
Cysteamine Supplementation In Vitro Remarkably Promoted Rumen Fermentation Efficiency towards Propionate Production via Prevotella Enrichment and Enhancing Antioxidant Capacity
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of CS Products
2.2. In Vitro Batch Cultures
2.3. Determination of In Vitro Dry Matter Digestibility, Volatile Fatty Acids, and Antioxidant Content
2.4. DNA Extraction and Sequencing
2.5. Chemical Analyses
2.6. Calculations
2.7. Statistical Analysis
3. Results
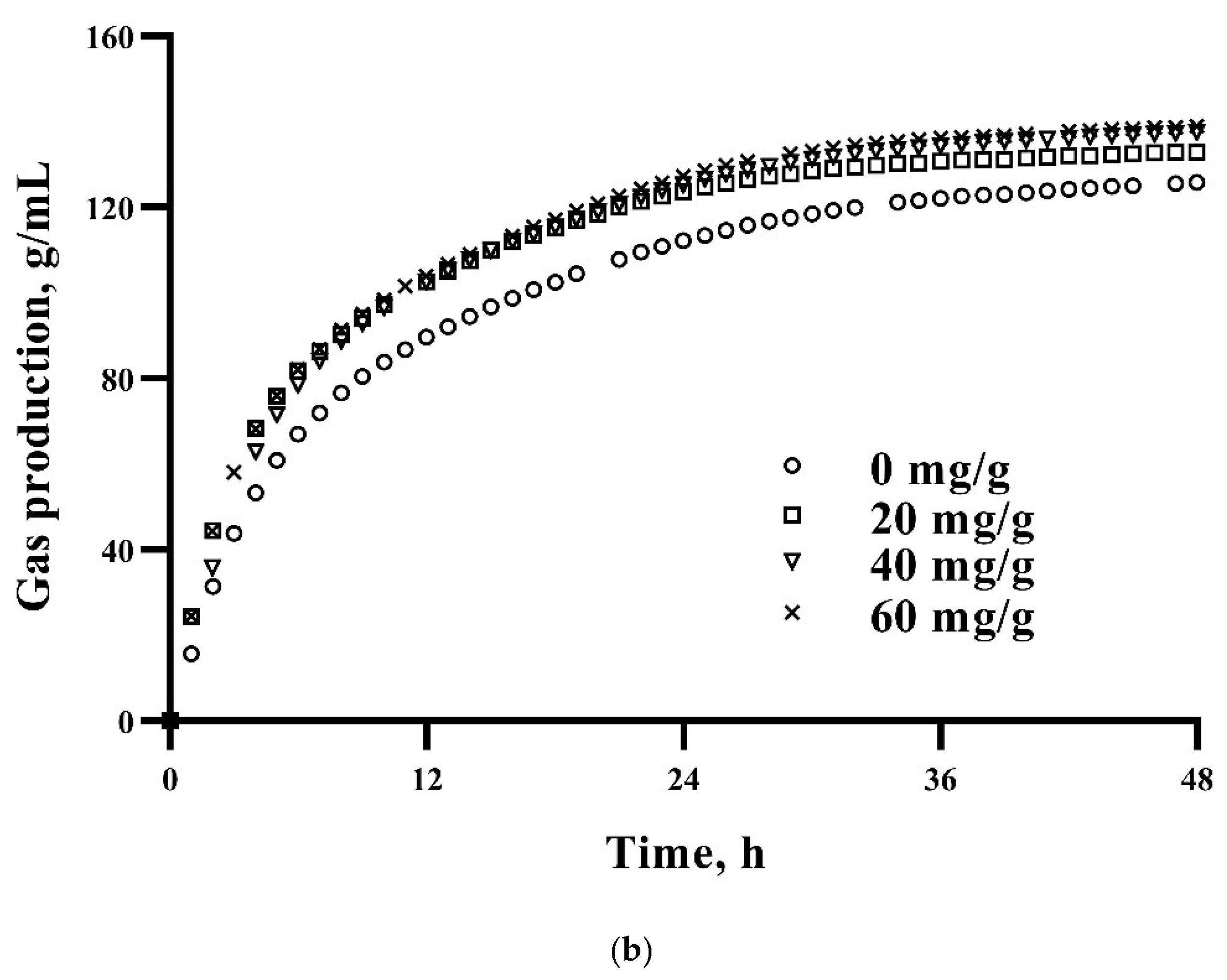
3.1. In Vitro Dry Matter Disappearance at 48 h and Kinetic Gas Production
3.2. Rumen Fermentation
3.3. Rumen Antioxidant Capacity
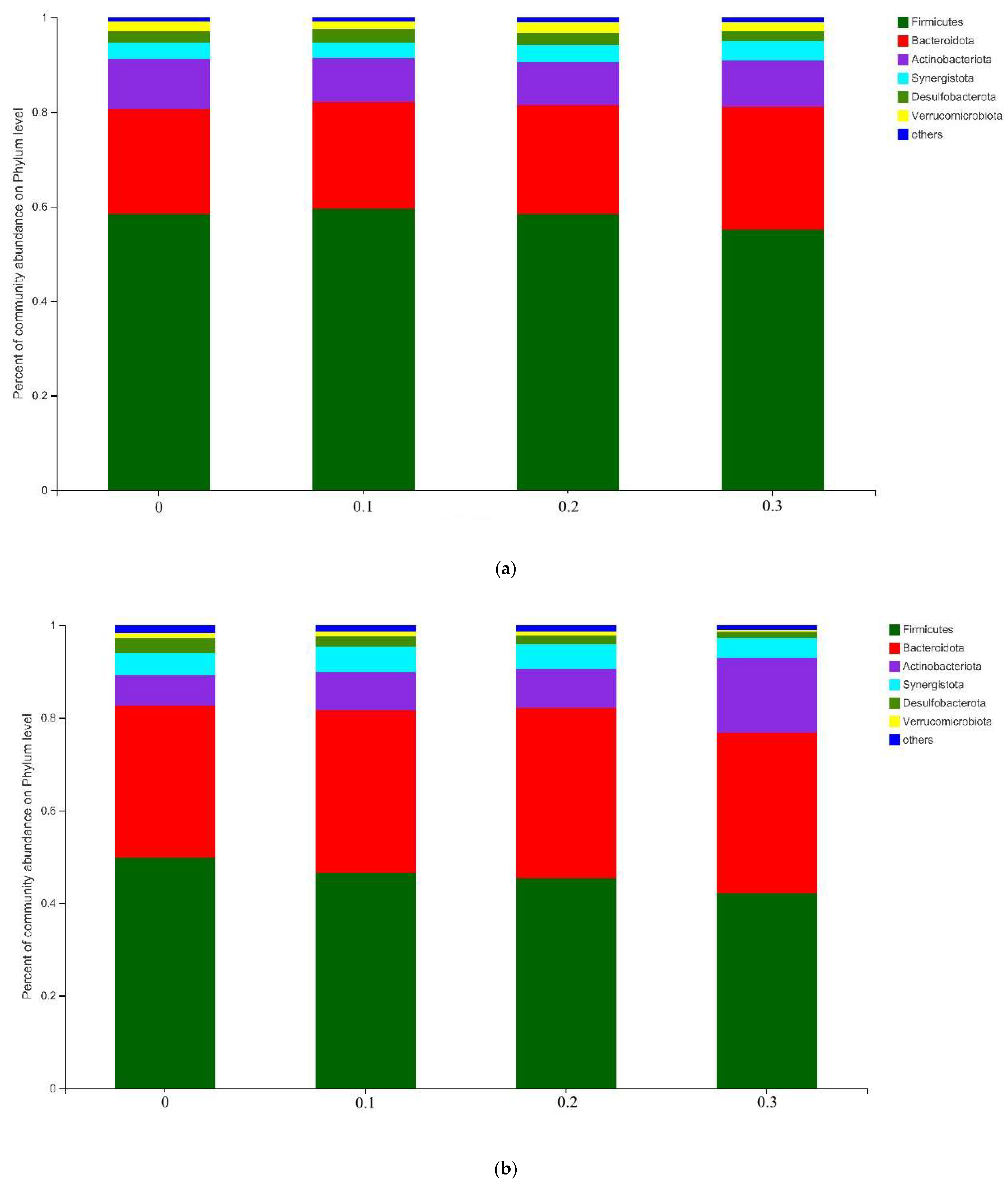
3.4. Rumen Bacteria Community
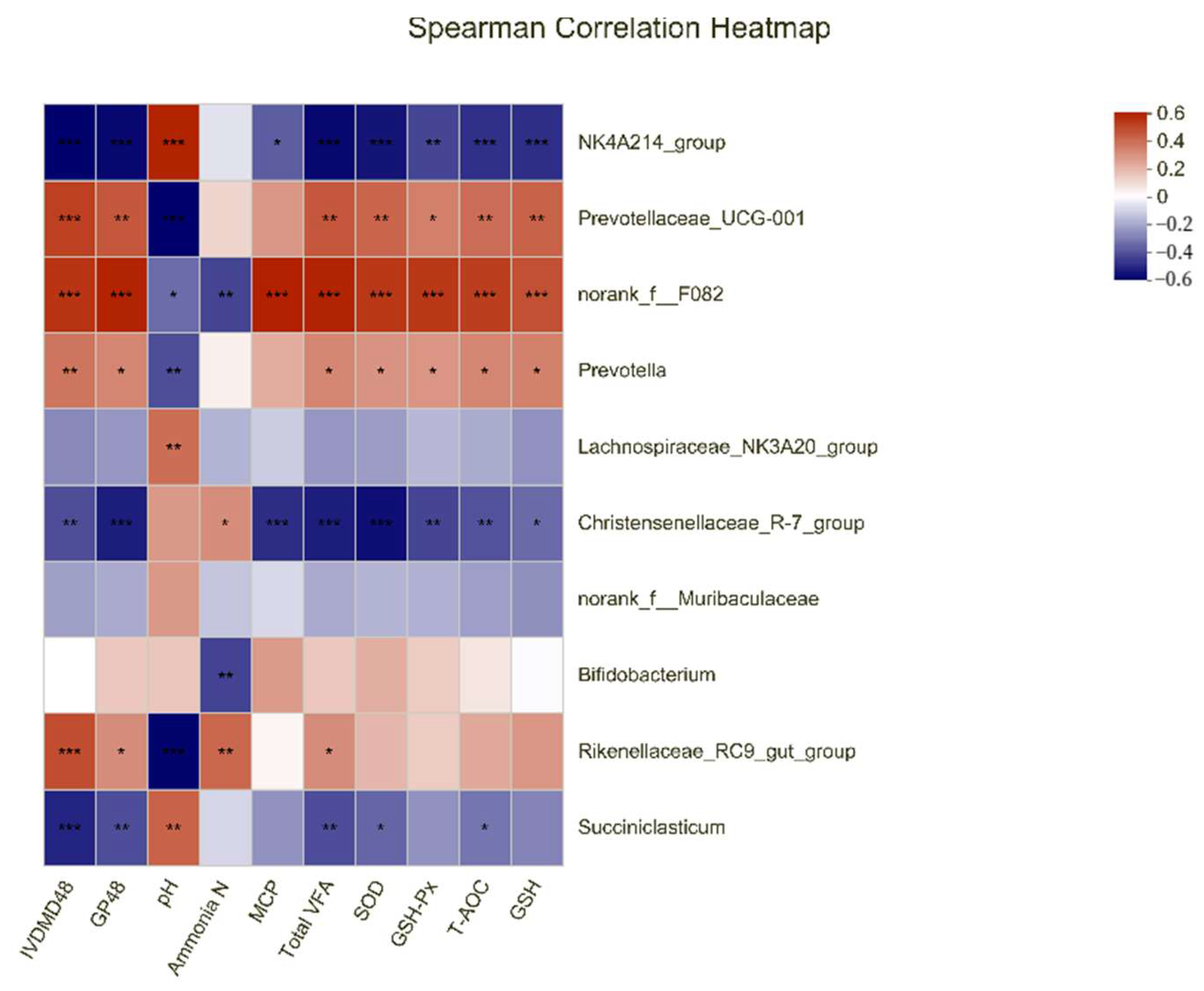
3.5. Correlations among the Top 10 Bacterial Genera and the Parameters of Gas Production Kinetic Parameters, Rumen Fermentation, and Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mamuad, L.L.; Lee, S.S.; Lee, S.S. Recent insight and future techniques to enhance rumen fermentation in dairy goats. Asian-Australas. J. Anim. Sci. 2019, 32, 1321–1330. [Google Scholar] [CrossRef]
- Robinson, C.J.; Bohannan, B.J.; Young, V.B. From structure to function: The ecology of host-associated microbial communities. Microbiol. Mol. Biol. Rev. 2010, 74, 453–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleason, C.B.; White, R.R. Variation in animal performance explained by the rumen microbiome or by diet composition. J. Anim. Sci. 2018, 96, 4658–4673. [Google Scholar] [CrossRef]
- Fiore, E.; Gianesella, M.; Arfuso, F.; Giudice, E.; Piccione, G.; Lora, M.; Stefani, A.; Morgante, M. Glucose infusion response on some metabolic parameters in dairy cows during transition period. Arch. Anim. Breed. 2014, 57, 3. [Google Scholar] [CrossRef] [Green Version]
- Fiore, E.; Arfuso, F.; Gianesella, M.; Vecchio, D.; Morgante, M.; Mazzotta, E.; Badon, T.; Rossi, P.; Bedin, S.; Piccione, G. Metabolic and hormonal adaptation in Bubalus bubalis around calving and early lactation. PLoS ONE 2018, 13, e0193803. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, W.; Yu, H.; Xu, L.; Qu, M.; Li, Y. Improved antioxidant activity and rumen fermentation in beef cattle under heat stress by dietary supplementation with creatine pyruvate. Anim. Sci. J. 2020, 91, e13486. [Google Scholar] [CrossRef]
- Abbate, J.M.; Macrì, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of Protein Hydrolysates from Anchovy (Engraulis Encrasicolus) Waste for Twelve Weeks Decreases Metabolic Dysfunction-Associated Fatty Liver Disease Severity in ApoE–/–Mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef]
- Armato, L.; Gianesella, M.; Morgante, M.; Fiore, E.; Rizzo, M.; Giudice, E.; Piccione, G. Rumen volatile fatty acids × dietary supplementation with live yeast and yeast cell wall in feedlot beef cattle. Acta Agr. Scand. A-Anim. Sci. 2016, 66, 119–124. [Google Scholar] [CrossRef]
- Monteverde, V.; Congiu, F.; Vazzana, I.; Dara, S.; Di Pietro, S.; Piccione, G. Serum lipid profile modification related to polyunsaturated fatty acid supplementation in thoroughbred horses. J. Appl. Anim. Res. 2017, 45, 615–618. [Google Scholar] [CrossRef]
- Benov, L. How Superoxide Radical Damages the Cell. Protoplasma 2001, 217, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y. Oxidant-Mediated Protein Amino Acid Conversion. Antioxidants 2019, 82, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, I.; Tomassoni, D.; Roy, P.; Di Cesare Mannelli, L.; Amenta, F.; Tayebati, S.K. Antioxidant Properties of Alpha-Lipoic (Thioctic) Acid Treatment on Renal and Heart Parenchyma in a Rat Model of Hypertension. Antioxidants 2021, 107, 1006. [Google Scholar] [CrossRef] [PubMed]
- Ayemele, A.G.; Tilahun, M.; Lingling, S.; Elsaadawy, S.A.; Guo, Z.; Zhao, G.; Xu, J.; Bu, D. Oxidative Stress in Dairy Cows: Insights into the Mechanistic Mode of Actions and Mitigating Strategies. Antioxidants 2021, 10, 1918. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.; Reichlin, S. Somatostatin in rat tissues is depleted by cysteamine administration. Endocrinology 1981, 109, 2255–2257. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.M.; Landry, D.; Millard, W.J.; Badger, T.M.; Arnold, M.A.; Martin, J.B. Depletion of somatostatin-like immunoreactivity in the rat central nervous system by cysteamine. J. Neurosci. 1982, 2, 225–231. [Google Scholar] [CrossRef]
- Hu, R.; Wang, Z.; Peng, Q.; Zou, H.; Wang, H.; Yu, X.; Jing, X.P.; Wang, Y.X.; Cao, B.H.; Bao, S.K.; et al. Effects of GHRP-2 and Cysteamine Administration on Growth Performance, Somatotropic Axis Hormone and Muscle Protein Deposition in Yaks (Bos grunniens) with Growth Retardation. PLoS ONE 2016, 11, e0149461. [Google Scholar] [CrossRef]
- Wu, Q.C.; Wang, W.K.; Zhang, F.; Li, W.J.; Wang, Y.L.; Lv, L.K.; Yang, H.J. Dietary Cysteamine Supplementation Remarkably Increased Feed Efficiency and Shifted Rumen Fermentation toward Glucogenic Propionate Production via Enrichment of Prevotella in Feedlot Lambs. Microorganisms 2022, 10, 1105. [Google Scholar] [CrossRef]
- Wang, C.; Dong, C.J.; Wang, Z.Q.; Yang, F.; Mao, H.L.; Wu, Z.; Zhou, Q.; Wang, H.F. Effect of cysteamine hydrochloride supplementation on the milk performance of dairy cow. Livest. Sci. 2015, 178, 94–99. [Google Scholar] [CrossRef]
- Issels, R.D.; Nagele, A.; Eckert, K.G.; Wilmanns, W. Promotion of cystine uptake and its utilization for glutathione biosynthesis induced by cysteamine and N-acetyl-cysteine. Biochem. Pharmacol. 1988, 37, 881–888. [Google Scholar] [CrossRef]
- Oyamada, T.; Fukui, Y. Oxygen tension and medium supplements for in vitro maturation of bovine oocytes cultured individually in a chemically defined medium. J. Reprod. Dev. 2004, 50, 107–117. [Google Scholar] [CrossRef]
- Sarıözkan, S.; Tuncer, P.B.; Büyükleblebici, S.; Bucak, M.N.; Cantürk, F.; Eken, A. Antioxidative effects of cysteamine, hyaluronan and fetuin on post-thaw semen quality, DNA integrity and oxidative stress parameters in the Brown Swiss bull. Andrologia 2015, 47, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M.; Williams, W.L. New, rapid, sensitive method for protein determination. Fed. Proc. 1976, 35, 274–284. [Google Scholar]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 19th ed.; Association of Official Analytical Chemists: Rockville, MD, USA, 2012. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- France, J.; Dijkstra, J.; Dhanoa, M.S.; Lopez, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: Derivation of models and other mathematical considerations. Br. J. Nutr. 2000, 83, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wu, Q.; Li, W.; Wang, Y.; Zhang, F.; Lv, L.; Li, S.; Yang, H. High-Gossypol Whole Cottonseed Exhibited Mediocre Rumen Degradability and Less Microbial Fermentation Efficiency than Cottonseed Hull and Cottonseed Meal with an In Vitro Gas Production Technique. Fermentation 2022, 8, 103. [Google Scholar] [CrossRef]
- Makkar, H.P. In Vitro Screening of Feed Resources for Efficiency of Microbial Protein Synthesis. In Vitro Screening of Plant Resources for Extra-Nutritional Attributes in Ruminants: Nuclear and Related Methodologies; Vercoe, P., Makkar, H., Schlink, A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 107–144. [Google Scholar] [CrossRef]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.P.; Fleury, M.J.J. Microbial antioxidant defense enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Janssen, P. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar]
- Kumar, S.; Dagar, S.S.; Sirohi, S.K.; Upadhyay, R.C.; Puniya, A.K. Microbial profiles, in vitro gas production and dry matter digestibility based on various ratios of roughage to concentrate. Ann. Microbiol. 2013, 63, 541–545. [Google Scholar] [CrossRef]
- Zhang, F.; Wu, Q.; Wang, W.; Guo, S.; Li, W.; Lv, L.; Chen, H.; Xiong, F.; Liu, Y.; Chen, Y.; et al. Inhibitory Effect Mediated by Deoxynivalenol on Rumen Fermentation under High-Forage Substrate. Fermentation 2022, 8, 369. [Google Scholar] [CrossRef]
- Wang, W.K.; Wang, Y.L.; Li, W.J.; Wu, Q.C.; Li, S.L.; Yang, H.J. Gossypol Exhibited Higher Detrimental Effect on Ruminal Fermentation Characteristics of Low-Forage in Comparison with High-Forage Mixed Feeds. Toxics 2021, 8, 51. [Google Scholar] [CrossRef]
- McCann, J.C.; Wiley, L.M.; Forbes, T.D.; Rouquette, F.M.; Tedeschi, L.O.; Zilberstein, D. Relationship between the rumen microbiome and residual feed intake-efficiency of brahman bulls stocked on bermudagrass pastures. PLoS ONE 2014, 9, e091864. [Google Scholar] [CrossRef] [Green Version]
- Heath, T. Dukes’ Physiology of Domestic Animals. Aust. Vet. J. 2010, 71, 187. [Google Scholar] [CrossRef]
- Wang, Q.J.; Mao, S.Y.; Zhang, H.X.; Zhu, W.Y. Effects of Cysteamine on in vitro Fermentation by Rumen Microbes from Goats. J. Huazhong Agric. Univ. 2002, 21, 535–539. [Google Scholar]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Song, T.Z.; Feng, J.; Xia, C.Y.; Yuhong, B.; Ke, F.; Guoqing, S. Effects of Cysteamine on pH, Total Dehydrogenase and TVFA of Rumen in Weaned Lambs of Tibetan Sheep. Anim. Feed. Sci. Technol. 2014, 35, 4–5. [Google Scholar]
- van Houter, M.F.J. The production and metabolism of volatile fatty acids by ruminants fed roughages: A review. Nature 1993, 43, 189–225. [Google Scholar] [CrossRef]
- Hu, R.; Zou, H.; Wang, Z.; Cao, B.; Peng, Q.; Jing, X.; Wang, Y.; Shao, Y.; Pei, Z.; Zhang, X.; et al. Nutritional Interventions Improved Rumen Functions and Promoted Compensatory Growth of Growth-Retarded Yaks as Revealed by Integrated Transcripts and Microbiome Analyses. Front. Microbiol. 2019, 21, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezerra, H.V.A.; Buarque, V.L.M.; Silva, L.S.B.; Leme, P.R.P.; Vidal, A.M.C.; Vaz, A.C.N.; Gallo, S.B.; Silva, S.L.; Leme, P.R. Effect of Castor and Cashew Nut Shell Oils, Selenium and Vitamin E as Antioxidants on the Health and Meat Stability of Lambs Fed a High-Concentrate Diet. Antioxidants 2020, 9, 1298. [Google Scholar] [CrossRef]
- Wang, S.; Wang, F.; Kong, F.; Cao, Z.; Wang, W.; Yang, H.; Wang, Y.; Bi, Y.; Li, S. Effect of Supplementing Different Levels of L-Glutamine on Holstein Calves during Weaning. Antioxidants 2022, 11, 542. [Google Scholar] [CrossRef]
- Kong, F.; Wang, S.; Dai, D.; Cao, Z.; Wang, Y.; Li, S.; Wang, W. Preliminary Investigation of the Effects of Rosemary Extract Supplementation on Milk Production and Rumen Fermentation in High-Producing Dairy Cows. Antioxidants 2022, 11, 1715. [Google Scholar] [CrossRef]
- Shokrollahi, B.; Fazli, A.; Morammazi, S.; Saadati, N.; Ahmad, H.I.; Hassan, F.U. Cysteamine administration in lambs grazing on mountain pastures: Effects on the body weight, antioxidant capacity, thyroid hormones and growth hormone secretion. Vet. Med. Sci. 2022, 8, 328–335. [Google Scholar] [CrossRef]
- Fink, R.C.; Scandalios, J.G. Molecular evolution and structure—Function relationships of the superoxide dismutase gene families in angiosperms and their relationship to other eukaryotic and prokaryotic superoxide dismutases. Arch. Biochem. Biophys. 2002, 399, 19–36. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, Q.; Zhou, Y.; Ahmad, H.; Wang, T. Effects of clinoptilolite on growth performance and antioxidant status in broilers. Biol. Trace Elem. Res. 2013, 155, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Bai, M.; Xu, K.; Shao, Y.; Yang, Z.; Xiong, X.; Huang, R.; Li, Y.; Liu, H. Effects of Coated Cysteamine on Oxidative Stress and Inflammation in Weaned Pigs. Animals 2021, 11, 2217. [Google Scholar] [CrossRef]
- Haenen, G.R.; Vermeulen, N.P.; Timmerman, H.; Bast, A. Effect of thiols on lipid peroxidation in rat liver microsomes. Chem.-Biol. Interact. 1989, 71, 201–212. [Google Scholar] [CrossRef]
- Ishii, T.; Hishimura, I.; Bannai, S.; Sugita, Y. Mechanism of growth promotion of L 1210 by 2-mercaptoethanol in vitro. J. Biol Chem. 1981, 256, 12387–12392. [Google Scholar] [CrossRef]
- Besouw, M.; Masereeuw, R.; van den Heuvel, L.; Levtchenko, E. Cysteamine: An old drug with new potential. Drug Discov. Today 2013, 18, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Schogor, A.L.; Huws, S.A.; Santos, G.T.; Scollan, N.D.; Hauck, B.D.; Winters, A.L.; Kim, E.J.; Petit, H.V. Ruminal Prevotella spp. may play an important role in the conversion of plant lignans into human health beneficial antioxidants. PLoS ONE 2014, 9, e87949. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Xie, T.Y.; Janssen, P.H.; Zhao Sun, X.; Beauchemin, K.A.; Tan, Z.L.; Gao, M. Shifts in Rumen Fermentation and Microbiota Are Associated with Dissolved Ruminal Hydrogen Concentrations in Lactating Dairy Cows Fed Different Types of Carbohydrates. J. Nutr. 2016, 146, 1714–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, M.R.; Chaudhary, L.C.; Figueres, L.; Wallace, R.J. Metabolism of polyunsaturated fatty acids and their toxicity to the microflora of the rumen. Antonie Van Leeuwenhoek 2007, 91, 303–314. [Google Scholar] [CrossRef]
- Chen, H.; Guo, B.; Yang, M.; Luo, J.; Hu, Y.; Qu, M.; Song, X. Response of Growth Performance, Blood Biochemistry Indices, and Rumen Bacterial Diversity in Lambs to Diets Containing Supplemental Probiotics and Chinese Medicine Polysaccharides. Front. Vet. Sci. 2021, 8, 681389. [Google Scholar] [CrossRef]
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide utilization by gut bacteria: Potential for new insights from genomic analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [CrossRef]
- Maekawa, M.; Beauchemin, K.A.; Christensen, D.A. Effect of concentrate level and feeding management on chewing activities, saliva production, and ruminal pH of lactating dairy cows. J. Dairy Sci. 2002, 85, 1165–1175. [Google Scholar] [CrossRef]
- Xue, F.; Pan, X.; Jiang, L.; Guo, Y.; Xiong, B. GC–MS analysis of the ruminal metabolome response to thiamine supplementation during high grain feeding in dairy cows. Metabolomics 2018, 14, 67. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Mu, Y.Y.; Zhang, R.Y.; Xue, Y.F.; Guo, C.Z.; Qi, W.P.; Zhanga, J.; Maoac, S. Responsive changes of rumen microbiome and metabolome in dairy cows with different susceptibility to subacute ruminal acidosis. Anim. Nutr. 2022, 8, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Yost, W.M.; Young, J.W.; Schmidt, S.P.; McGilliard, A.D. Gluconeogenesis in Ruminants: Propionic Acid Production from a High-Grain Diet Fed to Cattle2. J. Nutr. 1977, 107, 2036–2043. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.N.; Méndez-García, C.; Geier, R.R.; Iakiviak, M.; Chang, J.; Cann, I.; Mackie, R.I. Metabolic networks for nitrogen utilization in Prevotella ruminicola 23. Sci. Rep. 2017, 7, 7851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokusaeva, K.; Fitzgerald, G.F.; Van Sinderen, D. Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 2011, 6, 285–306. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, B.; Wright, A.D.G. Comparative metagenomic analysis of bacterial populations in three full-scale mesophilic anaerobic manure digesters. Appl. Microbiol. Biotechnol. 2014, 98, 2709–2717. [Google Scholar] [CrossRef]
- Sutton, J.D.; Dhanoa, M.S.; Morant, S.V.; France, J.; Napper, D.J.; Schuller, E. Rates of Production of Acetate, Propionate, and Butyrate in the Rumen of Lactating Dairy Cows Given Normal and Low-Roughage Diets. J. Dairy Sci. 2003, 86, 3620–3633. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items 1 | Low-Forage (LF) | High-Forage (HF) |
|---|---|---|
| Fermentation substrates composition | ||
| L. chinensis hay | 300 | 700 |
| Corn meal | 700 | 300 |
| Nutrition concentrations | ||
| CP | 81.2 | 73.9 |
| EE | 26.3 | 16.3 |
| NDF | 153.1 | 547.2 |
| ADF | 294.0 | 323.8 |
| NFC | 727.3 | 350.2 |
| Ash | 12.1 | 12.4 |
| Item 1 | CS Supplementation Level (mg/g Based on Substrate Weight) | SEM | p-Value 2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | S | I | L | Q | |||
| IVDMD48, g/kg DM | LF | 71.8 | 73.1 | 75.9 | 77.7 | 2.34 | <0.01 | 0.16 | 0.04 | 0.42 |
| HF | 56.9 | 59.0 | 62.2 | 61.5 | ||||||
| GP48, mL/g DM | LF | 133.7 | 139.8 | 152.5 | 154.5 | 2.06 | <0.01 | 0.038 | <0.01 | 0.319 |
| HF | 126.9 | 131.5 | 137.6 | 138.1 | ||||||
| A, mL/g DM | LF | 148.7 | 156.4 | 165.6 | 167.5 | 2.31 | <0.01 | 0.441 | <0.01 | 0.267 |
| HF | 136.3 | 140.8 | 151.4 | 151.8 | ||||||
| B | LF | 1.05 | 1.21 | 1.37 | 1.35 | 0.023 | <0.01 | 0.121 | <0.01 | <0.01 |
| HF | 0.96 | 1.07 | 1.20 | 1.21 | ||||||
| C, h | LF | 6.40 | 5.43 | 3.93 | 3.87 | 0.094 | <0.01 | 0.457 | <0.01 | <0.01 |
| HF | 6.83 | 5.64 | 4.36 | 4.40 | ||||||
| AGPR, mL/h | LF | 6.11 | 8.75 | 14.49 | 14.66 | 0.503 | <0.01 | <0.01 | <0.01 | 0.03 |
| HF | 4.80 | 6.70 | 10.47 | 10.43 | ||||||
| Item 1 | CS Supplementation Level (mg/g Based on Substrate Weight) | SEM | p-Value 2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | S | I | L | Q | |||
| pH | LF | 6.20 | 6.14 | 6.13 | 6.16 | 0.045 | <0.01 | 0.08 | 0.18 | 0.11 |
| HF | 6.54 | 6.41 | 6.40 | 6.43 | ||||||
| Ammonia N, mg/dL | LF | 44.5 | 42.2 | 37.9 | 34.6 | 0.58 | 0.01 | 0.680 | <0.01 | 0.395 |
| HF | 42.1 | 40.6 | 36.7 | 34.1 | ||||||
| MCP, mg/mL | LF | 0.49 | 0.57 | 0.74 | 0.81 | 0.017 | <0.01 | <0.01 | <0.01 | 0.395 |
| HF | 0.45 | 0.56 | 0.64 | 0.67 | ||||||
| Total VFA, mmol/L | LF | 101.2 | 113.6 | 126.1 | 128.8 | 12.03 | <0.01 | 0.179 | <0.01 | 0.079 |
| HF | 93.1 | 96.7 | 111.7 | 112.5 | ||||||
| VFA patterns, % molar | ||||||||||
| Acetate | LF | 55.9 | 53.4 | 51.9 | 50.0 | 2.97 | <0.01 | 0.342 | <0.01 | 0.079 |
| HF | 60.1 | 54.7 | 52.9 b | 51.3 | ||||||
| Propionate | LF | 16.2 | 17.5 | 19.0 | 20.0 | 0.39 | 0.041 | 0.035 | <0.01 | 0.019 |
| HF | 15.5 | 17.2 | 18.2 | 19.8 | ||||||
| Butyrate | LF | 2.54 | 2.43 | 2.52 | 2.51 | 0.085 | 0.914 | 0.248 | 0.714 | 0.395 |
| HF | 2.52 | 2.56 | 2.40 | 2.51 | ||||||
| A:P | LF | 3.97 | 3.06 | 2.72 | 2.52 | 0.072 | 0.022 | 0.107 | <0.01 | <0.01 |
| HF | 3.90 | 3.34 | 2.91 | 2.60 | ||||||
| Item 1 | CS Supplementation Level (mg/g Based on Substrate Weight) | SEM | p-Value 2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | S | I | L | Q | |||
| SOD, U/mL | LF | 93.9 | 99.3 | 99.7 | 104.8 | 1.31 | <0.01 | 0.39 | <0.01 | 0.62 |
| HF | 90.7 | 94.2 | 95.4 | 96.5 | ||||||
| MDA, nmoL/mL | LF | 1.63 a | 1.53 | 1.49 | 1.39 | 0.035 | <0.01 | 0.39 | <0.01 | 0.54 |
| HF | 1.82 | 1.73 | 1.63 | 1.63 | ||||||
| CAT, U/mL | LF | 10.63 | 10.66 | 11.07 | 11.14 | 0.139 | 0.35 | <0.01 | <0.01 | 0.58 |
| HF | 9.98 | 11.00 | 11.14 | 11.83 | ||||||
| GSH-Px, U/mL | LF | 1095 | 1159 | 1178 | 1201 | 20.5 | <0.01 | <0.01 | <0.01 | 0.44 |
| HF | 988 | 992 | 1124 | 1232 | ||||||
| T-AOC, U/mL | LF | 5.43 | 5.82 | 6.02 | 6.20 | 0.129 | <0.01 | <0.01 | <0.01 | 0.29 |
| HF | 4.78 | 5.03 | 5.32 | 6.35 | ||||||
| GSH, U/mL | LF | 3.79 | 4.09 | 4.25 | 4.23 | 0.135 | 0.83 | <0.01 | <0.01 | 0.08 |
| HF | 3.51 | 3.74 | 3.76 | 5.27 | ||||||
| GR, U/mL | LF | 7.93 | 8.14 | 8.17 | 8.27 | 0.126 | <0.01 | 0.04 | 0.032 | 0.06 |
| HF | 7.32 | 7.53 | 8.24 | 7.57 | ||||||
| Item | CS Supplementation Level (mg/g Based on Substrate Weight) | SEM | p-Value 1 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | S | I | L | Q | |||
| Coverage | LF | 0.99 | 0.99 | 0.99 | 0.99 | <0.01 | 0.715 | 0.089 | 0.289 | 0.793 |
| HF | 0.99 | 0.99 | 0.99 | 0.99 | ||||||
| Chao | LF | 1316 | 1314 | 1208 | 17.7 | <0.01 | 0.951 | 0.508 | 0.064 | |
| HF | 1193 | 1203 | 1240 | 1237 | ||||||
| Ace | LF | 1287 | 1285 | 1281 | 1203 | 16.1 | <0.01 | 0.759 | 0.358 | 0.094 |
| HF | 1179 | 1190 | 1213 | 1200 | ||||||
| Simpson | LF | 0.020 | 0.024 | 0.027 | 0.038 | <0.01 | <0.01 | 0.139 | 0.292 | 0.903 |
| HF | 0.035 | 0.044 | 0.035 | 0.032 | ||||||
| Shannon | LF | 5.00 | 4.89 | 4.87 | 4.70 | 0.056 | <0.01 | 0.157 | 0.278 | 0.953 |
| HF | 4.71 | 4.65 | 4.75 | 4.77 | ||||||
| Sobs | LF | 1031 | 1019 | 1029 | 968 | 11.1 | <0.01 | 0.557 | 0.357 | 0.136 |
| HF | 968 | 967 | 995 | 980 | ||||||
| Item, % | CS Supplementation Level (mg/g Based on Substrate Weight) | SEM | p-Value 1 | ||||
|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | L | Q | ||
| Succiniclasticum | 18.9 | 21.2 | 20.4 | 19.1 | 3.066 | 0.982 | 0.564 |
| Bifidobacterium | 8.13 | 6.79 | 6.29 | 7.21 | 0.679 | 0.296 | 0.113 |
| norank_f_Muribaculaceae | 7.86 | 6.36 | 6.77 | 6.36 | 0.716 | 0.160 | 0.727 |
| Rikenellaceae_RC9_gut | 7.23 | 6.17 | 7.03 | 7.16 | 0.982 | 0.886 | 0.554 |
| Christensenellaceae_R-7 | 5.59 | 5.37 | 5.12 | 5.04 | 0.479 | 0.384 | 0.897 |
| Lachnospiraceae_NK3A20 | 5.95 | 4.85 | 4.78 | 4.95 | 0.420 | 0.119 | 0.148 |
| Prevotella | 2.35 | 3.92 | 3.17 | 5.09 | 0.699 | 0.027 | 0.810 |
| NK4A214_group | 3.07 | 2.71 | 2.72 | 2.52 | 0.199 | 0.080 | 0.664 |
| norank_f_F082 | 2.47 | 2.66 | 2.98 | 3.20 | 0.287 | 0.028 | 0.373 |
| Desulfovibrio | 2.40 | 2.85 | 2.52 | 2.04 | 0.209 | 0.146 | 0.038 |
| norank_f_Eubacterium_coprostanoligenes | 2.61 | 2.73 | 2.21 | 1.96 | 0.200 | 0.013 | 0.366 |
| Family_XIII_AD3011 | 2.47 | 2.52 | 1.98 | 1.75 | 0.242 | 0.022 | 0.559 |
| Ruminococcus | 1.74 | 2.32 | 2.38 | 2.13 | 0.187 | 0.154 | 0.039 |
| Fretibacterium | 1.38 | 1.36 | 1.84 | 2.03 | 0.312 | 0.098 | 0.733 |
| norank_o_WCHB1-41 | 1.77 | 1.19 | 1.93 | 1.65 | 0.541 | 0.873 | 0.780 |
| Prevotellaceae_UCG-001 | 0.64 | 1.82 | 1.36 | 2.42 | 0.486 | 0.037 | 0.899 |
| Eubacterium_nodatum | 1.61 | 1.45 | 1.62 | 1.50 | 0.199 | 0.865 | 0.921 |
| Pyramidobacter | 1.10 | 1.14 | 1.33 | 1.74 | 0.250 | 0.075 | 0.462 |
| Veillonellaceae_UCG-001 | 0.99 | 1.18 | 1.32 | 1.23 | 0.231 | 0.427 | 0.577 |
| Item 1 | CS Supplementation Level (mg/g Based on Substrate Weight) | SEM | p-Value 1 | ||||
|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | L | Q | ||
| Rikenellaceae_RC9_gut | 12.9 | 13.5 | 12.2 | 12.4 | 1.041 | 0.322 | 0.127 |
| Succiniclasticum | 12.4 | 13.8 | 12.3 | 9.16 | 2.912 | 0.402 | 0.439 |
| Bifidobacterium | 4.90 | 6.51 | 6.66 | 14.16 | 1.132 | <0.01 | 0.017 |
| Muribaculaceae | 5.87 | 5.85 | 7.55 | 8.33 | 1.741 | 0.262 | 0.797 |
| Prevotella | 5.87 | 6.00 | 6.58 | 7.51 | 1.594 | <0.01 | 0.066 |
| Lachnospiraceae_NK3A20 | 5.06 | 4.27 | 4.11 | 6.49 | 0.666 | 0.181 | 0.028 |
| Christensenellaceae_R-7 | 6.24 | 5.01 | 4.54 | 3.71 | 0.448 | <0.01 | 0.667 |
| norank_f_F082 | 2.88 | 2.90 | 3.66 | 5.61 | 0.348 | <0.01 | 0.012 |
| Prevotellaceae_UCG-001 | 2.81 | 4.63 | 4.06 | 4.35 | 0.891 | <0.01 | 0.089 |
| Fretibacterium | 2.87 | 3.70 | 3.46 | 3.03 | 1.025 | 0.961 | 0.548 |
| NK4A214_group | 2.40 | 1.97 | 1.95 | 2.45 | 0.208 | 0.889 | 0.035 |
| Desulfovibrio | 3.19 | 2.19 | 1.85 | 1.21 | 0.221 | <0.01 | 0.431 |
| norank_f_Eubacterium_coprostanoligenes | 2.03 | 1.94 | 1.77 | 2.45 | 0.208 | 0.255 | 0.082 |
| Ruminococcus | 2.46 | 1.92 | 2.02 | 1.13 | 0.247 | <0.01 | 0.485 |
| Family_XIII_AD3011 | 2.20 | 1.97 | 1.52 | 1.18 | 0.271 | <0.01 | 0.844 |
| Pyramidobacter | 1.03 | 1.15 | 1.53 | 1.16 | 0.342 | 0.623 | 0.482 |
| VeillonellaceaeUCG001 | 0.99 | 1.44 | 1.24 | 1.09 | 0.367 | 0.956 | 0.426 |
| Prevotellaceae_UCG-003 | 1.14 | 0.91 | 1.13 | 0.75 | 0.278 | 0.445 | 0.779 |
| Sharpea | 0.59 | 0.83 | 0.76 | 1.71 | 0.150 | <0.01 | 0.028 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Chen, H.; Zhang, F.; Wang, W.; Xiong, F.; Liu, Y.; Lv, L.; Li, W.; Bo, Y.; Yang, H. Cysteamine Supplementation In Vitro Remarkably Promoted Rumen Fermentation Efficiency towards Propionate Production via Prevotella Enrichment and Enhancing Antioxidant Capacity. Antioxidants 2022, 11, 2233. https://doi.org/10.3390/antiox11112233
Wu Q, Chen H, Zhang F, Wang W, Xiong F, Liu Y, Lv L, Li W, Bo Y, Yang H. Cysteamine Supplementation In Vitro Remarkably Promoted Rumen Fermentation Efficiency towards Propionate Production via Prevotella Enrichment and Enhancing Antioxidant Capacity. Antioxidants. 2022; 11(11):2233. https://doi.org/10.3390/antiox11112233
Chicago/Turabian StyleWu, Qichao, Hewei Chen, Fan Zhang, Weikang Wang, Fengliang Xiong, Yingyi Liu, Liangkang Lv, Wenjuan Li, Yukun Bo, and Hongjian Yang. 2022. "Cysteamine Supplementation In Vitro Remarkably Promoted Rumen Fermentation Efficiency towards Propionate Production via Prevotella Enrichment and Enhancing Antioxidant Capacity" Antioxidants 11, no. 11: 2233. https://doi.org/10.3390/antiox11112233