0.012) |
| MMP11 | −2.498 (p = 0.021) | −0.270 (p = 0.766) | −2.338 (p = 0.012) | 0.342 (p = 0.620) | 1.556 (p = 0.115) | 0.762 (p > 0.999) |
| MMP14 | −0.194 (p = 0.400) | 0.823 (p = 0.370) | −1.098 (p = 0.318) | 0.031 (p = 0.642) | −0.176 (p = 0.165) | 0.573 (p = 0.024) |
| TIMP1 | −0.066 (p = 0.661) | 0.668 (p = 0.456) | −0.341 (p = 0.776) | 0.451 (p = 0.762) | −0.297 (p = 0.515) | 0.987 (p = 0.024) |
| TIMP2 | 0.297 (p = 0.905) | 1.348 (p = 0.456) | −1.056 (p = 0.559) | 0.979 (p = 0.292) | 0.160 (p = 0.209) | 1.659 (p = 0.012) |
| Nrf2 | 0.177 (p = 0.661) | 0.934 (p = 0.031) | −0.615 (p = 0.042) | 0.318 (p = 0.504) | −0.600 (p = 0.046) | 1.449 (p = 0.048) |
| IκB | 1.087 (p = 0.211) | 0.819 (p = 0.109) | 0.538 (p = 0.491) | −0.288 (p = 0.940) | −0.954 (p = 0.043) | 0.759 (p = 0.049) |
| NOX | −0.241 (p = 0.423) | 0.233 (p = 0.762) | −0.323 (p = 0.074) | −1.219 (p = 0.049) | 0.016 (p = 0.373) | 0.535 (p = 0.499) |
| Peroxynitrite | 0.127 (p = 0.917) | −0.071 (p = 0.747) | −0.751 (p = 0.223) | −1.634 (p = 0.214) | 0.198 (p = 0.829) | 0.631 (p = 0.460) |
| 3-nitrotyrosine | 0.471 (p = 0.645) | −0.083 (p = 0.965) | −0.899 (p = 0.219) | −0.775 (p = 0.497) | 0.594 (p = 0.036) | 1.096 (p = 0.048) |
| MDA | 1.342 (p = 0.139) | 1.042 (p = 0.042) | −0.759 (p = 0.339) | −0.614 (p = 0.683) | −0.318 (p = 0.905) | 0.877 (p = 0.383) |
| CAS-9 | −2.477 (p = 0.057) | −1.015 (p = 0.633) | −2.455 (p = 0.047) | −2.541 (p = 0.214) | 1.498 (p = 0.399) | −1.049 (p = 0.036) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Supruniuk, E.; Baczewska, M.; Żebrowska, E.; Maciejczyk, M.; Lauko, K.K.; Dajnowicz-Brzezik, P.; Milewska, P.; Knapp, P.; Zalewska, A.; Chabowski, A. Redox Biomarkers and Matrix Remodeling Molecules in Ovarian Cancer. Antioxidants 2024, 13, 200. https://doi.org/10.3390/antiox13020200
Supruniuk E, Baczewska M, Żebrowska E, Maciejczyk M, Lauko KK, Dajnowicz-Brzezik P, Milewska P, Knapp P, Zalewska A, Chabowski A. Redox Biomarkers and Matrix Remodeling Molecules in Ovarian Cancer. Antioxidants. 2024; 13(2):200. https://doi.org/10.3390/antiox13020200
Chicago/Turabian StyleSupruniuk, Elżbieta, Marta Baczewska, Ewa Żebrowska, Mateusz Maciejczyk, Kamil Klaudiusz Lauko, Patrycja Dajnowicz-Brzezik, Patrycja Milewska, Paweł Knapp, Anna Zalewska, and Adrian Chabowski. 2024. "Redox Biomarkers and Matrix Remodeling Molecules in Ovarian Cancer" Antioxidants 13, no. 2: 200. https://doi.org/10.3390/antiox13020200