
What Is the Relationship between Antioxidant Efficacy, Functional Composition, and Genetic Characteristics in Comparing Soybean Resources by Year?
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation Method for Soybean Resources and Experimental Materials
2.2. Preparation of Soybean Seed Extract
2.3. Antioxidant Activity Assay
2.3.1. ABTS Radical Scavenging Activity
2.3.2. DPPH Radical Scavenging Activity
2.4. Cell Potency Assay
2.4.1. Estrogen and ER Alpha Activity
2.4.2. UCP-1 Activity
2.4.3. NO Inhibition Activity
2.5. Characterization of Isoflavone in Soybean Sources
2.5.1. Chemicals and Reagents
2.5.2. Isoflavone Extraction and Solid Phase Extraction (SPE) Process
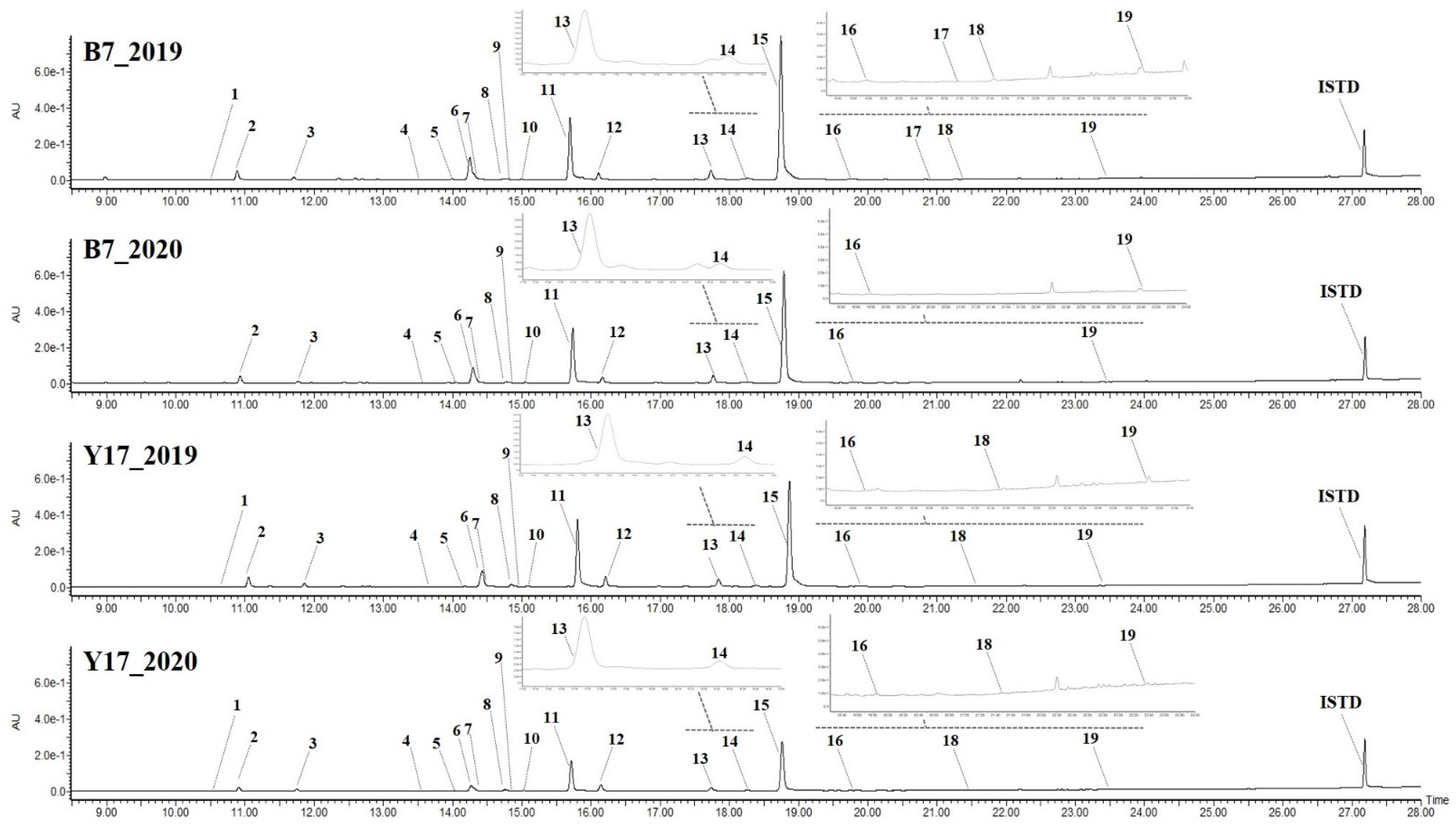
2.5.3. Isolation and Component Analysis of Individual Isoflavone Derivatives
2.6. Statistical Analysis
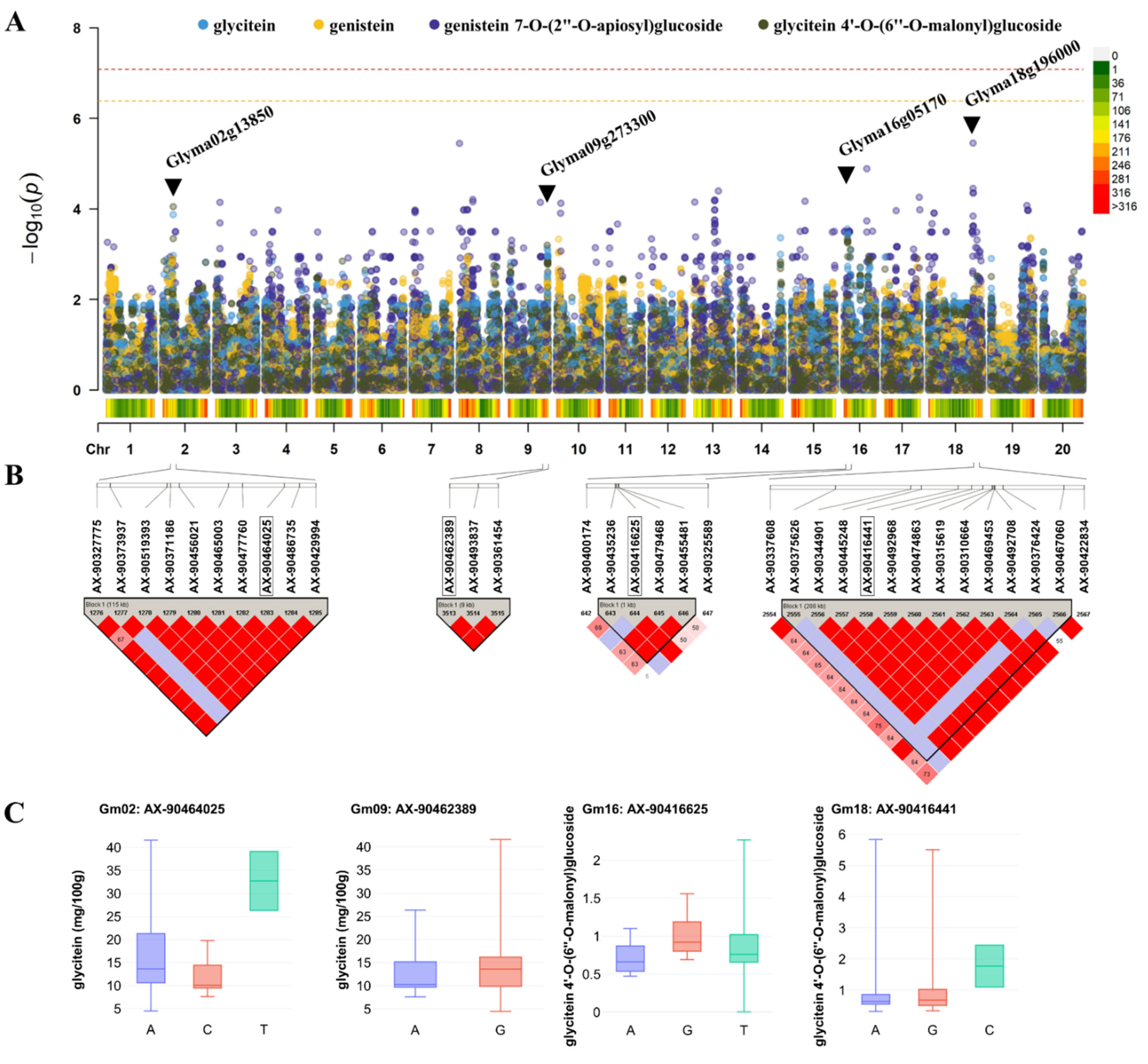
2.7. DNA Extraction, Genotyping, and Genome-Wide Association Study (GWAS)
3. Results
3.1. Physiological Activity Analysis of Soybean Seeds
3.1.1. Antioxidant Activity of Soybean Seeds (Radical Scavenging Activity)
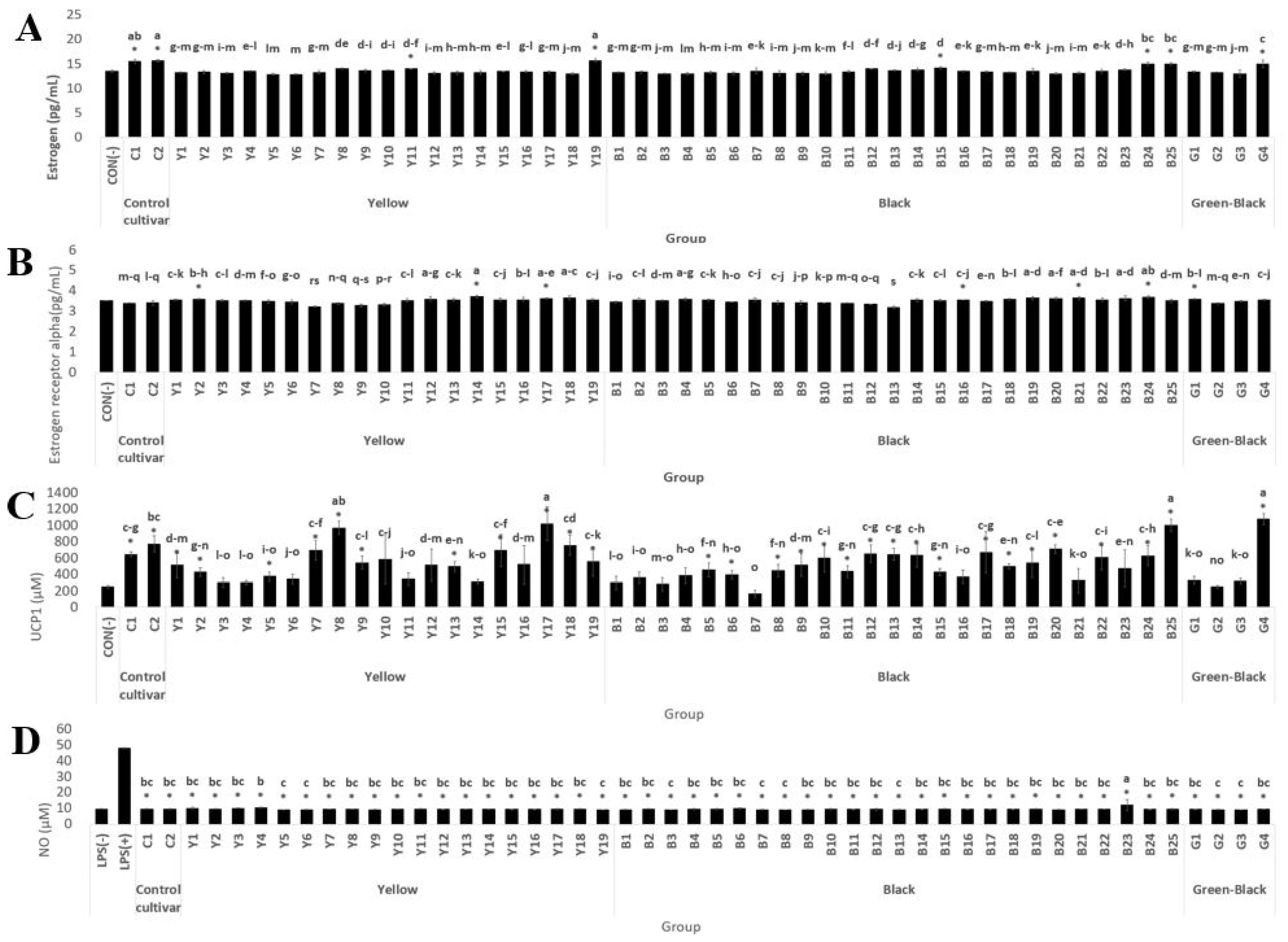
3.1.2. Estrogen and Estrogen Receptor Alpha Activity in MCF-7 Cells
3.1.3. Antiobesity Activity by Brown Fat Conversion Activity (UCP-1 Activity of C3H10T1/2 Cells)
3.1.4. NO Production Inhibition Activity of RAW 264.7 Cells
3.2. Composition and Content of Isoflavones in Soybean Seed Resources
3.3. PCA and Correlation Analysis between Efficacy and Components of Soybean Samples
3.4. Understanding the Relationship between Individual Isoflavone Derivatives and the High Efficacy of Soybean Resources and Genetic Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Messina, M. A brief historical overview of the past two decades of soy and isoflavone research. J. Nutr. 2010, 140, 1350S–1354S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.P.; Lee, J.; Gourley, L.; Duffy, S.W.; Day, N.E.; Estève, J. Dietary effects on breast-cancer risk in Singapore. Lancet 1991, 337, 1197–1200. [Google Scholar] [CrossRef]
- Isanga, J.; Zhang, G.N. Soybean bioactive components and their implications to health—A review. Food Rev. Int. 2008, 24, 252–276. [Google Scholar] [CrossRef]
- Basson, A.R.; Ahmed, S.; Almutairi, R.; Seo, B.; Cominelli, F. Regulation of intestinal inflammation by soybean and soy-derived compounds. Foods 2021, 10, 774. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Kim, C.H.; Yang, W.S. Physiologically active molecules and functional properties of soybeans in human health—A current perspective. Int. J. Mol. Sci. 2021, 22, 4054. [Google Scholar] [CrossRef]
- Jun, S.H.; Shin, W.K.; Kim, Y. Association of soybean food intake and cardiometabolic syndrome in Korean women: Korea national health and nutrition examination survey (2007 to 2011). Diabetes Metab. J. 2020, 44, 143–157. [Google Scholar] [CrossRef]
- Lichtenstein, A.H. Soy protein, isoflavones and cardiovascular disease risk. J. Nutr. 1998, 128, 1589–1592. [Google Scholar] [CrossRef] [Green Version]
- Jung, E.S.; Kim, H.R.; Hwang, Y.J.; Jang, K.A.; Seo, M.K.; Chu, H.N. Antioxidant, anti-inflammatory, anti-obesity and estrogen-like activities of soybean pod extracts. J. Korean Soc. Food Cult. 2021, 36, 649–660. [Google Scholar]
- Kim, I.S. Current perspectives on the beneficial effects of soybean isoflavones and their metabolites for humans. Antioxidants 2021, 10, 1064. [Google Scholar] [CrossRef]
- Chu, H.-N.; Lee, S.-J.; Wang, X.; Lee, S.-H.; Yoon, H.-M.; Hwang, Y.-J.; Jung, E.-S.; Kwon, Y.; Wee, C.-D.; Jang, K.-A.; et al. A correlation study on in vitro physiological activities of soybean cultivars, 19 individual isoflavone derivatives, and genetic characteristics. Antioxidants 2021, 10, 2027. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, H.W.; Lee, S.; Na, H.; Kwon, R.H.; Kim, J.H.; Yoon, H.; Choi, Y.-M.; Wee, C.-D.; Yoo, S.M.; et al. Characterization of isoflavones from seed of selected soybean (Glycine max L.) resources using high-resolution mass spectrometry. Korean J. Food Nutr. 2020, 33, 655–665. [Google Scholar]
- Hu, C.; Wong, W.T.; Wu, R.; Lai, W.F. Biochemistry and use of soybean isoflavones in functional food development. Crit. Rev. Food Sci. Nutr. 2020, 60, 2098–2112. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.J.; Anthony, M.S.; Cline, J.M.; Washburn, S.A.; Garner, S.C. Health potential of soy isoflavones for menopausal women. Public Health Nutr. 1999, 2, 489–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabbir, U.; Tyagi, A.; Ham, H.J.; Elahi, F.; Oh, D.H. Effect of fermentation on the bioactive compounds of the black soybean and their anti-Alzheimer’s activity. Front. Nutr. 2022, 9, 880361. [Google Scholar] [CrossRef]
- Day, L. Proteins from land plants–potential resources for human nutrition and food security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]
- Kulkarni, K.P.; Tayade, R.; Asekova, S.; Song, J.T.; Shannon, J.G.; Lee, J.D. Harnessing the potential of forage legumes, alfalfa, soybean, and cowpea for sustainable agriculture and global food security. Front. Plant Sci. 2018, 9, 1314. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.M.; Cary, T.R.; Nelson, R.L.; Diers, B.W. Confirmation of a seed yield QTL in soybean. Crop Sci. 2015, 55, 992–998. [Google Scholar] [CrossRef]
- Shim, S.; Kim, M.Y.; Ha, J.; Lee, Y.H.; Lee, S.H. Identification of QTLs for branching in soybean (Glycine max (L.) Merrill). Euphytica 2017, 213, 1–9. [Google Scholar] [CrossRef]
- Li, C.; Zou, J.; Jiang, H.; Yu, J.; Huang, S.; Wang, X.; Qi, Z. Identification and validation of number of pod-and seed-related traits QTLs in soybean. Plant Breed. 2018, 137, 730–745. [Google Scholar] [CrossRef]
- Li, M.W.; Wang, Z.; Jiang, B.; Kaga, A.; Wong, F.L.; Zhang, G.; Han, T.; Chung, G.; Nguyen, H.; Lam, H.M. Impacts of genomic research on soybean improvement in East Asia. Theor. Appl. Genet. 2020, 133, 1655–1678. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.J.; Park, M.R.; Yun, H.T.; Park, C.H. Analysis on variation and stability of agricultural characteristics in soybean landraces and cultivars. J. Korean Soc. Int. Agric. 2017, 29, 271–281. [Google Scholar] [CrossRef]
- Yoon, H.; Yi, J.; Desta, K.T.; Shin, M.J.; Lee, Y.; Lee, S. Yearly variation of isoflavone composition and yield-related traits of 35 Korean soybean germplasm. Korean J. Breed Sci. 2021, 53, 411–423. [Google Scholar] [CrossRef]
- Zhang, E.J.; Ng, K.M.; Luo, K.Q. Extraction and purification of isoflavones from soybeans and characterization of their estrogenic activities. J. Agric. Food Chem. 2007, 55, 6940–6950. [Google Scholar] [PubMed]
- Lee, S.H.; Jeong, Y.S.; Song, J.; Hwang, K.A.; Noh, G.M.; Hwang, I.G. Chemical composition and physiological activity of Opuntia ficus-indica depending on different cultivation regions. Korean J. Food Nutr. 2016, 29, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kwon, R.H.; Kim, J.H.; Na, H.; Lee, S.-J.; Choi, Y.-M.; Yoon, H.; Kim, S.Y.; Kim, Y.-S.; Lee, S.H.; et al. Changes in isoflavone profile from soybean seeds during Cheonggukjang fermentation based on high-resolution UPLC-DAD-QToF/MS: New succinylated and phosphorylated conjugates. Molecules 2022, 27, 4120. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Jeong, N.; Kim, J.H.; Lee, K.; Kim, K.H.; Pirani, A.; Jeong, S.C. Development, validation and genetic analysis of a large soybean SNP genotyping array. Plant J. 2015, 81, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Altshuler, D. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; Lee, D.J.; You, S. Radical scavenging activity of ethanol extracts and solvent partitioned fractions from various red seaweeds. Ocean Polar Res. 2012, 34, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.Y.; Lee, Y.J.; Song, H.S.; Hong, H.D.; Lim, J.H.; Choi, H.S.; Kang, S.-N.; Lee, O.-H. Antioxidant effects and nitrite scavenging ability of extract from Acanthopanax cortex shoot. Korean J. Food Nutr. 2012, 25, 793–799. [Google Scholar] [CrossRef]
- Noh, Y.H.; Lee, J.W.; Park, J.; Lee, S.H.; Lee, J.Y.; Kim, S.S.; Park, K.-K.; Kim, T.J.; Myung, S.-C.; Jeong, Y. Natural substance MS-10 improves women’s health via regulation of estrogen receptor. J. Korean Soc. Food Sci. Nutr. 2016, 45, 903–910. [Google Scholar]
- Porter, C. Quantification of UCP1 function in human brown adipose tissue. Adipocyte 2017, 6, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Goto, T.; Yu, R.; Uchida, K.; Tominaga, M.; Kano, Y.; Takahashi, N.; Kawada, T. Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci. Rep. 2015, 5, 18013. [Google Scholar]
- Lee, J.E.; Lee, J.Y.; Choi, J.I.; Kim, C.K.; Kim, S.J. Suppression of nitric oxide and interleukin-6 production by methanol extract of Sophorae Flos in macrophage cells. J. Korean Acad. Periodontol. 2005, 35, 9–19. [Google Scholar]
- Cross, R.K.; Wilson, K.T. Nitric oxide in inflammatory bowel disease. Inflamm. Bowel Dis. 2003, 9, 179–189. [Google Scholar] [CrossRef]
- Rural Development Administration. Study on Overall Characteristics and Health Benefits of Soybean Flavonoids. 2022. Available online: https://atis.rda.go.kr/rdais/rsrchReportInfo/rsrchReportInfoDetail.vw (accessed on 1 July 2022).
- Mitros, T.; Nelson, W.; Jackson, S.A. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar]
- Jia, M.; Ding, N.; Zhang, Q.; Xing, S.; Wei, L.; Zhao, Y.; Jia, W. A FERONIA-like receptor kinase regulates strawberry (Fragaria × ananassa) fruit ripening and quality formation. Front. Plant Sci. 2017, 8, 1099. [Google Scholar] [PubMed]
- Zhang, F.; Huang, J.; Guo, H.; Yang, C.; Li, Y.; Shen, S.; Luo, J. OsRLCK160 contributes to flavonoid accumulation and UV-B tolerance by regulating OsbZIP48 in rice. Sci. China Life Sci. 2022, 65, 1380–1394. [Google Scholar] [CrossRef]
- Egin-Bühler, B.; Ebel, J. Improved purification and further characterization of acetyl-CoA carboxylase from cultured cells of parsley (Petroselinum hortense). Eur. J. Biochem. 1983, 133, 335–339. [Google Scholar] [CrossRef]
- Kim, G.E.; Kim, M.S.; Sung, J. UVB Irradiation-induced transcriptional changes in lignin-and flavonoid biosynthesis and indole/tryptophan-auxin-responsive genes in rice seedlings. Plants 2022, 11, 1618. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ohmori, R.; Kiyose, C.; Momiyama, Y.; Ohsuzu, F.; Kondo, K. Antioxidant activities of black and yellow soybeans against low density lipoprotein oxidation. J. Agric. Food Chem. 2005, 53, 4578–4582. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-M.; Yoon, H.; Shin, M.-J.; Lee, Y.; Hur, O.S.; Lee, B.C.; Ha, B.-K.; Wang, X.; Desta, K.T. Metabolite contents and antioxidant activities of soybean (Glycine max (L.) Merrill) seeds of different seed coat colors. Antioxidants 2021, 10, 1210. [Google Scholar] [CrossRef] [PubMed]
- Eum, H.L.; Park, Y.; Yi, T.G.; Lee, J.W.; Ha, K.S.; Choi, I.Y.; Park, N.I. Effect of germination environment on the biochemical compounds and anti-inflammatory properties of soybean cultivars. PLoS ONE 2020, 15, e0232159. [Google Scholar] [CrossRef]
- Phippen, W.B.; Phippen, M.E. Soybean seed yield and quality as a response to field pennycress residue. Crop Sci. 2012, 52, 2767–2773. [Google Scholar] [CrossRef]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef]
- Dennis, T.; Li, X.; Xiao, X.; Deng, J.; Ajayo, B.S.; Long, X.; Zhang, Q.; Zhang, X.; Hu, B.; Wang, X.; et al. Spatiotemporal shading regulates anthocyanin, proanthocyanidin, and sucrose accumulation in black soybean seeds. Agron. J. 2020, 112, 708–718. [Google Scholar] [CrossRef]
- Lee, W.; Lee, S.H.; Ahn, R.S.; Park, M.J. Effect of genistein on the sexual maturation in immature female rats. Clin. Exp. Pediatr. 2009, 52, 111–118. [Google Scholar]
- Zhang, X.; Li, X.; Fang, H.; Guo, F.; Li, F.; Chen, A.; Huang, S. Flavonoids as inducers of white adipose tissue browning and thermogenesis: Signalling pathways and molecular triggers. Nutr. Metab. 2019, 16, 47. [Google Scholar] [CrossRef] [Green Version]
- Bonet, M.L.; Mercader, J.; Palou, A. A nutritional perspective on UCP1-dependent thermogenesis. Biochimie 2017, 134, 99–117. [Google Scholar] [CrossRef]
- Shin, S.; Ajuwon, K.M. Effects of diets differing in composition of 18-C fatty acids on adipose tissue thermogenic gene expression in mice fed high-fat diets. Nutrients 2018, 10, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, G.; Ablett, G.R.; Pauls, K.P.; Rajcan, I. Environmental effects on fatty acid levels in soybean seed oil. J. Am. Oil Chem. Soc. 2006, 83, 759–763. [Google Scholar] [CrossRef]
- Lee, J.D.; Woolard, M.; Sleper, D.A.; Smith, J.R.; Pantalone, V.R.; Nyinyi, C.N.; Cardinal, A.; Shannon, J.G. Environmental effects on oleic acid in soybean seed oil of plant introductions with elevated oleic concentration. Crop Sci. 2009, 49, 1762–1768. [Google Scholar] [CrossRef] [Green Version]
- Bellaloui, N.; Bruns, H.A.; Abbas, H.K.; Mengistu, A.; Fisher, D.K.; Reddy, K.N. Agricultural practices altered soybean seed protein, oil, fatty acids, sugars, and minerals in the Midsouth USA. Front. Plant Sci. 2015, 6, 31. [Google Scholar] [CrossRef]
- Jang, H.H.; Lee, Y.M.; Choe, J.S.; Kwon, O. Validation of soy isoflavone intake and its health effects: A review of the development of exposure biomarkers. Nutr. Res. Pract. 2021, 15, 1–11. [Google Scholar] [CrossRef]
- Nazari-Khanamiri, F.; Ghasemnejad-Berenji, M. Cellular and molecular mechanisms of genistein in prevention and treatment of diseases: An overview. J. Food Biochem. 2021, 45, e13972. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seed Coat Color | Code | Introduction Number | Name | Origin | Cultivar Type |
|---|---|---|---|---|---|
| Control | C1 (Yellow) | IT 212859 | Daewon | Korea | Control cultivar |
| C2 (Black) | IT 213192 | Cheongja 2 | Korea | Control cultivar | |
| Yellow | Y1 | IT 024099 | YJ208-1 | Korea | Landrace |
| Y2 | IT 104690 | Kongnamul Kong | Korea | Landrace | |
| Y3 | IT 113218 | Kongnamul Kong | Korea | Landrace | |
| Y4 | IT 153844 | KLS 87248 | Korea | Landrace | |
| Y5 | IT 155963 | Nongrim 51 | Japan | Variety | |
| Y6 | IT 171080 | PI 467319 | China | Variety | |
| Y7 | IT 195514 | Jang Kong | Korea | Landrace | |
| Y8 | IT 219581 | Myungjunamul Kong | Korea | Variety | |
| Y9 | IT 229418 | Danmi 2 | Korea | Variety | |
| Y10 | IT 229421 | Hoseo | Korea | Variety | |
| Y11 | IT 231360 | Kongnamul Kong | Korea | Landrace | |
| Y12 | IT 263155 | Sinpaldalkong 2 | Korea | Variety | |
| Y13 | IT 263167 | Uram | Korea | Variety | |
| Y14 | IT 263852 | Chungnamyeongi-1997-3 | Korea | Landrace | |
| Y15 | IT 269982 | Milyang 247 | Korea | Breeding line | |
| Y16 | IT 270002 | Jungmo 3008 | Korea | Variety | |
| Y17 | IT 274571 | GNU-2007-14613 | Korea | Landrace | |
| Y18 | IT 274592 | GNU-2007-14723 | Korea | Landrace | |
| Y19 | IT 324099 | CS 00728 | China | Breeding line | |
| Black | B1 | IT 021665 | PI 90763 | China | Landrace |
| B2 | IT 143347 | KLS86185 | Korea | Landrace | |
| B3 | IT 161904 | PI 84578 | Korea | Landrace | |
| B4 | IT 177271 | Geomjeong Kong-5 | Korea | Landrace | |
| B5 | IT 177573 | Geomjeong Kong-5 | Korea | Landrace | |
| B6 | IT 177709 | Geomjeong Kong-4 | Korea | Landrace | |
| B7 | IT 178054 | Geomjeong Kong-1 | Korea | Landrace | |
| B8 | IT 186183 | Kongnamul Kong | Korea | Landrace | |
| B9 | IT 189215 | 94yuja4 | Korea | Landrace | |
| B10 | IT 194560 | Geomjeong Kong | Korea | Landrace | |
| B11 | IT 212805 | Chungnamseocheon-1999-98 | Korea | Landrace | |
| B12 | IT 224192 | Jeonbukgunsansujip | Korea | Landrace | |
| B13 | IT 228822 | 409 | Korea | Landrace | |
| B14 | IT 231544 | Jwinuni Kong | Korea | Landrace | |
| B15 | IT 239896 | Jwineori Kong | Korea | Landrace | |
| B16 | IT 252252 | Neoljeokseoritae | Korea | Landrace | |
| B17 | IT 252748 | 294 | Korea | Landrace | |
| B18 | IT 252768 | 326 | Korea | Landrace | |
| B19 | IT 263853 | Geomen Kong | Korea | Landrace | |
| B20 | IT 274515 | GNU-2007-14502 | Korea | Landrace | |
| B21 | IT 275005 | 197 | Korea | Landrace | |
| B22 | IT 308619 | Junyeori Kong | Korea | Landrace | |
| B23 | IT 311261 | Jwinuni Kong | Korea | Landrace | |
| B24 | K 137773 | Heugseong | Korea | Variety | |
| B25 | IT 194558 | Geomjeong Kong | Korea | Landrace | |
| Green-Black | G1 | IT 154351 | KAS579-1 | Korea | Landrace |
| G2 | IT 154724 | KAS651-21 | Korea | Landrace | |
| G3 | IT 178160 | Geomjeong Kong | Korea | Landrace | |
| G4 | IT 186048 | Gangwonyanggu-1994-3709 | Korea | Landrace |
| Seed Coat Color | Effects (Unit) | Year | Mean | S.D. | p Value | ||
|---|---|---|---|---|---|---|---|
| Year | Seed Coat Color | Year and Seed Coat Color (1) | |||||
| Yellow, Black, Green- Black | ABTS (2) (mg AA eq/g) | 2019 | 4.14 | 2.00 | 0.435 | <0.001 | 0.206 |
| 2020 | 4.88 | 2.81 | |||||
| DPPH (2) (mg AA eq/g) | 2019 | 1.85 | 1.06 | 0.069 | <0.001 | 0.038 | |
| 2020 | 2.77 | 1.70 | |||||
| Estrogen (2) (%, vs. Control) | 2019 | 99.96 | 2.75 | 0.998 | 0.980 | 0.776 | |
| 2020 | 99.59 | 4.47 | |||||
| ER (2) (%, vs. Control) | 2019 | 118.88 | 12.05 | <0.001 | 0.339 | 0.336 | |
| 2020 | 99.91 | 3.29 | |||||
| UCP-1 (2) (%, vs. Control) | 2019 | 273.40 | 91.25 | 0.002 | 0.821 | 0.346 | |
| 2020 | 206.14 | 82.3 | |||||
| NO (2) (%, vs. Control) | 2019 | 12.94 | 14.87 | <0.001 | 0.552 | 0.473 | |
| 2020 | 80.44 | 0.96 | |||||
| Seed Coat Color | Code | ABTS (mg AA eq/g) | DPPH (mg AA eq/g) |
|---|---|---|---|
| Control cultivar | C1 | 4.45 ± 0.24 l,m | 2.68 ± 0.21 l–n |
| C2 | 5.91 ± 0.41 h | 3.77 ± 0.16 gh | |
| Range | 4.45–5.91 | 2.68–3.77 | |
| Mean | 5.18 | 3.23 | |
| Yellow | Y1 | 3.86 ± 0.23 n–q | 2.48 ± 0.41 m–p |
| Y2 | 2.08 ± 0.13 z | 1.35 ± 0.15 v–x | |
| Y3 | 2.26 ± 0.13 yz | 1.04 ± 0.10 x | |
| Y4 | 3.93 ± 0.20 n–p | 1.99 ± 0.19 q–t | |
| Y5 | 3.22 ± 0.11 s–w | 1.74 ± 0.03 s–v | |
| Y6 | 3.22 ± 0.08 s–w | 1.64 ± 0.14 t–w | |
| Y7 | 3.78 ± 0.14 n–r | 1.83 ± 0.12 r–v | |
| Y8 | 3.39 ± 0.13 q–u | 1.83 ± 0.11 r–v | |
| Y9 | 3.74 ± 0.17 n–r | 2.06 ± 0.12 p–t | |
| Y10 | 3.88 ± 0.17 n–q | 2.38 ± 0.13 m–q | |
| Y11 | 3.47 ± 0.13 p–u | 1.45 ± 0.13 u–x | |
| Y12 | 4.13 ± 0.03 mn | 1.90 ± 0.19 r–u | |
| Y13 | 3.97 ± 0.35 no | 1.89 ± 0.17 r–u | |
| Y14 | 1.98 ± 0.04 z | 1.02 ± 0.10 x | |
| Y15 | 1.93 ± 0.19 z | 1.45 ± 0.02 u–x | |
| Y16 | 5.49 ± 0.39 hi | 3.00 ± 0.10 j–l | |
| Y17 | 3.65 ± 0.17 n–s | 2.38 ± 0.12 m–q | |
| Y18 | 3.05 ± 0.10 u–x | 2.22 ± 0.06 n–r | |
| Y19 | 3.08 ± 0.08 t–w | 1.93 ± 0.19 q–u | |
| Range | 1.93–5.91 | 1.02–3.77 | |
| Mean | 3.37 | 1.87 | |
| Black | B1 | 10.40 ± 0.43 d | 6.31 ± 0.27 c |
| B2 | 2.81 ± 0.17 wx | 1.36 ± 0.09 v–x | |
| B3 | 3.53 ± 0.08 o–u | 1.69 ± 0.13 s–w | |
| B4 | 4.69 ± 0.12 j–l | 2.48 ± 0.38 m–p | |
| B5 | 4.76 ± 0.21 j–l | 2.66 ± 0.39 l–n | |
| B6 | 15.89 ± 0.44 a | 9.23 ± 0.49 a | |
| B7 | 3.44 ± 0.23 p–u | 1.94 ± 0.25 q–t | |
| B8 | 12.60 ± 0.50 b | 7.41 ± 0.26 b | |
| B9 | 5.01 ± 0.31 i–k | 2.77 ± 0.56 l,m | |
| B10 | 5.54 ± 0.13 h | 3.54 ± 0.40 hi | |
| B11 | 3.70 ± 0.19 n–s | 1.79 ± 0.08 r–v | |
| B12 | 3.30 ± 0.16 r–v | 1.61 ± 0.04 t–w | |
| B13 | 5.10 ± 0.09 ij | 2.83 ± 0.05 k–m | |
| B14 | 7.59 ± 0.20 f | 4.10 ± 0.18 fg | |
| B15 | 8.72 ± 0.34 e | 4.97 ± 0.21 d | |
| B16 | 3.44 ± 0.24 p–u | 1.81 ± 0.28 r–v | |
| B17 | 10.93 ± 0.40 c | 6.27 ± 0.30 c | |
| B18 | 4.57 ± 0.20 k–m | 2.70 ± 0.22 l,m | |
| B19 | 6.55 ± 0.12 g | 3.80 ± 0.15 gh | |
| B20 | 4.13 ± 0.18 mn | 2.57 ± 0.13 l–o | |
| B21 | 5.65 ± 0.26 h | 3.37 ± 0.54 ij | |
| B22 | 5.77 ± 0.10 h | 3.44 ± 0.28 g–i | |
| B23 | 7.96 ± 0.55 f | 4.59 ± 0.31 de | |
| B24 | 4.92 ± 0.59 i–k | 3.22 ± 0.32 i–k | |
| B25 | 6.64 ± 0.33 g | 4.39 ± 0.32 ef | |
| Range | 2.81–15.89 | 1.36–9.23 | |
| Mean | 6.31 | 3.63 | |
| Green-Black | G1 | 3.67 ± 0.18 n–s | 2.17 ± 0.13 o–s |
| G2 | 2.88 ± 0.22 v–x | 1.24 ± 0.04 wx | |
| G3 | 2.61 ± 0.16 xy | 1.40 ± 0.35 v–x | |
| G4 | 3.55 ± 0.20 o–t | 1.90 ± 0.29 r–u | |
| Range | 2.61–3.67 | 1.24–2.17 | |
| Mean | 3.17 | 1.68 |
| Seed Coat Color | Components | Year | Mean (mg/100 g) | S.D. (mg/100 g) | p Value | ||
|---|---|---|---|---|---|---|---|
| Year | Seed Coat Color | Year and Seed Coat Color (1) | |||||
| Yellow, Black, Green- Black | Genistein 5-O-glucoside (F1) | 2019 | 0.36 | 0.35 | 0.291 | 0.379 | 0.978 |
| 2020 | 0.27 | 0.28 | |||||
| Daidzein 7-O-glucoside (daidzin) (F2) | 2019 | 12.79 | 6.35 | 0.003 | 0.637 | 0.259 | |
| 2020 | 9.32 | 4.47 | |||||
| Glycitein 7-O-glucoside (glycitin) (F3) | 2019 | 4.67 | 2.84 | 0.772 | 0.166 | 0.362 | |
| 2020 | 4.47 | 2.57 | |||||
| Genistein 7-O-(6″-O-apiosyl)glucoside (ambosin) (F4) | 2019 | 0.53 | 0.28 | 0.518 | 0.451 | 0.239 | |
| 2020 | 0.51 | 0.31 | |||||
| Genistein 7-O-(2″-O-apiosyl)glucoside (F5) | 2019 | 1.40 | 1.50 | 0.363 | 0.065 | 0.974 | |
| 2020 | 1.05 | 1.18 | |||||
| Genistein 7-O-glucoside (genistin) (F6) | 2019 | 19.75 | 8.40 | <0.001 | <0.001 | 0.237 | |
| 2020 | 14.19 | 7.20 | |||||
| Daidzein 4′-O-(6″-O-malonyl)glucoside (F7) | 2019 | 6.66 | 2.97 | 0.026 | 0.723 | 0.360 | |
| 2020 | 5.39 | 2.31 | |||||
| Genistein 5-O-(6″-O-malonyl)glucoside (F8) | 2019 | 1.76 | 1.72 | 0.994 | 0.219 | 0.885 | |
| 2020 | 1.91 | 1.81 | |||||
| Daidzein 7-O-(4″-O-malonyl)glucoside (4″-O-malonyldaidzin) (F9) | 2019 | 1.11 | 0.62 | <0.001 | 0.321 | 0.352 | |
| 2020 | 0.64 | 0.29 | |||||
| Glycitein 4′-O-(6″-O-malonyl)glucoside (F10) | 2019 | 0.86 | 0.46 | 0.828 | 0.257 | 0.346 | |
| 2020 | 0.85 | 0.40 | |||||
| Daidzein 7-O-(6″-O-malonyl)glucoside (6″-O-malonyldaidzin) (F11) | 2019 | 75.39 | 32.16 | 0.069 | 0.873 | 0.325 | |
| 2020 | 66.94 | 26.89 | |||||
| Glycitein 7-O-(6″-O-malonyl)glucoside (6″-O-malonylglycitin) (F12) | 2019 | 10.18 | 5.98 | 0.457 | 0.229 | 0.527 | |
| 2020 | 9.25 | 5.20 | |||||
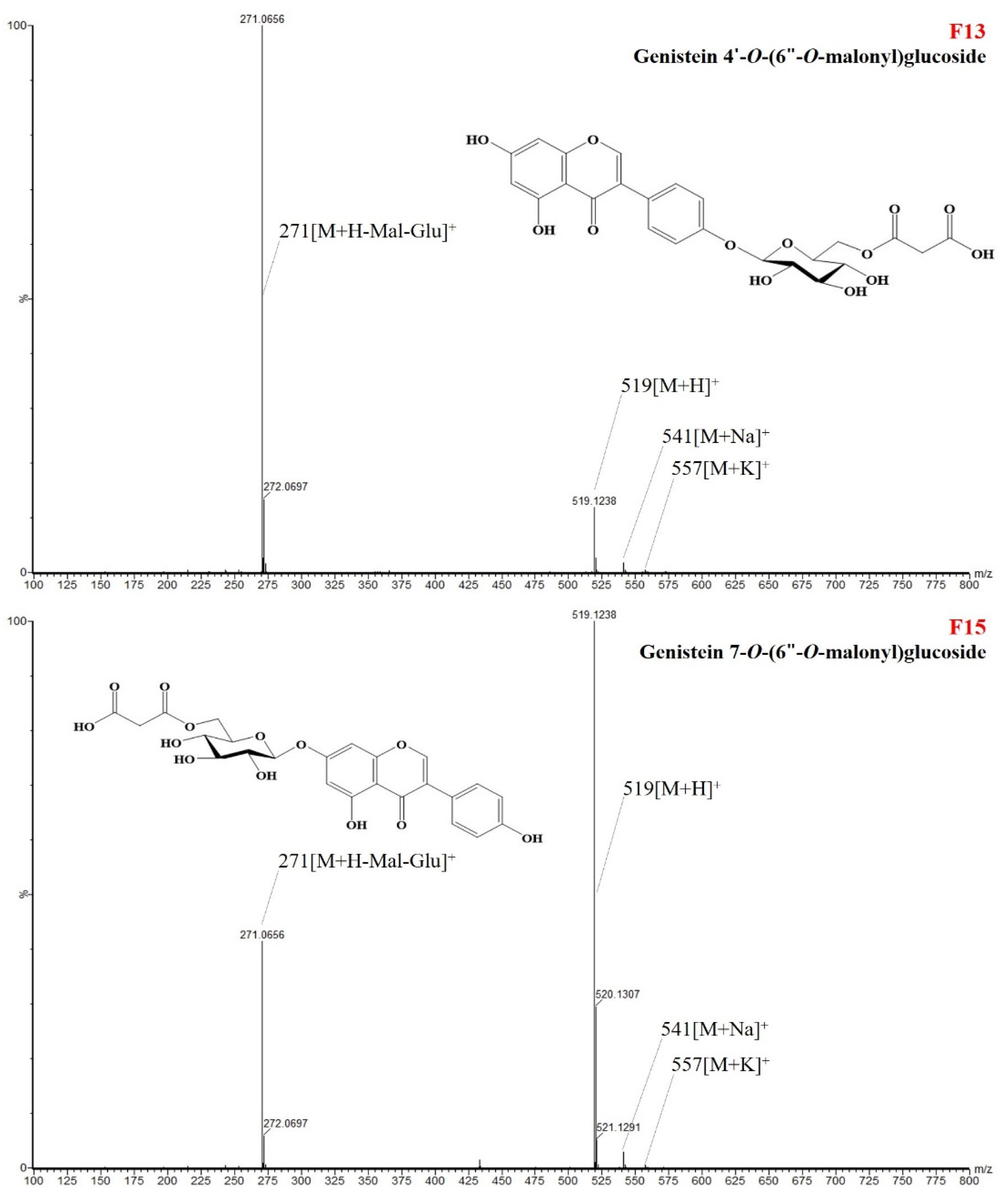
| Genistein 4′-O-(6″-O-malonyl)glucoside (F13) | 2019 | 9.78 | 3.14 | <0.001 | 0.008 | 0.474 | |
| 2020 | 6.99 | 3.14 | |||||
| Genistein 7-O-(4″-O-malonyl)glucoside (4″-O-malonylgenistin) (F14) | 2019 | 1.84 | 0.67 | <0.001 | 0.955 | 0.274 | |
| 2020 | 1.10 | 0.40 | |||||
| Genistein 7-O-(6″-O-malonyl)glucoside (6″-O-malonylgenistin) (F15) | 2019 | 134.71 | 41.78 | 0.019 | 0.004 | 0.505 | |
| 2020 | 116.05 | 43.28 | |||||
| Daidzein (F16) | 2019 | 0.77 | 0.50 | <0.001 | 0.425 | 0.904 | |
| 2020 | 0.31 | 0.21 | |||||
| Glycitein (F17) | 2019 | 0.16 | 0.24 | 0.288 | 0.314 | 0.752 | |
| 2020 | 0.07 | 0.22 | |||||
| Genistein 7-O-(6″-O-acetyl)glucoside (6″-O-acetylgenistin) (F18) | 2019 | 0.29 | 0.51 | 0.748 | 0.319 | 0.809 | |
| 2020 | 0.23 | 0.67 | |||||
| Genistein (F19) | 2019 | 0.76 | 0.37 | 0.185 | 0.064 | 0.113 | |
| 2020 | 0.55 | 0.29 | |||||
| Daidzein group | 2019 | 96.73 | 41.91 | 0.036 | 0.928 | 0.316 | |
| 2020 | 82.59 | 33.57 | |||||
| Genistein group | 2019 | 171.17 | 53.57 | 0.004 | 0.003 | 0.393 | |
| 2020 | 142.85 | 53.73 | |||||
| Glycitein group | 2019 | 15.88 | 9.28 | 0.544 | 0.224 | 0.457 | |
| 2020 | 14.63 | 8.13 | |||||
| Total (F20) | 2019 | 283.78 | 89.76 | 0.010 | 0.156 | 0.336 | |
| 2020 | 240.07 | 85.78 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, H.-N.; Lee, S.; Wang, X.; Wee, C.-D.; Yoon, H.-M.; Jung, E.-S.; Seo, M.-K.; Kwon, Y.; Jang, K.-A.; Kim, H.-R. What Is the Relationship between Antioxidant Efficacy, Functional Composition, and Genetic Characteristics in Comparing Soybean Resources by Year? Antioxidants 2022, 11, 2249. https://doi.org/10.3390/antiox11112249
Chu H-N, Lee S, Wang X, Wee C-D, Yoon H-M, Jung E-S, Seo M-K, Kwon Y, Jang K-A, Kim H-R. What Is the Relationship between Antioxidant Efficacy, Functional Composition, and Genetic Characteristics in Comparing Soybean Resources by Year? Antioxidants. 2022; 11(11):2249. https://doi.org/10.3390/antiox11112249
Chicago/Turabian StyleChu, Han-Na, Suji Lee, Xiaohan Wang, Chi-Do Wee, Hye-Myeong Yoon, Eun-Suk Jung, Mi-Kyung Seo, Yongseok Kwon, Kyeong-A Jang, and Haeng-Ran Kim. 2022. "What Is the Relationship between Antioxidant Efficacy, Functional Composition, and Genetic Characteristics in Comparing Soybean Resources by Year?" Antioxidants 11, no. 11: 2249. https://doi.org/10.3390/antiox11112249