
Anti-Inflammatory Effects of Idebenone Attenuate LPS-Induced Systemic Inflammatory Diseases by Suppressing NF-κB Activation
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Animal Experimentations
2.3. Hematoxylin and Eosin (HE) Staining
2.4. Immunohistochemistry for iNOS
2.5. Cells and Cell Culture
2.6. Assessment of NO Metabolite Levels
2.7. Western Blot Analysis
2.8. Preparation of Cytoplasmic and Nuclear Fractions
2.9. Determination of the Concentrations of TNF-α, IL-1β, IL-6, and PGE2
2.10. Immunofluorescence Assay
2.11. Measurement of ROS Production
2.12. Lipid Peroxidation Assay
2.13. Statistical Analysis
3. Results
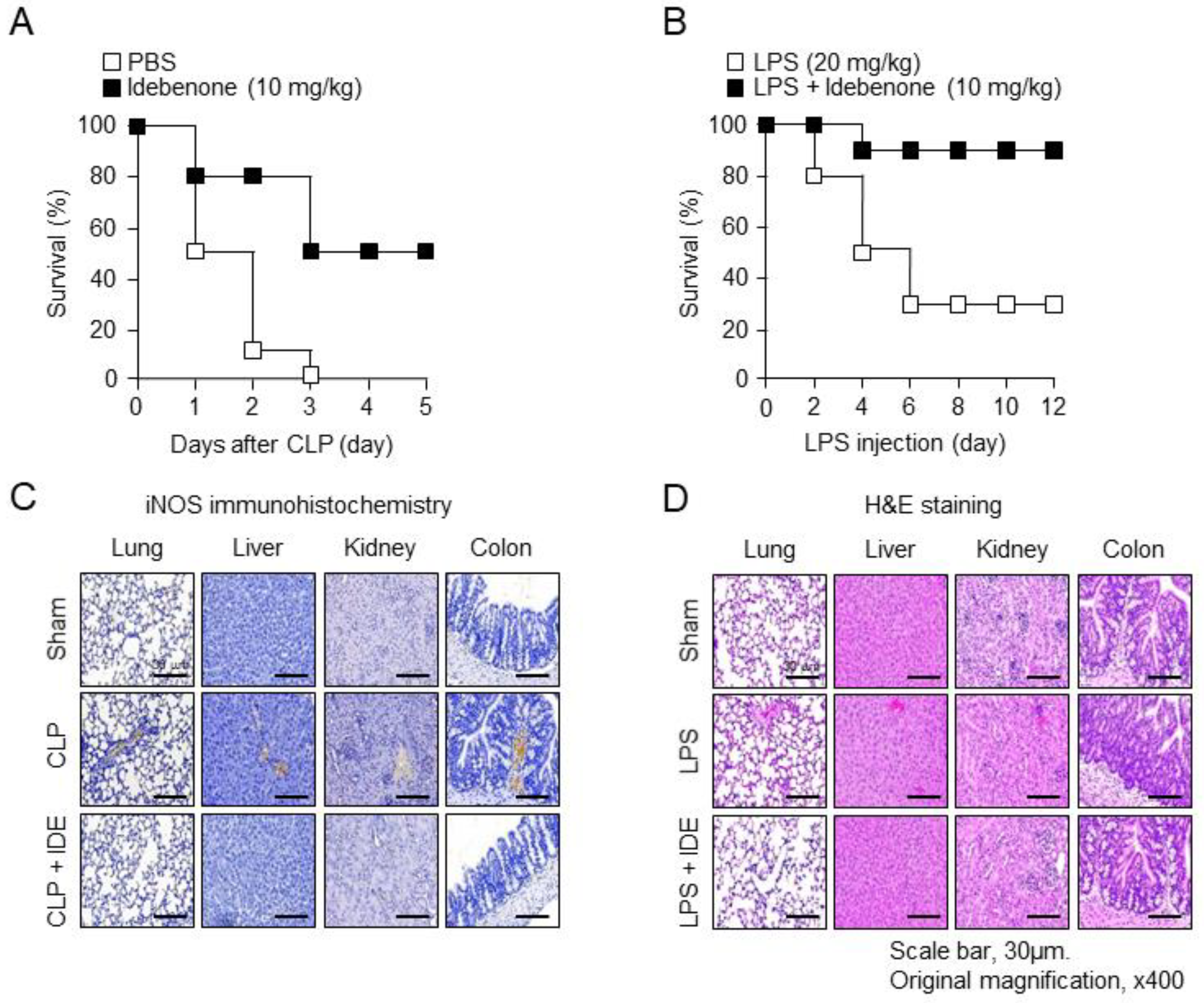
3.1. Idebenone Reduces Inflammatory Disease-Related Mortality and Protects against Tissue Damage In Vivo
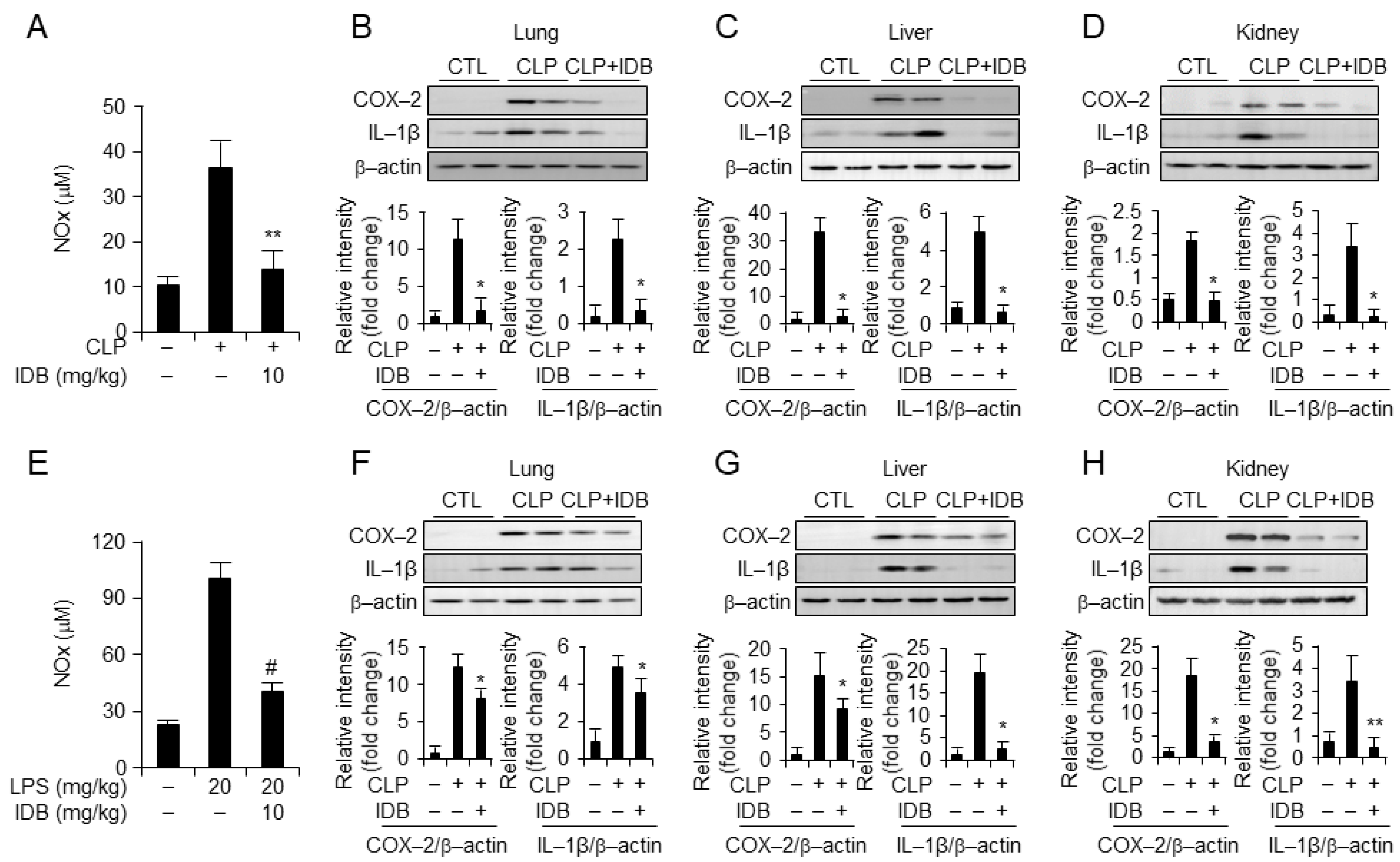
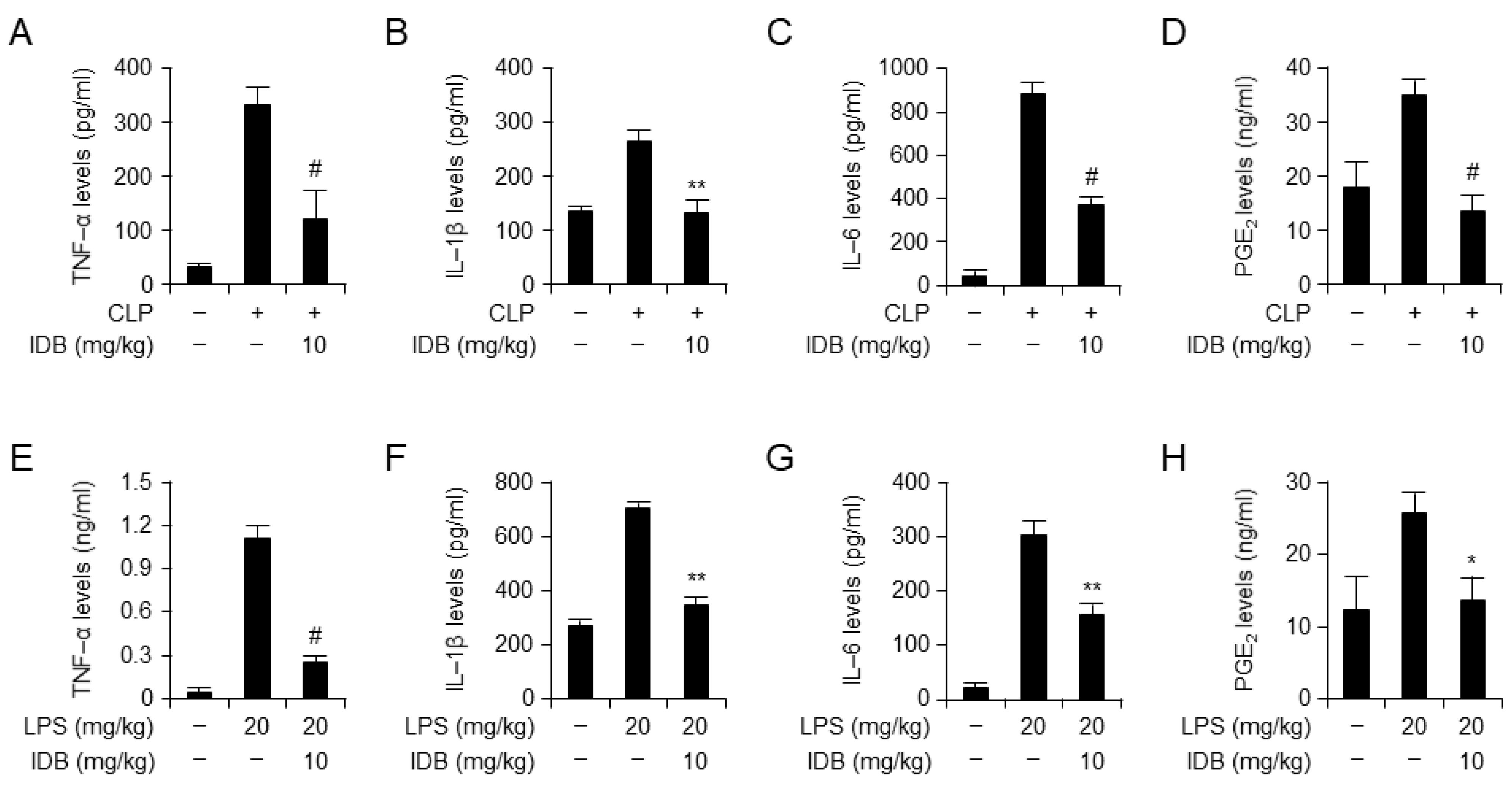
3.2. Idebenone Inhibits Inflammatory Responses in Mouse Models of CLP- and LPS-Induced Inflammation
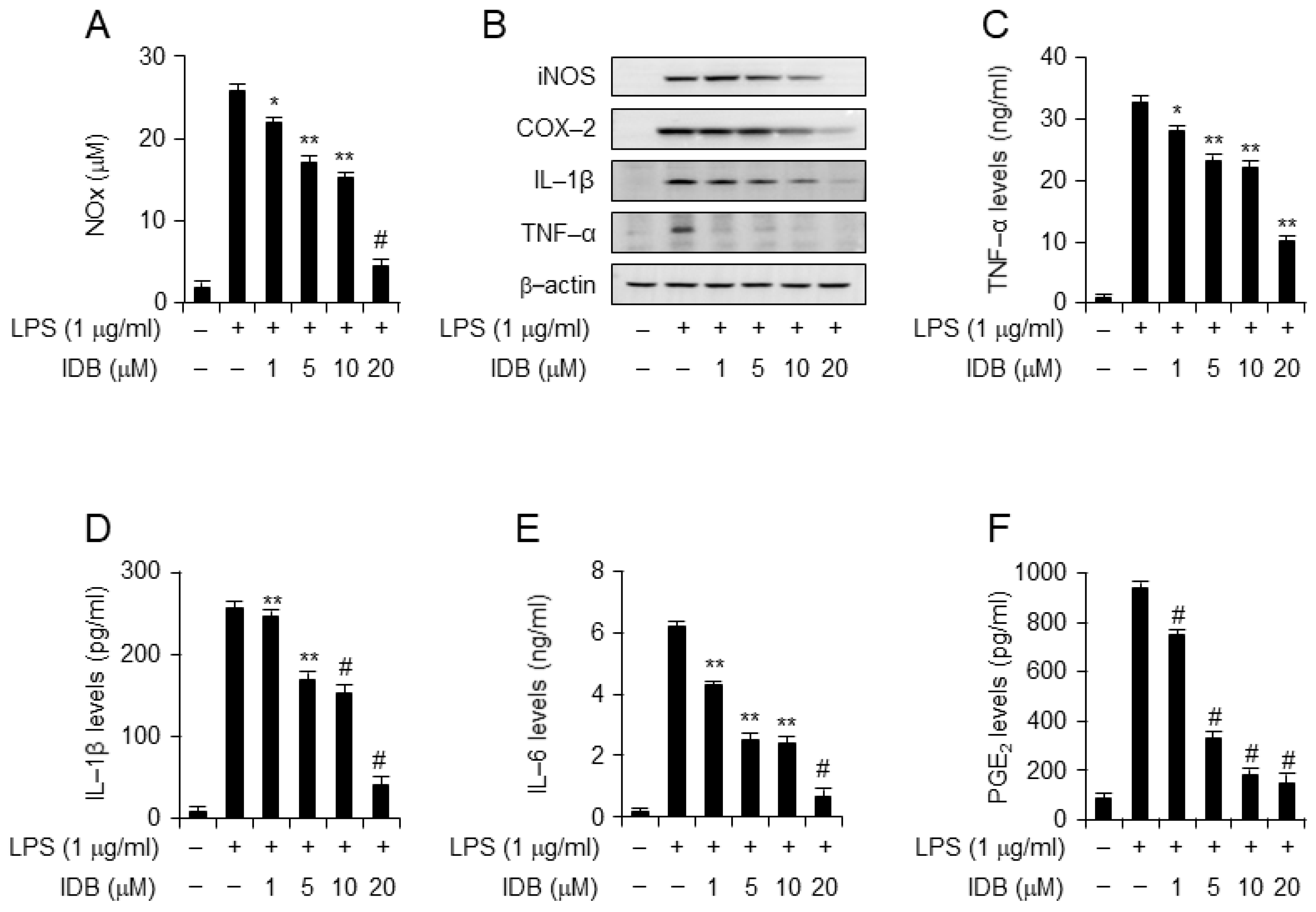
3.3. Idebenone Inhibits LPS-Induced Expression of Proinflammatory Mediators in Macrophages
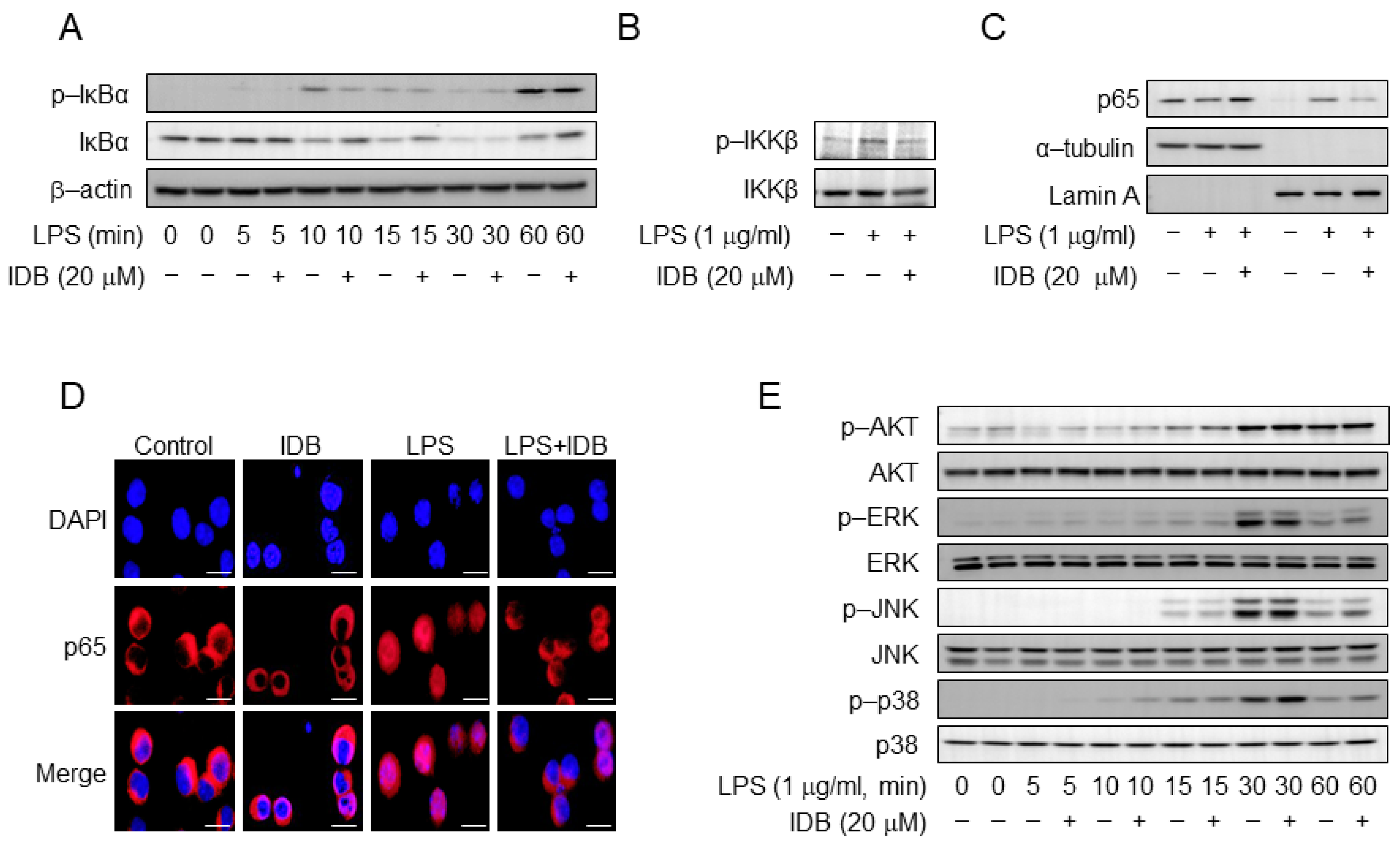
3.4. Idebenone Inhibites LPS-Induced NF-κB Activation in Macrophages
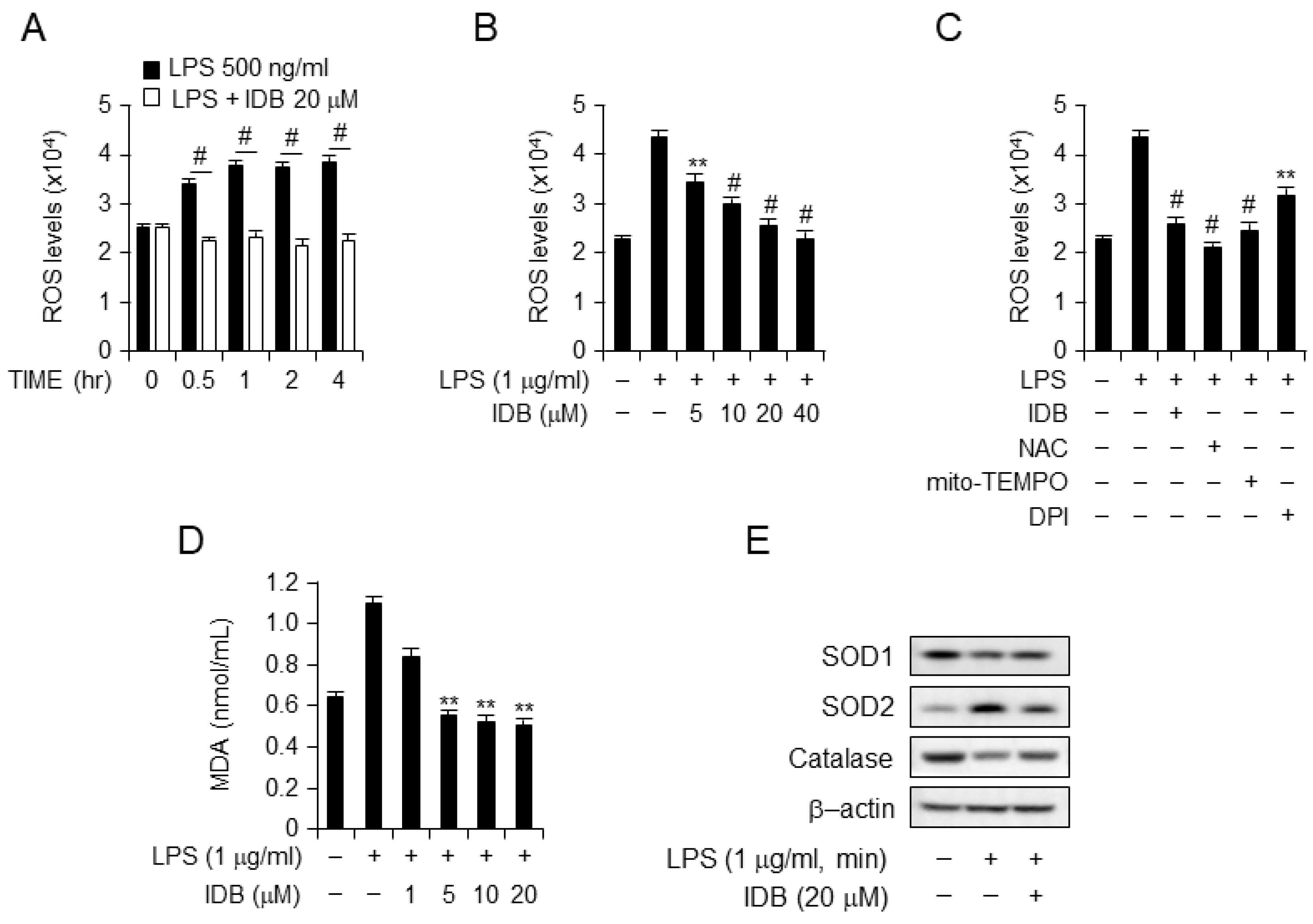
3.5. Idebenone Suppresses ROS Generation and Lipid Peroxidation in LPS-Stimulated RAW264.7 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Feghali, C.A.; Wright, T.M. Cytokines in acute and chronic inflammation. Front. Biosci. 1997, 2, d12–d26. [Google Scholar] [CrossRef] [PubMed]
- Delano, M.J.; Ward, P.A. The immune system’s role in sepsis progression, resolution, and long-term outcome. Immunol. Rev. 2016, 274, 330–353. [Google Scholar] [CrossRef]
- Horiguchi, H.; Loftus, T.J.; Hawkins, R.B.; Raymond, S.L.; Stortz, J.A.; Hollen, M.K.; Weiss, B.P.; Miller, E.S.; Bihorac, A.; Larson, S.D. Innate immunity in the persistent inflammation, immunosuppression, and catabolism syndrome and its implications for therapy. Front. Immunol. 2018, 9, 595. [Google Scholar] [CrossRef] [PubMed]
- Van der Poll, T.; van de Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The immunopathology of sepsis and potential therapeutic targets. Nat. Rev. Immunol. 2017, 17, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, M.; Evans, L.; Levy, M.; Rhodes, A. Sepsis and septic shock. Lancet 2018, 392, 75–87. [Google Scholar] [CrossRef]
- Huang, M.; Cai, S.; Su, J. The pathogenesis of sepsis and potential therapeutic targets. Int. J. Mol. Sci. 2019, 20, 5376. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The role of the reactive oxygen species and oxidative stress in the pathomechanism of the age-related ocular diseases and other pathologies of the anterior and posterior eye segments in adults. Oxid. Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef]
- Ranneh, Y.; Ali, F.; Akim, A.M.; Hamid, H.A.; Khazaai, H.; Fadel, A. Crosstalk between reactive oxygen species and pro-inflammatory markers in developing various chronic diseases: A review. Appl. Biol. Chem. 2017, 60, 327–338. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Brière, J.-J.; Schlemmer, D.; Chretien, D.; Rustin, P. Quinone analogues regulate mitochondrial substrate competitive oxidation. Biochem. Biophys. Res. Commun. 2004, 316, 1138–1142. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Mariscal, F.M.; Arenas-de Larriva, A.P.; Limia-Perez, L.; Romero-Cabrera, J.L.; Yubero-Serrano, E.M.; López-Miranda, J. Coenzyme Q10 supplementation for the reduction of oxidative stress: Clinical implications in the treatment of chronic diseases. Int. J. Mol. Sci. 2020, 21, 7870. [Google Scholar] [CrossRef] [PubMed]
- Bhagavan, H.N.; Chopra, R.K. Plasma coenzyme Q10 response to oral ingestion of coenzyme Q10 formulations. Mitochondrion 2007, 7, S78–S88. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Liu, Z.; Song, L.; Wang, X.; Zhang, Y.; Wu, N.; Lin, J.; Liu, Y.; Liu, Z. Idebenone alleviates neuroinflammation and modulates microglial polarization in LPS-stimulated BV2 cells and MPTP-induced Parkinson’s disease mice. Front. Cell Neurosci. 2019, 12, 529. [Google Scholar] [CrossRef] [PubMed]
- Senin, U.; Parnetti, L.; Barbagallo-Sangiorgi, G.; Bartorelli, L.; Bocola, V.; Capurso, A.; Cuzzupoli, M.; Denaro, M.; Marigliano, V.; Tammaro, A.E. Idebenone in senile dementia of Alzheimer type: A multicentre study. Arch. Gerontol. Geriatr. 1992, 15, 249–260. [Google Scholar] [CrossRef]
- Jaber, S.; Polster, B.M. Idebenone and neuroprotection: Antioxidant, pro-oxidant, or electron carrier? J. Bioenerg. Biomembr. 2015, 47, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Gueven, N.; Woolley, K.; Smith, J. Border between natural product and drug: Comparison of the related benzoquinones idebenone and coenzyme Q10. Redox. Biol. 2015, 4, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Shastri, S.; Shinde, T.; Sohal, S.S.; Gueven, N.; Eri, R. Idebenone protects against acute murine colitis via antioxidant and anti-inflammatory mechanisms. Int. J. Mol. Sci. 2020, 21, 484. [Google Scholar] [CrossRef]
- Meier, T.; Buyse, G. Idebenone: An emerging therapy for Friedreich ataxia. J. Neurol. 2009, 256, 25–30. [Google Scholar] [CrossRef]
- Heitz, F.D.; Erb, M.; Anklin, C.; Robay, D.; Pernet, V.; Gueven, N. Idebenone protects against retinal damage and loss of vision in a mouse model of Leber’s hereditary optic neuropathy. PLoS ONE 2012, 7, e45182. [Google Scholar] [CrossRef]
- McDaniel, D.; Neudecker, B.; DiNardo, J.; Lewis, J.; Maibach, H. Clinical efficacy assessment in photodamaged skin of 0.5% and 1.0% idebenone. J. Cosmet. Dermatol. 2005, 4, 167–173. [Google Scholar] [CrossRef]
- Hausse, A.; Aggoun, Y.; Bonnet, D.; Sidi, D.; Munnich, A.; Rötig, A.; Rustin, P. Idebenone and reduced cardiac hypertrophy in Friedreich’s ataxia. Heart 2002, 87, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Park, G.L.; Park, M.; Min, J.-K.; Park, Y.-J.; Chung, S.W.; Lee, S.-J. Anisomycin protects against sepsis by attenuating IκB kinase-dependent NF-κB activation and inflammatory gene expression. BMB Rep. 2021, 54, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Al-Rasheed, N.M.; Mohamed, A.M.; Baky, N.A.A.; Al-Rasheed, N.M.; Mohammad, R.A. Potential impact of quercetin and idebenone against immuno-inflammatory and oxidative renal damage induced in rats by titanium dioxide nanoparticles toxicity. J. Oleo Sci. 2013, 62, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hong, Y.S.; Jeong, J.H.; Yang, E.J.; Jhun, J.Y.; Park, M.K.; Jung, Y.O.; Min, J.K.; Kim, H.Y.; Park, S.H. Coenzyme Q10 ameliorates pain and cartilage degradation in a rat model of osteoarthritis by regulating nitric oxide and inflammatory cytokines. PLoS ONE 2013, 8, e69362. [Google Scholar] [CrossRef] [PubMed]
- Akpinar, E.; Kutlu, Z.; Kose, D.; Aydin, P.; Tavaci, T.; Bayraktutan, Z.; Yuksel, T.N.; Yildirim, S.; Eser, G.; Dincer, B. Protective effects of idebenone against sepsis induced acute lung damage. J. Investig. Surg. 2022, 35, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Shastri, S.; Shinde, T.; Perera, A.P.; Gueven, N.; Eri, R. Idebenone protects against spontaneous chronic murine colitis by alleviating endoplasmic reticulum stress and inflammatory response. Biomedicines 2020, 8, 384. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Jia, Z.; Mahaney, J.E.; Ross, D.; Misra, H.P.; Trush, M.A.; Li, Y. The highly expressed and inducible endogenous NAD (P) H: Quinone oxidoreductase 1 in cardiovascular cells acts as a potential superoxide scavenger. Cardiovasc. Toxicol. 2007, 7, 202–211. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.-J.; Jana, C.K.; Das, N. Role of catalase in oxidative stress-and age-associated degenerative diseases. Oxid. Med. Cell Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef]
- Yang, H.; Yu, S.; Kim, J.; Baek, K.; Lee, Y.-R.; Lee, H.S.; Choi, W.I.; Sung, D. Facile Solvent-Free Preparation of Antioxidant Idebenone-Loaded Nanoparticles for Efficient Wound Healing. Pharmaceutics 2022, 14, 521. [Google Scholar] [CrossRef] [PubMed]
- Nikitaras, V.; Zander-Fox, D.; McPherson, N.O. Improving Sperm Oxidative Stress and Embryo Quality in Advanced Paternal Age Using Idebenone In Vitro—A Proof-of-Concept Study. Antioxidants 2021, 10, 1079. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.-g. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Marchi, S.; Guilbaud, E.; Tait, S.W.; Yamazaki, T.; Galluzzi, L. Mitochondrial control of inflammation. Nat. Rev. Immunol. 2023, 23, 159–173. [Google Scholar] [CrossRef]
- Lee, H.-j.; Park, J.-H.; Hoe, H.-S. Idebenone Regulates Aβ and LPS-Induced Neurogliosis and Cognitive Function Through Inhibition of NLRP3 Inflammasome/IL-1β Axis Activation. Front. Immunol. 2022, 13, 749336. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.; Cho, Y.-L.; Park, S.; Park, M.; Hong, K.-S.; Park, Y.J.; Lee, I.-A.; Chung, S.W.; Lee, H.; Lee, S.-J. Anti-Inflammatory Effects of Idebenone Attenuate LPS-Induced Systemic Inflammatory Diseases by Suppressing NF-κB Activation. Antioxidants 2024, 13, 151. https://doi.org/10.3390/antiox13020151
Choi Y, Cho Y-L, Park S, Park M, Hong K-S, Park YJ, Lee I-A, Chung SW, Lee H, Lee S-J. Anti-Inflammatory Effects of Idebenone Attenuate LPS-Induced Systemic Inflammatory Diseases by Suppressing NF-κB Activation. Antioxidants. 2024; 13(2):151. https://doi.org/10.3390/antiox13020151
Chicago/Turabian StyleChoi, Yumin, Young-Lai Cho, Sujeong Park, Minkyung Park, Keun-Seok Hong, Young Jun Park, In-Ah Lee, Su Wol Chung, Heedoo Lee, and Seon-Jin Lee. 2024. "Anti-Inflammatory Effects of Idebenone Attenuate LPS-Induced Systemic Inflammatory Diseases by Suppressing NF-κB Activation" Antioxidants 13, no. 2: 151. https://doi.org/10.3390/antiox13020151