
On the Role of ROS and Glutathione in the Mode of Action Underlying Nrf2 Activation by the Hydroxyanthraquinone Purpurin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Nrf2 CALUX Reporter Gene Assay
2.4. DCF-DA Assay for Intracellular ROS Formation
2.5. Modulation of Intracellular GSH Level in the Nrf2 CALUX Cells
2.6. WST-1 Assay
2.7. Acid Dissociation Constant (pKa) Prediction
2.8. pH-Dependent Incubation of Purpurin with Glutathione
2.9. Ultra-Performance Liquid Chromatography (UPLC) Analysis of Purpurin Glutathione Adduct Formation
2.10. Liquid Chromatography-Time of Flight-Mass Spectrometry (LC-TOF-MS) Analysis of Purpurin Glutathione Adduct Formation
2.11. Liquid Chromatography Mass Spectrometry (LC-MS/MS) Analysis of GSH and GSSG
2.12. DCF-DA Assay for Cell-Free ROS Formation of Purpurin in the Absence or Presence of GSH at Different Concentrations and Different pH Values
2.13. Data Analysis
3. Results
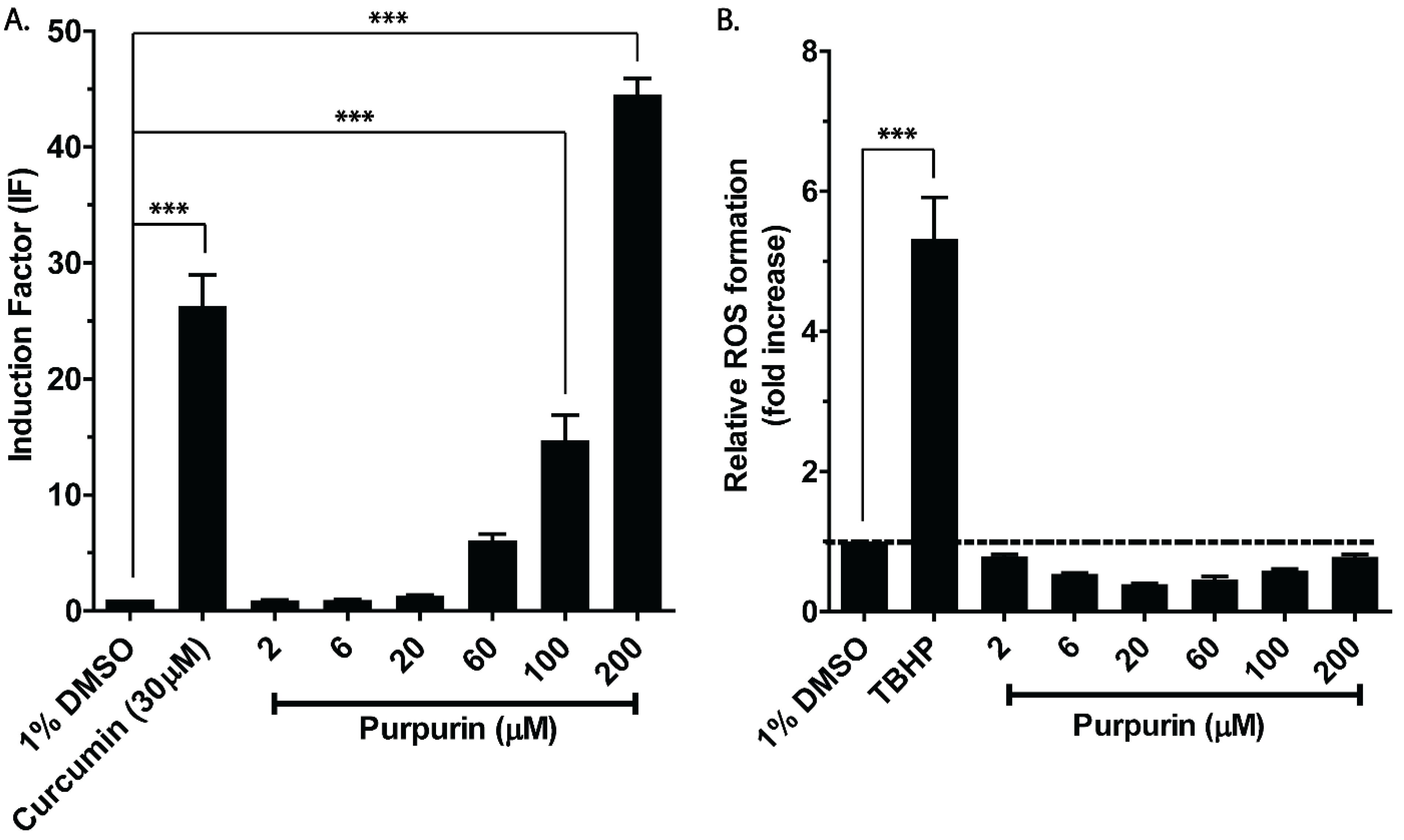
3.1. Nrf2 Activation and Intracellular ROS Formation of Purpurin
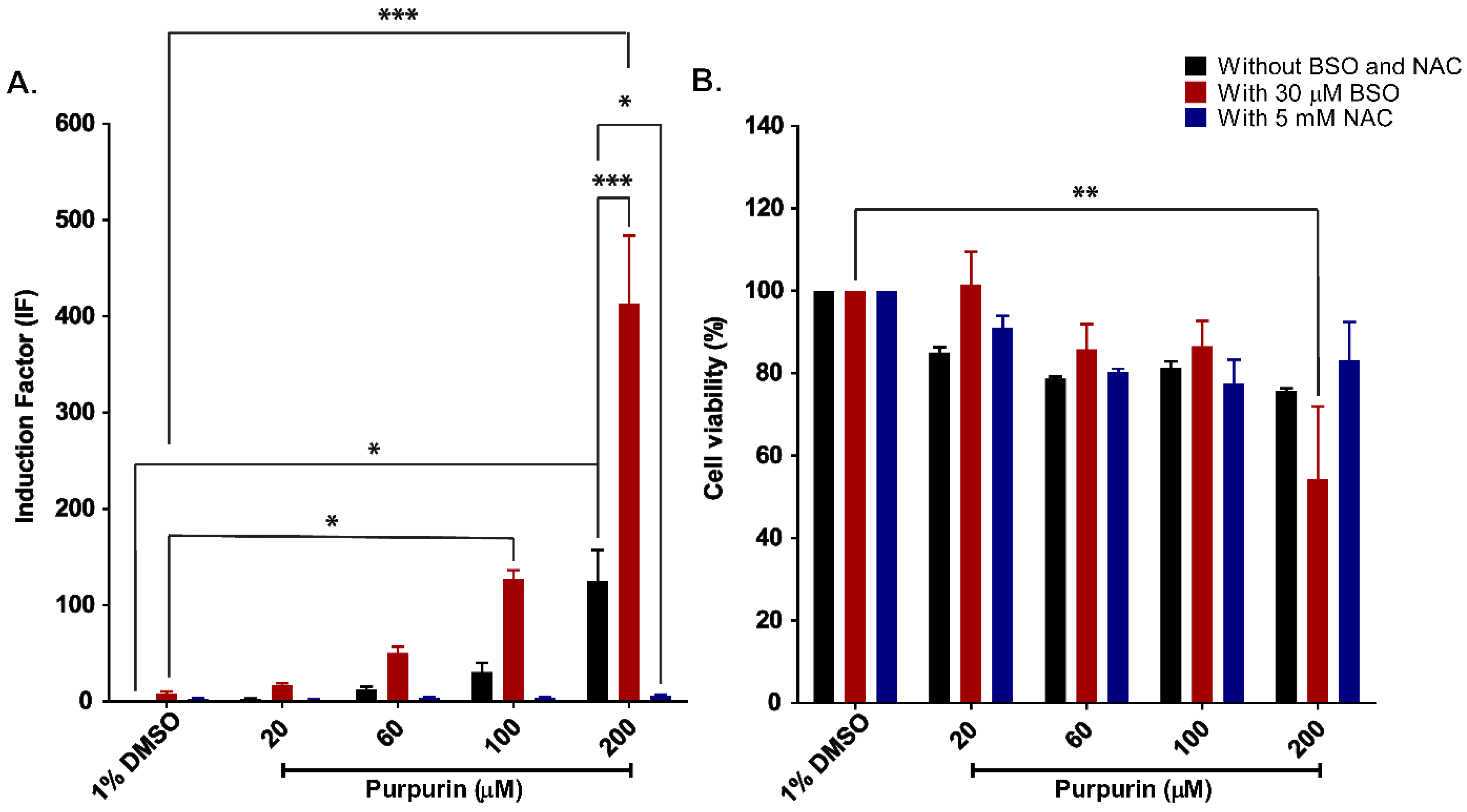
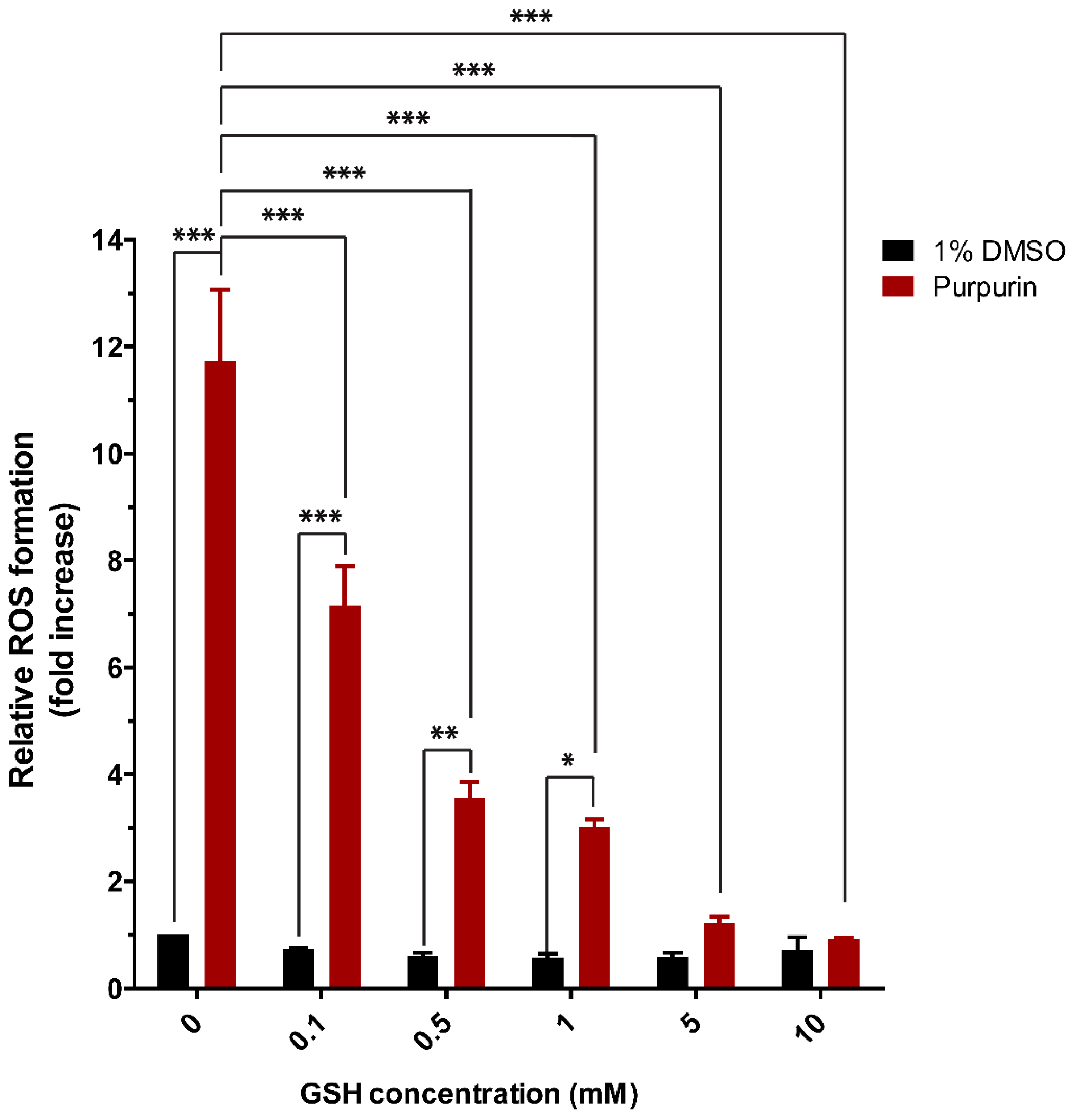
3.2. Nrf2 Activation and Cytotoxicity of Purpurin in Nrf2 CALUX Cells with and without Modulated Intracellular GSH Levels
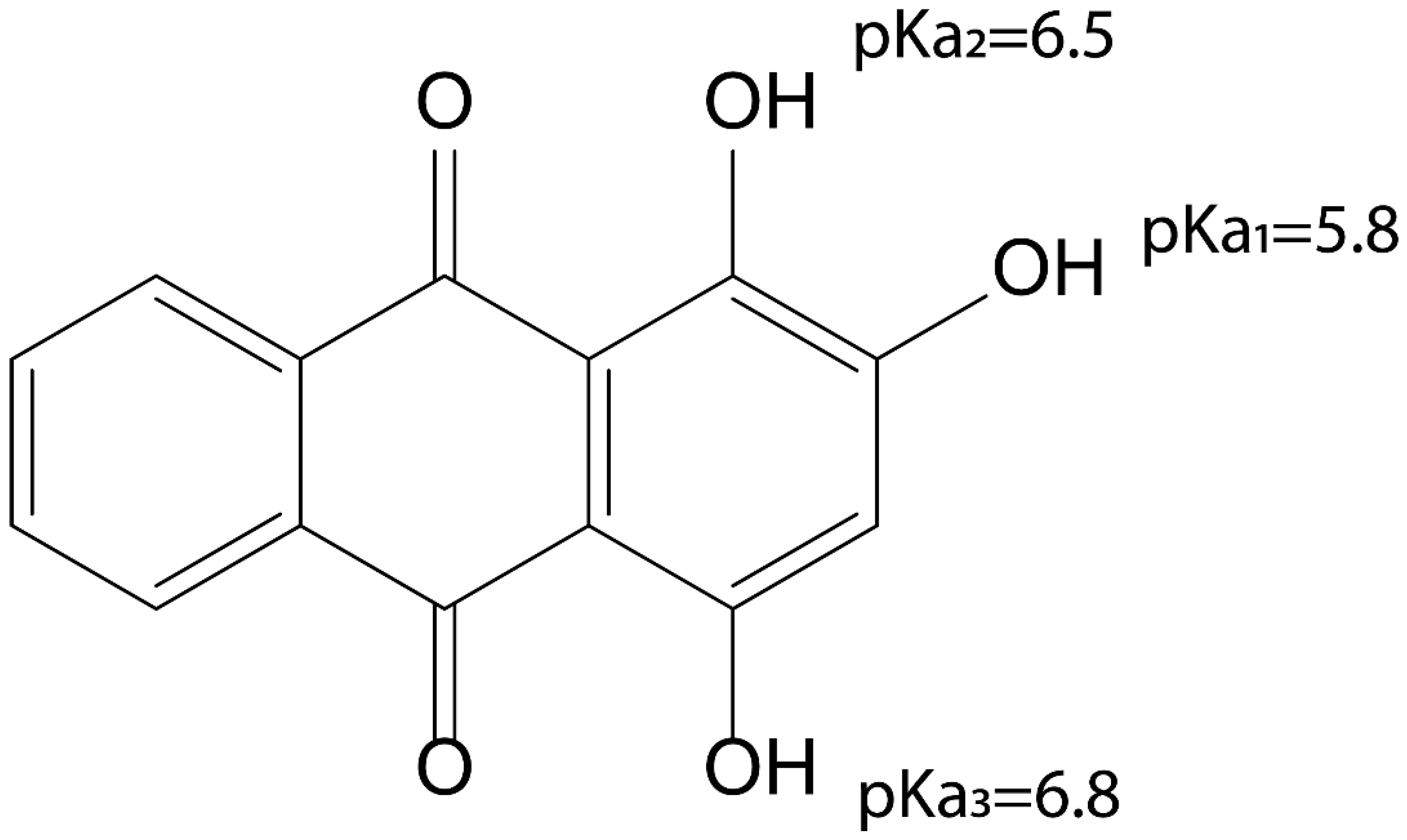
3.3. pKa of Purpurin
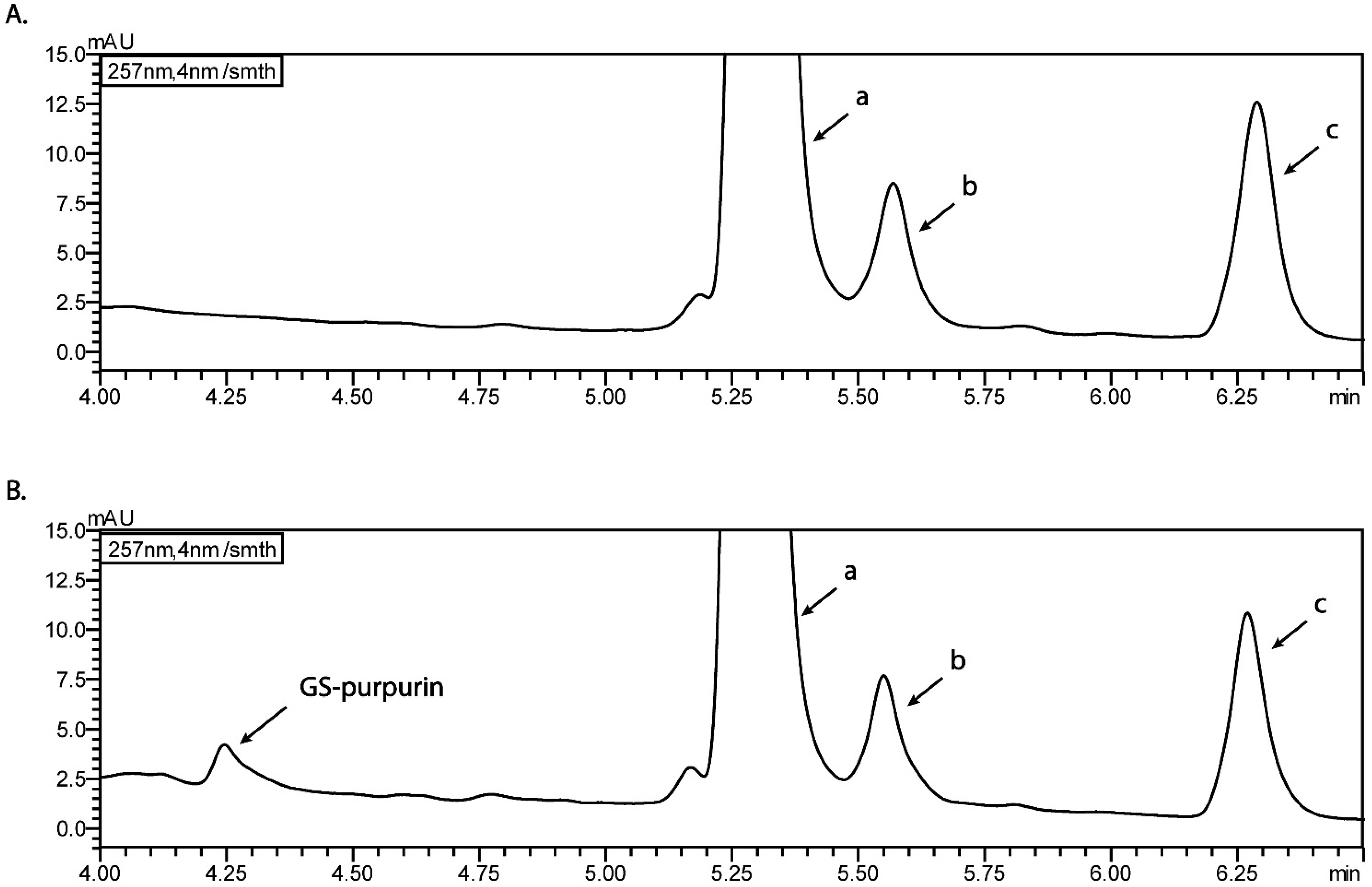
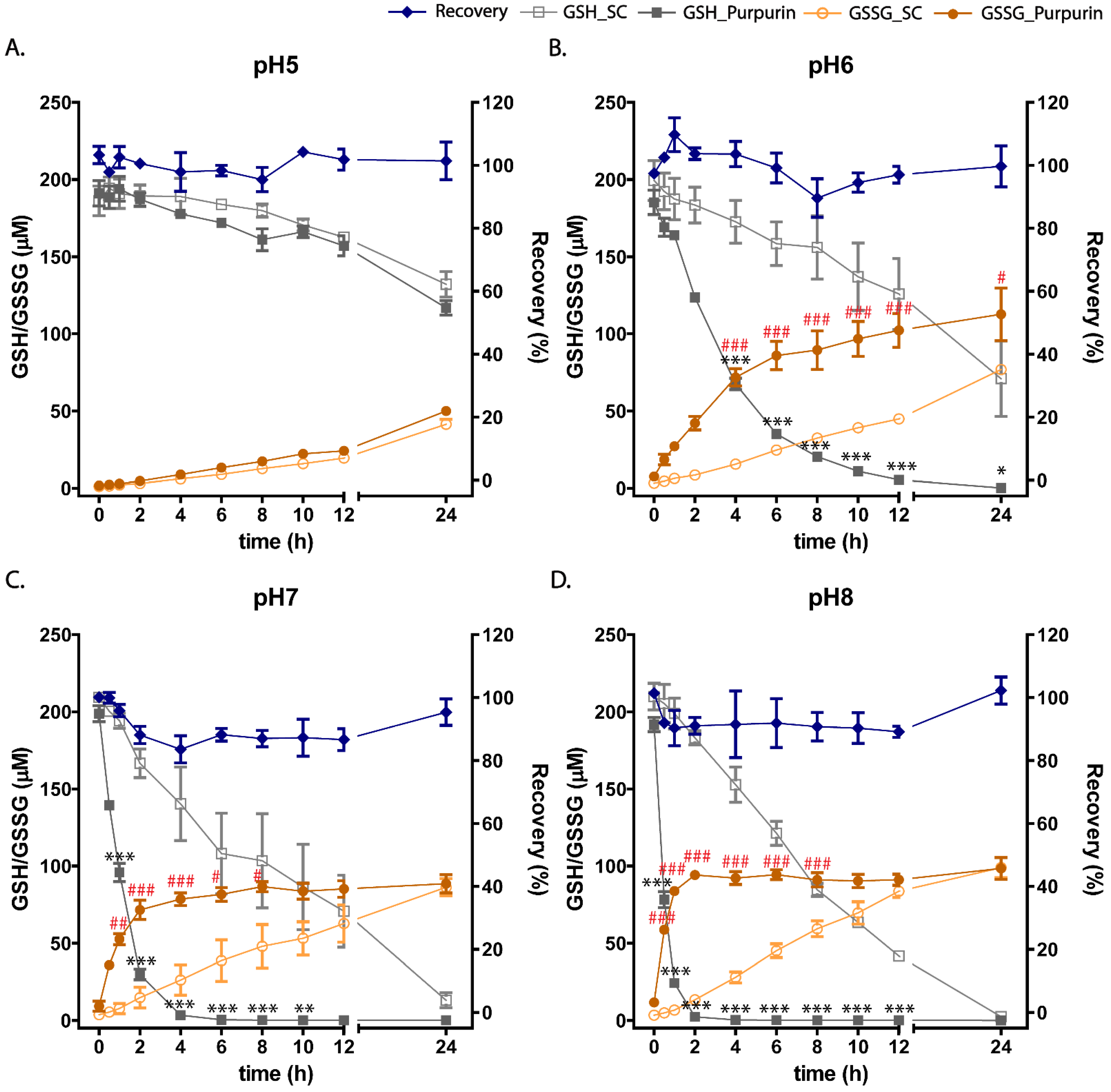
3.4. pH Dependent Cell-Free Incubation Purpurin and GSH
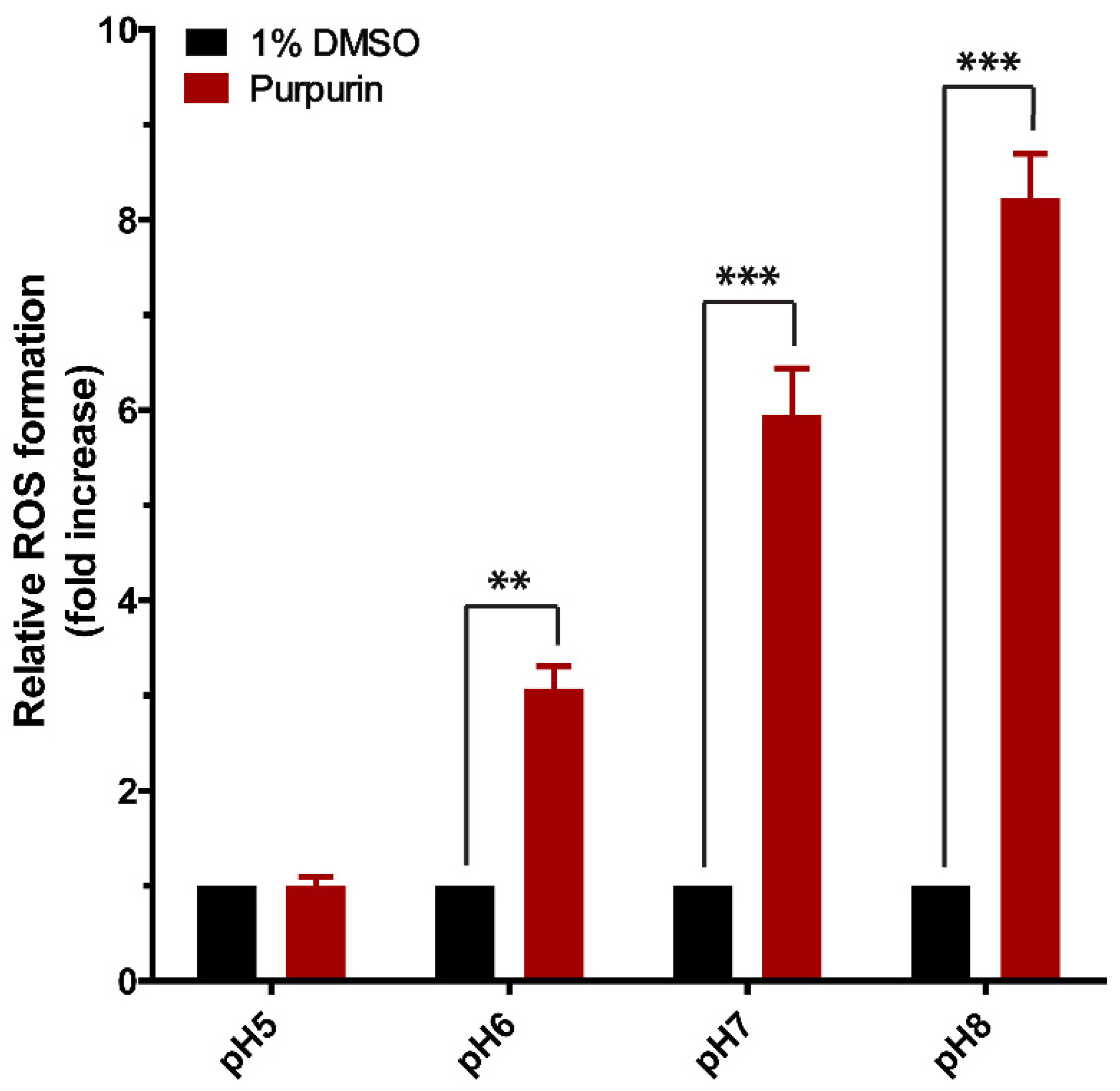
3.5. pH Dependent Cell-Free ROS Formation Induced by Purpurin
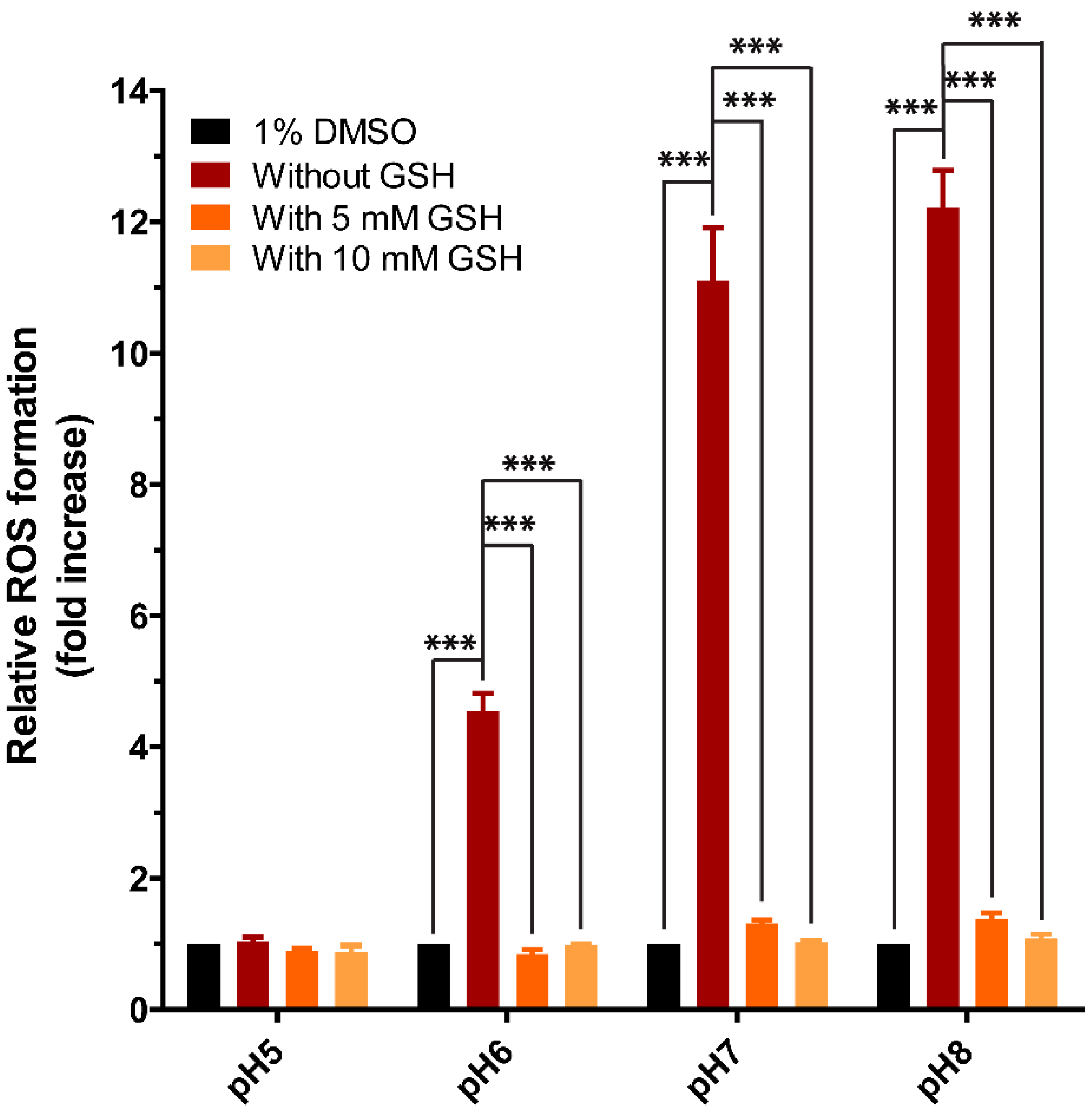
3.6. Cell-Free ROS Formation in Incubations of Purpurin with GSH at Different pH
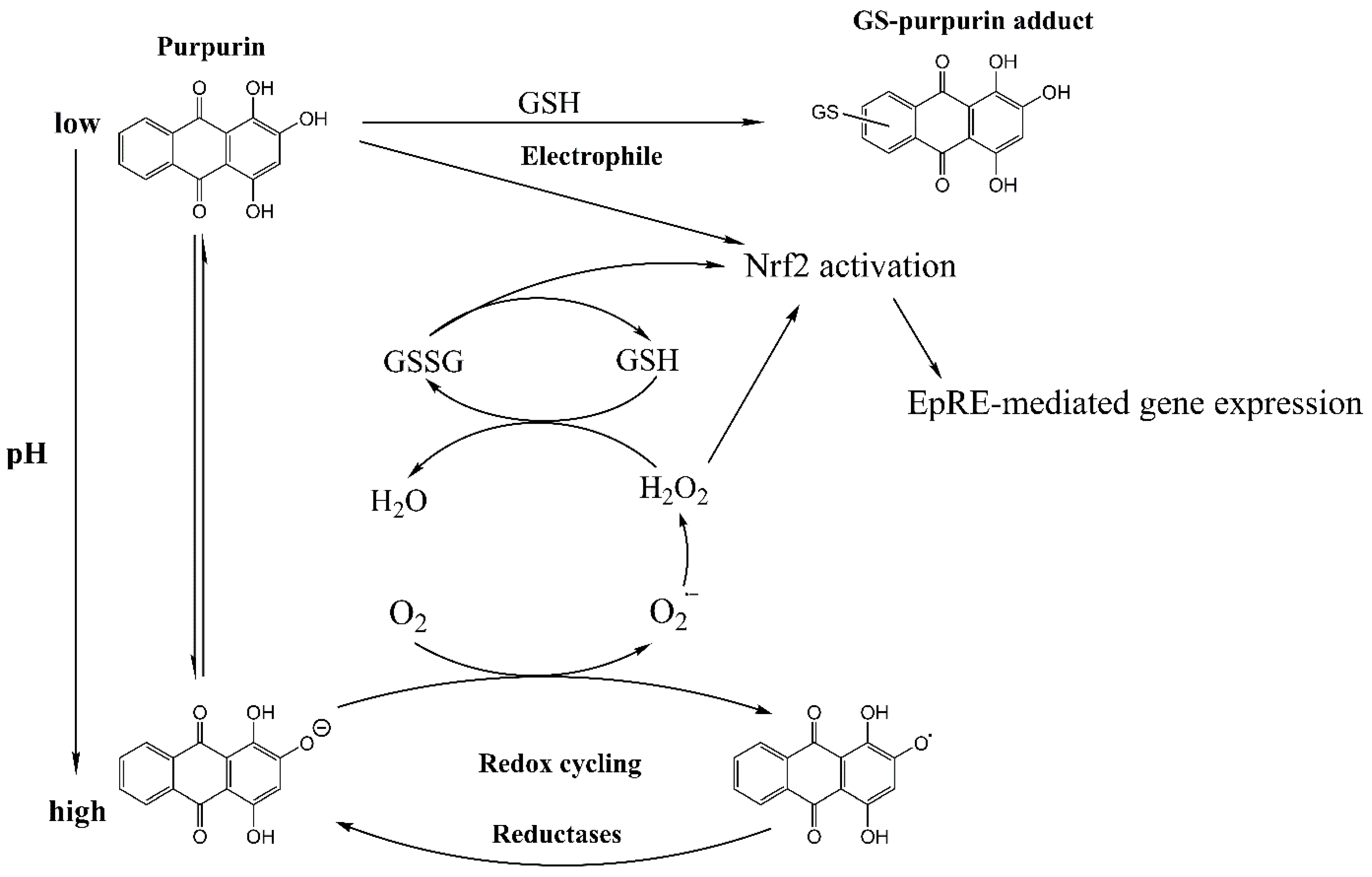
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Awad, H.M.; Boersma, M.G.; Boeren, S.; van Bladeren, P.J.; Vervoort, J.; Rietjens, I.M.C.M. The Regioselectivity of Glutathione Adduct Formation with Flavonoid Quinone/Quinone Methides Is pH-Dependent. Chem. Res. Toxicol. 2002, 15, 343–351. [Google Scholar] [CrossRef]
- Bo, S.; Lai, J.; Lin, H.; Luo, X.; Zeng, Y.; Du, T. Purpurin, a anthraquinone induces ROS-mediated A549 lung cancer cell apoptosis via inhibition of PI3K/AKT and proliferation. J. Pharm. Pharmacol. 2021, 73, 1101–1108. [Google Scholar] [CrossRef]
- Bolton, J.L.; Trush, M.A.; Penning, T.M.; Dryhurst, G.; Monks, T.J. Role of Quinones in Toxicology. Chem. Res. Toxicol. 2000, 13, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Chun, K.-S.; Raut, P.K.; Kim, D.-H.; Surh, Y.-J. Role of chemopreventive phytochemicals in NRF2-mediated redox homeostasis in humans. Free Radic. Biol. Med. 2021, 172, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Cores, Á.; Carmona-Zafra, N.; Clerigué, J.; Villacampa, M.; Menéndez, J.C. Quinones as Neuroprotective Agents. Antioxidants 2023, 12, 1464. [Google Scholar] [CrossRef]
- Devi Priya, M.; Siril, E.A. Traditional and modern use of indian madder (Rubia cordifolia L.): An overview. Int. J. Pharm. Sci. Rev. Res. 2014, 25, 154–164. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Zhang, Y.; Naidu, S.D.; Kostov, R.V.; Pheely, A.; Calabrese, V. Sulfhydryl-reactive phytochemicals as dual activators of transcription factors NRF2 and HSF1. In 50 Years of Phytochemistry Research; Springer: Cham, Switzerland, 2013; Volume 43, pp. 95–119. [Google Scholar] [CrossRef]
- Dong, X.; Fu, J.; Yin, X.; Qu, C.; Yang, C.; He, H.; Ni, J. Induction of Apoptosis in HepaRG Cell Line by Aloe-Emodin through Generation of Reactive Oxygen Species and the Mitochondrial Pathway. Cell. Physiol. Biochem. 2017, 42, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Eggler, A.L.; Savinov, S.N. Chemical and biological mechanisms of phytochemical activation of Nrf2 and importance in disease prevention. In 50 Years of Phytochemistry Research; Springer: Cham, Switzerland, 2013; Volume 43, pp. 121–155. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, J.; Rodrigues, A.; Laires, A.; Silva, F.; Costa, S.; Monteiro, M.J.; Monteiro, C.; Rueff, J. On the mechanisms of genotoxicity and metabolism of quercetin. Mutagenesis 1994, 9, 445–449. [Google Scholar] [CrossRef]
- Geiger, T.; Wehner, A.; Schaab, C.; Cox, J.; Mann, M. Comparative Proteomic Analysis of Eleven Common Cell Lines Reveals Ubiquitous but Varying Expression of Most Proteins. Mol. Cell. Proteom. 2012, 11, M111.014050. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.-P.; Sun, Y.-Y.; Chen, W.; Guo, X.; Guan, J.-K.; Li, D.-Y.; Du, G. Anti-diarrheal and anti-inflammatory activities of aqueous extract of the aerial part of Rubia cordifolia. BMC Complement. Altern. Med. 2017, 17, 20. [Google Scholar] [CrossRef] [Green Version]
- Güneş, E.; Durak, Y.; Uysal, A. Elucidation of biological properties of some commercial anthraquinones: Mutagenic/antimutagenic and antimicrobial activity approaches. J. Res. Pharm. 2019, 23, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.H.; Grunnet, N. Taurine, Glutathione and Bioenergetics. In Taurine 8; El Idrissi, A.; L’Amoreaux, W.J., Translators; Springer: New York, NY, USA, 2013; pp. 3–12. [Google Scholar]
- Holland, R.; Hawkins, A.E.; Eggler, A.L.; Mesecar, A.D.; Fabris, D.; Fishbein, J.C. Prospective Type 1 and Type 2 Disulfides of Keap1 Protein. Chem. Res. Toxicol. 2008, 21, 2051–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, Y.; Singh, J.; Raza, W.; Meena, A.; Rajak, S.; Sinha, R.A.; Luqman, S. Purpurin ameliorates alcohol-induced hepatotoxicity by reducing ROS generation and promoting Nrf2 expression. Life Sci. 2022, 309, 120964. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Kim, H.S.; Seo, Y.R. Understanding of ROS-inducing strategy in anticancer therapy. Oxid. Med. Cell. Longev. 2019, 2019, 5381692. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kwon, H.J.; Jung, H.Y.; Hahn, K.R.; Yoon, Y.S.; Hwang, I.K.; Choi, S.Y.; Kim, D.W. Neuroprotective Effects of Purpurin Against Ischemic Damage via MAPKs, Bax, and Oxidative Stress Cascades in the Gerbil Hippocampus. Mol. Neurobiol. 2022, 59, 2580–2592. [Google Scholar] [CrossRef] [PubMed]
- Lee-Hilz, Y.Y.; Boerboom, A.-M.J.F.; Westphal, A.H.; van Berkel, W.J.H.; Aarts, J.M.M.J.G.; Rietjens, I.M.C.M. Pro-Oxidant Activity of Flavonoids Induces EpRE-Mediated Gene Expression. Chem. Res. Toxicol. 2006, 19, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Meister, A.; Anderson, M.E. Glutathione. Ann. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Memar, M.Y.; Ghotaslou, R.; Samiei, M.; Adibkia, K. Antimicrobial use of reactive oxygen therapy: Current insights. Infect. Drug Resist. 2018, 11, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Hasumi, K.; Woo, J.-T.; Nagai, K.; Wachi, M. Generation of hydrogen peroxide primarily contributes to the induction of Fe (II)-dependent apoptosis in Jurkat cells by (−)-epigallocatechin gallate. Carcinogenesis 2004, 25, 1567–1574. [Google Scholar] [CrossRef] [Green Version]
- Nam, W.; Kim, S.; Nam, S.; Friedman, M. Structure-Antioxidative and Anti-Inflammatory Activity Relationships of Purpurin and Related Anthraquinones in Chemical and Cell Assays. Molecules 2017, 22, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, B.; Liu, Y.; Gao, X.; Cai, M.; Fu, J.; Yin, X.; Ni, J.; Dong, X. Isoliquiritigenin attenuates emodin-induced hepatotoxicity in vivo and in vitro through Nrf2 pathway. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 261, 109430. [Google Scholar] [CrossRef]
- Niki, E. Oxidative stress and antioxidants: Distress or eustress? Arch. Biochem. Biophys. 2016, 595, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Papadi, G.; Wesseling, S.; Troganis, A.N.; Vervoort, J.; Rietjens, I.M.C.M. Induction of EpRE-mediated gene expression by a series of mediterranean botanicals and their constituents. J. Ethnopharmacol. 2019, 240, 111940. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Qin, B.; Xu, Y.; Chen, J.; Huang, W.; Peng, Y.; Zheng, J. Chemical Reactivity of Emodin and Its Oxidative Metabolites to Thiols. Chem. Res. Toxicol. 2016, 29, 2114–2124. [Google Scholar] [CrossRef]
- Ren, Q.; Bakker, W.; de Haan, L.; Rietjens, I.M.; Bouwmeester, H. Induction of Nrf2-EpRE-mediated gene expression by hydroxyanthraquinones present in extracts from traditional Chinese medicine and herbs. Food Chem. Toxicol. 2023, 176, 113802. [Google Scholar] [CrossRef] [PubMed]
- Roesslein, M.; Hirsch, C.; Kaiser, J.-P.; Krug, H.; Wick, P. Comparability of in Vitro Tests for Bioactive Nanoparticles: A Common Assay to Detect Reactive Oxygen Species as an Example. Int. J. Mol. Sci. 2013, 14, 24320–24337. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Hussain, Y.; Luqman, S.; Meena, A. Purpurin: A natural anthraquinone with multifaceted pharmacological activities. Phytother. Res. 2021, 35, 2418–2428. [Google Scholar] [CrossRef]
- Spiegel, M.; Cel, K.; Sroka, Z. The mechanistic insights into the role of pH and solvent on antiradical and prooxidant properties of polyphenols—Nine compounds case study. Food Chem. 2023, 407, 134677. [Google Scholar] [CrossRef]
- Steele, M.L.; Fuller, S.; Patel, M.; Kersaitis, C.; Ooi, L.; Münch, G. Effect of Nrf2 activators on release of glutathione, cysteinylglycine and homocysteine by human U373 astroglial cells. Redox Biol. 2013, 1, 441–445. [Google Scholar] [CrossRef]
- Stoll, V.S.; Blanchard, J.S. Chapter 6 Buffers: Principles and practice. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1990; pp. 24–38. [Google Scholar] [CrossRef]
- Sun, Y.; Xin, X.; Zhang, K.; Cui, T.; Peng, Y.; Zheng, J. Cytochrome P450 mediated metabolic activation of chrysophanol. Chem.-Biol. Interact. 2018, 289, 57–67. [Google Scholar] [CrossRef]
- van der Linden, S.C.; von Bergh, A.R.M.; van Vught-Lussenburg, B.M.A.; Jonker, L.R.A.; Teunis, M.; Krul, C.A.M.; van der Burg, B. Development of a panel of high-throughput reporter-gene assays to detect genotoxicity and oxidative stress. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2014, 760, 23–32. [Google Scholar] [CrossRef]
- Wang, A.; Jiang, H.; Liu, Y.; Chen, J.; Zhou, X.; Zhao, C.; Chen, X.; Lin, M. Rhein induces liver cancer cells apoptosis via activating ROS-dependent JNK/Jun/caspase-3 signaling pathway. J. Cancer 2020, 11, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, Q.; Yang, Q.; Yan, X.; Feng, S.; Wang, Z. Comparison of Anthraquinones, Iridoid Glycosides and Triterpenoids in Morinda officinalis and Morinda citrifolia Using UPLC/Q-TOF-MS and Multivariate Statistical Analysis. Molecules 2019, 25, 160. [Google Scholar] [CrossRef] [Green Version]
- Wisniak, J. Pierre-jean robiquet. Educ. Quím. 2013, 24, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Mao, X.; Qin, B.; Peng, Y.; Zheng, J. In vitro and in vivo metabolic activation of rhein and characterization of glutathione conjugates derived from rhein. Chem.-Biol. Interact. 2018, 283, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Su, X.; Liang, Y.; Zhang, J.; Shi, C.; Lu, Y.; Gu, L.; Fu, L. Emodin azide methyl anthraquinone derivative triggers mitochondrial-dependent cell apoptosis involving in caspase-8-mediated Bid cleavage. Mol. Cancer Ther. 2008, 7, 1688–1697. [Google Scholar] [CrossRef] [Green Version]
- Zafarullah, M.; Li, W.Q.; Sylvester, J.; Ahmad, M. Molecular mechanisms of N-acetylcysteine actions. Cell. Mol. Life Sci. (CMLS) 2003, 60, 6–20. [Google Scholar] [CrossRef]
- Zenkov, N.K.; Menshchikova, E.B.; Tkachev, V.O. Keap1/Nrf2/ARE redox-sensitive signaling system as a pharmacological target. Biochemistry 2013, 78, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; van Dongen, K.C.W.; Bakker, W.; Miro Estruch, I.; Rietjens, I.M.C.M. The Influence of Intracellular Glutathione Levels on the Induction of Nrf2-Mediated Gene Expression by α-Dicarbonyl Precursors of Advanced Glycation End Products. Nutrients 2022, 14, 1364. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Q.; Bakker, W.; Wesseling, S.; Bouwmeester, H.; Rietjens, I.M.C.M. On the Role of ROS and Glutathione in the Mode of Action Underlying Nrf2 Activation by the Hydroxyanthraquinone Purpurin. Antioxidants 2023, 12, 1544. https://doi.org/10.3390/antiox12081544
Ren Q, Bakker W, Wesseling S, Bouwmeester H, Rietjens IMCM. On the Role of ROS and Glutathione in the Mode of Action Underlying Nrf2 Activation by the Hydroxyanthraquinone Purpurin. Antioxidants. 2023; 12(8):1544. https://doi.org/10.3390/antiox12081544
Chicago/Turabian StyleRen, Qiuhui, Wouter Bakker, Sebastiaan Wesseling, Hans Bouwmeester, and Ivonne M. C. M. Rietjens. 2023. "On the Role of ROS and Glutathione in the Mode of Action Underlying Nrf2 Activation by the Hydroxyanthraquinone Purpurin" Antioxidants 12, no. 8: 1544. https://doi.org/10.3390/antiox12081544