
Health Benefits and Pharmacological Properties of Stigmasterol
 ,
,  , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
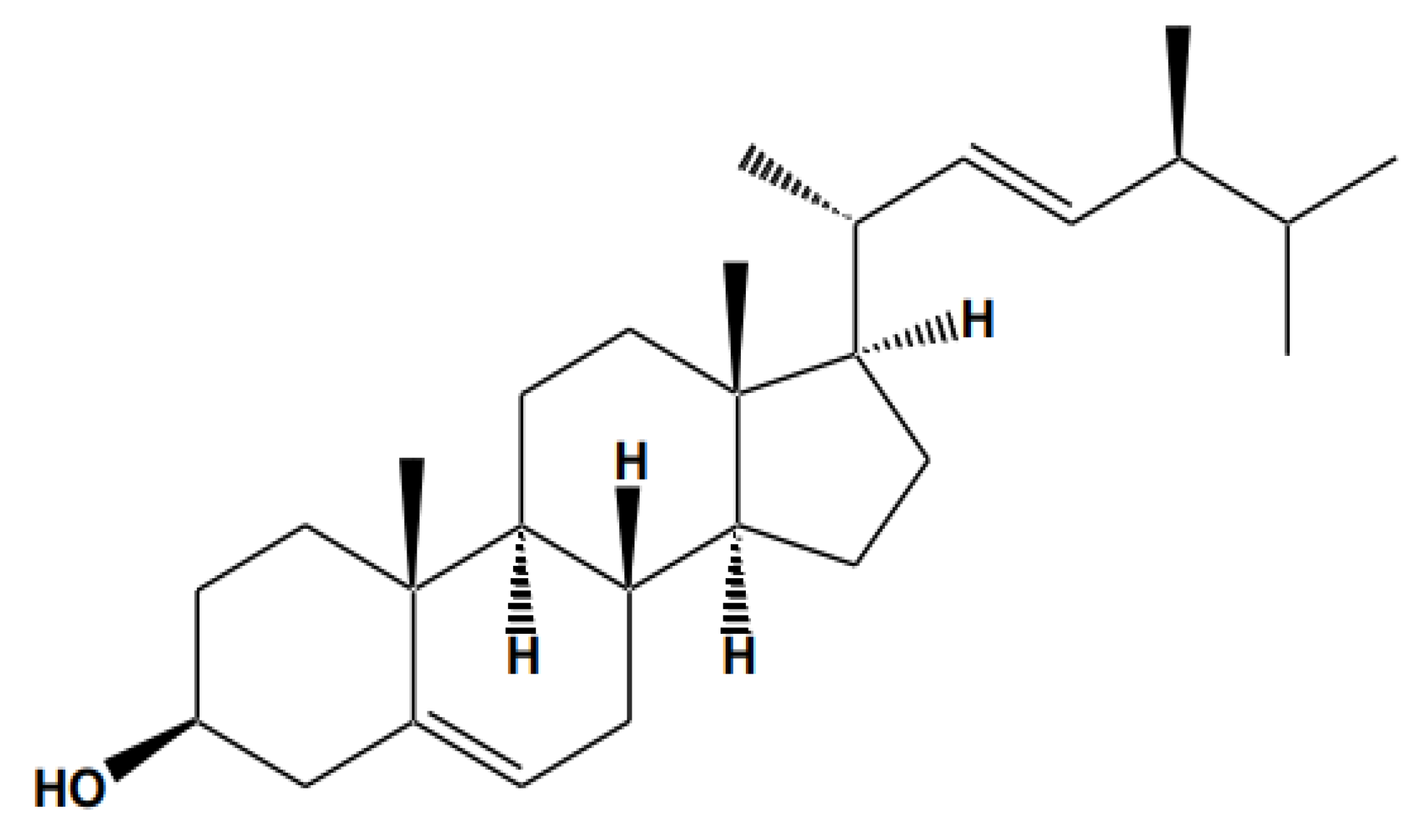
2. Sources and Physicochemical Properties of Stigmasterol
3. Anticancer Properties

{kind=link}
{kind=link}
| Cell Lines | Key Results | Ref |
|---|---|---|
| A2780 human ovarian cancer cell line SKOV3 human ovarian cancer cell line | A2780 cells (IC50 = 69.24 ± 7.31 for 24 h) SKOV3 cells (IC50 = 83.39 ± 3.75 for 24 h) Inhibited cell migration in A2780/SKOV3 cells Inhibited cell proliferation in A2780/SKOV3 cells Down-regulated the expression of p-PI3K/p-Akt protein levels Up-regulated the expression of PI3K/Akt/PTEN protein levels in SKOV3 cells Reduced the levels of PI3K and Akt in A2780/SKOV3cells Increased the expression of PTEN in A2780/SKOV3cells | [88] |
| Human gastric cancer cell line SGC-7901, MGC-803, and normal GES-1 cell line | Inhibited cell proliferation in SGC-7901/MGC-803 cells Induced apoptosis and autophagy in vitro Blocked Akt/mTOR signaling pathway Suppressed tumor growth of gastric cancer in vivo | [95] |
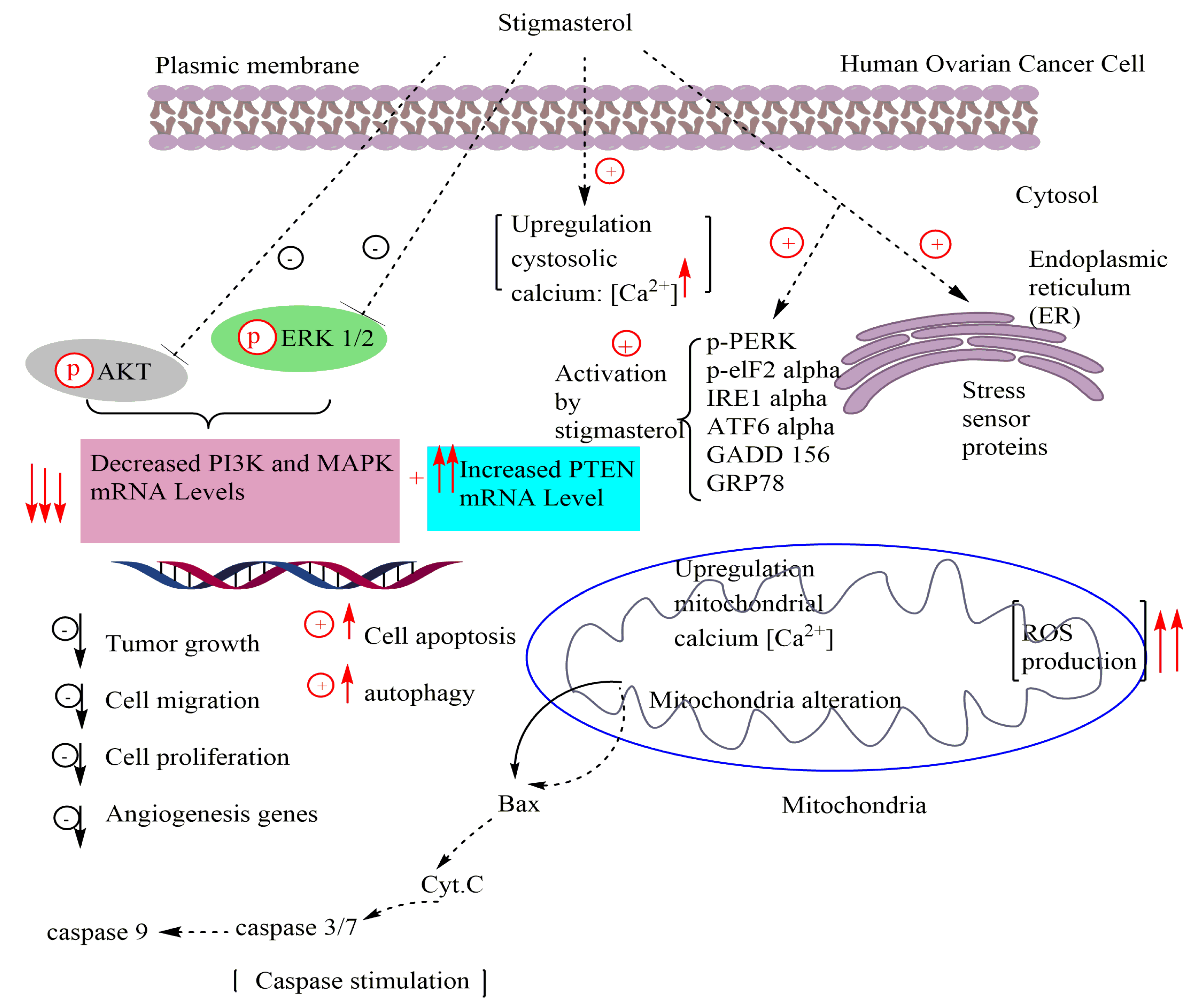
| Human ovarian cancer cells | Inhibited development of ES2 and OV90 cells Induced cell apoptosis Suppressed cell migration Inhibited angiogenesis genes Increased ROS production Increased calcium levels in the cytosol and mitochondria Increased mitochondrial depolarization Stimulated cell death Activated the ER-mitochondrial axis Inactivated PI3K and MAPK signal cascades | [87] |
| Human endometrial cancer cell lines, Ishikawa and SPEC2 | Inhibited Nrf2/NQO1/HO1 expression in endometrial cancer cells Suppressed Nrf2-ARE activity when combined with cisplatin Enhanced early apoptosis when combined with cisplatin Inhibited Tet1 expression/Tet1-induced hydroxymethylation Enhanced the inhibitory effect of cisplatin on migration and invasion | [36] |
| Human breast cancer cells (MDA-MB-231) and normal (HEK-293) cell lines | IC50 = 24.5 μg/mL Induced strong cytotoxic effects on MDA-MB-231 cells Increased ROS production Activated caspase-3 and -9 Induced apoptosis in MDA-MB-231 cells Increased levels of p53, Bax, caspase-3 and -9 Declined Bcl-2 gene expression Induced MDA-MB-231 cell death via apoptotic pathway | [34] |
| Molecular docking simulation In silico ADMET approach Molecular targets of breast cancer: ER-α, PR, HER2, and EGFR | Inhibited molecular targets of breast cancer: PR/ER-α/EGFR/HER2 Exhibited stronger binding affinities towards PR/ER-α/EGFR/HER2 Showed hydrophobic interaction with amino acid residue of PR/ER-α/EGFR/HER2 Identified as a substrate for P-glycoprotein and CYP3A4 Demonstrated high permeability for human intestinal absorption and Caco-2 cells Showed high blood–brain barrier permeability Showed no carcinogenicity | [42] |
| Human breast cancer cell lines MCF7 and MDA-MB-231 | Showed low cytotoxicity Significantly reduced levels of p-ERK/NF-Kb/VEGF/VEGFR-2 (p ≤ 0.05) Reduced Ki67 levels when combined with sorafenib (p ≤ 0.05) Markedly decreased VEGFR-2 mRNA gene expression Increased caspase-3 activity Decreased Bcl2 levels | [38] |
| Human gastric cancer cell line SNU-1, GES-1 normal cell line | IC50 = 15 µM Inhibited gastric cancer growth Inducted mitochondrial-mediated apoptosis Inhibited cancer cell migration Induced cell apoptosis Triggered G2/M cell cycle arrest in a dose-dependent manner Inhibited the JAK/STAT signalling pathway Exhibited minimal anticancer effects on the normal GES-1 cells Suppressed the phosphorylation of pSTAT1/pSTAT 2/pJAK1/pJAk2 | [89] |
| Lung cancer cell lines | Inhibited the proliferation Promoted the apoptosis Modulated retinoic acid-related orphan receptor C (RORC) Increased levels of caspase-3 and caspase-9 | [31] |
| Human hepatoma HepG2 cells | Showed potent cytotoxicity against HepG2 cells in a dose-dependent manner Up-regulated the levels of Bax, p53 Down-regulated the levels of Bcl-2 Induced apoptosis: caspase-8, 9 Increased apoptotic cell numbers Induced morphological changes Induced DNA damage | [32] |
| DMBA croton oil induced skin carcinoma | Reduced tumor size and cumulative number of papillomas Significantly increased the average latency period Decreased the levels of ASAT/ALAT/ALP Significantly increased glutathione (GSH)/superoxide dismutase (SOD)/catalase (CAT) Significantly inhibited high levels of lipid peroxide and DNA damage | [20] |
| Human gall bladder cancer cells | Up-regulated p27 expression gene Down-regulated Jab1 expression gene Activated caspase-3 Induced apoptosis Increased apoptotic cells and DNA | [92] |
| Ehrlich Ascites Carcinoma mice | Decreased tumor volume Decreased packed cell volume and viable cell count Increased mean survival time Increased life span of EAC tumor bearing mice Decreased the levels of lipid peroxidation Increased the levels of GSH/SOD/CAT Increased LDH activity in ascitic fluid Decreased membrane microviscosity Activated protein phosphatase 2A by ceramide Protected the heamoto-poietic system Increased the hemoglobin content/RBC count Reduced WBC count Increased platelet count | [93] |
| Human umbilical vein endothelial cells (HUVECs) Human cholangiocarcinoma (CCA) cells | IC50 = 21.1 ± 2.1 μM Suppressed cell viability, migration, and morphogenesis on HUVECs, but not CCA cells Significantly reduced the transcript level of TNF-α Decreased levels of VEGFR-2/Src/Akt/PCL/FAK Disrupted tumor angiogenesis (in vivo) Reduced the growth of CCA cells (in vivo) Decreased CD31-positive vessel content Decreased macrophage recruitment | [41] |
| HeLa cervical cancer cells | IC50 = 26.42 µM Exhibited cytotoxic effects Reduced cell viability | [35] |
| Human prostate cancer cells DU-145 | Inhibited prostate cancer growth Increased p53 protein expression Inhibited carcinoma development Decreased p21 and p27 protein expression | [94] |
| Molecular docking In silico Approach (ADMET screening) Human papillomavirus type (HPV-18 E1) protein (PDB ID: 1R9W) MD simulations | Showed high binding affinity: docking score of −8.7 kJ/mol Inhibited HPV-18 (1R9W protein) Showed high stability inside the binding pocket of 1R9W | [40] |
| Cervical cancer cells (HeLa) and Vero cells (L-929) In silico molecular docking | HeLa cells: IC50 = 11.58 µg/mL Vero cells: IC50 = 173.8 µg/mL Showed great in silico molecular docking activity Exhibited in vitro anticancer activity Exhibited potent cytotoxic activity at a concentration of 100 μg/mL Strongly occupied the active location of 3F81 Exhibited substantial interaction energy values against the protein target vaccinia H1-related (VHR) phosphatise Reduced the proliferation of cervical cancer cells (RCSB PDB ID: 3F81) | [7] |
4. Anti-Osteoarthritis Effects
| Experimental Approaches | Key Results | References |
|---|---|---|
| Experimental rabbit osteoarthritis (OA) model induced by anterior cruciate ligament transection (ACLT) | Reduced cartilage damage progression Inhibited cartilage degradation Down-regulated MMP-1/MMP-3/MMP-13 expression Up-regulated TIMP-1 Regulated the balance between MMPs and TIMPs | [44] |
| Monosodium-iodoacetate (MIA)-induced rat model of OA | Significantly corrected the OA lesions Enhanced cartilage repair and regeneration Provided a suitable milieu in situ Aided in cartilage regeneration Activated the resident progenitors Enhanced tissue repair and healing | [45] |
| Interleukin-1 beta (IL-1β)-induced inflammation in rat chondrocytes | Reduced iNOS/IL-6/MMP-3/MMP-13/ADAMTS-5 expression Increased COL2A1 expression Inhibited IL-1β-induced NF-κB activation | [46] |
| A model of newborn mouse chondrocytes and human OA | Inhibited pro-inflammatory/matrix degradation mediators Decreased MMP-3/MMP-13/ADAMTS-4/PGE2 Reduced Type II collagen and aggrecan mRNA levels Counteracted the IL-1β-induced NF-κB pathway | [96] |
| Interleukin (IL)-1β-induced chondrocytes | Revealed no significant effect on the viability of ATDC5 cells Reduced IL-1β-induced ATDC5 cell damage and ferroptosis Enhanced the inhibitory effect of ferroptosis inhibitors Down-regulated SREBF2 | [47] |
| Molecular docking analysis Network pharmacological approach and methods | Retarded the development of OA Minimized the associated pathological damage Involved in cell apoptosis/PI3K/AKT/IL-17 signaling pathways/viral infection Reduced chondrocyte apoptosis Protected the synovial lining and cartilage Targeted IL-6/EGFR/MAPK8/CCND1/CASP3/ESR1/VEGFA/MYC | [97] |
| Chick chorioallantoic membrane (CAM) assay | LC50 = 87.7 µg/mL Showed anti-arthritic activity by mTORC1 signalling pathway Deactivated certain inflammatory mediators Increased cell recovery and joint health Reduced the inflammation caused in cartilages of synovial joint Inhibited the formation of the IL and TNF-α pathways | [98] |
| Network Pharmacology Approach Thin-Layer Chromatography Analysis Enrichment analysis | Exhibited potential pharmacological effect against OA Involved Toll-like receptor (TLR)/IL-6 signaling pathways | [99] |
| Molecular docking analysis Bioinformatics analysis | Exerted beneficial effects on cartilage degeneration Targeted NCOA2/PGR/PTGS1/PTGS2/RXRA/NR3C2 Involved PI3K-Akt/TNF signaling pathways | [100] |
5. Anti-Inflammatory Effects
6. Immunomodulatory Effects
7. Neuroprotective Effects
8. Antibacterial Activity of Stigmasterol
9. Antifungal Activity
10. Antioxidant Activity
11. Anti-Diabetic Effects
12. Antiparasitic Activity
13. Antiviral Activity of Stigmasterol
14. Bioaccessibility and Bioavailability
15. Toxicology and Clinical Investigations
16. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Bassols, A.; Costa, C.; Eckersall, P.D.; Osada, J.; Sabria, J.; Tibau, J. The Pig as an Animal Model for Human Pathologies: A Proteomics Perspective. Proteom. Clin. Appl. 2014, 8, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Ozleyen, A.; Tumer, T.B.; Adetunji, C.O.; El Omari, N.; Balahbib, A.; Taheri, Y.; Bouyahya, A.; Martorell, M.; Martins, N. Natural Products and Synthetic Analogs as a Source of Antitumor Drugs. Biomolecules 2019, 9, 679. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; El Omari, N.; Elmenyiy, N.; Guaouguaou, F.-E.; Balahbib, A.; Belmehdi, O.; Salhi, N.; Imtara, H.; Mrabti, H.N.; El-Shazly, M. Moroccan Antidiabetic Medicinal Plants: Ethnobotanical Studies, Phytochemical Bioactive Compounds, Preclinical Investigations, Toxicological Validations and Clinical Evidences; Challenges, Guidance and Perspectives for Future Management of Diabetes Worldwide. Trends Food Sci. Technol. 2021, 115, 147–254. [Google Scholar]
- Bouyahya, A.; Mechchate, H.; Oumeslakht, L.; Zeouk, I.; Aboulaghras, S.; Balahbib, A.; Zengin, G.; Kamal, M.A.; Gallo, M.; Montesano, D. The Role of Epigenetic Modifications in Human Cancers and the Use of Natural Compounds as Epidrugs: Mechanistic Pathways and Pharmacodynamic Actions. Biomolecules 2022, 12, 367. [Google Scholar] [CrossRef]
- Vanmierlo, T.; Bogie, J.F.; Mailleux, J.; Vanmol, J.; Lütjohann, D.; Mulder, M.; Hendriks, J.J. Plant Sterols: Friend or Foe in CNS Disorders? Prog. Lipid Res. 2015, 58, 26–39. [Google Scholar] [CrossRef]
- Chen, M.-H.; Chen, X.-J.; Wang, M.; Lin, L.-G.; Wang, Y.-T. Ophiopogon Japonicus—a Phytochemical, Ethnomedicinal and Pharmacological Review. J. Ethnopharmacol. 2016, 181, 193–213. [Google Scholar] [CrossRef]
- Ahamed, J.I.; Francy, K.; Priya, A.V.; PremaKumari, J.; Steiny, R.P.; Kamalarajan, P.; Venkatadri, B. Computational and Biological Efficacy of Stigmasterol against HeLa Cells and Vero Cells-First Time Isolated from the Ethanolic Extract of Annonamuricata Linn Leaves. J. Mol. Struct. 2022, 1252, 132186. [Google Scholar] [CrossRef]
- Ghosh, S.; Kumar, A.; Sachan, N.; Chandra, P. Evaluation of the Antidepressant-like Effect of Total Sterols Fraction and Stigmasterol Isolated from Leaves of Aegle Marmelos and Possible Mechanism(s) of Action Involved. Curr. Drug Discov. Technol. 2022, 19, 26–40. [Google Scholar] [CrossRef]
- Astuti, A.D.; Yasir, B.; Rahim, A.; Natzir, R.; Subehan, S.; Goto, K.N.; Alam, G. Isolation and Characterization of Stigmasterol and β-Sitosterol from Plectranthus Scutellarioides Var. Color Blaze Dark Star and Cytotoxicity of Its Fraction. Egypt. J. Chem. 2022, 65, 1–2. [Google Scholar] [CrossRef]
- Ayele, T.T.; Gurmessa, G.T.; Abdissa, Z.; Kenasa, G.; Abdissa, N. Oleanane and Stigmasterol-Type Triterpenoid Derivatives from the Stem Bark of Albizia Gummifera and Their Antibacterial Activities. J. Chem. 2022, 2022, 9003143. [Google Scholar] [CrossRef]
- Muhammad, A.A.; Hassan, H.S.; Sani, Y.M.; Jimoh, A.A.; Bakare, L.O.; Sadam, A.A. Isolation and Characterization of Lupeol and Stigmasterol from Methanol Root Extract of Combretum Hypopolinum (Diels.) Okafor (Combretaceae). J. Pharm. Allied Sci. 2021, 18, 3547–3553. [Google Scholar]
- Arora, M.; Kalia, A.N. Isolation and Characterization of Stigmasterol and β-Sitosterol-D-Glycoside from Ethanolic Extract of the Stems of Salvadora Persica Linn. Int. J. Pharm. Pharm. Sci. 2013, 5, 245–249. [Google Scholar]
- Padmasri, G.; Sarada, D.V.L. Isolation and Characterization of Stigmasterol and β-Sitosterol from Acacia nilotica (L.) Delile ssp. Indica (Benth.) Brenan. J. Pharm. Res. 2011, 4, 3601–3602. [Google Scholar]
- Yusuf, A.J.; Abdullahi, M.I.; Haruna, A.K.; Idris, A.Y.; Musa, A.M. Isolation and Characterization of Stigmasterol and Bis-(5, 7-Diacetyl-Catechin-4’-α-Rhamnopyranoside) from the Stem Bark of Neocarya Macrophylla (Sabine) Prance (Chrysobalanaceae). Niger. J. Basic Appl. Sci. 2015, 23, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Michael, H.; Arora, S.; Harkonen, P.L.; Kumar, S. Studies on Correlation of Antimutagenic and Antiproliferative Activities of Juglans regia L. J. Environ. Pathol. Toxicol. Oncol. 2003, 22, 57–66. [Google Scholar] [CrossRef]
- Kamboj, A.; Saluja, A.K. Isolation of Stigmasterol and β-Sitosterol from Petroleum Ether Extract of Aerial Parts of Ageratum Conyzoides (Asteraceae). Int. J. Pharm. Pharm. Sci. 2011, 3, 94–96. [Google Scholar]
- Mailafiya, M.M.; Yusuf, A.J.; Abdullahi, M.I.; Aleku, G.A.; Ibrahim, I.A.; Yahaya, M.; Abubakar, H.; Sanusi, A.; Adamu, H.W.; Alebiosu, C.O. Antimicrobial Activity of Stigmasterol from the Stem Bark of Neocarya Macrophylla. J. Med. Plants Econ. Dev. 2018, 2, 1–5. [Google Scholar]
- Khan, M.E.; Odokpe, A.U.; Tor-Anyiin, T.A. Isolation and Characterization of Stigmasterol and B-Sitosterol from Cassia Sieberiana (Fabaceae) Leaf Extract. J. Chem. Soc. Niger. 2020, 45, 135–142. [Google Scholar]
- Gomes, A.; Saha, A.; Chatterjee, I.; Chakravarty, A.K. Viper and Cobra Venom Neutralization by β-Sitosterol and Stigmasterol Isolated from the Root Extract of Pluchea Indica Less.(Asteraceae). Phytomedicine 2007, 14, 637–643. [Google Scholar] [CrossRef]
- Ali, H.; Dixit, S.; Ali, D.; Alqahtani, S.M.; Alkahtani, S.; Alarifi, S. Isolation and Evaluation of Anticancer Efficacy of Stigmasterol in a Mouse Model of DMBA-Induced Skin Carcinoma. Drug Des. Dev. Ther. 2015, 9, 2793. [Google Scholar] [CrossRef]
- Alawode, T.T.; Lajide, L.; Olaleye, M.; Owolabi, B. Stigmasterol and β-Sitosterol: Antimicrobial Compounds in the Leaves of Icacina Trichantha Identified by GC–MS. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 1–8. [Google Scholar] [CrossRef]
- Fidan, H.; Stefanova, G.; Kostova, I.; Stankov, S.; Damyanova, S.; Stoyanova, A.; Zheljazkov, V.D. Chemical Composition and Antimicrobial Activity of Laurus nobilis L. Essential Oils from Bulgaria. Molecules 2019, 24, 804. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Anwar, F.; Ashraf, M.; Uddin, M.K. Characterization of High-Value Bioactives in Some Selected Varieties of Pakistani Rice (Oryza sativa L.). Int. J. Mol. Sci. 2012, 13, 4608–4622. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Chaudhary, J.; Jain, A.; Kishore, L. Stigmasterol: A Comprehensive Review. Int. J. Pharm. Sci. Res. 2011, 2, 2259. [Google Scholar]
- Idowu, T.O.; Ogundaini, A.O.; Adesanya, S.A.; Onawunmi, G.O.; Osungunna, M.O.; Obuotor, E.M.; Abegaz, B.M. Isolation and Characterization of Chemical Constituents from Chrysophyllum Albidum g. Don-Holl. Stem-Bark Extracts and Their Antioxidant and Antibacterial Properties. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 182–189. [Google Scholar] [CrossRef]
- Khanam, S.; Sultana, R. Isolation of β-Sitosterol & Stigmasterol as Active Immunomodulatory Constituents from Fruits of Solanum Xanthocarpum (Solanaceae). IJPSR 2012, 3, 1057–1060. [Google Scholar]
- Cabral, C.E.; Klein, M.R.S.T. Phytosterols in the Treatment of Hypercholesterolemia and Prevention of Cardiovascular Diseases. Arq. Bras. Cardiol. 2017, 109, 475–482. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Medina-Pastor, P.; Triacchini, G. The 2018 European Union Report on Pesticide Residues in Food. EFSA J. 2020, 18, e06057. [Google Scholar]
- Ashraf, R.; Bhatti, H.N. Stigmasterol. In A Centum of Valuable Plant Bioactives; Elsevier: Lahore, Pakistan, 2021; pp. 213–232. [Google Scholar]
- Nualkaew, S.; Padee, P.; Talubmook, C. Hypoglycemic Activity in Diabetic Rats of Stigmasterol and Sitosterol-3-O--D-Glucopyranoside Isolated from Pseuderanthemum Palatiferum (Nees) Radlk. Leaf Extract. J. Med. Plants Res. 2015, 9, 629–635. [Google Scholar]
- Dong, Y.; Chen, C.; Zhang, C.; Zhang, L.; Zhang, Y.; Li, Y.; Dong, Z. Stigmasterol Inhibits the Progression of Lung Cancer by Regulating Retinoic Acid-Related Orphan Receptor C. Histol. Histopathol. 2021, 36, 18388. [Google Scholar]
- Kim, Y.-S.; Li, X.-F.; Kang, K.-H.; Ryu, B.; Kim, S.K. Stigmasterol Isolated from Marine Microalgae Navicula Incerta Induces Apoptosis in Human Hepatoma HepG2 Cells. BMB Rep. 2014, 47, 433. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.M.; Overbeek-Wager, E.; Grumbo, R.J. Diagnosis and Management of Endometrial Cancer. Am. Fam. Physician 2016, 93, 468–474. [Google Scholar] [PubMed]
- Farshori, N.N.; Al-Oqail, M.M.; Al-Sheddi, E.S.; Al-Massarani, S.M.; Alam, P.; Siddiqui, M.A.; Ahmad, J.; Al-Khedhairy, A.A. HPTLC Estimation and Anticancer Potential of Aloe Perryi Petroleum Ether Extract (APPeE): A Mechanistic Study on Human Breast Cancer Cells (MDA-MB-231). J. King Saud Univ. Sci. 2022, 34, 101968. [Google Scholar] [CrossRef]
- Kurniasih, N.; Supriadin, A.; Harneti, D.; Abdulah, R.; Taib, M.N.A.M.; Supratman, U. Ergosterol Peroxide and Stigmasterol from the Stembark of Aglaia Simplicifolia (Meliaceae) and Their Cytotoxic against HeLa Cervical Cancer Cell Lines. J. Kim. Val. 2021, 7, 46–51. [Google Scholar] [CrossRef]
- Liao, H.; Zhu, D.; Bai, M.; Chen, H.; Yan, S.; Yu, J.; Zhu, H.; Zheng, W.; Fan, G. Stigmasterol Sensitizes Endometrial Cancer Cells to Chemotherapy by Repressing Nrf2 Signal Pathway. Cancer Cell Int. 2020, 20, 1–10. [Google Scholar]
- Mabuchi, S.; Kuroda, H.; Takahashi, R.; Sasano, T. The PI3K/AKT/MTOR Pathway as a Therapeutic Target in Ovarian Cancer. Gynecol. Oncol. 2015, 137, 173–179. [Google Scholar] [CrossRef]
- Omran, G.; Abd-Alhaseeb, M.; Houssen, M. The Anti-Tumor Effect of Stigmasterol on Sorafenib Treated Human Breast Cancer Cell Lines. Res. Sq. 2021, 1–14. [Google Scholar] [CrossRef]
- Araldi, R.P.; Sant’Ana, T.A.; Módolo, D.G.; De Melo, T.C.; Spadacci-Morena, D.D.; De Cassia Stocco, R.; Cerutti, J.M.; De Souza, E.B. The Human Papillomavirus (HPV)-Related Cancer Biology: An Overview. Biomed. Pharmacother. 2018, 106, 1537–1556. [Google Scholar] [CrossRef]
- Salaria, D.; Rolta, R.; Mehta, J.; Awofisayo, O.; Fadare, O.A.; Kaur, B.; Kumar, B.; Da Costa, R.A.; Chandel, S.R.; Kaushik, N. Phytoconstituents of Traditional Himalayan Herbs as Potential Inhibitors of Human Papillomavirus (HPV-18) for Cervical Cancer Treatment: An In Silico Approach. PLoS ONE 2022, 17, e0265420. [Google Scholar] [CrossRef]
- Kangsamaksin, T.; Chaithongyot, S.; Wootthichairangsan, C.; Hanchaina, R.; Tangshewinsirikul, C.; Svasti, J. Lupeol and Stigmasterol Suppress Tumor Angiogenesis and Inhibit Cholangiocarcinoma Growth in Mice via Downregulation of Tumor Necrosis Factor-α. PLoS ONE 2017, 12, e0189628. [Google Scholar] [CrossRef]
- Spriha, S.E.; Rahman, S.A. In Silico Evaluation of Selected Compounds from Bergenia Ciliata (Haw.) Sternb against Molecular Targets of Breast Cancer. Indian J. Pharm. Educ. Res. 2022, 56, S105–S114. [Google Scholar] [CrossRef]
- Sacitharan, P.K. Ageing and Osteoarthritis. Biochem. Cell Biol. Ageing Part II Clin. Sci. 2019, 91, 123–159. [Google Scholar]
- Chen, W.-P.; Yu, C.; Hu, P.-F.; Bao, J.-P.; Tang, J.-L.; Wu, L.-D. Stigmasterol Blocks Cartilage Degradation in Rabbit Model of Osteoarthritis. Acta Biochim. Pol. 2012, 59, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Sampath, S.J.P.; Kotikalapudi, N.; Venkatesan, V. A Novel Therapeutic Combination of Mesenchymal Stem Cells and Stigmasterol to Attenuate Osteoarthritis in Rodent Model System—A Proof of Concept Study. Stem Cell Investig. 2021, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Sampath, S.J.P.; Rath, S.N.; Kotikalapudi, N.; Venkatesan, V. Beneficial Effects of Secretome Derived from Mesenchymal Stem Cells with Stigmasterol to Negate IL-1β-Induced Inflammation in-Vitro Using Rat Chondrocytes—OA Management. Inflammopharmacology 2021, 29, 1701–1717. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.; Xu, P.; Li, H. Stigmasterol Alleviates Interleukin-1beta-Induced Chondrocyte Injury by down-Regulatingsterol Regulatory Element Binding Transcription Factor 2 to Regulateferroptosis. Bioengineered 2021, 12, 9332–9340. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, D.K.; Kanui, T.I.; Mbugua, P.M.; Githinji, C.G. Analgesic and Anti-Inflammatory Activities of 9-Hexacosene and Stigmasterol Isolated from Mondia Whytei. Phytopharmacology 2012, 2, 212–223. [Google Scholar]
- Antwi, A.O.; Obiri, D.D.; Osafo, N.; Essel, L.B.; Forkuo, A.D.; Atobiga, C. Stigmasterol Alleviates Cutaneous Allergic Responses in Rodents. BioMed Res. Int. 2018, 2018, 3984068. [Google Scholar] [CrossRef]
- Morais, M.G.; Saldanha, A.A.; Azevedo, L.S.; Mendes, I.C.; Rodrigues, J.P.C.; Amado, P.A.; De Souza Farias, K.; Zanuncio, V.S.S.; Cassemiro, N.S.; Da Silva, D.B. Antioxidant and Anti-Inflammatory Effects of Fractions from Ripe Fruits of Solanum Lycocarpum St. Hil.(Solanaceae) and Putative Identification of Bioactive Compounds by GC–MS and LC-DAD-MS. Food Res. Int. 2022, 156, 111145. [Google Scholar] [CrossRef]
- Morgan, L.V.; Petry, F.; Scatolin, M.; De Oliveira, P.V.; Alves, B.O.; Zilli, G.A.L.; Volfe, C.R.B.; Oltramari, A.R.; De Oliveira, D.; Scapinello, J. Investigation of the Anti-Inflammatory Effects of Stigmasterol in Mice: Insight into Its Mechanism of Action. Behav. Pharmacol. 2021, 32, 640–651. [Google Scholar] [CrossRef]
- Olajubutu, O.G.; Ogunremi, B.I.; Adewole, A.H.; Awotuya, O.I.; Fakola, E.G.; Anyim, G.; Faloye, K.O. Topical Anti-Inflammatory Activity of Petiveria Alliacea, Chemical Profiling and Computational Investigation of Phytoconstituents Identified from Its Active Fraction. Chem. Afr. 2022, 5, 557–565. [Google Scholar] [CrossRef]
- Wen, S.; He, L.; Zhong, Z.; Zhao, R.; Weng, S.; Mi, H.; Liu, F. Stigmasterol Restores the Balance of Treg/Th17 Cells by Activating the Butyrate-PPARγ Axis in Colitis. Front. Immunol. 2021, 12, 4201. [Google Scholar] [CrossRef] [PubMed]
- Ralf, K.; Gabriel, B.; Ermilo, A.-A.V.; Martha, M.-G.; Mirbella, C.-F.; Rocio, B.-A. Anti-Inflammatory and Immunomodulatory Effects of Critonia Aromatisans Leaves: Downregulation of pro-Inflammatory Cytokines. J. Ethnopharmacol. 2016, 190, 174–182. [Google Scholar]
- Sharif, M.; Anjum, I.; Shabbir, A.; Mushtaq, M.N. Immunomodulatory and Anti-Inflammatory Effects of Aerva Lanata in Ovalbumin Induced Allergic Asthmatic Mice. J. Ethnopharmacol. 2022, 289, 115087. [Google Scholar] [CrossRef] [PubMed]
- Xiaoyue, S.; Yanbin, L. Myasthenia Gravis: The Pharmacological Basis of Traditional Chinese Medicine for Its Clinical Application. BioFactors 2022, 48, 228–238. [Google Scholar] [CrossRef]
- Li, H.-L.; Zhou, J.-P.; Deng, J.-M. Therapeutic Mechanism of Xiaoqinglong Decoction against COVID-19 Based on Network Pharmacology and Molecular Docking Technology. Comb. Chem. High Throughput Screen. 2022. [Google Scholar] [CrossRef]
- Soares, J.J.; Rodrigues, D.T.; Gonçalves, M.B.; Lemos, M.C.; Gallarreta, M.S.; Bianchini, M.C.; Gayer, M.C.; Puntel, R.L.; Roehrs, R.; Denardin, E.L. Paraquat Exposure-Induced Parkinson’s Disease-like Symptoms and Oxidative Stress in Drosophila Melanogaster: Neuroprotective Effect of Bougainvillea Glabra Choisy. Biomed. Pharmacother. 2017, 95, 245–251. [Google Scholar] [CrossRef]
- Yadav, M.; Parle, M.; Jindal, D.K.; Dhingra, S. Protective Effects of Stigmasterol against Ketamine-Induced Psychotic Symptoms: Possible Behavioral, Biochemical and Histopathological Changes in Mice. Pharmacol. Rep. 2018, 70, 591–599. [Google Scholar] [CrossRef]
- Pattarachotanant, N.; Prasansuklab, A.; Tencomnao, T. Momordica charantia L. Extract Protects Hippocampal Neuronal Cells against PAHs-Induced Neurotoxicity: Possible Active Constituents Include Stigmasterol and Vitamin E. Nutrients 2021, 13, 2368. [Google Scholar] [CrossRef]
- Pratiwi, R.; Nantasenamat, C.; Ruankham, W.; Suwanjang, W.; Prachayasittikul, V.; Prachayasittikul, S.; Phopin, K. Mechanisms and Neuroprotective Activities of Stigmasterol against Oxidative Stress-Induced Neuronal Cell Death via Sirtuin Family. Front. Nutr. 2021, 8, 648995. [Google Scholar] [CrossRef]
- Haque, M.N.; Hannan, M.A.; Dash, R.; Choi, S.M.; Moon, I.S. The Potential LXRβ Agonist Stigmasterol Protects against Hypoxia/Reoxygenation Injury by Modulating Mitophagy in Primary Hippocampal Neurons. Phytomedicine 2021, 81, 153415. [Google Scholar] [CrossRef] [PubMed]
- Adebiyi, O.E.; Olopade, J.O.; Olayemi, F.O. Neuroprotective Effect of Grewia Carpinifolia Extract against Vanadium Induced Behavioural Impairment. Folia Vet. 2016, 60, 5–13. [Google Scholar] [CrossRef]
- Kumar, G.; Mukherjee, S.; Patnaik, R. Identification of Withanolide-M and Stigmasterol as Potent Neuroprotectant and Dual Inhibitor of Inducible/Neuronal Nitric Oxide Synthase by Structure-Based Virtual Screening. J. Biol. Eng. Res. Rev. 2017, 4, 9–13. [Google Scholar]
- Brimson, J.M.; Brimson, S.J.; Brimson, C.A.; Rakkhitawatthana, V.; Tencomnao, T. Rhinacanthus Nasutus Extracts Prevent Glutamate and Amyloid-β Neurotoxicity in HT-22 Mouse Hippocampal Cells: Possible Active Compounds Include Lupeol, Stigmasterol and β-Sitosterol. Int. J. Mol. Sci. 2012, 13, 5074–5097. [Google Scholar] [CrossRef] [PubMed]
- Burg, V.K.; Grimm, H.S.; Rothhaar, T.L.; Grösgen, S.; Hundsdörfer, B.; Haupenthal, V.J.; Zimmer, V.C.; Mett, J.; Weingärtner, O.; Laufs, U. Plant Sterols the Better Cholesterol in Alzheimer’s Disease? A Mechanistical Study. J. Neurosci. 2013, 33, 16072–16087. [Google Scholar] [CrossRef]
- Ibrahim, N.; Yaacob, W.A. Transcriptome Analysis of Methicillin-Resistant Staphylococcus Aureus in Response to Stigmasterol and Lupeol. J. Glob. Antimicrob. Resist. 2017, 8, 48–54. [Google Scholar]
- Odiba, J.; Musa, A.; Hassan, H.; Yahay, S.; Okolo, E. Antimicrobial Activity of Isolated Stigmast-5-En-3-β-Ol (β-Sitosterol) from Honeybee Propolis from North-Western, Nigeria. Int. J. Pharm. Sci. Res. 2014, 5, 908–918. [Google Scholar]
- Kanokmedhakul, K.; Kanokmedhakul, S.; Phatchana, R. Biological Activity of Anthraquinones and Triterpenoids from Prismatomeris Fragrans. J. Ethnopharmacol. 2005, 100, 284–288. [Google Scholar] [CrossRef]
- Correa, G.; Abreu, V.D.C.; Martins, D.; Takahashi, J.A.; Fontoura, H.D.S.; Cara, D.C.; Pilo-Veloso, D.; Alcântara, A.F.D.C. Anti-Inflammatory and Antimicrobial Activities of Steroids and Triterpenes Isolated from Aerial Parts of Justicia Acuminatissima (Acanthaceae). Int. J. Pharm. Pharm. Sci. 2014, 6, 75–81. [Google Scholar]
- Yinusa, I.; George, N.I.; Shuaibu, U.O.A.; Ayo, R.G. Bioactivity of Stigmasterol Isolated from the Aerial Part of Spillanthes Acmella (Murr) on Selected Microorganism. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 475–479. [Google Scholar]
- Mbambo, B.; Odhav, B.; Mohanlall, V. Antifungal Activity of Stigmasterol, Sitosterol and Ergosterol from Bulbine Natalensis Baker (Asphodelaceae). J. Med. Plants Res. 2012, 6, 5135–5141. [Google Scholar]
- Aminu, R.; Umar, I.A.; Rahman, M.A.; Ibrahim, M.A. Stigmasterol Retards the Proliferation and Pathological Features of Trypanosoma Congolense Infection in Rats and Inhibits Trypanosomal Sialidase in Vitro and in Silico. Biomed. Pharmacother. 2017, 89, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, R.; Shayanfar, P.; Maniati, M. In Vitro Antileishmanial Effects of Saffron Compounds, Crocin and Stigmasterol, on Iranian Strain of Leishmania Major (MHOM/IR/75/ER). Iran. J. Parasitol. 2021, 16, 151. [Google Scholar] [CrossRef] [PubMed]
- De-Eknamkul, W.; Potduang, B. Biosynthesis of β-Sitosterol and Stigmasterol in Croton Sublyratus Proceeds via a Mixed Origin of Isoprene Units. Phytochemistry 2003, 62, 389–398. [Google Scholar] [CrossRef]
- Gao, J.J.; Cheng, D.L.; Liu, X.P. Chemical Constituents of Emilia Sonchifolia L. DC. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Medica 1993, 18, 102–103. [Google Scholar]
- Han, Y.; Xia, C.; Cheng, X.; Xiang, R.; Liu, H.; Yan, Q.; Xu, D. Preliminary Studies on Chemical Constituents and Pharmacological Action of Eclipta prostrata L. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Medica 1998, 23, 680–703. [Google Scholar]
- Jamaluddin, F.; Mohamed, S.; Lajis, M.N. Hypoglycaemic Effect of Parkia Speciosa Seeds Due to the Synergistic Action of β-Sitosterol and Stigmasterol. Food Chem. 1994, 49, 339–345. [Google Scholar] [CrossRef]
- Li, C.; Bu, P.-B.; Yue, D.-K.; Sun, Y. Chemical Constituents from Roots of Ficus Hirta. China J. Chin. Mater. Med. 2006, 31, 131–133. [Google Scholar]
- Li, X.L.; Wang, H.; Liu, G.; Zhang, X.Q.; Ye, W.C.; Zhao, S.X. Study on Chemical Constituents from Desmodium Styracifolium. Zhong Yao Cai Zhongyaocai J. Chin. Med. Mater. 2007, 30, 802–805. [Google Scholar]
- Liu, G.; Zheng, J.; Yu, Z.; Zhang, J.; Lin, R. Study on Sterols and Triterpenes from the Stems of Akebia Quinata. Zhong Yao Cai Zhongyaocai J. Chin. Med. Mater. 2005, 28, 1060–1062. [Google Scholar]
- Peng, T.; Dong, X.; Deng, Y.; Tu, Y.; Li, X. Research on Chemical Ingredients of the Root from Cultivar Aralia Cordata Thunb. Zhong Yao Cai Zhongyaocai J. Chin. Med. Mater. 2005, 28, 996–998. [Google Scholar]
- Stevens, M.M.; Honerkamp-Smith, A.R.; Keller, S.L. Solubility Limits of Cholesterol, Lanosterol, Ergosterol, Stigmasterol, and β-Sitosterol in Electroformed Lipid Vesicles. Soft Matter 2010, 6, 5882–5890. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhong, Y.; Luo, H.; Ding, X.; Zuo, C. Studies on Chemical Constituents of the Roots of Gypsophila Oldhamiana Miq. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Med. 1999, 24, 680–681. [Google Scholar]
- Yang, X.-W.; Guo, Q.-M. Studies on Chemical Constituents in Fruits of Eucalyptus Globulus. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Medica 2007, 32, 496–500. [Google Scholar]
- Zhang, M.; Chen, Y. Chemical Constituents of Eclipta alba (L.) Hassk. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Med. 1996, 21, 480–481. [Google Scholar]
- Bae, H.; Song, G.; Lim, W. Stigmasterol Causes Ovarian Cancer Cell Apoptosis by Inducing Endoplasmic Reticulum and Mitochondrial Dysfunction. Pharmaceutics 2020, 12, 488. [Google Scholar] [CrossRef]
- Li, M.; Zhang, W.; Yang, L.; Wang, H.; Wang, Y.; Huang, K.; Zhang, W. The Mechanism of Xiaoyao San in the Treatment of Ovarian Cancer by Network Pharmacology and the Effect of Stigmasterol on the PI3K/Akt Pathway. Dis. Markers 2021, 2021, 4304507. [Google Scholar] [CrossRef]
- Li, K.; Yuan, D.; Yan, R.; Meng, L.; Zhang, Y.; Zhu, K. Stigmasterol Exhibits Potent Antitumor Effects in Human Gastric Cancer Cells Mediated via Inhibition of Cell Migration, Cell Cycle Arrest, Mitochondrial Mediated Apoptosis and Inhibition of JAK/STAT Signalling Pathway. J. BUON 2018, 23, 1420–1425. [Google Scholar]
- Alzahrani, A.S. PI3K/Akt/MTOR Inhibitors in Cancer: At the Bench and Bedside. In Proceedings of the Seminars in Cancer Biology; Elsevier: Alpharetta, GA, USA, 2019; Volume 59, pp. 125–132. [Google Scholar]
- Zhao, H.; Zhang, X.; Wang, M.; Lin, Y.; Zhou, S. Stigmasterol Simultaneously Induces Apoptosis and Protective Autophagy by Inhibiting Akt/MTOR Pathway in Gastric Cancer Cells. Front. Oncol. 2021, 11, 629008. [Google Scholar] [CrossRef]
- Pandey, P.; Bajpai, P.; Siddiqui, M.H.; Sayyed, U.; Tiwari, R.; Shekh, R.; Mishra, K.; Kapoor, V.K. Elucidation of the Chemopreventive Role of Stigmasterol against Jab1 in Gall Bladder Carcinoma. Endocr. Metab. Immune Disord. Drug Targets Former. Curr. Drug Targets Immune Endocr. Metab. Disord. 2019, 19, 826–837. [Google Scholar] [CrossRef]
- Ghosh, T.; Maity, T.K.; Singh, J. Evaluation of Antitumor Activity of Stigmasterol, a Constituent Isolated from Bacopa Monnieri Linn Aerial Parts against Ehrlich Ascites Carcinoma in Mice. Orient. Pharm. Exp. Med. 2011, 11, 41–49. [Google Scholar] [CrossRef]
- Scholtysek, C.; Krukiewicz, A.A.; Sharma, K.P.; Sharma, P.C.; Goldmann, W.H. Characterizing Components of the Saw Palmetto Berry Extract (SPBE) on Prostate Cancer Cell Growth and Traction. Biochem. Biophys. Res. Commun. 2009, 379, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, X.; Liu, J.; Pan, X.; Zhao, Q. Stigmasterol Exerts Neuro-Protective Effect against Ischemic/Reperfusion Injury through Reduction of Oxidative Stress and Inactivation of Autophagy. Neuropsychiatr. Dis. Treat. 2019, 15, 2991. [Google Scholar] [CrossRef] [PubMed]
- Gabay, O.; Sanchez, C.; Salvat, C.; Chevy, F.; Breton, M.; Nourissat, G.; Wolf, C.; Jacques, C.; Berenbaum, F. Stigmasterol: A Phytosterol with Potential Anti-Osteoarthritic Properties. Osteoarthr. Cartil. 2010, 18, 106–116. [Google Scholar] [CrossRef]
- Zhang, L.; Shi, X.; Huang, Z.; Mao, J.; Mei, W.; Ding, L.; Zhang, L.; Xing, R.; Wang, P. Network Pharmacology Approach to Uncover the Mechanism Governing the Effect of Radix Achyranthis Bidentatae on Osteoarthritis. BMC Complement. Med. Ther. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Gowri, K.; Chitra, V. In-Vitro and Ex-Vivo Evaluation of Plant Sterol in Osteoarthritis by CAM Assay. J. Med. Pharm. Allied Sci. 1899, 11, 4288–4293. [Google Scholar]
- Yu, M.-X.; Ma, X.-Q.; Song, X.; Huang, Y.-M.; Jiang, H.-T.; Wang, J.; Yang, W.-H. Validation of the Key Active Ingredients and Anti-Inflammatory and Analgesic Effects of Shenjin Huoxue Mixture against Osteoarthritis by Integrating Network Pharmacology Approach and Thin-Layer Chromatography Analysis. Drug Des. Dev. Ther. 2020, 14, 1145. [Google Scholar] [CrossRef]
- Chen, W.; Lin, T.; He, Q.; Yang, P.; Zhang, G.; Huang, F.; Wang, Z.; Peng, H.; Li, B.; Liang, D. Study on the Potential Active Components and Molecular Mechanism of Xiao Huoluo Pills in the Treatment of Cartilage Degeneration of Knee Osteoarthritis Based on Bioinformatics Analysis and Molecular Docking Technology. J. Orthop. Surg. 2021, 16, 1–19. [Google Scholar] [CrossRef]
- Khan, M.A.; Sarwar, A.H.M.G.; Rahat, R.; Ahmed, R.S.; Umar, S. Stigmasterol Protects Rats from Collagen Induced Arthritis by Inhibiting Proinflammatory Cytokines. Int. Immunopharmacol. 2020, 85, 106642. [Google Scholar] [CrossRef]
- He, D.; Wang, S.; Fang, G.; Zhu, Q.; Wu, J.; Li, J.; Shi, D.; Lian, X. LXRs/ABCA1 Activation Contribute to the Anti-Inflammatory Role of Phytosterols on LPS-Induced Acute Lung Injury. J. Funct. Foods 2022, 89, 104966. [Google Scholar] [CrossRef]
- Antwi, A.O.; Obiri, D.D.; Osafo, N. Stigmasterol Modulates Allergic Airway Inflammation in Guinea Pig Model of Ovalbumin-Induced Asthma. Mediat. Inflamm. 2017, 2017, 2953930. [Google Scholar] [CrossRef] [PubMed]
- Le, C.-F.; Kailaivasan, T.H.; Chow, S.-C.; Abdullah, Z.; Ling, S.-K.; Fang, C.-M. Phytosterols Isolated from Clinacanthus Nutans Induce Immunosuppressive Activity in Murine Cells. Int. Immunopharmacol. 2017, 44, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Karim, N.; Khan, I.; Abdelhalim, A.; Halim, S.A.; Khan, A.; Al-Harrasi, A. Stigmasterol Can Be New Steroidal Drug for Neurological Disorders: Evidence of the GABAergic Mechanism via Receptor Modulation. Phytomedicine 2021, 90, 153646. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Weon, J.B.; Ma, C.J. Neuroprotective Activity of Phytosterols Isolated from Artemisia Apiacea. Korean J. Pharmacogn. 2014, 45, 214–219. [Google Scholar]
- Tan, M.A.; Gonzalez, S.J.B.; Alejandro, G.J.D.; An, S.S.A. Neuroprotective Effects of Vomifoliol, Isolated from Tarenna Obtusifolia Merr.(Rubiaceae), against Amyloid-Beta1-42-Treated Neuroblastoma SH-SY5Y Cells. 3 Biotech 2020, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Adebiyi, O.E.; Olopade, J.O.; Olayemi, F.O. Sodium Metavanadate Induced Cognitive Decline, Behavioral Impairments, Oxidative Stress and down Regulation of Myelin Basic Protein in Mice Hippocampus: Ameliorative Roles of β-Spinasterol, and Stigmasterol. Brain Behav. 2018, 8, e01014. [Google Scholar] [CrossRef]
- Tamokou, J.D.D.; Kuiate, J.R.; Tene, M.; Nwemeguela, T.J.K.; Tane, P. The Antimicrobial Activities of Extract and Compounds Isolated from Brillantaisia Lamium. Iran. J. Med. Sci. 2011, 36, 24. [Google Scholar]
- Edilu, A.; Adane, L.; Woyessa, D. In Vitro Antibacterial Activities of Compounds Isolated from Roots of Caylusea Abyssinica. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 1–8. [Google Scholar] [CrossRef]
- Awouafack, M.D.; McGaw, L.J.; Gottfried, S.; Mbouangouere, R.; Tane, P.; Spiteller, M.; Eloff, J.N. Antimicrobial Activity and Cytotoxicity of the Ethanol Extract, Fractions and Eight Compounds Isolated from Eriosema Robustum (Fabaceae). BMC Complement. Altern. Med. 2013, 13, 289. [Google Scholar] [CrossRef]
- Kaigongi, M.M.; Lukhoba, C.W.; Yaouba, S.; Makunga, N.P.; Githiomi, J.; Yenesew, A. In Vitro Antimicrobial and Antiproliferative Activities of the Root Bark Extract and Isolated Chemical Constituents of Zanthoxylum Paracanthum Kokwaro (Rutaceae). Plants 2020, 9, 920. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Hashem, H.A.; Khalil, R.R. Stigmasterol Treatment Increases Salt Stress Tolerance of Faba Bean Plants by Enhancing Antioxidant Systems. Plant Omics 2012, 5, 476–485. [Google Scholar]
- Osuntokun, O.T.; Oluduro, A.O.; Idowu, T.O.; Omotuyi, A.O. Assessment of Nephrotoxicity, Anti-Inflammatory and Antioxidant Properties of Epigallocatechin, Epicatechin and Stigmasterol Phytosterol (Synergy) Derived from Ethyl Acetate Stem Bark Extract of Spondias Mombin on Wistar Rats Using Molecular Method of Ana. J. Mol. Microbiol. 2017, 1, 103. [Google Scholar]
- Liang, Q.; Yang, J.; He, J.; Chen, X.; Zhang, H.; Jia, M.; Liu, K.; Jia, C.; Pan, Y.; Wei, J. Stigmasterol Alleviates Cerebral Ischemia/Reperfusion Injury by Attenuating Inflammation and Improving Antioxidant Defenses in Rats. Biosci. Rep. 2020, 40, BSR20192133. [Google Scholar] [CrossRef]
- Shanthakumar, B.; Sathish, M.; Suresh, A.J. In Vitro Antioxidant Activity of Extracts and Stigmasterol from Leaves of Clerodendrum Inerme Linn. Res. J. Pharm. Biol. Chem. Sci. 2013, 4, 1411–1418. [Google Scholar]
- Wang, J.; Huang, M.; Yang, J.; Ma, X.; Zheng, S.; Deng, S.; Huang, Y.; Yang, X.; Zhao, P. Anti-Diabetic Activity of Stigmasterol from Soybean Oil by Targeting the GLUT4 Glucose Transporter. Food Nutr. Res. 2017, 61, 1364117. [Google Scholar] [CrossRef] [PubMed]
- Ramu, R.; Shirahatti, P.S.; Nayakavadi, S.; Vadivelan, R.; Zameer, F.; Dhananjaya, B.L.; Prasad, N. The Effect of a Plant Extract Enriched in Stigmasterol and β-Sitosterol on Glycaemic Status and Glucose Metabolism in Alloxan-Induced Diabetic Rats. Food Funct. 2016, 7, 3999–4011. [Google Scholar] [CrossRef]
- Panda, S.; Jafri, M.; Kar, A.; Meheta, B.K. Thyroid Inhibitory, Antiperoxidative and Hypoglycemic Effects of Stigmasterol Isolated from Butea Monosperma. Fitoterapia 2009, 80, 123–126. [Google Scholar] [CrossRef]
- Ward, M.G.; Li, G.; Barbosa-Lorenzi, V.C.; Hao, M. Stigmasterol Prevents Glucolipotoxicity Induced Defects in Glucose-Stimulated Insulin Secretion. Sci. Rep. 2017, 7, 9536. [Google Scholar] [CrossRef] [Green Version]
- Reza, M.S.; Shuvo, M.S.R.; Hassan, M.M.; Basher, M.A.; Islam, M.A.; Naznin, N.E.; Jafrin, S.; Ahmed, K.S.; Hossain, H.; Daula, A.F.M.S.U. Antidiabetic and Hepatoprotective Potential of Whole Plant Extract and Isolated Compounds of Aeginetia Indica. Biomed. Pharmacother. 2020, 132, 110942. [Google Scholar] [CrossRef]
- Soekamto, N.H.; Ahmad, F.; Appa, F.E. Potential of Stigmasterol from EtOAc Extract Melochia Umbellata (Houtt) Stapf Var. Visenia as Dengue Antivirus. In Proceedings of the Journal of Physics: Conference Series; IOP Publishing: Bristol, UK, 2019; Volume 1341, p. 32044. [Google Scholar]
- Petrera, E.; Níttolo, A.G.; Alché, L.E. Antiviral Action of Synthetic Stigmasterol Derivatives on Herpes Simplex Virus Replication in Nervous Cells in Vitro. BioMed Res. Int. 2014, 2014, 947560. [Google Scholar] [CrossRef]
- Dos Santos Souza Marinho, R.; Ramos, C.J.B.; Leite, J.P.G.; Teixeira, V.L.; De Palmer Paixão, I.C.N.; Belo, C.A.D.; Pereira, A.B.; Pinto, A.M.V. Antiviral Activity of 7-Keto-Stigmasterol Obtained from Green Antarctic Algae Prasiola Crispa against Equine Herpesvirus 1. J. Appl. Phycol. 2017, 29, 555–562. [Google Scholar] [CrossRef]
- Fakih, O.; Sanver, D.; Kane, D.; Thorne, J.L. Exploring the Biophysical Properties of Phytosterols in the Plasma Membrane for Novel Cancer Prevention Strategies. Biochimie 2018, 153, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Hellner, G.; Hoke, E.R.; Nagy, V.; Szakács, G.; Poppe, L. Integrated Enzymatic Production of Specific Structured Lipid and Phytosterol Ester Compositions. Process Biochem. 2010, 45, 1245–1250. [Google Scholar] [CrossRef]
- Schneider, K.; DiNovi, M.; Baines, J.; Schlatter, J. Phytosterols, Phytostanols and Their Esters. Saf. Eval. Certain Food Addit. 2009, 117, 3–573. [Google Scholar]
- Moghadasian, M.H.; Tan, Z.; Le, K.; Shahidi, F. Anti-Atherogenic Effects of Phytosteryl Oleates in Apo-E Deficient Mice. J. Funct. Foods 2016, 21, 97–103. [Google Scholar] [CrossRef]
- Wang, S.; Yu, D.; Shi, Y.; Jiang, L.; Yang, F.; Yu, G. Investigation into the Bioavailability of Synthesized Phytosterol Esters in Vitro and in Vivo Using Caco-2 Cell Model and Wistar Rats. Food Sci. Technol. 2021, 42. [Google Scholar] [CrossRef]
- Makran, M.; Faubel, N.; López-García, G.; Cilla, A.; Barberá, R.; Alegría, A.; Garcia-Llatas, G. Sterol Bioaccessibility in a Plant Sterol-Enriched Beverage Using the INFOGEST Digestion Method: Influence of Gastric Lipase, Bile Salts and Cholesterol Esterase. Food Chem. 2022, 382, 132305. [Google Scholar] [CrossRef]
- Kritchevsky, D.; Chen, S.C. Phytosterols—Health Benefits and Potential Concerns: A Review. Nutr. Res. 2005, 25, 413–428. [Google Scholar] [CrossRef]
- Heinemann, T.; Axtmann, G.; Bergmann, K. von Comparison of Intestinal Absorption of Cholesterol with Different Plant Sterols in Man. Eur. J. Clin. Investig. 1993, 23, 827–831. [Google Scholar] [CrossRef]
- Hamada, T.; Goto, H.; Yamahira, T.; Sugawara, T.; Imaizumi, K.; Ikeda, I. Solubility in and Affinity for the Bile Salt Micelle of Plant Sterols Are Important Determinants of Their Intestinal Absorption in Rats. Lipids 2006, 41, 551–556. [Google Scholar] [CrossRef]
- EFSA Panel on Food additives and Nutrient Sources added to Food (ANS). Added to Scientific Opinion on the Safety of Stigmasterol-Rich Plant Sterols as Food Additive. EFSA J. 2012, 10, 2659. [Google Scholar] [CrossRef]
- Sioen, I.; Matthys, C.; Huybrechts, I.; Van Camp, J.; De Henauw, S. Consumption of Plant Sterols in Belgium: Consumption Patterns of Plant Sterol-Enriched Foods in Flanders, Belgium. Br. J. Nutr. 2011, 105, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Hepburn, P.A.; Horner, S.A.; Smith, M. Safety Evaluation of Phytosterol Esters. Part 2. Subchronic 90-Day Oral Toxicity Study on Phytosterol Esters–a Novel Functional Food. Food Chem. Toxicol. 1999, 37, 521–532. [Google Scholar] [CrossRef]
- Kim, J.-C.; Kang, B.-H.; Shin, C.-C.; Kim, Y.-B.; Lee, H.-S.; Kim, C.-Y.; Han, J.; Kim, K.-S.; Chung, D.-W.; Chung, M.-K. Subchronic Toxicity of Plant Sterol Esters Administered by Gavage to Sprague–Dawley Rats. Food Chem. Toxicol. 2002, 40, 1569–1580. [Google Scholar] [CrossRef]


| Experimental Approaches | Key Results | References |
|---|---|---|
| Collagen-induced arthritis (CIA) induced inflammation in rats | Significantly suppressed TNF-α/IL-6/IL-1β/Inos/COX-2 expression Increased the expression of anti-inflammatory cytokine IL-10 Down-regulated the expression of NF-kBp65/p38MAPK Inhibited p-IKB-α activation | [101] |
| IL-1β-induced inflammation in rat chondrocytes | Decreased the expression of iNOS and IL-6 | [46] |
| Dimethylbenzene-induced ear edema test | Inhibition = 50.34% at 30 mg/kg dose Inhibited ear edema Showed significant anti-inflammatory activity (p < 0.01) | [48] |
| Carrageenan-induced peritonitis and arachidonic acid-induced paw edema Molecular docking Acetic acid-induced writhing test | Decreased leukocyte infiltration in peritonitis assay Reduced paw edema elicited by arachidonic acid Prevented abdominal writhes and paw licking Reduced the number of crossings Presented anti-inflammatory effects mediated by glucocorticoid receptors | [51] |
| Dermatitis induced by 12-O-tetradecanoylphorbol-13-acetate (TPA) | Controlled inflammatory features such as ear skin oedema and neutrophilia Inhibited the expression of pro-inflammatory cytokines Reduced serum levels of TNF-α | [49] |
| Induced rat paw oedema model | Inhibition = 79.86 ± 0.16% Inhibited the production of pro-inflammatory proteins Exhibited the best binding energy (6.5 kcal/mol) against TNF-α Observed to be an effective molecule against the COX-2 enzyme Used as a topical anti-inflammatory drug Enhanced electrophilic index | [52] |
| Ovalbumin induced allergic asthmatic mice | Improved the number of inflammatory Decreased the level of inflammatory modulator TNF-α and IgE antibodies Reduced IL- 4, IL-5, and IL-13 Increased the expression levels of AQP1 and AQP5 | [55] |
| LPS-stimulated RAW264.7 cells LPS-induced acute lung injury in C57BL/6J mice | Activated the LXRs/ABCA1 signaling pathway Affected the activation of the TLR4/NF-κB pathway | [102] |
| Carrageenan-induced hind paw edema model | Displayed anti-inflammatory effect (p < 0.001) Decreased NO production Inhibited TNF-α, IL-1β, IL-6, and COX-2 production | [54]. |
| Experimental Approaches | Key Results | References |
|---|---|---|
| Cyclophosphamide-induced immunosuppression in mice | Exhibited effective immunomodulatory effect Potentiated non-specific immune response Increased haematological parameter Increased neutrophil adhesion test | [26] |
| Ovalbumin-induced airway | Decreased the proliferation of lymphocytes, eosinophils, and monocytes Decreased the perivascular, peribronchiolar, and alveolar infiltration of inflammatory cells Reduced the expression of VCAM-1/OVA sIgE levels | [103] |
| Dextran sodium sulfate (DSS)-induced colitis model | Reduced DSS-induced inflammatory responses Regulated the Treg/Th17 balance Up-regulated IL-10 and TGF-β expression Decreased the expression of IL-17A Suppressed the Th17 cell response Promoted the development of Treg cells Provoked higher generation of gut microbiota-derived short-chain fatty acids (SCFAs), specifically butyrate Activated PPARγ by butyrate restored Treg/Th17 cells balance | [53] |
| Carrageenan-induced hind paw edema model in NIH mice | Reduced macrophage release of TNF-α/NO/COX-2/IL-1β/IL-6 Did not alter macrophage viability | [54] |
| Network pharmacology data | Targeted PI3K-Akt signaling pathway, the TNF-α and IL-17 signaling pathway, and Th17 cell differentiation Protected against myasthenia gravis | [56] |
| Biological network analysis | Targeted STAT3, CCL2, MAPK1, FOS, VEGFA, CASP3, IL-6, MAPK3, MAPK8, and CASP8 Acted against COVID-19 Controlled immune, inflammatory response and apoptosis | [57] |
| Murine cells | Suppressed Concanavalin A (ConA)-induced T-cell proliferation Did not block T helper 2 (Th2) and cytokine (IL-4 and IL-10) secretion Showed no effect on the secretion of Th1 cytokines (IL-2 and IFN-γ) | [104] |
| Experimental Approaches | Key Results | References |
|---|---|---|
| In silico docking Recombinant GABAA receptor subtypes expressed in Xenopus laevis oocytes | Up-regulated GABA-induced currents at the ternary α2β2γ2L, α4β3δ, and binary α4β3 subtypes of GABAAR | [105] |
| Human neuronal cells (SH-SY5Y cells) | Maintained ROS levels inside the cells Prevented oxidative stress-induced cell death Facilitated the up-regulation of FoxO3a/CAT/Bcl-2 in the neurons Increased the expression levels of sirtuin 1 (SIRT1) Decreased acetylated lysine levels Stimulated SIRT1 activity | [61] |
| Glutamate-induced neurotoxicity in hippocampal HT-22 cell line | Exhibited neuroprotective activity Inhibited ROS production Inhibited Ca2+ production | [106] |
| Hypoxia/reoxygenation (H/R)-induced excitotoxicity in hippocampal neurons | Protected against excitotoxicity Attenuated oxidative stress Attenuated mitochondrial dysfunction and DNA damage Controlled vesicle exocytosis Reduced VGLUT1 expression and vesicle pool size Suppressed GluN2B expression Up-regulated mitophagy-related proteins: LC3BII/p62/PINK1 | [62] |
| Ketamine-induced psychotic symptoms in mice | Decreased locomotor activity Reduced stereotypic behaviors Decreased immobility duration Increased step-down latency Increased GABA and GSH levels Decreased dopamine/MDA/TNF-α/AChE activity Did not found to cause catalepsy | [59] |
| Hippocampal neuronal cell line (HT22) In silico analysis | Reduced ROS production Down-regulated cyclin D1/p53/p38/MAPK protein expressions Inhibited cell apoptosis Normalized cell cycle progression Inhibited cytochrome P450 (CYP: CYP1A1, CYP1A2, and CYP1B1) Revealed high binding to CYP1A1/CYP1A2 | [60] |
| Vanadium-induced toxicity in mice | Attenuated spatial learning deficits Reduced escape latency Reduced grooming and rearing Reduced stretch-attend posture frequency Increased activities of CAT/SOD Decreased oxidative stress markers Increased MBP expression Crossed the blood-brain barrier Exhibited potent antioxidant Exhibited neuroprotective activities | [63] |
| Paraquat exposure-induced Parkinson’s disease-like symptoms and oxidative stress in Drosophila melanogaster | Decreased AChE activity and ROS generation Prevented mortality and dopamine deficiency Enhanced locomotor performance and lipid peroxidation | [58] |
| Induced Cerebral Ischemic/Reperfusion Injury in rats | Reduced neurological deficits and infarct damage Improved histopathology changes Restored the levels of the endogenous antioxidant defense system Depressed the expression level of beclin1, and the conversion of LC3 I to LC3 II Promoted the phosphorylation of mTOR Inhibited the phosphorylation of AMPK/JNK and expression of JNK | [95] |
| Structure-based molecular docking | Showed higher selectivity for iNOS and nNOS Exhibited similar binding conformations Inhibited iNOS and nNOS without inhibiting eNOS | [64] |
| Neuroblastoma SH-SY5Y cells | Inhibition % = 13.74% at 50 μM Did not exhibit neuroprotective potential in Aβ1–42-treated SH-SY5Y cells | [107] |
| Vanadium-induced neurotoxicity | Decreased escape latency 28.01 ± 0.02 Increased swimming time in target quadrant 98.24 ± 17.38 Increased exploration and latency Increased activities of antioxidant enzymes Decreased oxidative stress markers and lipid peroxidation Increased MBP expression | [108] |
| HT-22 mouse hippocampal cells | Attenuated the neuron cell death Protected against glutamate toxicity | [65] |
| - | Reduced Aβ generation Decreased β-secretase activity Reduced expression of all γ-secretase components Reduced cholesterol and presenilin distribution in lipid rafts Decreased BACE1 internalization to endosomal compartments | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakrim, S.; Benkhaira, N.; Bourais, I.; Benali, T.; Lee, L.-H.; El Omari, N.; Sheikh, R.A.; Goh, K.W.; Ming, L.C.; Bouyahya, A. Health Benefits and Pharmacological Properties of Stigmasterol. Antioxidants 2022, 11, 1912. https://doi.org/10.3390/antiox11101912
Bakrim S, Benkhaira N, Bourais I, Benali T, Lee L-H, El Omari N, Sheikh RA, Goh KW, Ming LC, Bouyahya A. Health Benefits and Pharmacological Properties of Stigmasterol. Antioxidants. 2022; 11(10):1912. https://doi.org/10.3390/antiox11101912
Chicago/Turabian StyleBakrim, Saad, Nesrine Benkhaira, Ilhame Bourais, Taoufiq Benali, Learn-Han Lee, Nasreddine El Omari, Ryan A. Sheikh, Khang Wen Goh, Long Chiau Ming, and Abdelhakim Bouyahya. 2022. "Health Benefits and Pharmacological Properties of Stigmasterol" Antioxidants 11, no. 10: 1912. https://doi.org/10.3390/antiox11101912