
Plant Glutathione Peroxidases: Non-Heme Peroxidases with Large Functional Flexibility as a Core Component of ROS-Processing Mechanisms and Signalling


Abstract
:1. Introduction
2. Phylogenetic Aspects of Plant GPXs
3. Structure, Biochemical Properties, and Main Activities of Plant GPX Proteins
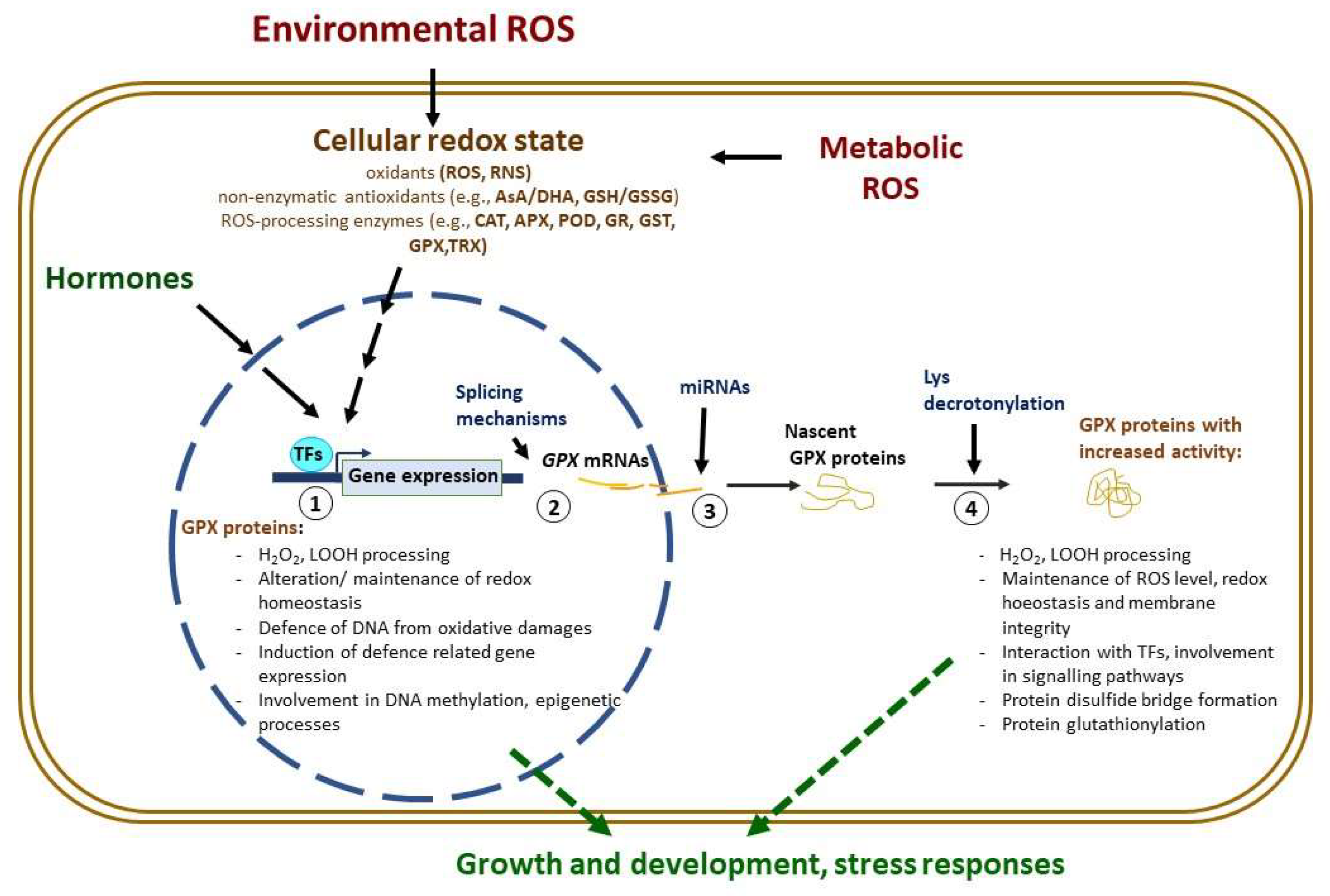
4. Involvement of GPXs in the Signalling Crosstalk under Abiotic Stress Responses
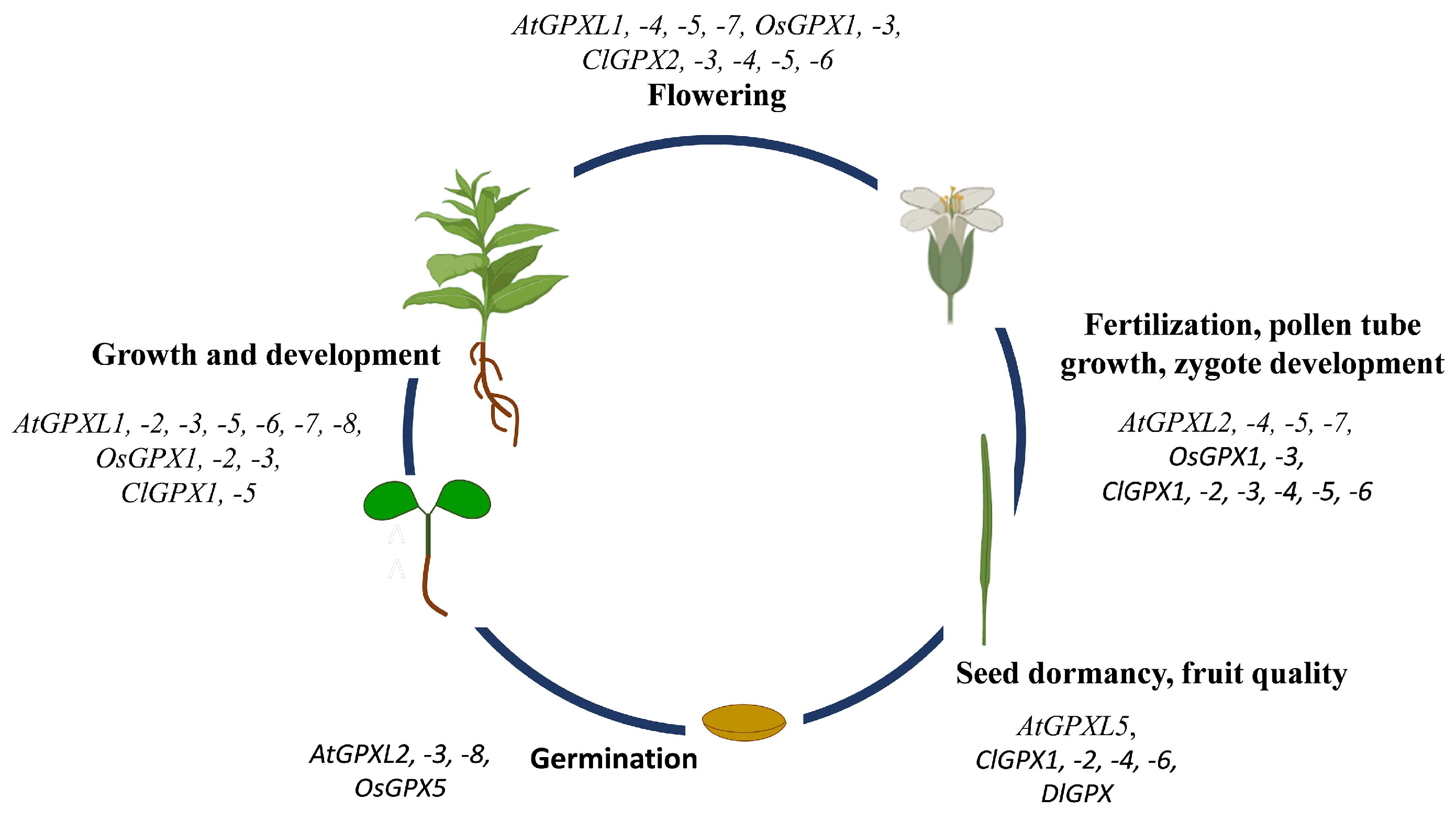
5. GPXs Regulates the Growth and Development of Plants
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Homeostasis and Antioxidant Signaling: A Metabolic Interface between Stress Perception and Physiological Responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, V.P.; Awasthi, M.; Singh, S.; Tiwari, S.; Dwivedi, U.N. A Comprehensive Review on Function and Application of Plant Peroxidases. Biochem. Anal. Biochem. 2017, 6, 308. [Google Scholar] [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases Have More Functions than a Swiss Army Knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef]
- Koua, D.; Cerutti, L.; Falquet, L.; Sigrist, C.J.A.; Theiler, G.; Hulo, N.; Dunand, C. PeroxiBase: A Database with New Tools for Peroxidase Family Classification. Nucleic Acids Res. 2009, 37, D261–D266. [Google Scholar] [CrossRef] [Green Version]
- Passardi, F.; Theiler, G.; Zamocky, M.; Cosio, C.; Rouhier, N.; Teixera, F.; Margis-Pinheiro, M.; Ioannidis, V.; Penel, C.; Falquet, L. PeroxiBase: The Peroxidase Database. Phytochemistry 2007, 68, 1605–1611. [Google Scholar] [CrossRef]
- Rouhier, N.; Jacquot, J.-P. The Plant Multigenic Family of Thiol Peroxidases. Free Radic. Biol. Med. 2005, 38, 1413–1421. [Google Scholar] [CrossRef]
- Herbette, S.; Roeckel-Drevet, P.; Drevet, J.R. Seleno-independent Glutathione Peroxidases: More than Simple Antioxidant Scavengers. FEBS J. 2007, 274, 2163–2180. [Google Scholar] [CrossRef]
- Dayer, R.; Fischer, B.B.; Eggen, R.I.L.; Lemaire, S.D. The Peroxiredoxin and Glutathione Peroxidase Families in Chlamydomonas Reinhardtii. Genetics 2008, 179, 41–57. [Google Scholar] [CrossRef] [Green Version]
- Attacha, S.; Solbach, D.; Bela, K.; Moseler, A.; Wagner, S.; Schwarzländer, M.; Aller, I.; Müller, S.J.; Meyer, A.J. Glutathione Peroxidase-like Enzymes Cover Five Distinct Cell Compartments and Membrane Surfaces in Arabidopsis Thaliana. Plant Cell Environ. 2017, 40, 1281–1295. [Google Scholar] [CrossRef]
- Flohé, L.; Toppo, S.; Orian, L. The Glutathione Peroxidase Family: Discoveries and Mechanism. Free Radic. Biol. Med. 2022, 187, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.R. The Glutathione Peroxidases. Cell. Mol. Life Sci. C 2001, 57, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Nakagawa, Y. Biological Significance of Phospholipid Hydroperoxide Glutathione Peroxidase (PHGPx, GPx4) in Mammalian Cells. Free Radic. Biol. Med. 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Mills, G.C. Hemoglobin Catabolism: I. Glutathione Peroxidase, an Erythrocyte Enzyme Which Protects Hemoglobin from Oxidative Breakdown. J. Biol. Chem. 1957, 229, 189–197. [Google Scholar] [CrossRef]
- Trenz, T.S.; Delaix, C.L.; Turchetto-Zolet, A.C.; Zamocky, M.; Lazzarotto, F.; Margis-Pinheiro, M. Going Forward and Back: The Complex Evolutionary History of the GPx. Biology 2021, 10, 1165. [Google Scholar] [CrossRef] [PubMed]
- Islam, T.; Manna, M.; Kaul, T.; Pandey, S.; Reddy, C.S.; Reddy, M.K. Genome-Wide Dissection of Arabidopsis and Rice for the Identification and Expression Analysis of Glutathione Peroxidases Reveals Their Stress-Specific and Overlapping Response Patterns. Plant Mol. Biol. Rep. 2015, 33, 1413–1427. [Google Scholar] [CrossRef]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione Peroxidase Family—An Evolutionary Overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione Peroxidases. Biochim. Biophys. Acta BBA-Gen. Subj. 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Benhar, M. Roles of Mammalian Glutathione Peroxidase and Thioredoxin Reductase Enzymes in the Cellular Response to Nitrosative Stress. Free Radic. Biol. Med. 2018, 127, 160–164. [Google Scholar] [CrossRef]
- Conrad, M.; Moreno, S.G.; Sinowatz, F.; Ursini, F.; Kolle, S.; Roveri, A.; Brielmeier, M.; Wurst, W.; Maiorino, M.; Bornkamm, G.W. The Nuclear Form of Phospholipid Hydroperoxide Glutathione Peroxidase Is a Protein Thiol Peroxidase Contributing to Sperm Chromatin Stability. Mol. Cell. Biol. 2005, 25, 7637–7644. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohe, R.; Kipp, A. Glutathione Peroxidases in Different Stages of Carcinogenesis. Biochim. Biophys. Acta BBA-Gen. Subj. 2009, 1790, 1555–1568. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Geng, Y.; Yang, Y.; Seim, I.; Yang, G. Oxidative Stress Drives Divergent Evolution of the Glutathione Peroxidase (GPX) Gene Family in Mammals. Integr. Zool. 2021, 16, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Krümmel, B.; Plötz, T.; Jörns, A.; Lenzen, S.; Mehmeti, I. The Central Role of Glutathione Peroxidase 4 in the Regulation of Ferroptosis and Its Implications for Pro-Inflammatory Cytokine-Mediated Beta-Cell Death. Biochim. Biophys. Acta BBA-Mol. Basis Dis. 2021, 1867, 166114. [Google Scholar] [CrossRef]
- Ursini, F.; Travain, V.B.; Cozza, G.; Miotto, G.; Roveri, A.; Toppo, S.; Maiorino, M. A White Paper on Phospholipid Hydroperoxide Glutathione Peroxidase (GPx4) Forty Years Later. Free Radic. Biol. Med. 2022, 188, 117–133. [Google Scholar] [CrossRef]
- Weaver, K.; Skouta, R. The Selenoprotein Glutathione Peroxidase 4: From Molecular Mechanisms to Novel Therapeutic Opportunities. Biomedicines 2022, 10, 891. [Google Scholar] [CrossRef] [PubMed]
- Navrot, N.; Collin, V.; Gualberto, J.; Gelhaye, E.; Hirasawa, M.; Rey, P.; Knaff, D.B.; Issakidis, E.; Jacquot, J.-P.; Rouhier, N. Plant Glutathione Peroxidases Are Functional Peroxiredoxins Distributed in Several Subcellular Compartments and Regulated during Biotic and Abiotic Stresses. Plant Physiol. 2006, 142, 1364–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, A.; Yabuta, Y.; Takeda, T.; Nakano, Y.; Shigeoka, S. Hydroperoxide Reduction by Thioredoxin-specific Glutathione Peroxidase Isoenzymes of Arabidopsis thaliana. FEBS J. 2006, 273, 5589–5597. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-Related Redox Regulation and Signaling in Plants. In Proceedings of the Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 80, pp. 3–12. [Google Scholar]
- Foyer, C.H.; Wilson, M.H.; Wright, M.H. Redox Regulation of Cell Proliferation: Bioinformatics and Redox Proteomics Approaches to Identify Redox-Sensitive Cell Cycle Regulators. Free Radic. Biol. Med. 2018, 122, 137–149. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Mase, K.; Tsukagoshi, H. Reactive Oxygen Species Link Gene Regulatory Networks During Arabidopsis Root Development. Front. Plant Sci. 2021, 12, 642. [Google Scholar] [CrossRef]
- Rattanawong, K.; Koiso, N.; Toda, E.; Kinoshita, A.; Tanaka, M.; Tsuji, H.; Okamoto, T. Regulatory Functions of ROS Dynamics via Glutathione Metabolism and Glutathione Peroxidase Activity in Developing Rice Zygote. Plant J. 2021, 108, 1097–1115. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Lv, D.; Wang, P.; Wang, X.-C.; Chen, J.; Miao, C.; Song, C.-P. An Arabidopsis Glutathione Peroxidase Functions as both a Redox Transducer and a Scavenger in Abscisic Acid and Drought Stress Responses. Plant Cell 2006, 18, 2749–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passaia, G.; Queval, G.; Bai, J.; Margis-Pinheiro, M.; Foyer, C.H. The Effects of Redox Controls Mediated by Glutathione Peroxidases on Root Architecture in Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 1403–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passaia, G.; Margis-Pinheiro, M. Glutathione Peroxidases as Redox Sensor Proteins in Plant Cells. Plant Sci. 2015, 234, 22–26. [Google Scholar] [CrossRef]
- Meyer, A.J.; Dreyer, A.; Ugalde, J.M.; Feitosa-Araujo, E.; Dietz, K.-J.; Schwarzländer, M. Shifting Paradigms and Novel Players in Cys-Based Redox Regulation and ROS Signaling in Plants-and Where to Go Next. Biol. Chem. 2021, 402, 399–423. [Google Scholar] [CrossRef]
- Ferro, D.; Bakiu, R.; Pucciarelli, S.; Miceli, C.; Vallesi, A.; Irato, P.; Santovito, G. Molecular Characterization, Protein–Protein Interaction Network, and Evolution of Four Glutathione Peroxidases from Tetrahymena Thermophila. Antioxidants 2020, 9, 949. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, B.; Miao, R.; Deng, X.; Duan, Y.; Cheng, Y.; Zhang, W.; Shi, M.; Huang, K.; Xia, X.-Q. Transcriptomic and Physiological Responses to Oxidative Stress in a Chlamydomonas Reinhardtii Glutathione Peroxidase Mutant. Genes 2020, 11, 463. [Google Scholar] [CrossRef]
- Ozyigit, I.I.; Filiz, E.; Vatansever, R.; Kurtoglu, K.Y.; Koc, I.; Öztürk, M.X.; Anjum, N.A. Identification and Comparative Analysis of H2O2-Scavenging Enzymes (Ascorbate Peroxidase and Glutathione Peroxidase) in Selected Plants Employing Bioinformatics Approaches. Front. Plant Sci. 2016, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Huai, X.; Li, P.; Raza, A.; Mubarik, M.S.; Habib, M.; Fiaz, S.; Zhang, B.; Pan, J.; Khan, R.S.A. Genome-Wide Characterization of Glutathione Peroxidase (GPX) Gene Family in Rapeseed (Brassica napus L.) Revealed Their Role in Multiple Abiotic Stress Response and Hormone Signaling. Antioxidants 2021, 10, 1481. [Google Scholar] [CrossRef]
- Sharma, A.; Kaur, A.; Tyagi, S.; Upadhyay, S.K. Glutathione Peroxidases in Plants: Innumerable Role in Abiotic Stress Tolerance and Plant Development. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Jang, M.-G.; Noh, H.-Y.; Lee, H.-J.; Sukweenadhi, J.; Kim, J.-H.; Kim, S.-Y.; Kwon, W.-S.; Yang, D.-C. Molecular Characterization of Two Glutathione Peroxidase Genes of Panax Ginseng and Their Expression Analysis against Environmental Stresses. Gene 2014, 535, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churin, Y.; Schilling, S.; Börner, T. A Gene Family Encoding Glutathione Peroxidase Homologues in Hordeum Vulgare (Barley). FEBS Lett. 1999, 459, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Aleem, M.; Aleem, S.; Sharif, I.; Wu, Z.; Aleem, M.; Tahir, A.; Atif, R.M.; Cheema, H.M.N.; Shakeel, A.; Lei, S. Characterization of SOD and GPX Gene Families in the Soybeans in Response to Drought and Salinity Stresses. Antioxidants 2022, 11, 460. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Han, X.-M.; Wang, W.; Yang, H.-L. Molecular and Catalytic Properties of Glutathione Peroxidase Family Proteins from Pinus tabulaeformis. Plant Mol. Biol. Rep. 2014, 32, 771–778. [Google Scholar] [CrossRef]
- Jana, G.A.; Yaish, M.W. Genome-Wide Identification and Functional Characterization of Glutathione Peroxidase Genes in Date Palm (Phoenix dactylifera L.) under Stress Conditions. Plant Gene 2020, 23, 100237. [Google Scholar] [CrossRef]
- Wang, X.; Liu, X.; An, Y.; Zhang, H.; Meng, D.; Jin, Y.; Huo, H.; Yu, L.; Zhang, J. Identification of Glutathione Peroxidase Gene Family in Ricinus Communis and Functional Characterization of RcGPX4 in Cold Tolerance. Front. Plant Sci. 2021, 12, 707127. [Google Scholar] [CrossRef]
- Zhou, Y.; Hu, L.; Ye, S.; Jiang, L.; Liu, S. Genome-Wide Identification of Glutathione Peroxidase (GPX) Gene Family and Their Response to Abiotic Stress in Cucumber. 3 Biotech 2018, 8, 159. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, J.; Wang, J.; Yang, W.; Yang, Y. Identification and Characterization of the Glutathione Peroxidase (GPX) Gene Family in Watermelon and Its Expression under Various Abiotic Stresses. Agronomy 2018, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Ramos, J.; Matamoros, M.A.; Naya, L.; James, E.K.; Rouhier, N.; Sato, S.; Tabata, S.; Becana, M. The Glutathione Peroxidase Gene Family of Lotus Japonicus: Characterization of Genomic Clones, Expression Analyses and Immunolocalization in Legumes. New Phytol. 2009, 181, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Akbudak, M.A.; Filiz, E.; Vatansever, R.; Kontbay, K. Genome-Wide Identification and Expression Profiling of Ascorbate Peroxidase (APX) and Glutathione Peroxidase (GPX) Genes under Drought Stress in Sorghum (Sorghum bicolor L.). J. Plant Growth Regul. 2018, 37, 925–936. [Google Scholar] [CrossRef]
- Mallikarjuna, M.G.; Sharma, R.; Veeraya, P.; Tyagi, A.; Rao, A.R.; Chandappa, L.H.; Chinnusamy, V. Evolutionary and Functional Characterisation of Glutathione Peroxidases Showed Splicing Mediated Stress Responses in Maize. Plant Physiol. Biochem. 2022, 178, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Milla, M.A.R.; Maurer, A.; Huete, A.R.; Gustafson, J.P. Glutathione Peroxidase Genes in Arabidopsis Are Ubiquitous and Regulated by Abiotic Stresses through Diverse Signaling Pathways. Plant J. 2003, 36, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Li, K.; Li, H.; Song, C.-P.; Miao, Y. The Glutathione Peroxidase Gene Family in Gossypium Hirsutum: Genome-Wide Identification, Classification, Gene Expression and Functional Analysis. Sci. Rep. 2017, 7, 44743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, S.; Sembi, J.K.; Upadhyay, S.K. Gene Architecture and Expression Analyses Provide Insights into the Role of Glutathione Peroxidases (GPXs) in Bread Wheat (Triticum aestivum L.). J. Plant Physiol. 2018, 223, 19–31. [Google Scholar] [CrossRef]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant Glutathione Peroxidases: Emerging Role of the Antioxidant Enzymes in Plant Development and Stress Responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Gupta, S.; Dong, Y.; Dijkwel, P.P.; Mueller-Roeber, B.; Gechev, T.S. Genome-Wide Analysis of ROS Antioxidant Genes in Resurrection Species Suggest an Involvement of Distinct ROS Detoxification Systems during Desiccation. Int. J. Mol. Sci. 2019, 20, 3101. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Chen, J.; Ma, T.; Li, H.; Wang, N.; Li, Z.; Zhang, Z.; Zhou, Y. The Glutathione Peroxidase Gene Family in Thellungiella Salsuginea: Genome-Wide Identification, Classification, and Gene and Protein Expression Analysis under Stress Conditions. Int. J. Mol. Sci. 2014, 15, 3319–3335. [Google Scholar] [CrossRef] [Green Version]
- San Koh, C.; Didierjean, C.; Navrot, N.; Panjikar, S.; Mulliert, G.; Rouhier, N.; Jacquot, J.-P.; Aubry, A.; Shawkataly, O.; Corbier, C. Crystal Structures of a Poplar Thioredoxin Peroxidase That Exhibits the Structure of Glutathione Peroxidases: Insights into Redox-Driven Conformational Changes. J. Mol. Biol. 2007, 370, 512–529. [Google Scholar]
- Toppo, S.; Vanin, S.; Bosello, V.; Tosatto, S.C.E. Evolutionary and Structural Insights into the Multifaceted Glutathione Peroxidase (Gpx) Superfamily. Antioxid. Redox Signal. 2008, 10, 1501–1514. [Google Scholar] [CrossRef]
- Herbette, S.; Lenne, C.; Leblanc, N.; Julien, J.; Drevet, J.R.; Roeckel-Drevet, P. Two GPX-like Proteins from Lycopersicon Esculentum and Helianthus Annuus Are Antioxidant Enzymes with Phospholipid Hydroperoxide Glutathione Peroxidase and Thioredoxin Peroxidase Activities. Eur. J. Biochem. 2002, 269, 2414–2420. [Google Scholar] [CrossRef]
- Jung, B.G.; Lee, K.O.; Lee, S.S.; Chi, Y.H.; Jang, H.H.; Kang, S.S.; Lee, K.; Lim, D.; Yoon, S.C.; Yun, D.-J. A Chinese Cabbage CDNA with High Sequence Identity to Phospholipid Hydroperoxide Glutathione Peroxidases Encodes a Novel Isoform of Thioredoxin-Dependent Peroxidase. J. Biol. Chem. 2002, 277, 12572–12578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiler, A.; Schneider, M.; Förster, H.; Roth, S.; Wirth, E.K.; Culmsee, C.; Plesnila, N.; Kremmer, E.; Rådmark, O.; Wurst, W. Glutathione Peroxidase 4 Senses and Translates Oxidative Stress into 12/15-Lipoxygenase Dependent-and AIF-Mediated Cell Death. Cell Metab. 2008, 8, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Bela, K.; Riyazuddin, R.; Horváth, E.; Hurton, Á.; Gallé, Á.; Takács, Z.; Zsigmond, L.; Szabados, L.; Tari, I.; Csiszár, J. Comprehensive Analysis of Antioxidant Mechanisms in Arabidopsis Glutathione Peroxidase-like Mutants under Salt-and Osmotic Stress Reveals Organ-Specific Significance of the AtGPXL’s Activities. Environ. Exp. Bot. 2018, 150, 127–140. [Google Scholar] [CrossRef]
- Chang, C.C.C.; Slesak, I.; Jordá, L.; Sotnikov, A.; Melzer, M.; Miszalski, Z.; Mullineaux, P.M.; Parker, J.E.; Karpinska, B.; Karpinski, S. Arabidopsis Chloroplastic Glutathione Peroxidases Play a Role in Cross Talk between Photooxidative Stress and Immune Responses. Plant Physiol. 2009, 150, 670–683. [Google Scholar] [CrossRef] [Green Version]
- Gaber, A. Arabidopsis Glutathione Peroxidase 8 Is a Key Enzyme in Response to Environmental Stresses. Arab. J. Biotechnol. 2011, 14, 213–224. [Google Scholar]
- Riyazuddin, R.; Bela, K.; Horváth, E.; Rigó, G.; Gallé, Á.; Szabados, L.; Fehér, A.; Csiszár, J. Overexpression of the Arabidopsis Glutathione Peroxidase-like 5 Gene (AtGPXL5) Resulted in Altered Plant Development and Redox Status. Environ. Exp. Bot. 2019, 167, 103849. [Google Scholar] [CrossRef]
- Toppo, S.; Flohé, L.; Ursini, F.; Vanin, S.; Maiorino, M. Catalytic Mechanisms and Specificities of Glutathione Peroxidases: Variations of a Basic Scheme. Biochim. Biophys. Acta BBA-Gen. Subj. 2009, 1790, 1486–1500. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Bela, K.; Poór, P.; Szepesi, Á.; Horváth, E.; Rigó, G.; Szabados, L.; Fehér, A.; Csiszár, J. Crosstalk between the Arabidopsis Glutathione Peroxidase-Like 5 Isoenzyme (AtGPXL5) and Ethylene. Int. J. Mol. Sci. 2022, 23, 5749. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Edwards, R. Glutathione Transferases. In The Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2010; Volume 8, p. e0131. [Google Scholar]
- Luo, D.; Smith, S.W.; Anderson, B.D. Kinetics and Mechanism of the Reaction of Cysteine and Hydrogen Peroxide in Aqueous Solution. J. Pharm. Sci. 2005, 94, 304–316. [Google Scholar] [CrossRef]
- Marinho, H.S.; Real, C.; Cyrne, L.; Soares, H.; Antunes, F. Hydrogen Peroxide Sensing, Signaling and Regulation of Transcription Factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef] [Green Version]
- Diao, Y.; Xu, H.; Li, G.; Yu, A.; Yu, X.; Hu, W.; Zheng, X.; Li, S.; Wang, Y.; Hu, Z. Cloning a Glutathione Peroxidase Gene from Nelumbo Nucifera and Enhanced Salt Tolerance by Overexpressing in Rice. Mol. Biol. Rep. 2014, 41, 4919–4927. [Google Scholar] [CrossRef] [PubMed]
- Passaia, G.; Fonini, L.S.; Caverzan, A.; Jardim-Messeder, D.; Christoff, A.P.; Gaeta, M.L.; de Araujo Mariath, J.E.; Margis, R.; Margis-Pinheiro, M. The Mitochondrial Glutathione Peroxidase GPX3 Is Essential for H2O2 Homeostasis and Root and Shoot Development in Rice. Plant Sci. 2013, 208, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Delaunay, A.; Pflieger, D.; Barrault, M.-B.; Vinh, J.; Toledano, M.B. A Thiol Peroxidase Is an H2O2 Receptor and Redox-Transducer in Gene Activation. Cell 2002, 111, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Paiva, A.L.S.; Passaia, G.; Jardim-Messeder, D.; Nogueira, F.C.S.; Domont, G.B.; Margis-Pinheiro, M. The Mitochondrial Isoform Glutathione Peroxidase 3 (OsGPX3) Is Involved in ABA Responses in Rice Plants. J. Proteom. 2021, 232, 104029. [Google Scholar] [CrossRef] [PubMed]
- Lima-Melo, Y.; Carvalho, F.E.L.; Martins, M.O.; Passaia, G.; Sousa, R.H.V.; Neto, M.C.L.; Margis-Pinheiro, M.; Silveira, J.A.G. Mitochondrial GPX1 Silencing Triggers Differential Photosynthesis Impairment in Response to Salinity in Rice Plants. J. Integr. Plant Biol. 2016, 58, 737–748. [Google Scholar] [CrossRef]
- Paiva, A.L.S.; Passaia, G.; Lobo, A.K.M.; Jardim-Messeder, D.; Silveira, J.A.G.; Margis-Pinheiro, M. Mitochondrial Glutathione Peroxidase (OsGPX3) Has a Crucial Role in Rice Protection against Salt Stress. Environ. Exp. Bot. 2019, 158, 12–21. [Google Scholar] [CrossRef]
- Li, K.; Jia, Q.; Guo, J.; Zhu, Z.; Shao, M.; Wang, J.; Li, W.; Dai, J.; Guo, M.; Li, R. The High Chlorophyll Fluorescence 244 (HCF244) Is Potentially Involved in Glutathione Peroxidase 7-Regulated High Light Stress in Arabidopsis thaliana. Environ. Exp. Bot. 2022, 195, 104767. [Google Scholar] [CrossRef]
- Gaber, A.; Ogata, T.; Maruta, T.; Yoshimura, K.; Tamoi, M.; Shigeoka, S. The Involvement of Arabidopsis Glutathione Peroxidase 8 in the Suppression of Oxidative Damage in the Nucleus and Cytosol. Plant Cell Physiol. 2012, 53, 1596–1606. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Cai, X.-T.; Du, J.; Zhao, T.-L.; Wang, P.-F.; Zhao, P.-X.; Liu, R.; Xie, Q.; Cao, X.-F.; Xiang, C.-B. PARAQUAT TOLERANCE3 Is an E3 Ligase That Switches off Activated Oxidative Response by Targeting Histone-Modifying PROTEIN METHYLTRANSFERASE4b. PLoS Genet. 2016, 12, e1006332. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Lin, P.; Luo, Y.; Bai, H.; Liao, X.; Li, X.; Tian, Y.; Jiang, B.; Pan, Y.; Zhang, F. Lysine Decrotonylation of Glutathione Peroxidase at Lysine 220 Site Increases Glutathione Peroxidase Activity to Resist Cold Stress in Chrysanthemum. Ecotoxicol. Environ. Saf. 2022, 232, 113295. [Google Scholar] [CrossRef]
- Ganie, S.A.; Reddy, A.S.N. Stress-Induced Changes in Alternative Splicing Landscape in Rice: Functional Significance of Splice Isoforms in Stress Tolerance. Biology 2021, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Zhai, C.-Z.; Zhao, L.; Yin, L.-J.; Chen, M.; Wang, Q.-Y.; Li, L.-C.; Xu, Z.-S.; Ma, Y.-Z. Two Wheat Glutathione Peroxidase Genes Whose Products Are Located in Chloroplasts Improve Salt and H2O2 Tolerances in Arabidopsis. PLoS ONE 2013, 8, e73989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, A.M.M.; Reis, S.P.M.; Gramacho, K.P.; Micheli, F. The Glutathione Peroxidase Family of Theobroma Cacao: Involvement in the Oxidative Stress during Witches’ Broom Disease. Int. J. Biol. Macromol. 2020, 164, 3698–3708. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, S.; Sui, X.; Wang, J.; Geng, Y.; Gao, F.; Zhou, Y. Genome-Wide Characterization, Evolution, and Expression Analysis of the Ascorbate Peroxidase and Glutathione Peroxidase Gene Families in Response to Cold and Osmotic Stress in Ammopiptanthus nanus. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Passaia, G.; Caverzan, A.; Fonini, L.S.; Carvalho, F.E.L.; Silveira, J.A.G.; Margis-Pinheiro, M. Chloroplastic and Mitochondrial GPX Genes Play a Critical Role in Rice Development. Biol. Plant. 2014, 58, 375–378. [Google Scholar] [CrossRef]
- Wu, F.; Jiang, G.; Yan, H.; Xiao, L.; Liang, H.; Zhang, D.; Jiang, Y.; Duan, X. Redox Regulation of Glutathione Peroxidase by Thioredoxin in Longan Fruit in Relation to Senescence and Quality Deterioration. Food Chem. 2021, 345, 128664. [Google Scholar] [CrossRef]
- Bashandy, T.; Guilleminot, J.; Vernoux, T.; Caparros-Ruiz, D.; Ljung, K.; Meyer, Y.; Reichheld, J.-P. Interplay between the NADP-Linked Thioredoxin and Glutathione Systems in Arabidopsis Auxin Signaling. Plant Cell 2010, 22, 376–391. [Google Scholar] [CrossRef] [Green Version]
- Diaz Vivancos, P.; Wolff, T.; Markovic, J.; Pallardo, F.V.; Foyer, C.H. A Nuclear Glutathione Cycle within the Cell Cycle. Biochem. J. 2010, 431, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New Roles in Redox Signaling for an Old Antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [Green Version]
- Reichheld, J.-P.; Khafif, M.; Riondet, C.; Droux, M.; Bonnard, G.; Meyer, Y. Inactivation of Thioredoxin Reductases Reveals a Complex Interplay between Thioredoxin and Glutathione Pathways in Arabidopsis Development. Plant Cell 2007, 19, 1851–1865. [Google Scholar] [CrossRef] [Green Version]
- Pagnussat, G.C.; Yu, H.-J.; Ngo, Q.A.; Rajani, S.; Mayalagu, S.; Johnson, C.S.; Capron, A.; Xie, L.-F.; Ye, D.; Sundaresan, V. Genetic and Molecular Identification of Genes Required for Female Gametophyte Development and Function in Arabidopsis. Development 2005, 132, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marty, L.; Siala, W.; Schwarzländer, M.; Fricker, M.D.; Wirtz, M.; Sweetlove, L.J.; Meyer, Y.; Meyer, A.J.; Reichheld, J.-P.; Hell, R. The NADPH-Dependent Thioredoxin System Constitutes a Functional Backup for Cytosolic Glutathione Reductase in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 9109–9114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Holmgren, A. The Thioredoxin Antioxidant System. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Bela, K.; Bangash, S.A.K.; Csiszár, J. Plant Glutathione Peroxidases: Antioxidant Enzymes in Plant Stress Responses and Tolerance. In Glutathione in Plant Growth, Development, and Stress Tolerance; Springer: Cham, Switzerland, 2017; pp. 113–126. [Google Scholar]
- Jiang, K.; Moe-Lange, J.; Hennet, L.; Feldman, L.J. Salt Stress Affects the Redox Status of Arabidopsis Root Meristems. Front. Plant Sci. 2016, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.-C.; Seeley, K.A.; Sung, Z.R. RML1 and RML2, Arabidopsis Genes Required for Cell Proliferation at the Root Tip. Plant Physiol. 1995, 107, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Vernoux, T.; Wilson, R.C.; Seeley, K.A.; Reichheld, J.-P.; Muroy, S.; Brown, S.; Maughan, S.C.; Cobbett, C.S.; Van Montagu, M.; Inzé, D. The ROOT MERISTEMLESS1/CADMIUM SENSITIVE2 Gene Defines a Glutathione-Dependent Pathway Involved in Initiation and Maintenance of Cell Division during Postembryonic Root Development. Plant Cell 2000, 12, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Frendo, P.; Harrison, J.; Norman, C.; Jiménez, M.J.H.; Van de Sype, G.; Gilabert, A.; Puppo, A. Glutathione and Homoglutathione Play a Critical Role in the Nodulation Process of Medicago Truncatula. Mol. Plant-Microbe Interact. 2005, 18, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Schnaubelt, D.; Queval, G.; Dong, Y.; Diaz-Vivancos, P.; Makgopa, M.E.; Howell, G.; De Simone, A.; Bai, J.; Hannah, M.A.; Foyer, C.H. Low Glutathione Regulates Gene Expression and the Redox Potentials of the Nucleus and Cytosol in Arabidopsis thaliana. Plant. Cell Environ. 2015, 38, 266–279. [Google Scholar] [CrossRef]
- Sato, A.; Yamamoto, K.T. Overexpression of the Non-canonical Aux/IAA Genes Causes Auxin-related Aberrant Phenotypes in Arabidopsis. Physiol. Plant. 2008, 133, 397–405. [Google Scholar] [CrossRef]
- Mravec, J.; Skůpa, P.; Bailly, A.; Hoyerová, K.; Křeček, P.; Bielach, A.; Petrášek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.-D. Subcellular Homeostasis of Phytohormone Auxin Is Mediated by the ER-Localized PIN5 Transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef]
- Hamaguchi, A.; Yamashino, T.; Koizumi, N.; Kiba, T.; Kojima, M.; Sakakibara, H.; Mizuno, T. A Small Subfamily of Arabidopsis RADIALIS-LIKE SANT/MYB Genes: A Link to HOOKLESS1-Mediated Signal Transduction during Early Morphogenesis. Biosci. Biotechnol. Biochem. 2008, 72, 2687–2696. [Google Scholar] [CrossRef] [Green Version]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.-P.; Riondet, C. Thioredoxin and Glutaredoxin Systems in Plants: Molecular Mechanisms, Crosstalks, and Functional Significance. Antioxid. Redox Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef] [PubMed]
- Shahpiri, A.; Svensson, B.; Finnie, C. The NADPH-Dependent Thioredoxin Reductase/Thioredoxin System in Germinating Barley Seeds: Gene Expression, Protein Profiles, and Interactions between Isoforms of Thioredoxin h and Thioredoxin Reductase. Plant Physiol. 2008, 146, 789. [Google Scholar] [CrossRef] [Green Version]
- Benitez-Alfonso, Y.; Cilia, M.; Roman, A.S.; Thomas, C.; Maule, A.; Hearn, S.; Jackson, D. Control of Arabidopsis Meristem Development by Thioredoxin-Dependent Regulation of Intercellular Transport. Proc. Natl. Acad. Sci. USA 2009, 106, 3615–3620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.; Wong, J.H.; Feldman, L.J.; Lemaux, P.G.; Buchanan, B.B. A Membrane-Associated Thioredoxin Required for Plant Growth Moves from Cell to Cell, Suggestive of a Role in Intercellular Communication. Proc. Natl. Acad. Sci. USA 2010, 107, 3900–3905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsova, B.; Hoja, U.; Wimmelbacher, M.; Greiner, E.; Üstün, Ş.; Melzer, M.; Petersen, K.; Lein, W.; Börnke, F. Plastidial Thioredoxin z Interacts with Two Fructokinase-like Proteins in a Thiol-Dependent Manner: Evidence for an Essential Role in Chloroplast Development in Arabidopsis and Nicotiana benthamiana. Plant Cell 2010, 22, 1498–1515. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Fang, G.; Yang, J.; Li, Y. A Thioredoxin-Dependent Glutathione Peroxidase (OsGPX5) Is Required for Rice Normal Development and Salt Stress Tolerance. Plant Mol. Biol. Report. 2017, 35, 333–342. [Google Scholar] [CrossRef]
- Zhou, B.; Yao, W.; Wang, S.; Wang, X.; Jiang, T. The Metallothionein Gene, TaMT3, from Tamarix Androssowii Confers Cd2+ Tolerance in Tobacco. Int. J. Mol. Sci. 2014, 15, 10398–10409. [Google Scholar] [CrossRef] [Green Version]
- Sarıkaya, E.; Doğan, S. Glutathione Peroxidase in Health and Diseases. In Glutathione System and Oxidative Stress in Health and Disease; IntechOpen: London, UK, 2020; p. 49. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Hu, T.; Zhu, S.; Mukaisho, K.; El-Rifai, W.; Peng, D.-F. Glutathione Peroxidase 7 Suppresses Cancer Cell Growth and Is Hypermethylated in Gastric Cancer. Oncotarget 2017, 8, 54345. [Google Scholar] [CrossRef] [Green Version]
- Gaber, A. The Importance of Arabidopsis Glutathione Peroxidase 8 for Protecting Arabidopsis Plant and E. Coli Cells against Oxidative Stress. GM Crops Food 2014, 5, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, M.; Sakamoto, W. Putative Phospholipid Hydroperoxide Glutathione Peroxidase Gene from Arabidopsis thaliana Induced by Oxidative Stress. Genes Genet. Syst. 1997, 72, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbette, S.; Le Menn, A.; Rousselle, P.; Ameglio, T.; Faltin, Z.; Branlard, G.; Eshdat, Y.; Julien, J.-L.; Drevet, J.R.; Roeckel-Drevet, P. Modification of Photosynthetic Regulation in Tomato Overexpressing Glutathione Peroxidase. Biochim. Biophys. Acta BBA-Gen. Subj. 2005, 1724, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Faltin, Z.; Holland, D.; Velcheva, M.; Tsapovetsky, M.; Roeckel-Drevet, P.; Handa, A.K.; Abu-Abied, M.; Friedman-Einat, M.; Eshdat, Y.; Perl, A. Glutathione Peroxidase Regulation of Reactive Oxygen Species Level Is Crucial for in Vitro Plant Differentiation. Plant Cell Physiol. 2010, 51, 1151–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Protein Structure | Active Site Cys/SeCys | Reducing Agent | Substrate | References | |
|---|---|---|---|---|---|
| Animal GPXs | tetramer, dimer, or monomer | SeCys or Cys | GSH, TRX, NADPH | H2O2, LOOH, ROOH | [11,12] |
| A. thaliana GPXLs | monomer | Cys | TRX | AlkylOOH CumOOH PCOOH PUFAOOH | [27] |
| GPXs from other plant species | monomer (dimer) | Cys | TRX, GSH | AlkylOOH CumOOH PCOOH PUFAOOH | [26,32,52,61,62] |
| Enzyme | Reported Function/Involvement | Organism | References | ||||
|---|---|---|---|---|---|---|---|
| Stress Responses | Redox Regulation/ Signalling | Normal Metabolism | Development | ||||
| Animal GPXs | |||||||
| HsGPX1, -2, -3, -5, -6 | H2O2 and lipid hydroperoxide processing, stress tolerance | Insulin signalling | H2O2 and lipid hydroperoxide processing + | Male fertility | Homo sapiens | [112] | |
| HsGPX4 | Lipid peroxidation, protein thiol oxidation | Cell death, Ferroptosis | Protein thiol oxidation | Spermatogenesis, chromatin condensation, | Homo sapiens | [23,24,25] | |
| HsGPX7 | General scavenging of ROS, lipid peroxides Anti-inflammatory | Anti-carcinogenesis | Homo sapiens | [18,21,113] | |||
| MmGPX4 | Anti-carcinogenesis | Spermatogenesis | Spermatogenesis, male fertility, embryogenic development | Mus musculus | [20] | ||
| A. thalianaGPXLs | AtGPXL1-8 | Diverse biotic and abiotic (heat, cold, salt, drought, osmotic and metal) stresses, ferroptosis | Maintenance of redox homeostasis, oxidative signal transducer in ABA and drought stress signalling, | Photosynthesis | Development in whole life cycle (embryogenesis, germination, root, shoot apical meristem, hypocotyl, root system, rosette leaves, flowering, pollen tube growth, fertilization, seed dormancy) | A. thaliana | [33,36,41,53,58,64,67,69,114,115] |
| GPXs from other plant species | OsGPX1-5 | Drought, salt, cold, oxidative stresses | Redox signalling, participation in the interaction between ER stress and redox homeostasis, crosstalk between mitochondria and chloroplast | Photosynthesis and cellular respiration | In development during the whole life cycle (embryogenesis, germination, root-, shoot apical meristem, hypocotyl, root system, seedling development, rosette leaves, inflorescence and silique, pollen tube growth, seed setting, grain filling, seed dormancy) | O. sativa | [16,32,34,39,74,77,87,110] |
| SlGPX1-5 (GSHPxle1-5) | Heat stress, cold, light stress | unknown | unknown | unknown | S. lycopersicum | [39,58,116] | |
| HvGPX1-3 | Oxidative stress, salt/osmotic stress, norflurazon, and paraquat resistance | unknown | unknown | unknown | H. vulgare | [43] | |
| BoGPX1-8 | Salinity, cold, waterlogging, and drought | Bn BoGPX genes might contribute to stress responses and hormone signaling pathways | unknown | In development of root, seed, leaf, stem, flower, and silique | B. oleracea | [40] | |
| BrGPX1-12 | Salinity, cold, waterlogging, and drought | BrGPX genes might contribute to stress responses and hormone signaling pathways | unknown | In development of root, seed, leaf, stem, flower, and silique | B. rapa | [40] | |
| BnGPX1-25 | Salinity, cold, waterlogging, and drought | BnGPX genes might contribute to stress responses and hormone signaling pathways | unknown | In development of root, seed, leaf, stem, flower, and silique | B. napus | [40] | |
| GhGPX1-13 | Salt stress, heat, sulphate solution | Importance of GhGPXs in hormone signalling, regulation of redox homeostasis | unknown | In regulation of plant growth and development | G. hirsutum | [54] | |
| TaGPX1-12 | Heat, drought and/or a combination, salt | Possible role of TaGPXs in development and stress response, putative roles in signal transductions | GSH biosynthetic and metabolic processes, DNA metabolic processes | Putative roles in plant growth and development, in leaf developmental stages, roots, stems, spikes, and grain | T. aestivum | [55] | |
| ZmGPX1-7 | Drought, waterlogging | Stress regulation through regulatory elements and splicing mechanisms | Growth, development | Z. mays | [52] | ||
| Function/Involvement in: | Heme-Peroxidases Family: Class III Peroxidases | Non-Heme Peroxidases Family: Glutathione Peroxidases |
|---|---|---|
| ROS metabolism | + | + |
| Redox signalling | + | + |
| Defence against pathogen infection | + | + |
| Defence against abiotic stresses | + | + |
| Wound healing | + | − |
| Cell wall metabolism | + | − |
| Lignification and suberization | + | − |
| Defence of membranes | − | + |
| Growth and development | + | + |
| Seed germination | + | + |
| Growth of roots | + | + |
| Growth of shoots | + | + |
| Flowering | + | + |
| Fertilization, pollen tube growth | + | + |
| Embryogenesis, seed development | + | + |
| Fruit growth and ripening | + | + |
| Interaction with plant hormones | + | + |
| Auxin catabolism | + | − |
| Auxin transport | − | + |
| Ethylene biosynthesis | + | + |
| Ethylene signalling | − | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bela, K.; Riyazuddin, R.; Csiszár, J. Plant Glutathione Peroxidases: Non-Heme Peroxidases with Large Functional Flexibility as a Core Component of ROS-Processing Mechanisms and Signalling. Antioxidants 2022, 11, 1624. https://doi.org/10.3390/antiox11081624
Bela K, Riyazuddin R, Csiszár J. Plant Glutathione Peroxidases: Non-Heme Peroxidases with Large Functional Flexibility as a Core Component of ROS-Processing Mechanisms and Signalling. Antioxidants. 2022; 11(8):1624. https://doi.org/10.3390/antiox11081624
Chicago/Turabian StyleBela, Krisztina, Riyazuddin Riyazuddin, and Jolán Csiszár. 2022. "Plant Glutathione Peroxidases: Non-Heme Peroxidases with Large Functional Flexibility as a Core Component of ROS-Processing Mechanisms and Signalling" Antioxidants 11, no. 8: 1624. https://doi.org/10.3390/antiox11081624