
Comparable Benefits of Stingless Bee Honey and Caffeic Acid in Mitigating the Negative Effects of Metabolic Syndrome on the Brain
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
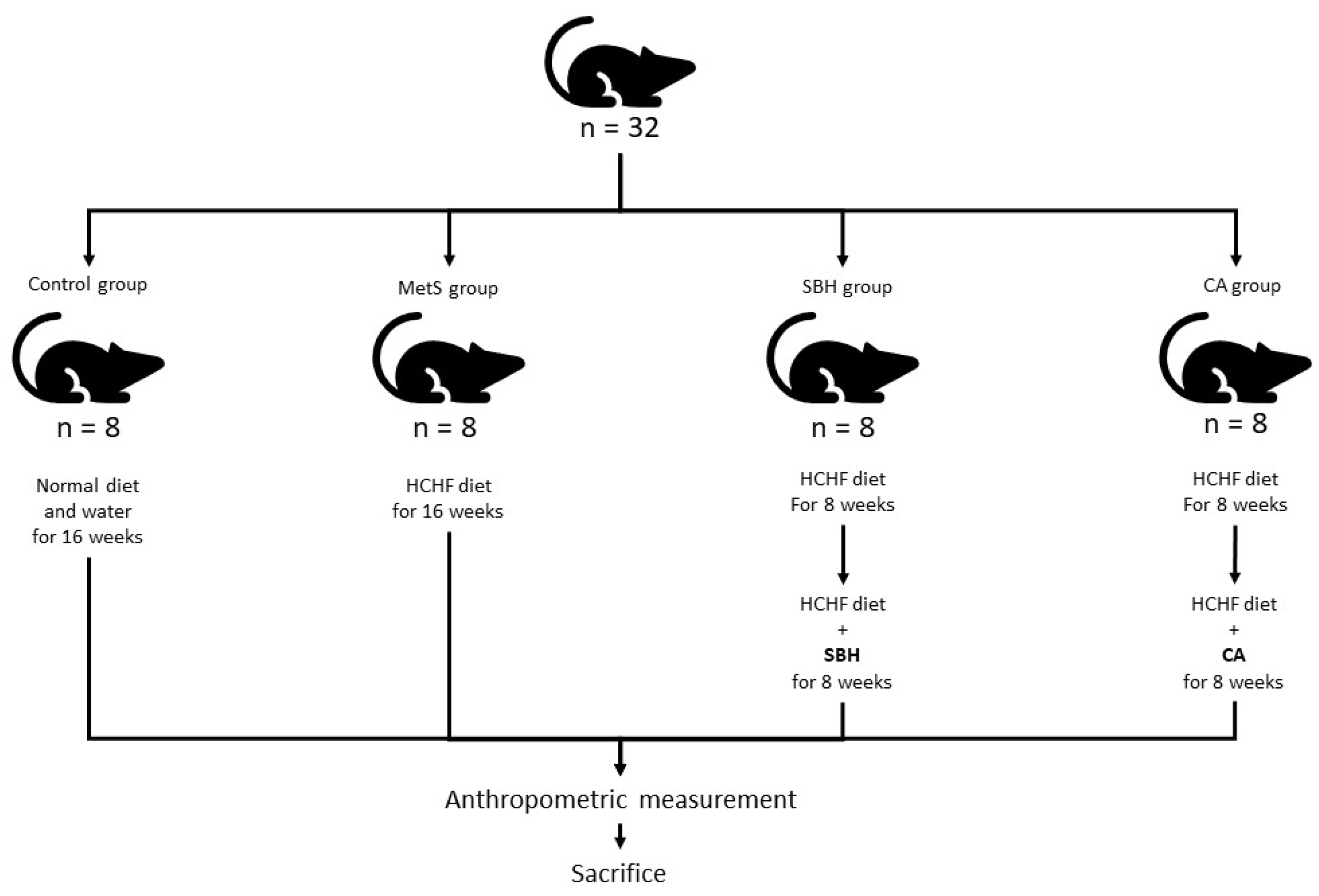
2.1. Animals and Treatment
2.2. Fructose Solution and High-Fructose/High-Fat Diet Preparation
2.3. Stingless Bee Honey and CA Solution
2.4. Anthropometric Measurement
2.5. Brain Extraction
2.6. Serum Biochemical Analyses
2.7. ELISA Test
2.8. Immunohistology
2.8.1. EdU (5-Ethynyl-2′-deoxyuridine)
2.8.2. TUNEL Assay
2.9. Statistical Analysis
3. Results
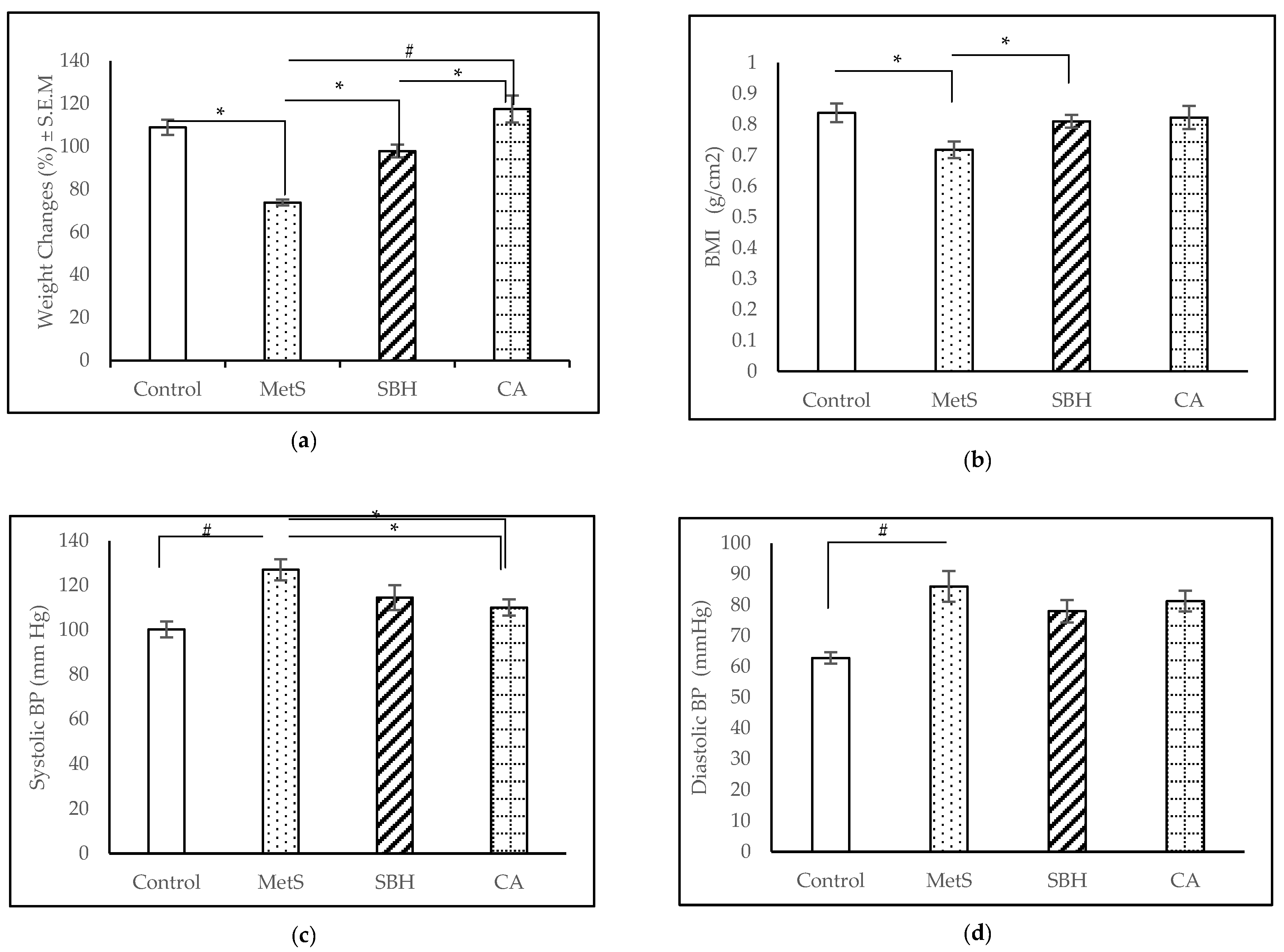
3.1. Anthropometrical and Physiological Measurements
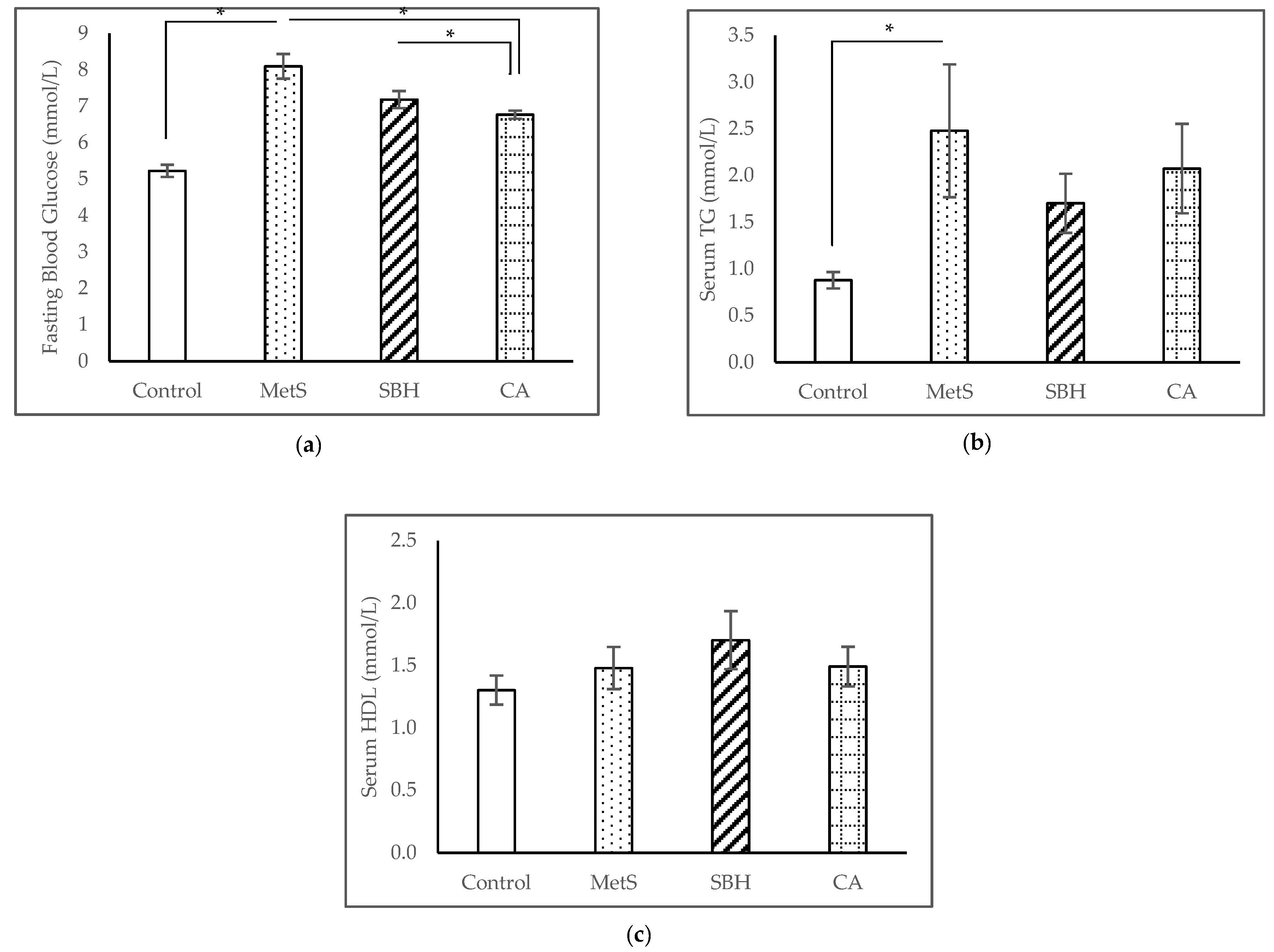
3.2. Serum Biochemical Analyses
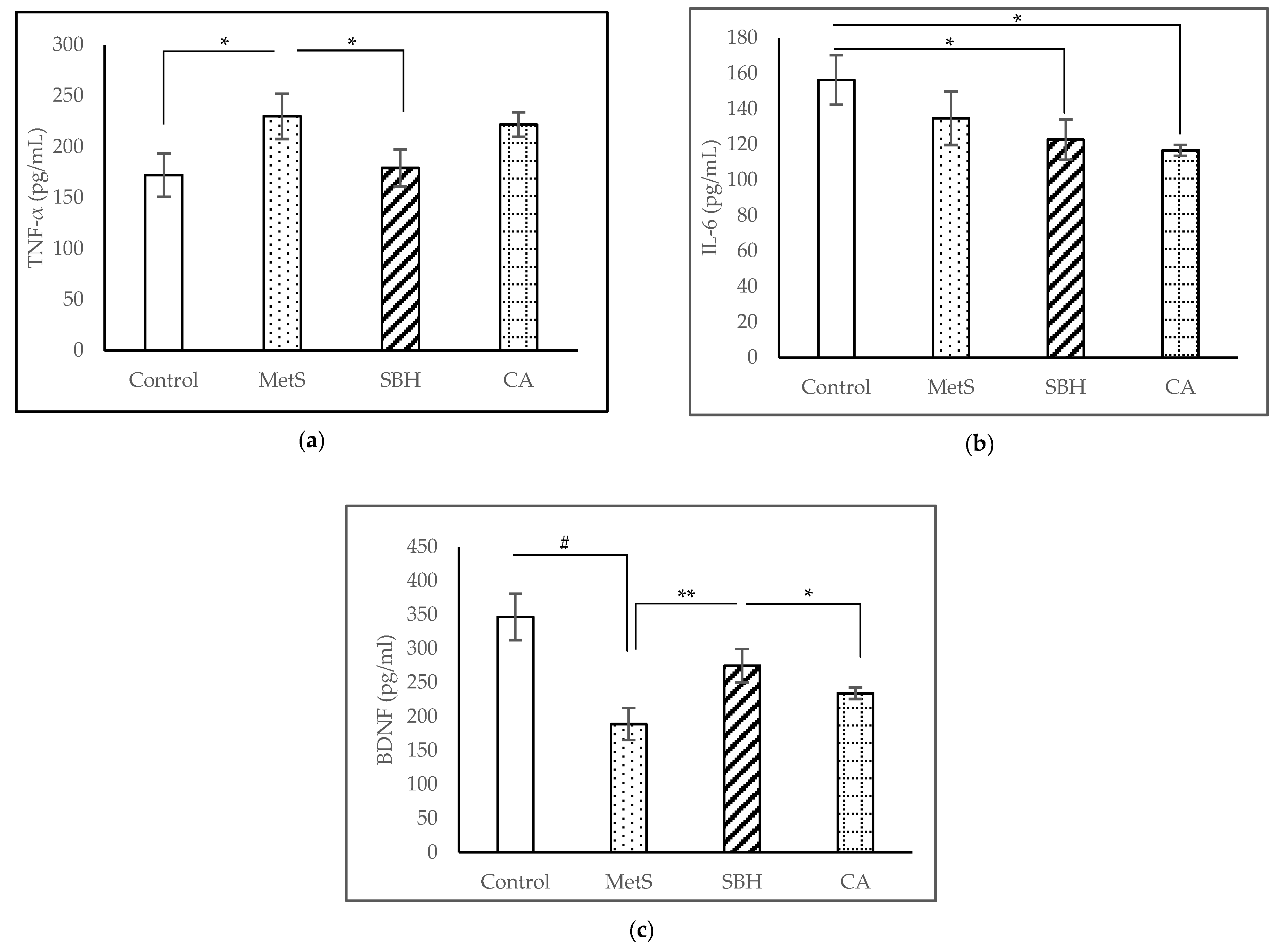
3.3. ELISA Studies
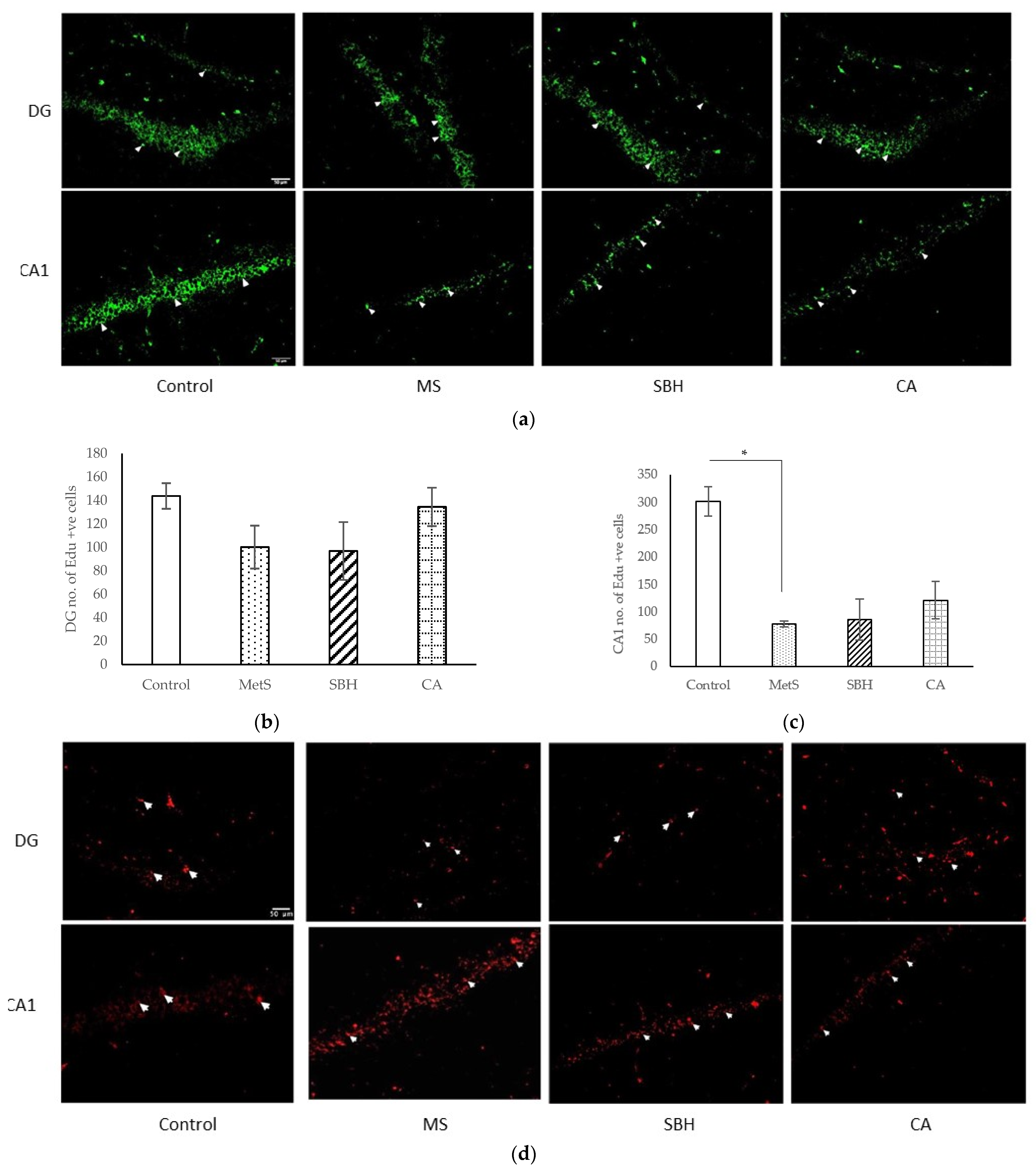
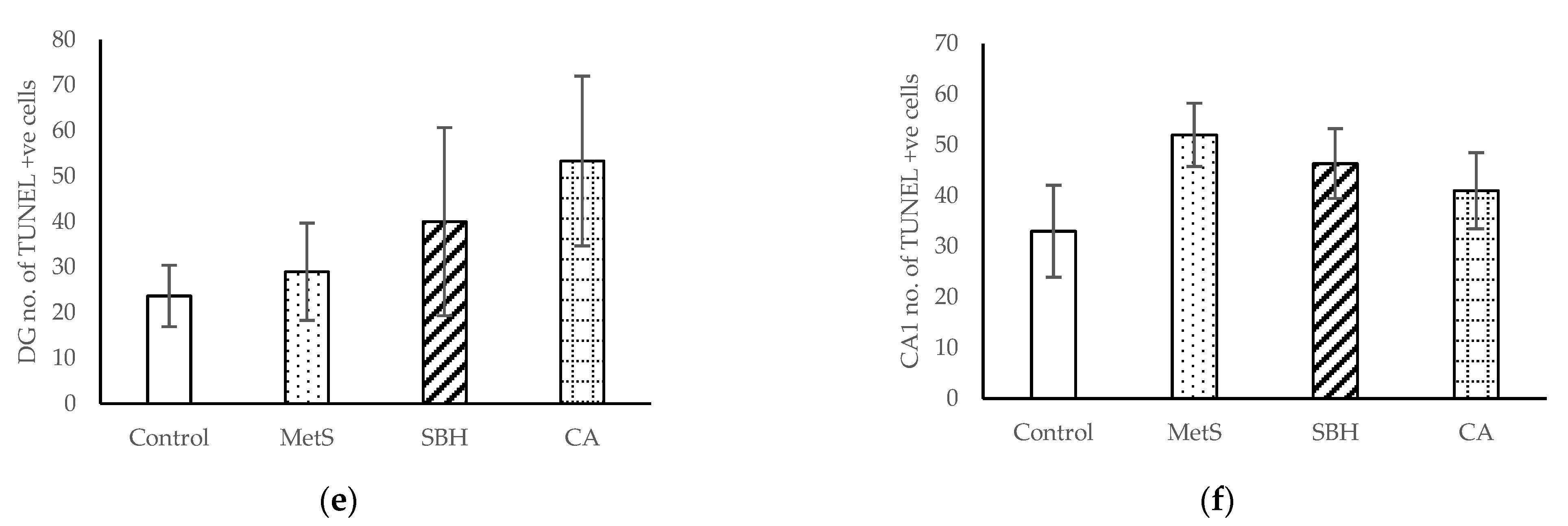
3.4. Immunohistological Studies
4. Discussion
4.1. MetS Induction with HCHF Diet
4.2. Effect of SBH and CA on MetS Model
4.3. Effect of SBH and CA on Inflammatory Status and BDNF Level of MetS Model
4.4. Effect of SBH and CA on Neurogenesis and Apoptosis of Brain Cells of MetS Model
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberti, K.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.-C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the Metabolic Syndrome. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dyken, P.; Lacoste, B. Impact of Metabolic Syndrome on Neuroinflammation and the Blood–Brain Barrier. Front. Neurosci. 2018, 12, 930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshad, N.; Lin, T.S.; Yahaya, M.F. Stingless Bee Honey Reduces Anxiety and Improves Memory of the Metabolic Disease-Induced Rats. CNS Neurol. Disord.—Drug Targets 2020, 19, 115–126. [Google Scholar] [CrossRef] [PubMed]
- McNay, E.C.; Recknagel, A.K. Reprint of: ‘Brain Insulin Signaling: A Key Component of Cognitive Processes and a Potential Basis for Cognitive Impairment in Type 2 Diabetes’. Neurobiol. Learn. Mem. 2011, 96, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Gancheva, S.; Zhelyazkova-Savova, M.; Galunska, B.; Chervenkov, T. Experimental Models of Metabolic Syndrome in Rats. Scr. Sci. Med. 2015, 47, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Arshad, N.; Lin, T.S.; Yahaya, M.F. Metabolic Syndrome and Its Effect on the Brain: Possible Mechanism. CNS Neurol. Disord. Drug Targets 2018, 17, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Otero, M.C.B.; Bernolo, L. Honey as Functional Food and Prospects in Natural Honey Production. In Functional Foods and Nutraceuticals: Bioactive Components, Formulations and Innovations; Egbuna, C., Dable Tupas, G., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 197–210. ISBN 978-3-030-42319-3. [Google Scholar]
- Yaacob, M.; Rajab, N.F.; Shahar, S.; Sharif, R. Stingless Bee Honey and Its Potential Value: A Systematic Review. Food Res. 2017, 2, 124–133. [Google Scholar] [CrossRef]
- Lage, L.G.A.; Coelho, L.L.; Resende, H.C.; Tavares, M.G.; Campos, L.A.O.; Fernandes-Salomão, T.M. Honey Physicochemical Properties of Three Species of the Brazilian Melipona. An. Acad. Bras. Ciênc. 2012, 84, 605–608. [Google Scholar] [CrossRef] [Green Version]
- Mohamad, M.A.M.; Mazlan, M.A.; Ibrahim, M.; Mat Yusof, A.; Shamsuddin, S.A.A.; Nik Hassan, N.F.; Muhammad, H.; Md Isa, M.L. The Effect of Malaysian Stingless Bee, Trigona Spp. Honey in Promoting Proliferation of the Undifferentiated Stem Cell. Asia Pac. J. Mol. Biol. Biotechnol. 2019, 27, 10–19. [Google Scholar] [CrossRef]
- Ranneh, Y.; Ali, F.; Zarei, M.; Akim, A.M.; Hamid, H.A.; Khazaai, H. Malaysian Stingless Bee and Tualang Honeys: A Comparative Characterization of Total Antioxidant Capacity and Phenolic Profile Using Liquid Chromatography-Mass Spectrometry. LWT 2018, 89, 1–9. [Google Scholar] [CrossRef]
- Putteeraj, M.; Lim, W.L.; Teoh, S.L.; Yahaya, M.F. Flavonoids and Its Neuroprotective Effects on Brain Ischemia and Neurodegenerative Diseases. Curr. Drug Targets 2018, 19, 1710–1720. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Lee, M.-K.; Park, Y.B.; Jeon, S.-M.; Choi, M.-S. Antihyperglycemic and Antioxidant Properties of Caffeic Acid in Db/Db Mice. J. Pharmacol. Exp. Ther. 2006, 318, 476–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agunloye, O.M.; Oboh, G. Hypercholesterolemia, Angiotensin Converting Enzyme and Ecto-Enzymes of Purinergic System: Ameliorative Properties of Caffeic and Chlorogenic Acid in Hypercholesterolemic Rats. J. Food Biochem. 2018, 42, e12604. [Google Scholar] [CrossRef]
- Bi, Q.; Hou, J.; Qi, P.; Ma, C.; Shen, Y.; Feng, R.; Yan, B.; Wang, J.; Shi, X.; Zheng, Y.; et al. Venenum Bufonis Induces Rat Neuroinflammation by Activiating NF-ΚB Pathway and Attenuation of BDNF. J. Ethnopharmacol. 2016, 186, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Brown, L. Rodent Models for Metabolic Syndrome Research. J. Biomed. Biotechnol. 2011, 2011, 351982. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. The Effects of a Modified High-Carbohydrate High-Fat Diet on Metabolic Syndrome Parameters in Male Rats. Exp. Clin. Endocrinol. Diabetes 2018, 126, 205–212. [Google Scholar] [CrossRef]
- Quesada, O.; Claggett, B.; Rodriguez, F.; Cai, J.; Moncrieft, A.E.; Garcia, K.; Del Rios Rivera, M.; Hanna, D.B.; Daviglus, M.L.; Talavera, G.A.; et al. Associations of Insulin Resistance with Systolic and Diastolic Blood Pressure: A Study From the HCHS/SOL. Hypertension 2021, 78, 716–725. [Google Scholar] [CrossRef]
- Panchal, S.K.; Ward, L.; Brown, L. Ellagic Acid Attenuates High-Carbohydrate, High-Fat Diet-Induced Metabolic Syndrome in Rats. Eur. J. Nutr. 2013, 52, 559–568. [Google Scholar] [CrossRef]
- Bargut, T.C.L.; Santos, L.P.; Machado, D.G.L.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Eicosapentaenoic Acid (EPA) vs. Docosahexaenoic Acid (DHA): Effects in Epididymal White Adipose Tissue of Mice Fed a High-Fructose Diet. Prostaglandins Leukot. Essent. Fat. Acids 2017, 123, 14–24. [Google Scholar] [CrossRef]
- Ramli, N.Z.; Chin, K.-Y.; Zarkasi, K.A.; Ahmad, F. The Beneficial Effects of Stingless Bee Honey from Heterotrigona Itama against Metabolic Changes in Rats Fed with High-Carbohydrate and High-Fat Diet. Int. J. Environ. Res. Public Health 2019, 16, E4987. [Google Scholar] [CrossRef]
- Mohd Kamal, D.A.; Ibrahim, S.F.; Kamal, H.; Kashim, M.I.A.M.; Mokhtar, M.H. Physicochemical and Medicinal Properties of Tualang, Gelam and Kelulut Honeys: A Comprehensive Review. Nutrients 2021, 13, 197. [Google Scholar] [CrossRef] [PubMed]
- Muhammad Abdul Kadar, N.N.; Ahmad, F.; Teoh, S.L.; Yahaya, M.F. Caffeic Acid on Metabolic Syndrome: A Review. Molecules 2021, 26, 5490. [Google Scholar] [CrossRef] [PubMed]
- Nasry, M.R.; Abo-Youssef, A.M.; Zaki, H.F.; El-Denshary, E.-E.-D.S. Effect of Caffeic Acid and Pioglitazone in an Experimental Model of Metabolic Syndrome. Int. J. Sci. Res. Publ. 2015, 5, 1–9. Available online: http://www.ijsrp.org/research-paper-1015.php?rp=P464679 (accessed on 7 February 2021).
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, E1321. [Google Scholar] [CrossRef] [Green Version]
- Mariana, B.D.; Tiago, L.S.; Ramon, R.P.P.B.d.M.; Jamile, M.F.; Tiago, S.M.; Richard, R.C.M.; Hector, G.R.; Dânya, B.L.; Alice, M.C.M.; Maria, G.R.d.Q. Caffeic Acid Reduces Lipid Accumulation and Reactive Oxygen Species Production in Adipocytes. Afr. J. Pharm. Pharmacol. 2018, 12, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Mohd Rafie, A.Z.; Syahir, A.; Wan Ahmad, W.A.N.; Mustafa, M.Z.; Mariatulqabtiah, A.R. Supplementation of Stingless Bee Honey from Heterotrigona Itama Improves Antiobesity Parameters in High-Fat Diet Induced Obese Rat Model. Evid.-Based Complement. Altern. Med. 2018, 2018, e6371582. [Google Scholar] [CrossRef] [Green Version]
- Ramli, N.Z.; Chin, K.-Y.; Zarkasi, K.A.; Ahmad, F. A Review on the Protective Effects of Honey against Metabolic Syndrome. Nutrients 2018, 10, 1009. [Google Scholar] [CrossRef] [Green Version]
- Hashim, K.-N.; Chin, K.-Y.; Ahmad, F. The Mechanism of Honey in Reversing Metabolic Syndrome. Molecules 2021, 26, 808. [Google Scholar] [CrossRef]
- Oboh, G.; Ojueromi, O.O.; Ademosun, A.O.; Omayone, T.P.; Oyagbemi, A.A.; Ajibade, T.O.; Adedapo, A.A. Effects of Caffeine and Caffeic Acid on Selected Biochemical Parameters in L-NAME-Induced Hypertensive Rats. J. Food Biochem. 2021, 45, e13384. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, L.; Gao, P.; Liu, D.; Zhu, Z. Caffeic Acid Ameliorates Angiotensin II-Induced Increase In Blood Pressure By Activating Vascular Sarco-/Endoplasmic Reticulum CA-ATPASE2A. J. Hypertens. 2021, 39, e248. [Google Scholar] [CrossRef]
- Aziz, M.S.A.; Giribabu, N.; Rao, P.V.; Salleh, N. Pancreatoprotective Effects of Geniotrigona Thoracica Stingless Bee Honey in Streptozotocin-Nicotinamide-Induced Male Diabetic Rats. Biomed. Pharmacother. 2017, 89, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, M.; Azizi-Soleiman, F. Honey and Glycemic Control: A Systematic Review. PharmaNutrition 2020, 11, 100180. [Google Scholar] [CrossRef]
- Pasupuleti, V.R.; Arigela, C.S.; Gan, S.H.; Salam, S.K.N.; Krishnan, K.T.; Rahman, N.A.; Jeffree, M.S. A Review on Oxidative Stress, Diabetic Complications, and the Roles of Honey Polyphenols. Oxid. Med. Cell. Longev. 2020, 2020, e8878172. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, F.; Salehi, S.; Kohanmoo, A.; Akhlaghi, M. Effect of Natural Honey on Glycemic Control and Anthropometric Measures of Patients with Type 2 Diabetes: A Randomized Controlled Crossover Trial. Int. J. Prev. Med. 2019, 10, 3. [Google Scholar] [CrossRef]
- Rashid, M.R.; Nor Aripin, K.N.; Syed Mohideen, F.B.; Baharom, N.; Omar, K.; Md Taujuddin, N.M.S.; Mohd Yusof, H.H.; Addnan, F.H. The Effect of Kelulut Honey on Fasting Blood Glucose and Metabolic Parameters in Patients with Impaired Fasting Glucose. J. Nutr. Metab. 2019, 2019, e3176018. [Google Scholar] [CrossRef]
- Majid, M.; Younis, M.A.; Naveed, A.K.; Shah, M.U.; Azeem, Z.; Tirmizi, S.H. Effects of Natural Honey on Blood Glucose and Lipid Profile in Young Healthy Pakistani Males. J. Ayub. Med. Coll Abbottabad 2013, 25, 44–47. [Google Scholar]
- Akhbari, M.; Jabbari, M.; Ayati, M.H.; Namazi, N. The Effects of Oral Consumption of Honey on Key Metabolic Profiles in Adult Patients with Type 2 Diabetes Mellitus and Nondiabetic Individuals: A Systematic Review of Clinical Trials. Evid. -Based Complement. Altern. Med. 2021, 2021, e6666832. [Google Scholar] [CrossRef]
- Nakamura, Y.; Natsume, M.; Yasuda, A.; Ishizaka, M.; Kawahata, K.; Koga, J. Fructooligosaccharides Suppress High-Fat Diet-Induced Fat Accumulation in C57BL/6J Mice. BioFactors 2017, 43, 145–151. [Google Scholar] [CrossRef]
- Alkhalifah, M.K.; Alabduljabbar, K.A.; Alkhenizan, A.H. Effect of Natural Honey on Lowering Lipid Profile. Saudi Med. J. 2021, 42, 473–481. [Google Scholar] [CrossRef]
- Nie, J.; Chang, Y.; Li, Y.; Zhou, Y.; Qin, J.; Sun, Z.; Li, H. Caffeic Acid Phenethyl Ester (Propolis Extract) Ameliorates Insulin Resistance by Inhibiting JNK and NF-ΚB Inflammatory Pathways in Diabetic Mice and HepG2 Cell Models. J. Agric. Food Chem. 2017, 65, 9041–9053. [Google Scholar] [CrossRef]
- Oršolić, N.; Sirovina, D.; Odeh, D.; Gajski, G.; Balta, V.; Šver, L.; Jazvinšćak Jembrek, M. Efficacy of Caffeic Acid on Diabetes and Its Complications in the Mouse. Molecules 2021, 26, 3262. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.; Mong, M.; Chan, K.; Yin, M. Anti-Glycative and Anti-Inflammatory Effects of Caffeic Acid and Ellagic Acid in Kidney of Diabetic Mice. Mol. Nutr. Food Res. 2010, 54, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Srikanthan, K.; Feyh, A.; Visweshwar, H.; Shapiro, J.I.; Sodhi, K. Systematic Review of Metabolic Syndrome Biomarkers: A Panel for Early Detection, Management, and Risk Stratification in the West Virginian Population. Int. J. Med. Sci. 2016, 13, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Xiao, L.; Wang, H.; Wang, G. Neurogenic Hypothesis of Positive Psychology in Stress-Induced Depression: Adult Hippocampal Neurogenesis, Neuroinflammation, and Stress Resilience. Int. Immunopharmacol. 2021, 97, 107653. [Google Scholar] [CrossRef]
- Lehrskov, L.L.; Christensen, R.H. The Role of Interleukin-6 in Glucose Homeostasis and Lipid Metabolism. Semin. Immunopathol. 2019, 41, 491–499. [Google Scholar] [CrossRef]
- Lang Lehrskov, L.; Lyngbaek, M.P.; Soederlund, L.; Legaard, G.E.; Ehses, J.A.; Heywood, S.E.; Wewer Albrechtsen, N.J.; Holst, J.J.; Karstoft, K.; Pedersen, B.K.; et al. Interleukin-6 Delays Gastric Emptying in Humans with Direct Effects on Glycemic Control. Cell Metab. 2018, 27, 1201–1211.e3. [Google Scholar] [CrossRef] [Green Version]
- Kalantari, N.; Ghasemi, M.; Bayani, M.; Ghaffari, S. Effect of Honey on MRNA Expression of TNF-α, IL-1β and IL-6 Following Acute Toxoplasmosis in Mice. Cytokine 2016, 88, 85–90. [Google Scholar] [CrossRef]
- Biluca, F.C.; da Silva, B.; Caon, T.; Mohr, E.T.B.; Vieira, G.N.; Gonzaga, L.V.; Vitali, L.; Micke, G.; Fett, R.; Dalmarco, E.M.; et al. Investigation of Phenolic Compounds, Antioxidant and Anti-Inflammatory Activities in Stingless Bee Honey (Meliponinae). Food Res. Int. 2020, 129, 108756. [Google Scholar] [CrossRef]
- Grabska-Kobylecka, I.; Kaczmarek-Bak, J.; Figlus, M.; Prymont-Przyminska, A.; Zwolinska, A.; Sarniak, A.; Wlodarczyk, A.; Glabinski, A.; Nowak, D. Presence of Caffeic Acid in Cerebrospinal Fluid: Evidence That Dietary Polyphenols Can Cross the Blood-Brain Barrier in Humans. Nutrients 2020, 12, 1531. [Google Scholar] [CrossRef]
- Mudgal, J.; Basu Mallik, S.; Nampoothiri, M.; Kinra, M.; Hall, S.; Grant, G.D.; Anoopkumar-Dukie, S.; Davey, A.K.; Rao, C.M.; Arora, D. Effect of Coffee Constituents, Caffeine and Caffeic Acid on Anxiety and Lipopolysaccharide-Induced Sickness Behavior in Mice. J. Funct. Foods 2020, 64, 103638. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Malheiros, R.T.; Delgado, H.O.; Felber, D.T.; Kraus, S.I.; dos Santos, A.R.S.; Manfredini, V.; da Silva, M.D. Mood Disorders Are Associated with the Reduction of Brain Derived Neurotrophic Factor in the Hypocampus in Rats Submitted to the Hipercaloric Diet. Metab. Brain Dis. 2021, 36, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Davarpanah, M.; Shokri-mashhadi, N.; Ziaei, R.; Saneei, P. A Systematic Review and Meta-Analysis of Association between Brain-Derived Neurotrophic Factor and Type 2 Diabetes and Glycemic Profile. Sci. Rep. 2021, 11, 13773. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, M.Z.; Zulkifli, F.N.; Fernandez, I.; Mariatulqabtiah, A.R.; Sangu, M.; Nor Azfa, J.; Mohamed, M.; Roslan, N. Stingless Bee Honey Improves Spatial Memory in Mice, Probably Associated with Brain-Derived Neurotrophic Factor (BDNF) and Inositol 1,4,5-Triphosphate Receptor Type 1 (Itpr1) Genes. Evid.-Based Complement. Altern. Med. 2019, 2019, e8258307. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Ben, X.; Xiao, L.; Peng, M.; Zhang, Y. AMP-Activated Protein Kinase Activation in Mediating Phenylalanine-Induced Neurotoxicity in Experimental Models of Phenylketonuria. J. Inherit. Metab. Dis. 2018, 41, 679–687. [Google Scholar] [CrossRef]
- Marosi, K.; Mattson, M.P. BDNF Mediates Adaptive Brain and Body Responses to Energetic Challenges. Trends Endocrinol. Metab. 2014, 25, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Abbott, L.C.; Nigussie, F. Adult Neurogenesis in the Mammalian Dentate Gyrus. Anat. Histol. Embryol. 2020, 49, 3–16. [Google Scholar] [CrossRef]
- Zhou, W.; Yao, Y.; Li, J.; Wu, D.; Zhao, M.; Yan, Z.; Pang, A.; Kong, L. TIGAR Attenuates High Glucose-Induced Neuronal Apoptosis via an Autophagy Pathway. Front. Mol. Neurosci. 2019, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Van der Borght, K.; Köhnke, R.; Göransson, N.; Deierborg, T.; Brundin, P.; Erlanson-Albertsson, C.; Lindqvist, A. Reduced Neurogenesis in the Rat Hippocampus Following High Fructose Consumption. Regul. Pept. 2011, 167, 26–30. [Google Scholar] [CrossRef]
- Velásquez-Jiménez, D.; Corella-Salazar, D.A.; Zuñiga-Martínez, B.S.; Domínguez-Avila, J.A.; Montiel-Herrera, M.; Salazar-López, N.J.; Rodrigo-Garcia, J.; Villegas-Ochoa, M.A.; González-Aguilar, G.A. Phenolic Compounds That Cross the Blood–Brain Barrier Exert Positive Health Effects as Central Nervous System Antioxidants. Food Funct. 2021, 12, 10356–10369. [Google Scholar] [CrossRef]
- Huang, D.; Refaat, M.; Mohammedi, K.; Jayyousi, A.; Al Suwaidi, J.; Abi Khalil, C. Macrovascular Complications in Patients with Diabetes and Prediabetes. Biomed. Res. Int. 2017, 2017, 7839101. [Google Scholar] [CrossRef] [PubMed]
- Olianas, M.C.; Dedoni, S.; Onali, P. Inhibition of TNF-α-Induced Neuronal Apoptosis by Antidepressants Acting through the Lysophosphatidic Acid Receptor LPA1. Apoptosis 2019, 24, 478–498. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rodríguez, D.R.; Blanco-Luquin, I.; Mendioroz, M. The Participation of Microglia in Neurogenesis: A Review. Brain Sci. 2021, 11, 658. [Google Scholar] [CrossRef]
- Kusumastuty, I.; Sembiring, F.; Andarini, S.; Handayani, D. High-Fat-High-Fructose Diet Decreases Hippocampal Neuron Number in Male Rats. Indones. Biomed. J. 2020, 12, 1–7. [Google Scholar] [CrossRef]
- Saxena, A.K.; Phyu, H.P.; Al-Ani, I.M.; Talib, N.A. Potential Protective Effect of Honey against Chronic Cerebral Hypoperfusion-Induced Neurodegeneration in Rats. J. Anat. Soc. India 2014, 63, 151–155. [Google Scholar] [CrossRef]
- Giri, B.; Dey, S.; Das, T.; Sarkar, M.; Banerjee, J.; Dash, S.K. Chronic Hyperglycemia Mediated Physiological Alteration and Metabolic Distortion Leads to Organ Dysfunction, Infection, Cancer Progression and Other Pathophysiological Consequences: An Update on Glucose Toxicity. Biomed. Pharmacother. 2018, 107, 306–328. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Calvani, R.; Coelho-Junior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial Dysfunction, Oxidative Stress, and Neuroinflammation: Intertwined Roads to Neurodegeneration. Antioxidants 2020, 9, 647. [Google Scholar] [CrossRef] [PubMed]
- Lau, F.C.; Shukitt-Hale, B.; Joseph, J.A. The Beneficial Effects of Fruit Polyphenols on Brain Aging. Neurobiol. Aging 2005, 26, 128–132. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muhammad Abdul Kadar, N.N.; Ahmad, F.; Teoh, S.L.; Yahaya, M.F. Comparable Benefits of Stingless Bee Honey and Caffeic Acid in Mitigating the Negative Effects of Metabolic Syndrome on the Brain. Antioxidants 2022, 11, 2154. https://doi.org/10.3390/antiox11112154
Muhammad Abdul Kadar NN, Ahmad F, Teoh SL, Yahaya MF. Comparable Benefits of Stingless Bee Honey and Caffeic Acid in Mitigating the Negative Effects of Metabolic Syndrome on the Brain. Antioxidants. 2022; 11(11):2154. https://doi.org/10.3390/antiox11112154
Chicago/Turabian StyleMuhammad Abdul Kadar, Nellysha Namela, Fairus Ahmad, Seong Lin Teoh, and Mohamad Fairuz Yahaya. 2022. "Comparable Benefits of Stingless Bee Honey and Caffeic Acid in Mitigating the Negative Effects of Metabolic Syndrome on the Brain" Antioxidants 11, no. 11: 2154. https://doi.org/10.3390/antiox11112154